

Implications of altered fire regimes for birds of dry sclerophyll forest under climate change
Michael J. M. Franklin A B * , Richard E. Major C and Ross A. Bradstock A
A B * , Richard E. Major C and Ross A. Bradstock A
A
B
C
Abstract
Climate change is driving increases in forest fire activity around the world. Consequently, recent and future research into the implications for affected biota, and its translation into policy and management, is of vital importance for the conservation of forest biodiversity. The dry sclerophyll forests of south-eastern Australia are affected by changing fire regimes, including having recently been extensively burnt. In this review, we synthesise contemporary research into the responses of dry sclerophyll forest birds to fire regimes, with a focus on contrasts in levels of time since fire, fire severity, fire frequency, and their interactions. We identify and discuss high-priority knowledge gaps and management issues in this domain. Recent studies have gained insights into the complexity of the effects of fire on birds by, for example, showing how avian fire responses are influenced by climate variability. In dry sclerophyll forests, most species that responded to time since fire were less likely to occur where it was short. There were mixed positive and negative responses to increasing fire severity and/or frequency among species in these forest bird assemblages. Given that quantification of the risk of extinction of rare and uncommon birds under increased fire activity is challenging, options for investigating their fire response are proposed. Availability of habitat for birds that flourish where fire is recent, severe and/or regular is unlikely to decline given current trends. We present a management approach that conceptualises large blocks of biologically connected forest with longer fire ages, but otherwise differing fire histories, as biodiversity reservoirs.
Keywords: avian ecology, climate change, dry sclerophyll forest, fire frequency, fire regime, fire severity, rare species, time since fire.
Introduction
Wildfires are major disturbances, which have long-term impacts on biota in forested landscapes. Further to the effects of any one fire, biological communities respond to fire regimes, which represent dynamic combinations of fire frequency, intensity, season of occurrence, and the spatial extent of fires at a given location through time (Gill 1975; Bond and Keeley 2005). Over the coming decades, climate change is predicted to continue to drive increases in fire activity in forested landscapes worldwide (de Groot et al. 2013; Dupuy et al. 2020), including those located in south-eastern Australia (Dowdy et al. 2019). These predictions have already manifested at continental scales, as evidenced in the 2019–2020 mega-fires in eastern Australia (Abram et al. 2021), which burnt extraordinary amounts of natural vegetation at high severity (Collins et al. 2021). These fires impacted substantial proportions of the distributions of many vertebrate species, including those with threatened status before the fires (Ward et al. 2020). A consequence of larger, more frequent fires is that the total amount of recently-burnt forest in a region is increased (Fairman et al. 2016), which has potential, major implications for fauna that require older habitat. For instance, particular bird species were more likely to occur where time since fire was longer in foothill–gully forest systems (Robinson et al. 2014), so there is potential for significant reductions in suitable habitat for those species.
The responses of individual bird species to fire regimes vary across habitats and are influenced by the effects of fire on vegetation, as well as environmental and climatic factors (Woinarski and Recher 1997; Bradstock et al. 2005; Valentine et al. 2012; Woinarski and Legge 2013; Kelly et al. 2017; Connell et al. 2022). However, potential habitat for some species may only exist under narrow combinations of fire regime variables (Hutto and Patterson 2016). Understanding the response of birds at the species level in their various habitats is necessary to maintain populations across species distributions (Driscoll et al. 2010; Connell et al. 2017). For example, the tropical savannas of northern Australia have experienced increases in the frequency of intense, late-dry season fires, to which 43 of 44 savanna bird species responded by showing range contractions in distribution modelling (Reside et al. 2012). Insights can be obtained by analysing information about species distributions in fire-prone landscapes in association with species traits and life histories (e.g. Gosper et al. 2019a).
Our understanding of the most suitable fire history patterns for the maintenance of avian diversity is incomplete or absent for many regions and ecosystems (Kelly et al. 2015). Because the responses of many bird species to fire are context-dependent, gaining and integrating understanding about species response mechanisms is important for fire management that aims to reduce the risk of extinction of any species (Driscoll et al. 2010). Identification of favourable fire regimes for different functional groups of bird species is an important area for future research (Kelly et al. 2017). However, insights will be reliant on a sound understanding of the life-cycle processes underpinning the fire responses of focal species, and the habitat factors that mediate them (Whelan et al. 2002).
Over decades, successive wildfires burn through dry sclerophyll forest landscapes at different levels of severity and frequency. Together with planned or prescribed burns, wildfires produce complex spatial patterns representing different fire histories in the habitat used by birds. Following a wildfire, regrowth of the vegetation is a function of time since fire and climatic conditions superimposed on the antecedent fire regime, and there is corresponding variation in the availability of resources used by birds. In these ways, fire regimes shape the structure and composition of vegetation, and birds respond in time and space to their fire-affected habitats (Bradstock et al. 2005; Woinarski and Legge 2013).
