

Short-term responses to sheep grazing in a Patagonian steppe
Magalí D. Valenta A F , Rodolfo A. Golluscio B C , Ana L. Frey A , Lucas A. Garibaldi D and Pablo A. Cipriotti C E
A F , Rodolfo A. Golluscio B C , Ana L. Frey A , Lucas A. Garibaldi D and Pablo A. Cipriotti C E
A Universidad de Buenos Aires, Facultad de Agronomía, Cátedra de Ovinotecnia, Buenos Aires, Argentina.
B Universidad de Buenos Aires, Facultad de Agronomía, Cátedra de Forrajicultura, Buenos Aires, Argentina.
C Universidad de Buenos Aires, Consejo Nacional de Investigaciones Científicas y Técnicas, Instituto de Investigaciones Fisiológicas y Ecológicas Vinculadas a la Agricultura (IFEVA), Facultad de Agronomía, Buenos Aires, Argentina.
D Instituto de Investigaciones en Recursos Naturales, Agroecología y Desarrollo Rural (IRNAD), Sede Andina, Universidad Nacional de Río Negro (UNRN) y Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Mitre 630, CP 8400, San Carlos de Bariloche, Río Negro, Argentina.
E Universidad de Buenos Aires, Facultad de Agronomía. Departamento de Métodos Cuantitativos y Sistemas de Información, Buenos Aires, Argentina.
F Corresponding author. Email: mvalenta@agro.uba.ar
The Rangeland Journal 42(1) 1-8 https://doi.org/10.1071/RJ19012
Submitted: 15 March 2019 Accepted: 16 December 2019 Published: 25 March 2020
Abstract
Grazing modifies ecosystem function through direct effects on plants, but also through indirect effects mediated by floristic changes induced by grazing. Although both types of effects occur in the long term, only the direct effects are evident in the short term. We evaluated the short-term direct effects of sheep (Ovis aries) grazing on a Patagonian steppe during one growing season. We measured plant aerial cover in permanent transects located at increasing distances from a watering point in three paddocks with different stocking rates through the growing season. We also measured frequency of defoliation for vegetative and reproductive phases of different plant species located along these transects. Sheep grazing directly (a) reduced aerial cover and/or increased frequency of defoliation of certain preferred grasses and perennial forbs, (b) did not increase the aerial cover of any life form, but only the proportion of bare soil, (c) did not change the litter aerial cover, and (d) defoliated the flowers of even the least preferred shrub. Result a) was coincident with previous plant aerial cover long-term studies; but results (b) and (c) were contrary to long-term studies, probably because they resulted from indirect rather than direct grazing effects. Result (d) was not detected by long-term studies, probably because flower defoliation through grazing is undetectable when measuring shrub plant aerial cover. Our study showed that grazing has short-term direct effects mainly on the most preferred species. This could be useful for rangeland management and conservation of Patagonian steppes because short-term effects may be more easily reversible than long-term ones, and may provide early warning of rangeland condition deterioration.
Additional keywords: degradation, defoliation, herbivory, non-preferred species, plant functional types, preferred species.
Introduction
Grazing may affect plant communities either directly or indirectly (Milchunas and Lauenroth 1993; Bisigato and Bertiller 1997; Golluscio et al. 2009). Direct effects, such as trampling and removal of photosynthetic or reproductive plant organs of certain species (Olofsson 2006), occur either in the short- or long-term (a few weeks or several years respectively). Indirect effects are given by successional changes in rangeland communities driven by grazing. Grazing alters the competitive interactions between neighbouring plants by changing resource availability and by selective consumption of some species (Olofsson 2006; Graff et al. 2007). These changes are evident only in the long term. The study of short-term responses may allow quantification of some of the most dynamic direct effects of grazing, almost without any confusion with indirect effects. Quantifying short-term direct grazing effects on plants is important for rangeland management and conservation issues, such as ecosystem services, because these effects may be more readily detected than long-term ones, and may provide early warning of condition deterioration. Most studies compared sites with different historical grazing intensities (Briske et al. 2008), and hence, represent the long-term response of plant communities to grazing, showing both direct and indirect effects of grazing on rangelands. Few studies have evaluated the response of plant communities to grazing in the medium term (several years) and discuss the direct and indirect grazing effects (Olofsson 2006; Austrheim et al. 2008; Oñatibia and Aguiar 2016). To our knowledge, there are few studies evaluating the direct effects of sheep grazing in the short term (one growing season).