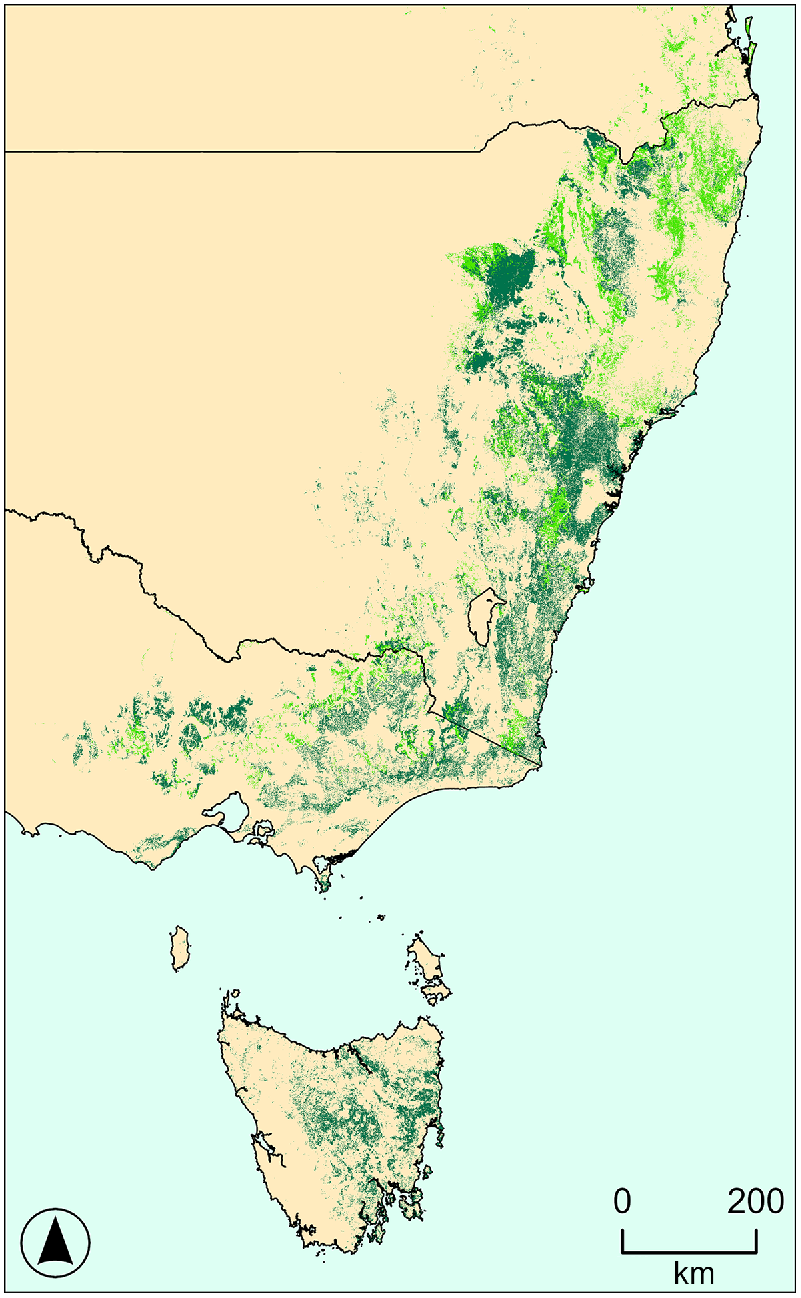
Recent advances have been made in understanding avian responses to time since fire (Gosper et al. 2019a), and components of fire regimes such as fire frequency (Kelly et al. 2017), in particular Australian systems, including dry sclerophyll forest (Fig. 1). However, we need to more fully understand the implications of changing fire regimes for dry sclerophyll forest birds in terms of their habitat requirements, response mechanisms, and population processes. In south-eastern Australia, extensive dry sclerophyll forest communities support the spread of wildfires on and adjacent to the Great Dividing Range, and are subject to widely varying fire regimes (Keith 2004). Many of the constituent species regenerate following fire by resprouting from epicormic or lignotuber buds, whereas others use a range of strategies involving seeds (Keith 2004). These forests are floristically and structurally diverse, and can have shrubby or grassy understories (Fig. 1), with canopies 10–30 m tall and 30–70% foliage cover. Dry sclerophyll forests are important habitats for birds (Smith et al. 2019).
The distribution of dry sclerophyll forests with shrubby (dark green) and grassy (light green) understoreys across south-eastern Australia (Department of Climate Change, Energy, the Environment and Water 2020; Department of Planning and Environment 2022).
Scope and methods
In this review, we synthesise knowledge gained since 2010 about the responses of dry sclerophyll forest birds to fire regimes, to better predict responses to likely increases in wildfire activity under climate change (Abram et al. 2021). Driscoll et al. (2010) outlined outstanding research questions relating to fire and biodiversity and evaluated methodological frameworks for use in future investigations. They found knowledge gaps in the broad areas of species fire response mechanisms, effects of other processes on fire responses, and the influence of fire regimes with different spatiotemporal attributes. Here we focus on subsequent avian studies that address this lack of information using the latter theme as a framework, with main sections on species and guild responses to variation in time since fire, fire severity, fire frequency, and their interactions. A general synthesis of the literature on avian fire ecology in Australian habitats was conducted by Woinarski and Recher (1997). We include their relevant key findings, and those from a limited number of studies conducted prior to 2010, as supporting information to the core review. To assemble a candidate set of literature for this narrative review, the Web of Science, Scopus and Google Scholar were searched using the terms listed in Supplementary material. The reference lists of the resulting literature were also checked for relevant material.
Where relevant, the scope of this review extends to cover avian responses to fire in other vegetation communities such as woodland, which share bird species with dry sclerophyll forests. Studies conducted on other continents have been included where they contain conceptual or methodological approaches that may be applied to address knowledge gaps in the context of dry sclerophyll forest. This additional information provides a broader base for exploration of mechanisms underpinning species and guild fire responses. Studies conducted in dry sclerophyll forest were not excluded from the review to accommodate inclusion of domestic or international studies conducted in other vegetation types. Recent reviews have focused on animal movement (Nimmo et al. 2019), predation (Doherty et al. 2022) and mortality (Jolly et al. 2022) in relation to wildfire, so we include only additional material pertinent to the study system on these topics. We identify and discuss high-priority knowledge gaps and management issues in this domain.
Avian responses to time since fire
The responses of birds to time since fire vary by species and according to context, ranging from preferences for burnt habitat immediately following fire to long unburnt vegetation. In avian assemblages, abundance and/or richness at the species or functional group level in relation to time since fire are influenced by variation in the availability of resources required by species as the vegetation regenerates and matures (Gosper et al. 2019a). As such, time since fire is a proxy for a range of processes that vary spatially in fire-prone forests. Causal modelling frameworks enable explicit evaluation of such processes, for example, by partitioning avian responses to direct (population processes) and indirect (habitat structure) effects of time since fire (Franklin et al. 2023). Climatic factors interact with time since fire to influence resource availability, such that individual species responses to time since fire can differ substantially during periods of drought and higher than average rainfall (Connell et al. 2022, Table 1).
| Current knowledge | Implication under climate change | Knowledge gap for priority research | |
|---|---|---|---|
| Many species require habitat with longer time since fire. | Such species are vulnerable to sustained habitat loss. | No knowledge is available for many species. Investigate fire-age habitat requirements of species in context. | |
| Resources required by birds change through time since fire. | Resources available to birds at longer fire ages may be greatly reduced in landscapes. | Primary research is required to address the many knowledge gaps in how resources change. | |
| Long and short-term climate influences species responses to time since fire. | Increased climate variability, including extreme events, and overarching climate trends may interact strongly with fire history. | A wider scope of work on how birds respond to interactions between climate and fire regime components is required. | |
| Species can respond differently to time since fire in different dry sclerophyll forest habitats. | Some habitats may be impacted more than others by increased fire activity. | Predicted impacts of future fire on different habitats need to be linked to avian time since fire responses. | |
| Many species respond to the effects of time since fire on habitat structure, rather than time since fire per se. | Structure is also influenced by changing climate, environment, and other fire regime components. | Methods such as structural equation modelling can incorporate habitat structure and fire history, and should be used to disentangle these complex relationships. | |
| Limited capacity to move to optimal habitat in fire-prone landscapes might obscure fire responses of exclusively sedentary species. | Sedentary species that require older forest are vulnerable to sustained habitat loss. | More needs to be known about the time since fire habitat requirements (type, amount, configuration) of sedentary species in a landscape context. |
See the section Avian responses to time since fire for details and supporting literature.
Plant reproductive mechanisms and time to maturity have been used to establish a fire return interval of 7–30 years to guide fire management of dry sclerophyll forests with shrubby understories in eastern Australia (Bradstock and Kenny 2003; Kenny et al. 2004). In the south-east of their range, dry eucalypt forests mature 10–35 years after fire (Cheal 2010). The following three subsections are structured broadly in relation to these timeframes, as short (<7 years), mid-range (7–30 years) and long (>30 years) time since fire.
Short time since fire
The responses of birds to recent fire in eucalypt forests can vary widely among species and reflect both the direct effects of wildfire, and the effects of fire on required resources. Direct effects include mortality and impacts on population processes (Whelan et al. 2002), which may contribute to rapid population declines if large fires burn substantial proportions of species distributions, particularly if those fires are severe (Legge et al. 2021; Jolly et al. 2022). Wildfires alter the availability of habitat resources used by birds, including vegetation structure (Karna et al. 2020; Doherty et al. 2022), hollows for nesting (Haslem et al. 2012), and food (Valentine et al. 2014). The absence of understorey vegetation immediately after fire in dry eucalypt forests can result in an influx of bird species that use open areas for foraging. However, regeneration of the vegetation commences promptly, and bird species present pre-fire, but absent post-fire, may return to burnt areas within 2 or 3 years (Woinarski and Recher 1997).
Total bird abundance in a range of habitats, including dry sclerophyll forest, had recovered to pre-fire numbers by 3 years after fire, but the degree of recovery at the species level varied (Loyn 1997). For example, numbers of honeyeaters were very low immediately following fire, which was probably a function of their high mobility and capacity to track resources at regional scales. In diverse foothill forests dominated by eucalypts that resprout post-fire, some bird species were more likely to occur where time since fire was short, whereas the probability of occurrence in recently-burnt habitat was reduced for other species (Kelly et al. 2017). Five years after fire, montane dry sclerophyll forest was unsuitable for most species that responded to time since fire. Many species required habitat at a more advanced state of regeneration to support their mostly low levels of occurrence in landscapes (Franklin et al. 2022).
Mid-range time since fire
The responses of species, guilds, and assemblages to short time since fire are often not the same a decade or more after fire. In the year after fire, in mixed eucalypt/White Cypress Pine (Callitris glaucophylla) forest in western NSW, there were more species and individuals in areas that had not been burnt by wildfire than in the burnt area (Turner 1992). When the same areas were surveyed again eight years after the fire, there were many more species and individuals in the burnt forest than in the unburnt areas. The additional species in the burnt areas were mainly foliage, understorey, and ground foraging insectivorous species.