This study was conducted in a Patagonian steppe, an ecosystem subject to severe desertification processes induced by selective sheep grazing as a result of inappropriate grazing practices (León and Aguiar 1985). Several studies have analysed long-term effects of grazing on cover, floristic composition, and plant density. These studies recorded a decrease in overall plant and litter cover, and specifically a decrease in the cover of preferred grass and forb species, together with an increase in that of some less preferred grass and shrub species (Soriano et al. 1980; Fernández et al. 1992; Perelman et al. 1997; Graff et al. 2007; Golluscio et al. 2009; Cesa and Paruelo 2011). Our conceptual model is that in the short-term sheep grazing may negatively affect preferred species (direct effect), in the long term causing floristic changes linked to the alteration of the competitive interactions favouring non-preferred over preferred species (indirect effects). However, it is not well known which of the preferred species are most responsive to grazing. Our objective was to analyse (a) the change of plant aerial cover – at the species and functional group organisation levels – through one growing season along a gradient of stocking rates, given by the combination of paddock stocking rate and the distance from the water point in each paddock, and (b) the frequency of defoliation for vegetative and reproductive phases of different plant species. Since grazing could not have affected floristic composition or biogeochemical cycles during this short period, we assumed our work evaluated mainly direct grazing effects.
Methods
Study site
The trial was conducted in three paddocks subject to different stocking rates with a common watering point at the Experimental Ranch of the National Institute of Agricultural Technology (INTA) at Río Mayo (45°24′38.7″S, 70°17′37.8″W), south of the Occidental District of the Patagonian Phytogeographic Province of Argentina (León et al. 1998). The paddocks were not used for more than 12 months before our study. The vegetation has an upper shrubby layer, and a lower herbaceous layer, composed of grasses and forbs (Soriano and Sala 1984). It is characterised as a shrub-grass steppe of Poa ligularis Nees ex. Steud., Pappostipa speciosa (Trin. & Rupr.) Romaschenko, Mulinum spinosum (Cav.) Pers., Adesmia volckmannii Phil. and Senecio filaginoides DC, on a loam to loam-sandy soil of moderate depth, with high stoniness (Paruelo et al. 1988). The climate is cold temperate, with mean monthly temperature ranging from 2°C (July) to 8.4°C (January) and a mean annual precipitation of 152 mm, 70% falling in autumn–winter (Jobbágy and Sala 2000).