Shifts in the structure of bird assemblages were observed in response to time since the last hazard reduction burn in eucalypt woodland of South Australia. In addition to sites last burnt at least 20 years ago having more birds overall, assemblages in recently-burnt sites had increased proportions of large, generalist species, but these effects were lessened if some long unburnt woodland was accessible (Prowse et al. 2017). In north-eastern Australian woodlands, the extent of habitat with fire ages >10 years in the landscape was the most important attribute for landscape-scale richness of frugivores, insectivores and canopy species (Burgess and Maron 2016). In montane dry sclerophyll forests, regeneration of canopy structure as time since fire increased from 5 to 16 years was a key driver of increases in the occurrence of individual canopy species, and the richness of canopy and total species (Franklin et al. 2023). Factors that may influence vegetation structure in addition to time since fire, including altered plant community composition owing to increased fire frequency (Fairman et al. 2016), or different precipitation patterns under climate change (Williamson et al. 2014; Connell et al. 2022), will be important for birds in dry sclerophyll forests.
Relatively slow rates of vegetation regeneration can result in sustained negative responses from birds. In alpine areas, the post-fire recovery of vegetation, including trees such as the Snowgum (Eucalyptus pauciflora niphophila), takes longer than many of their low altitude counterparts owing to different regeneration mechanisms (Green 2013). Ten years after wildfire in the Snowy Mountains of south-east Australia, the Snowgum canopy had not regrown, which negatively impacted species that would forage in that space, especially granivores. Nectivorous birds were also less common, which corresponded with reduced availability of flowers from the obligate-seeding Grevillia victoriae (Green 2013).
Long time since fire
Birds continue to respond to time since fire up to and beyond 100 years in particular Australian ecosystems. In south-east Australian eucalypt forests, the majority of species that responded to time since fire up to 70 years, including the migratory Rose Robin (Petroica rosea), Yellow-faced Honeyeater (Lichenostomus chrysops) and Golden Whistler (Pachycephala pectoralis), were more abundant in long unburnt areas (Kelly et al. 2017). The responses of individual species in sites representing a 100-year gradient of time since fire differed in two types of regional mallee eucalypt community (Watson et al. 2012a). Only four of 16 species that responded significantly to time since fire did so in both vegetation communities, but the shape of the response curve was the same in each community for only two species, both of which were honeyeaters. Despite the variation in the linear and non-linear patterns of species responses, habitat with a time since fire of at least 20 years was important for many species in the region (Watson et al. 2012a). Similarly, in obligate-seeding Australian eucalypt woodland, most bird species were more abundant, and assemblages were richer, with increasing time since fire (Gosper et al. 2019a). Species responses to different aspects of fire regimes, such as time since fire, may vary along rainfall and temperature gradients (Kelly et al. 2017). For example, in south-east Australian eucalypt forests, the probability of occurrence of the Australian King Parrot (Alisterus scapularis) peaked in areas with longer time since fire, but only where summer temperatures were at the low end of the regional range (Kelly et al. 2017).
Long unburnt forest in the landscape
Individuals of many species may use habitat with different fire ages, providing that there is some long unburnt forest within their home range. For example, migratory species may engage in landscape complementation (Nimmo et al. 2019) during the breeding season, by accessing resources in dry sclerophyll forests at different fire ages, including long unburnt forest (Franklin et al. 2021a). In south Australian woodlands, long unburnt habitat in landscapes is necessary to maintain avian diversity under frequent burning regimes (Prowse et al. 2017). Older forest in the landscape can provide refuge from more recent fires for many species (Robinson et al. 2014). However, the size and spatial configuration of unburnt patches determines their utility as refugia from wildfire and as post-fire habitat for birds (Berry et al. 2015), in combination with the capacity and propensity of individual species to move in fire-affected landscapes (Nimmo et al. 2019). Unburnt habitat external to or within the spatial extent of a wildfire represents a base from which particular species may exploit resources in burnt areas, and source habitat for delayed recolonisation contingent on sufficient regeneration (Watson et al. 2012b).
Avian responses to fire severity
High and low-severity wildfires have different effects on forest habitat (Bennett et al. 2016), and positive and negative responses to increased fire severity have been recorded among species in avian assemblages in a variety of forests globally (Hutto and Patterson 2016; Knaggs et al. 2020; Franklin et al. 2022). Fire severity refers to the extent to which vegetation is scorched or consumed by fire (Bradstock 2008). Here we include studies that contrast avian responses to one or more levels of fire severity with those in nearby unburnt areas, where vegetation remained intact. Much of what we know about the effects of fire severity on birds is based on the period shortly after fire (Table 2).
| Current knowledge | Implication under climate change | Knowledge gap for priority research | |
|---|---|---|---|
| Short-term avian responses to high-severity fire can be negative, but less is known about medium or long-term responses. | High-severity fire is a strong disturbance, and greater proportions of landscapes will be impacted. | Investigations of avian responses to the time since fire × high-severity fire interaction are required for many areas. | |
| There is some evidence that species responses to high-severity fire persist well into the second decade post-fire. | Habitat loss will have a sustained temporal dimension for these species. | Unchanged bird responses through time suggest that high-severity fires may have lasting, but unknown, effects on habitat that require investigation. | |
| Larger fires are likely to result in more area burnt at high severity, and fires are predicted to increase in frequency. | The amount of forest burnt frequently at high severity will increase. | Little is known about how birds respond to multiple high-severity fires through time in a location, but recent remote sensing methods/products could facilitate this research. | |
| In other systems, particular bird guilds have been identified as vulnerable to high-severity and/or high-frequency fire. | Guild-level understanding has potential to support fire management of dry sclerophyll forest at large spatial scales. | Much more work is required to build understanding of the risks of increased high-severity fire and high fire frequency to dry sclerophyll forest bird guilds. |
See the section Avian responses to fire severity for details and supporting literature.