Study design
Forty Australian Merino wethers (average weight = 40.2 kg) grazed each of three paddocks from October 2005 to February 2006. Since paddocks varied in size (100, 136 and 170 ha), we achieved a stocking rate, of 1.2, 0.9 and 0.7 sheep equivalent (SE) per ha during the occupation period, equivalent to an annual stocking rate of 0.4, 0.3 and 0.2 SE ha−1 respectively. These stocking rates are higher, equal and lower, respectively, than the average annual stocking rate in the region (0.3 SE ha−1; Fernández et al. 1992), and all are higher than the usual stocking rate of the Experimental Ranch. The sheep equivalent represents the average annual requirements of a 50 kg pregnant ewe and which rears a lamb to three months, including the forage consumed by the lamb (Cocimano et al. 1977). To increase the range of the grazing pressure gradient analysed, we stratified our sampling by marking four permanent transects of 60 m within each paddock, tangent to concentric circles located at increasing distances from the common watering point (250, 500, 750 and 1000 m). The distance between transects (250 m) allowed covering the steppe heterogeneity present in the three paddocks. The localised stocking rate is usually higher than the mean stocking rate near the watering point and decreases with the distance from the watering point. Except in areas very close to watering or resting points, dung density is considered an acceptable indicator of local grazing intensity (Lange and Willcocks 1978; Bisigato and Bertiller 1997). Dung pellets along each transect were counted in an area of 24 m2 (60 m × 0.4 m), and from this the localised stocking rate for each transect (LSR, SE ha−1) was estimated as the product of the average stocking rate of each paddock and the ratio between fresh dung density of each transect and mean fresh dung density of each paddock:
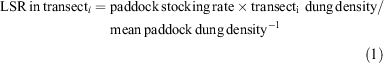
Data collection
Along each transect we measured aerial cover of plant species, litter, and bare soil using the linear intercept method (Greig Smith 1983). Although basal cover is more directly related with long-term degradation than aerial cover, we measured aerial cover because it is more variable in the short term, the focal point of our work. We made the measurements in October, November, December, January and February (5, 50, 80, 110 and 140 days since the start of grazing respectively). We pooled aerial cover data at different levels of organisation (a) litter, bare soil, preferred and non-preferred species (specific quality index (SQI) of each species ≥2 and <2 respectively; Elissalde et al. 2002), (b) plant functional types resulting from classifying the 24 species explaining 96% of the total aerial cover (Golluscio et al. 2005) by subdividing the four life forms (grasses, shrubs, perennial forbs and annual forbs), into preferred and non-preferred types, and (c) the twenty four species mentioned above. For the seven most abundant species of the community which accounted for 82% of total aerial cover (Bromus pictus Hook., P. ligularis and P. speciosa (preferred grasses), Pappostipa major (Speg.) Romaschenko (non-preferred grass), A. volckmannii and M. spinosum (preferred shrubs), and S. filaginoides (non-preferred shrub; Golluscio et al. 2005)), we also measured the frequency of defoliation during vegetative (November) and reproductive (February) phases, as another measurement of direct effects of grazing. To measure frequency of defoliation, at 10 points regularly distanced on each transect, we noted signs of defoliation in the three nearest individuals of each species, and estimated the frequency of defoliated plants ((no. defoliated plants × no. total plants−1) × 100). In the case of grasses, defoliated organs were leaves, and in shrubs, they were leaves during the vegetative phase, but flowers during the reproductive phase.
Data analyses
We analysed changes in aerial cover of litter, bare soil, preferred and non-preferred species; the main functional groups (i.e. preferred and non-preferred grasses, preferred and non-preferred shrubs, preferred and non-preferred perennial forbs, and non-preferred annual forbs; no preferred annual forb species were reported for this plant community), and the 24 most conspicuous plant species of the steppe community to analyse the vegetation dynamics through a growing season and under different localised stocking rates. We adopted a linear mixed model with fixed and random effects to accommodate the temporal correlation of repeated-measurements in the same transects and paddocks. We tested for the fixed effects of time (days since the start of grazing) and the localised stocking rate at the transect level (LSR), and their interaction for each functional group or species by using mixed-effect models (R software, nlme package; Pinheiro et al. 2009). Random effects were considered through a random intercept model for the paddocks and transects included the temporal correlation (Zuur et al. 2009). Our model considered that measurements from plants were nested within transects, and transects were nested within paddocks.
For frequency of defoliation at vegetative and reproductive phases of the above-mentioned seven most abundant species, we adopted a generalised linear mixed model with fixed and random effects to accommodate the random nature of transects and paddocks. We adopted a binomial family and link function logit to reflect the nature of response variable at plant level (0 = no defoliated, 1 = defoliated). First, we tested for a global model the fixed effects of phenological phase and plant species and their interaction by using Chi-square tests (R software, lme4 package, Bates et al. 2015). Second, we conducted single models to assess the role of phenological phase across different species or the differences among species for a phenological phase. We built the punctual predictions and confidence intervals (α = 5%) based on estimates of fixed and random effects of fitted mixed models.