Short-term responses of bird species and assemblages to increasing levels of fire severity can be negative in particular south-east Australian habitats. Eastern Bristlebirds (Dasyornis brachypterus) were less abundant post-fire in coastal habitat burnt at high severity, which was probably due to the complete loss of the required dense shrub layer (Bain et al. 2008). In a range of habitats in the same region, avian species richness measured 1 year after wildfire declined with increasing fire severity and fire frequency (Lindenmayer et al. 2008). In montane forests, where the dominant trees were obligate-seeding eucalypts, the effects of fire severity on birds varied considerably among species, however, increases in the extent of severely burnt area negatively affected the majority of species (Lindenmayer et al. 2014). In mixed eucalypt forest subject to frequent fire, bird species richness 2–3 years after the most recent fire was 29% greater in sites where only the understorey was burnt, compared with those where the crown was also burnt (Robinson et al. 2014). This was not the case in other parts of the study area where the fire return interval was longer, but in these areas the composition of assemblages and the occurrence of 11 species differed between sites burnt at low and high severity in the last fire.
Particular species are associated with the early post-fire habitat state because of their life history traits, but the level of fire severity influences their occurrence. In eucalypt forests, species such as the Flame Robin (Petroica phoenicia), Buff-rumped Thornbill (Acanthiza reguloides) and Eastern Whipbird (Psophodes olivaceus) may be present in higher numbers 2–3 years after fire in sites where the tree canopy has been either scorched or burnt (Robinson et al. 2014). Flame Robins are altitudinal migrants (Smith et al. 2019) that may colonise recently-burnt forest and so become more abundant shortly after fire (Kelly et al. 2017), because they forage in open areas created by fire (Stokes 1975; Lindenmayer et al. 2014). At 5 years since fire in montane dry sclerophyll forest, the sedentary Variegated Fairy-wren (Malurus lamberti) had a probability of occurrence of ~0.25 where historical high-severity fire had been limited in extent, but was unlikely to occur at all where it had been extensive (Franklin et al. 2022). Numbers of the Superb Lyrebird (Menura novaehollandiae) were greater in areas of eucalypt woodland and open forest where the understorey had been recently removed by a low-intensity hazard reduction burn (Doty et al. 2015). This was presumably because foraging was more efficient in the absence of low-level vegetation structure. In other parts of the world, species such as the Black-backed Woodpecker (Picoides arcticus) depend on food resources in the form of wood-boring beetles that disperse into recently-burnt forest. The post-fire colonisation and persistence of this species was maximal in Californian forest that had burnt recently, and highest levels of colonisation occurred in severely burnt areas (Tingley et al. 2018).
Very little is known about the extent to which avian responses to fire severity persist or change through time since fire, although a limited number of recent studies have addressed this gap. Bird assemblages were similar in patches of North American montane conifer forest that were either unburnt, or very recently burnt to different levels of severity (Tingley et al. 2016). Over the ensuing 10 years, the bird assemblages associated with the original levels of fire severity differentiated, rather than becoming more similar. This may be at least partly explained by the fact that the responses of the vegetation to different fire severities resulted in divergence of plant communities over the period (Tingley et al. 2016). In montane dry sclerophyll forests, both positive and negative responses to historical high-severity fire were observed among species, but species responses did not change over an 11-year period commencing at 5 years since the last fire (Franklin et al. 2022). This result indicates that some effects of high-severity fires on habitat may be persistent. Those species that were less likely to occur where historical high-severity fire had been extensive may be potentially vulnerable to future fire regimes, particularly because their responses did not change through time (Franklin et al. 2022).
Avian guilds and fire severity
When a fire regime shifts towards increasing amounts of high-severity fire, populations of bird species may be threatened, and some of these species may have guild or functional group membership in common. Severe fires are becoming more frequent in the tropical savannas of northern Australia, and the species that are potentially vulnerable are those that (1) use the ground, understorey, or tree hollows, (2) are frugivorous or granivorous, (3) have poor dispersal capacity, and/or (4) exhibit long lifespans and low reproductive rates (Noske 1992; Woinarski and Legge 2013). These attributes are among those that can make species susceptible to high-severity fires in diverse ecosystems. Of all guilds assessed in Sumatran tropical rainforest, understorey insectivores were the only group to be negatively affected by increasing fire severity (Adeney et al. 2006). Species abundance and richness in most foraging guilds in Amazonian forest, including insectivorous guilds that forage on the ground or at low levels in the vegetation, were relatively low at the highest level of fire severity (Barlow and Peres 2004). In North American conifer forests, the richness of tree and primary cavity-nesting bird guilds declined as both patches burnt at high severity increased in size, and as the distance to areas burnt at lower severity increased (Steel et al. 2022).