Results
Monthly temperature during the trial was similar to that from the 1985–2004 historic record, but precipitation during the October–February period was higher (61 vs 36 mm, for 2005–2006 and historic record, respectively, Fig. 1).

|
Aerial cover of preferred species significantly decreased (P < 0.01), whereas that of non-preferred species did not change during the growing season (P > 0.05) (Table 1; Fig. 2a). The proportion of bare soil increased throughout the experiment (0.01 < P < 0.05), whereas litter aerial cover was stable (Table 1; Fig. 2b). The aerial cover of preferred species and bare soil was unaffected by LSR (P > 0.05) (Table 1). Aerial cover of preferred grasses and non-preferred annual forbs decreased, whereas that of non-preferred grasses, and preferred and non-preferred shrubs and perennial forbs did not change during the experimental period. Preferred grasses were also negatively affected by LSR (Table 1; Fig. 2c, e, g, h) and positively affected by the interaction between time and LSR. The positive interaction slope suggests that decrease in aerial cover as days under grazing and LSR increased is less pronounced at high than at low values of both variables (Table 1).

|

|
At the species level, aerial cover only decreased in two of the preferred grasses, P. ligularis and P. speciosa (Table 1; Fig. 2d). Aerial cover also decreased in Microsteris gracilis within the annual forbs. LSR mainly reduced plant aerial cover in preferred grasses, specifically P. ligularis and P. speciosa, but increased aerial cover in the preferred perennial forb Calceolaria polyrhiza, perhaps by the relaxation of competition exerted by P. ligularis and P. speciosa as LSR increased (Table 1). The slight aerial cover increase of the preferred grass and shrub species recorded around day 100 may have been related to rain events in January and February 2006 (Fig. 1).
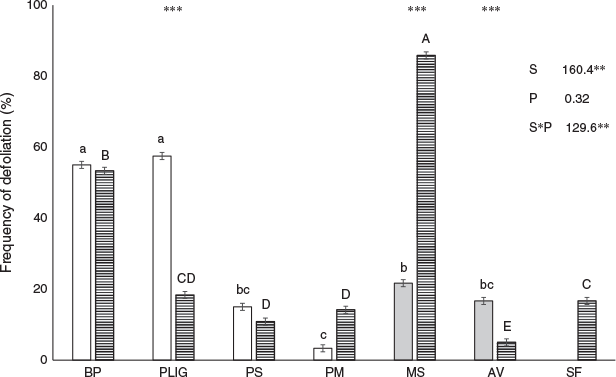
The ranking of frequency of defoliation among species was different in both phenological phases (Fig. 3). In the vegetative phase, the grasses B. pictus and P. ligularis were the most defoliated, with similar defoliation frequency (Fig. 3). Shrubs were less defoliated than these grasses, but the preferred shrubs Mulinun spinosum and A. volckmannii were more defoliated than the non-preferred S. filaginoides, which was remained undefoliated during this phase (Fig. 3). In the reproductive phase, defoliation of some shrubs was similar to that of the most defoliated grasses. Mulinum spinosum had the highest defoliation frequency, followed by B. pictus. Conversely, in the reproductive phase, the preferred A. volckmannii was the least defoliated shrub while, S. filaginoides had a higher defoliation frequency than A. volckmannii (only slightly lower than M. spinosum) (Fig. 3).
|
Discussion
Short-term grazing reduced the aerial cover of preferred species as grazing duration increased, and this was more evident in grasses, specifically in the preferred species P. ligularis and the less preferred – but dominant – P. speciosa, than in shrubs and forbs. In contrast to the effect of grazing duration, our trial only was able to detect significant effects of LSR in the case of the preferred grasses, specifically in the above-mentioned P. ligularis and P. speciosa. As grazing duration increased, total aerial cover of preferred grass species decreased, while total aerial cover of non-preferred species was stable over the growing season. The aerial cover decrease in annual non-preferred forbs and of M. gracilis within them may have been associated with their phenological pattern: towards January–February annual forbs lose most of their leaves, senesce and disperse their seeds (Golluscio et al. 2005). The lack of a clear and significant response in the aerial cover of shrubs cannot be considered as a decisive direct effect because the aerial cover change depends on population processes. Although individual plants of some shrub species can show clear biomass change, this is not the case in the three shrub species we studied, where population biomass depends more on population density than on individual biomass, and population density in turn depends on mortality and recruitment which operate at time scales much longer than one year. Conversely, in grasses, tillering, perhaps the main recruitment strategy, operates at short time periods (Loeser et al. 2004). Bromus pictus was more defoliated than P. ligularis which in turn was more defoliated than P. speciosa. The most defoliated grass species showed an increase in defoliation frequency, but no change in aerial cover during the grazing period. At the other end of the gradient, P. speciosa, third in the preference ranking, showed a low frequency of defoliation but a 50% decrease of aerial cover. P. ligularis, located between these two extremes in the grazing preference gradient, was the only species to show both an aerial cover decrease and a high defoliation frequency (Figs 2d, 3; Table 1). This sensible grazing of P. ligularis, together with its response to LSR reinforces the usual acceptance of P. ligularis a valuable key species.
From November to January sheep diet changes from a predominance of preferred grass and shrub leaves, which lose quality when blooming, to a predominance of shrub reproductive organs, together with the leaves of the most preferred grasses. Advanced maturity reduces digestibility and toughens grass leaves, so sheep select other forage sources, mainly shrub flowers. Leaves of A. volckmannii and M. spinosum have physical defences (thorns), whereas those of S. filaginoides have chemical defences (Cavagnaro et al. 2003; Cavagnaro and Golluscio 2017). These defences enable them to evade grazing through a reduction in the accessibility and palatability of their leaves, respectively. This study demonstrated that sheep defoliated the leaves of shrubs with fewer chemical defence (A. volckmannii and M. spinosum), and the flowers of all.
Sheep defoliated flowers of the three most abundant shrubs (direct effect), but this did not affect the shrub aerial cover. Some studies have found a similar lack of shrub cover change with grazing but findings vary. Recorded cover increased in the least preferred S. filaginoides (Fernández et al. 1992; Bertiller and Bisigato 1998; Cesa and Paruelo 2011), whereas cover decreased in the preferred M. spinosum and A. volckmannii (Fernández et al. 1992; Cesa and Paruelo 2011). Cover increases in the preferred shrubs have also been recorded (Soriano et al. 1980; Bertiller and Bisigato 1998). We suggest that the net effect of grazing on the aerial cover of shrub species not only operates at time scales longer than one year, but also depends on the balance between direct and indirect grazing effects. We propose that under low localised stocking rates, the aerial cover of S. filaginoides may decrease because of damage caused by grazing to their reproductive organs (direct effect). However, at higher localised stocking rates, the aerial cover of S. filaginoides may increase because of the resource liberation brought about by the loss of aerial cover of more preferred grasses, shrubs and perennial forbs (indirect effect).
Our results are similar to those of several studies analysing the long-term effects of grazing in Patagonian steppe: an increase in the proportion of bare soil (Soriano et al. 1980; Bottaro 2007; Golluscio et al. 2009), an aerial cover decrease of preferred grasses (Bertiller and Bisigato 1998; Bottaro 2007; Graff et al. 2007), and a loss of forb species (Perelman et al. 1997; Bottaro 2007; Golluscio et al. 2009; Cesa and Paruelo 2011). However, in contrast to our study, previous long-term studies recorded a decrease in aerial cover of litter and standing dead biomass (Soriano et al. 1980; Golluscio et al. 2009; Cesa and Paruelo 2011), an increase of aerial cover of non-preferred grasses (Bertiller and Bisigato 1998; Bottaro 2007; Cesa and Paruelo 2011), the replacement of preferred by non-preferred grasses (Perelman et al. 1997; Golluscio et al. 2009; Cesa and Paruelo 2011), and an increase in shrub cover (Soriano et al. 1980; Bertiller and Bisigato 1998), resulting in significant changes in floristic composition of the steppe. Most of these long-term changes may have been the result of indirect grazing effects rather than direct effects, i.e. the floristic change induced by the loss of preferred species favouring the increase in aerial cover of non-preferred species, and the alteration in the N cycle triggered by this floristic change (Golluscio et al. 2009). The detection of short-term direct effects is the first step to avoid such undesirable long-term effects.