Recurrent high-severity fire
To support avian diversity and forest ecosystem function more broadly, greater understanding is required about how dry sclerophyll forest bird assemblages are affected by repeated high-severity wildfires through time. The amount and distribution of area burnt at different levels of severity within the spatial extent of wildfires has been shown to have a strong influence on birds (Lindenmayer et al. 2014; Steel et al. 2022). Furthermore, the frequency of fire and variation in the severity of the most recent fire can interact to influence birds (Robinson et al. 2014). Mapping the severity of the entire extent of major wildfires at fine spatial scales (e.g. Gibson et al. 2020) has provided unprecedented opportunities for investigation of the cumulative or longer-term effects of fire severity on components of ecosystems. These data have enabled investigations into the effects of fire regimes on birds that accounted for the contributions of the severity of a chronological series of fires to the extent of historical high-severity fire over large forest patches (Franklin et al. 2022). Attributes of avian habitat such as the density of live trees, distribution of tree hollows, and the amount of coarse woody debris are affected by frequent fires and are likely to be important for particular bird species (Woinarski and Recher 1997; Kelly et al. 2017). However, it is unclear about the extent to which frequent high-severity fires may diminish these aspects of avian habitat (Prior et al. 2016).
Avian responses to fire frequency
The frequency of fire can have important consequences for the biota in natural systems (Bradstock 2008), including the occurrence of forest birds (Kelly et al. 2017). Fires that occur frequently in a location have the potential to disrupt population processes of species that are residents for at least part of the annual cycle (Biodiversity Conservation Act 2016 (NSW), s.4). Following wildfire, the vegetation structure and resources used by forest birds take time to regenerate to resemble their pre-fire state. Changes in the structure and composition of the vegetation may occur if subsequent fires burn these areas before plants are able to reach maturity (Le Breton et al. 2022). The effects of climate change on fire regimes can result in a shift in forest composition toward plants that can tolerate increased fire activity (Fairman et al. 2016; Nolan et al. 2021). This in turn could detrimentally affect bird species that have strong associations with negatively impacted plants and associated vegetation structure (Table 3).
| Current knowledge | Implication under climate change | Knowledge gap for priority research | |
|---|---|---|---|
| Where fire is frequent, locations where the last fire was more severe may hold fewer species. | Alpha, beta, and gamma diversity structures may be simplified in landscapes. | Further work on the interactive effects of fire frequency and fire severity on birds is required. | |
| Avian responses to very high-frequency wildfire are poorly understood in these systems. | Areas burnt at very high-frequency are expected to become more prominent in landscapes. | Species’ tolerance thresholds in relation to very high fire frequency need to be established. | |
| High fire frequency can change vegetation community composition. | Bird species with strong associations with negatively-impacted plants could be detrimentally affected. | Predicted vegetation community changes need to be linked to avian plant associations. | |
| Frequent fires influence the availability of resources used by birds, including hollows, coarse woody debris, and the density of live trees. | The amount of forest frequently burnt at high severity is likely to increase. | Understanding is required on the extent to which frequent high-severity fires can diminish these aspects of avian habitat. |
See the section Avian responses to fire frequency for details and supporting literature.
In eucalypt forest bird assemblages, the responses of species to fire frequency vary according to their habitat requirements and foraging behaviours. In a range of eucalypt forests of south-eastern Australia, the insectivorous Eastern Yellow Robin (Eopsaltria australis) and White-browed Scrubwren (Sericornis frontalis), and the omnivorous Silvereye (Zosterops lateralis) were more common when fire frequency was low (Kelly et al. 2017). There is some evidence that time since fire may buffer positive or negative effects of fire frequency on dry sclerophyll forest birds. For example, at 16 years since fire, there was little difference in the probability of occurrence of species in forest burnt either once or four times in the preceding 31 years (Franklin et al. 2021a). Many common bird species that make up assemblages in eucalypt forest with a history of anthropogenic disturbance may be generally resilient to frequent low-intensity fires that only burn the forest understory, if the fires are small in extent. In Victorian foothill eucalypt forests, of the 27 bird species assessed in relation to variation in the frequency of low-intensity hazard reduction fires, only four species responded significantly, and they were all insectivores (Loyn and McNabb 2015). The inter-fire intervals of the replicated burnt patch treatments for this long-term (27 year) experiment were 3 years and 10 years. The insectivorous Brown Thornbill (Acanthiza pusilla) and Scarlet Robin (Petroica boodang), which forage in the shrub layer and on the ground respectively, were less abundant in the frequently-burnt sites. Australian Magpies (Cracticus tibicen) also forage on open ground but were more common in frequently-burnt patches, as was the Rufous Whistler (Pachycephala rufiventris), which feeds in the tree canopy (Loyn and McNabb 2015).
Despite these few examples, little data is available on the effects of extremely low and high wildfire frequency on dry sclerophyll forest birds, partly because these habitats are currently limited in extent and often located in very remote areas. This presents challenges to those intending to use space-for-time substitution studies to explore the effects of potential extreme fire frequency on birds. This challenge is pressing given that avian communities may face increased exposure to high fire frequencies later in the 21st century. However, detailed, longitudinal studies of the population biology of particular species in frequently-burnt habitat can shed light on local extinction risk for those species (Woinarski and Recher 1997). Brooker and Brooker (1994) predicted that isolated populations of the Splendid Fairy-wren (Malurus splendens), in heathy woodland habitat, would no longer be viable when the area exposed to frequent fire (probability of fire 0.2 per annum) exceeded 2000 ha. This modelling was based on an 18-year study, during which the behaviour and population dynamics of the wrens were measured in relation to rainfall, nest parasitism, nest predation, and a fire regime including multiple fire events (Rowley and Brooker 1987; Brooker and Rowley 1991; Russell and Rowley 1993).