Conclusions
This research demonstrates direct short-term responses of Patagonian steppes to herbivory by using two variables: the aerial cover dynamics and the frequency of defoliation of different plant species throughout one growing season. Patagonian grasslands show high to very high risk of desertification (Perelman et al. 1997; Del Valle et al. 1998). The early detection of the patterns and processes that lead to degradation of the arid and semiarid grasslands is a key technological tool because most signs of degradation are evident late, when is very expensive to apply reversion practices (Milton et al. 1994). In terms of adaptive grazing management, it is important to have indicators of defoliation in plants that show signs in the short term and act as early warning indicators of the use of forage resources mainly in those systems characterised by continuous grazing. Therefore, it is useful to detect which species would be key in detecting direct effects of grazing in the short term. This study confirmed the known value of P. ligularis as key species for management because it is an abundant – but not dominant – preferred grass with high defoliation frequency, with the most pronounced aerial cover decreases in the short term as a function of both grazing duration and LSR. Quantifying the direct effects of sheep grazing on plants is important for rangeland monitoring, management and for the conservation of ecosystem services because short-term effects may be more easily reversible than long-term ones and provide early warnings against degradation processes.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
Magalí Valenta wants to thank Mirta Rabadán, Fernando Cavagnaro and Guillermo García Martínez for their cooperation in data gathering, Martín Aguiar and two anonymous reviewers for their commentaries and support, technicians of the INTA Experimental Stations of Esquel, Trelew and Bariloche. We also thank INTA for permission to work in the Experimental Ranch of Río Mayo (Chubut, Argentina). UBA (G044), ANPCYT (PICT 00463 and 2243), and CONICET (PIP 5963/05) provided economic support for this project, and M.D.V. received a CONICET postgraduate fellowship.
References
Austrheim, G., Mysterud, A., Pedersen, B., Halvorsen, R., Hassel, K., and Evju, M. (2008). Large scale experimental effects of three levels of sheep densities on an alpine ecosystem. Oikos 117, 837–846.| Large scale experimental effects of three levels of sheep densities on an alpine ecosystem.Crossref | GoogleScholarGoogle Scholar |
Bates, D., Maechler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67, 1–48.
| Fitting linear mixed-effects models using lme4.Crossref | GoogleScholarGoogle Scholar |
Bertiller, M. B., and Bisigato, A. (1998). Vegetation dynamics under grazing disturbance. The state-and-transition model for the Patagonian steppes. Ecología Austral 8, 191–199.
Bisigato, A. J., and Bertiller, M. B. (1997). Grazing effects on patchy dryland vegetation in northern Patagonia. Journal of Arid Environments 36, 639–653.
| Grazing effects on patchy dryland vegetation in northern Patagonia.Crossref | GoogleScholarGoogle Scholar |
Bottaro, H. S. (2007). Influencias de la productividad, la calidad de la vegetación y el deterioro sobre la receptividad ganadera en el NO de la Patagonia. thesis, Facultad de Agronomía, Universidad de Buenos Aires, Buenos Aires, Argentina.