Avian guilds and fire frequency
Fire occurs frequently in particular northern and western Australian ecosystems, and several studies of the effects of fire frequency on biota have revealed some consistent themes in terms of avian responses at the guild level. Frequent fire can detrimentally affect frugivores, granivores, insectivores and ground and hollow nesting species, because of a corresponding reduction in food resources and/or habitat (Yates et al. 2008; Valentine et al. 2012; Woinarski and Legge 2013). In particular, species that are negatively impacted by frequent fire may have requirements for suitably distributed unburnt vegetation, fire-sensitive vegetation and/or diversity of grasses (Woinarski and Legge 2013). For example, granivores and frugivores were not detected in semi-arid Western Australian shrublands with a time since fire of less than 44 years (Davis et al. 2016), so habitat with shorter inter-fire intervals is probably not suitable for these birds. Species with poor dispersal ability and small home ranges are also vulnerable to frequent fire in these and other types of fire-affected ecosystems (Yates et al. 2008; Nimmo et al. 2019). Further research into the vulnerability of species with these traits to increased fire frequency in dry sclerophyll forests is required.
Discussion
We have synthesised knowledge obtained since 2010 about avian responses to fire regimes in dry sclerophyll forest, which can be integrated into management to achieve conservation objectives. Despite these advances, large gaps remain in our understanding about the processes and mechanisms that underpin the fire responses of most bird species (Clarke 2020). Avian responses to fire regimes are context specific (Rainsford et al. 2021) and complex, because factors such as extreme climatic episodes (Connell et al. 2022) and regional land use (Gosper et al. 2019b) interact with fire history to influence avian occurrence. Fire activity is increasing across the distribution of dry sclerophyll forests under climate change (Abram et al. 2021), so continued research in this domain is required to support management of these habitats for avian diversity. For example, the 2019–2020 mega-fires burnt an unprecedented amount of avian habitat, including dry sclerophyll forest, which stimulated focused work on the impacts of those fires on vertebrates (Legge et al. 2021). Many bird species were found to warrant listing or uplisting under national threatened species legislation, based on estimates of population impacts from expert elicitation, and the proportions of their known distribution that were burnt at different levels of severity in that fire season (Legge et al. 2021).
Future fire regimes
Future interactions between fire regime components are likely to be particularly important for dry sclerophyll forest birds, but few studies have been set up to explicitly model interactions between components. More forest is likely to be subject to reduced intervals between high-severity fires, because fires are predicted to increase in frequency (Bradstock 2010), and larger fires are likely to result in greater area burnt at high severity (Collins et al. 2021). Where fire is frequent in mixed eucalypt forest, species richness can be substantially lower where fire is also relatively severe (Robinson et al. 2014), which may ultimately lead to reduced avian diversity across landscapes subject to increased fire activity. Further work on the interactive effects of fire frequency and fire severity on birds in dry sclerophyll forest is required, because populations still recovering from severe fire are likely to be vulnerable to negative effects of short fire return intervals. With time since fire held constant at 16 years, the occurrence of individual species differed only slightly in forest that had been burnt once or four times in the previous 31 years (Franklin et al. 2021a). Fire frequency may be more influential on birds at shorter time since fire, so research that investigates fire frequency effects at different levels of time since fire would be valuable. Increasing time since fire to 16 years did not raise the probability of occurrence of most species that were averse to severely burnt forest at 5 years since fire (Franklin et al. 2022). Therefore, high-severity fire does not necessarily have to be frequent to diminish available habitat across wide areas for affected species.
Consideration of scale in bird–fire research
Birds are highly mobile organisms, and the apparent responses of birds to fire depend on the spatial scale at which predictor variables are expressed (Kelly et al. 2017) and bird responses are measured (Burgess and Maron 2016). Accordingly, study sites, or the areas within which fire history is quantified in relation to the location where bird assemblages are surveyed, need to be of a size and shape that match species home ranges/space use and the ecological effects of fire on the habitat. The responses of vertebrates, including birds, to fire frequency were stronger within a region than when considered across multiple regions (Kelly et al. 2017). Over larger areas it is likely that there is greater variation in other environmental factors that also influence bird distributions. Furthermore, birds respond to the effects of fire on attributes of habitat that exhibit high local variability, such as the average diameter of trees, or leaf litter depth (Kelly et al. 2017). The responses of foraging guilds to fire regimes and environmental attributes may also vary depending on whether bird responses are evaluated and expressed at the site or landscape scale. For instance, at the landscape scale, the extent of habitat with time since fire of >10 years had the strongest influence on richness of insectivores, frugivores and canopy species, but range in elevation was the best predictor of richness of these groups at the patch scale (Burgess and Maron 2016).
Unknown effects of fire regimes on rare and uncommon birds
The long-term preservation of species is the fundamental aim in conservation management at local, regional and global scales, but many species are rare, necessitating targeted approaches to monitoring their distributions (Jeliazkov et al. 2022). When a population is small and a species occurs at low levels in the landscape, there is potential for the attribution of null fire responses to that species, which are inaccurate in a broader context (Connell et al. 2022). To better understand the risk of extinction of rare species under changing fire regimes, study designs should be based on what is known about individual species ecology. For example, call playback can be an effective method of detecting rare species in vegetation communities they are known to occupy (Brown et al. 2009).