Briske, D. D., Derner, J. D., Brown, J. R., Fublendorf, S. D., Teaque, W. R., Havstad, K. M., Gillen, R. L., Ash, A. J., and Willms, W. D. (2008). Rotational grazing on rangelands: reconciliation of perception and experimental evidence. Rangeland Ecology and Management 61, 3–17.
| Rotational grazing on rangelands: reconciliation of perception and experimental evidence.Crossref | GoogleScholarGoogle Scholar |
Cavagnaro, F. P., and Golluscio, R. A. (2017). Structural anti-herbivore defense reduction of two Patagonian spiny shrubs in response to long time exclusion of large herbivores. Journal of Arid Environments 142, 36–40.
| Structural anti-herbivore defense reduction of two Patagonian spiny shrubs in response to long time exclusion of large herbivores.Crossref | GoogleScholarGoogle Scholar |
Cavagnaro, F. P., Golluscio, R. A., Wassner, D. F., and Ravetta, D. A. (2003). Caracterización química de arbustos patagónicos con diferente preferencia para los herbívoros. Ecología Austral 13, 215–222.
Cesa, A., and Paruelo, J. M. (2011). Changes in vegetation structure induced by domestic grazing in Patagonia (Southern Argentina). Journal of Arid Environments 75, 1129–1135.
| Changes in vegetation structure induced by domestic grazing in Patagonia (Southern Argentina).Crossref | GoogleScholarGoogle Scholar |
Cocimano, M., Lange, A., and Menvielle, E. (1977). Equivalencias ganaderas para vacunos de carne y ovinos. (AA-CREA: Buenos Aires, Argentina.)
Del Valle, H. F., Elissalde, N. O., Gagliardini, D. A., and Milovich, J. (1998). Status of desertification in the Patagonian Region: assessment and mapping from satellite imagery. Arid Soil Research and Rehabilitation 12, 95–122.
Elissalde, N., Escobar, J., and Nakamatsu, V. (2002). ‘Inventario y evaluación de pastizales naturales de la zona árida y semiárida de la Patagonia. Programa de acción nacional de lucha contra la desertificación.’ (INTA-GTZ: Chubut, Argentina.)
Fernández, R. J., Nuñez, A. H., and Soriano, A. (1992). Contrasting demography of two Patagonian shrubs under different conditions of sheep grazing and resource supply. Oecologia 91, 39–46.
| Contrasting demography of two Patagonian shrubs under different conditions of sheep grazing and resource supply.Crossref | GoogleScholarGoogle Scholar | 28313371PubMed |
Golluscio, R. A., Oesterheld, M., and Aguiar, M. R. (2005). Relationship between phenology and life form: a test with twenty five Patagonian species. Ecography 28, 273–282.
| Relationship between phenology and life form: a test with twenty five Patagonian species.Crossref | GoogleScholarGoogle Scholar |
Golluscio, R. A., Austin, A. T., García Martínez, G. C., Gonzalez-Polo, M., Sala, O. E., and Jackson, R. B. (2009). Sheep grazing decreases organic carbon and nitrogen pools in the Patagonian steppe: combination of direct and indirect effects. Ecosystems 12, 686–697.
| Sheep grazing decreases organic carbon and nitrogen pools in the Patagonian steppe: combination of direct and indirect effects.Crossref | GoogleScholarGoogle Scholar |
Graff, P., Aguiar, M. R., and Chaneton, J. C. (2007). Shifts in positive and negative plant interactions along a grazing intensity gradient. Ecology 88, 188–199.
| Shifts in positive and negative plant interactions along a grazing intensity gradient.Crossref | GoogleScholarGoogle Scholar | 17489467PubMed |
Greig Smith, P. (1983). ‘Quantitative Plant Ecology.’ 3rd edn. (Blackwell Scientific Publications: Oxford, UK.)
Jobbágy, E., and Sala, O. E. (2000). Controls of grass and shrub aboveground production in the Patagonian steppe. Ecological Applications 10, 541–549.
| Controls of grass and shrub aboveground production in the Patagonian steppe.Crossref | GoogleScholarGoogle Scholar |
Lange, R. T., and Willcocks, M. C. (1978). The relation between sheep-time spent and egesta accumulated within in arid zone paddock. Australian Journal of Experimental Agriculture and Animal Husbandry 18, 764–767.
| The relation between sheep-time spent and egesta accumulated within in arid zone paddock.Crossref | GoogleScholarGoogle Scholar |
León, R. J. C., and Aguiar, M. R. (1985). El deterioro por uso pastoril en estepas herbáceas patagónicas. Phytocoenologia 13, 181–196.
León, R. J. C., Bran, D., Collantes, M., Paruelo, J. M., and Soriano, A. (1998). Grandes unidades de vegetación de la Patagonia extra andina. Ecología Austral 8, 125–144.
Loeser, M. R., Crews, T. E., and Sisk, T. D. (2004). Defoliation increased above-ground productivity in a semi-arid grassland. Journal of Range Management 57, 442–447.
| Defoliation increased above-ground productivity in a semi-arid grassland.Crossref | GoogleScholarGoogle Scholar |
Milchunas, D. G., and Lauenroth, W. K. (1993). Quantitative effect of grazing on vegetation and soils over a global range of environments. Ecological Monographs 63, 327–366.
| Quantitative effect of grazing on vegetation and soils over a global range of environments.Crossref | GoogleScholarGoogle Scholar |
Milton, S. J., Dean, W. R. J., du Plessis, M. A., and Siegfried, W. R. (1994). A conceptual model of arid rangeland degradation. The escalating cost of declining productivity. Bioscience 44, 70–76.
| A conceptual model of arid rangeland degradation. The escalating cost of declining productivity.Crossref | GoogleScholarGoogle Scholar |
Olofsson, J. (2006). Short- and long-term effects of changes in reindeer grazing pressure on tundra heath vegetation. Journal of Ecology 94, 431–440.
| Short- and long-term effects of changes in reindeer grazing pressure on tundra heath vegetation.Crossref | GoogleScholarGoogle Scholar |
Oñatibia, G. R., and Aguiar, M. R. (2016). Continuous moderate grazing management promotes biomass production in Patagonian arid rangelands. Journal of Arid Environments 125, 73–79.
| Continuous moderate grazing management promotes biomass production in Patagonian arid rangelands.Crossref | GoogleScholarGoogle Scholar |
Paruelo, J. M., Aguiar, M. R., and Golluscio, R. A. (1988). Soil water availability in the Patagonian and steppe gravel content effect. Arid Soil Research and Rehabilitation 2, 67–74.
| Soil water availability in the Patagonian and steppe gravel content effect.Crossref | GoogleScholarGoogle Scholar |
Perelman, S., León, R., and Bussacca, J. (1997). Floristic changes related to grazing intensity in a Patagonian shrub steppe. Ecography 20, 400–406.
| Floristic changes related to grazing intensity in a Patagonian shrub steppe.Crossref | GoogleScholarGoogle Scholar |
Pinheiro, J., Bates, D., DebRoy, S., and Sarkar, D. (2009). The R Core Team, nlme: Linear and Nonlinear Mixed Effects Models. R package ver. 3.1-96.
Sistema de Información Patagonia Sur (SIPA) (2019). Resumen Anual de Boletín Meteorológico Estación Meteorológica Río Mayo. Available at: http://sipas.inta.gob.ar/?q=agrometeorologia-boletines-estacion&idEstacion=19 (accessed 20 June 2019).
Soriano, A., and Sala, O. E. (1984). Ecological strategies in a Patagonian arid steppe. Vegetation 56, 9–15.
| Ecological strategies in a Patagonian arid steppe.Crossref | GoogleScholarGoogle Scholar |
Soriano, A., Sala, O. E., and León, R. J. C. (1980). Vegetación actual y vegetación potencial en el pastizal de coirón amargo (Stipa spp.) del S.W. de Chubut. Boletín de la Sociedad Argentina de Botánica 19, 309–314.
Zuur, A. F., Ieno, E. N., Walker, N. J., Saveliev, A. A., and Smith, G. M. (2009). ‘Mixed Effects Models and Extensions in Ecology with R.’ (Springer: Berlin.)