The distribution of a rare species in a landscape is influenced by the combination of its local density, and the extent to which it is spatially dispersed or clumped within a narrow or broad range, hence species can be characterised by their type of rarity (Rabinowitz 1981). Tailored sampling and modelling approaches should be incorporated in designs of studies focused on rare species responses to fire, depending on the type of rarity (Jeliazkov et al. 2022). For example, random sampling across a region may not yield sufficient observations to model fire responses of a rare species with a spatially clumped distribution (Jeliazkov et al. 2022). To maximise the effectiveness of sampling effort and minimise research costs, the methodology for data collection needs to be optimal for the target species. For example, programmable acoustic recorders can capture extended temporal samples in the field at very low cost once deployed, thus increasing the potential for detection of rare species through their calls (Franklin et al. 2021b). This functionality is complemented by the ongoing, rapid development of methods for automated processing of large acoustic datasets to identify individual species (Darras et al. 2019).
Genetic analysis of rare and uncommon bird species in fire-prone forests has the potential to provide valuable information to support management of their populations, but these methods have been underutilised to date. A limited number of valuable studies of population genetics in relation to fire have been conducted on amphibians (Potvin et al. 2017), mammals (Pereoglou et al. 2013; Banks et al. 2017), reptiles (Neuwald and Templeton 2013; Smith et al. 2014) and birds (Brown et al. 2013). Genetic data can be used to infer the source locations of individuals recolonising burnt areas, which may shed light on species dispersal and movement patterns in fire-affected landscapes (Brown et al. 2013; Pereoglou et al. 2013; Banks et al. 2017). Furthermore, genetic information has potential to inform whether sub- populations are functionally connected or isolated (Sitters and Di Stefano 2020). Genetic analyses may also reveal underlying population trends, such as inbreeding, that may be masked when post-fire surveys suggest population stability (Potvin et al. 2017). These applications have particular relevance for dry sclerophyll forest birds, because proportions of habitat burnt recently, severely and frequently have recently increased across the distribution of this vegetation formation (Fairman et al. 2016; Collins et al. 2021; Franklin et al. 2023).
Fire management for avian diversity
To conserve biodiversity, fire management and planning for natural landscapes has often aimed to maintain a mosaic of patches with contrasting fire histories. However, this approach potentially lacks specificity and may achieve sub-optimal results (Bradstock et al. 2005; Taylor et al. 2012). For example, the responses of many of the avifauna of semi-arid, multi-stemmed (mallee) eucalypt ecosystems have shown that a higher proportion of older vegetation, rather than an even spatial distribution of a range of fire-age classes, would better support avian diversity (Taylor et al. 2012, 2013; Connell et al. 2017). Greater species richness (Taylor et al. 2012), relative abundance (Kelly et al. 2015), and probability of occurrence of many species, including a number of threatened species (Connell et al. 2017), were associated with longer time since fire. In addition to requiring long unburnt vegetation, a number of species were also restricted in their distributions to areas with particular climate and vegetation characteristics (Connell et al. 2017). Recent work has raised awareness of the importance of dry sclerophyll forest at mid-range and longer time since fire for many species in the avian assemblages occupying these habitats (Kelly et al. 2017; Franklin et al. 2021a, 2022). For instance, of 79 bird species studied in relation to fire, 18% were averse to forest where time since fire was short (Franklin et al. 2022), and/or where there was an absence of long unburnt forest in the surrounding landscape (Franklin et al. 2021a). Larger, more frequent fires threaten these species, because such fires will increase the proportion of forest habitat at short fire ages in landscapes, which may remain in an unsuitable state for many years.
Fire management and planning now need to consider that proportions of dry sclerophyll forest with longer fire ages are declining in south-east Australian landscapes, but many forest birds require habitat at longer time since fire. We argue that there is a need to sustain substantial, functionally connected areas of long unburnt forest across landscapes, that are otherwise undisturbed by practices such as timber harvesting. Proportions of these areas would ideally be burnt infrequently and at low severity, for the purpose of biodiversity conservation. The increases in fire activity that are currently underway will provide for bird species with requirements for early successional, and frequently or severely burnt habitat, that could be actively managed, as required. The 2019–2020 mega-fires in the forests of south-eastern Australia were evidence of increased fire activity driven by climate change (Abram et al. 2021). As a result of these fires, vast areas of forest estate in the region currently have a fire age of 3 years, which presents challenges for management that aims to support species populations and biodiversity (Legge et al. 2022). Forest canopy structure is less complex at short time since fire, but many forest bird species require the denser canopies associated with longer time since fire (Franklin et al. 2023). Many bird species respond to the effects of time since fire on habitat structure instead of time since fire per se, so spatial definition of both habitat structure and fire history would enhance specificity of avian fire management (Sitters et al. 2014; Lindenmayer et al. 2016; Franklin et al. 2023). Avian responses to fire can differ under periods of drought and above-average rainfall (Connell et al. 2022). Such knowledge presents opportunities for nuanced fire management to support biodiversity. For example, management fires could be scheduled to occur during periods when climatic conditions are favourable for avian populations (Clarke 2008).
Conclusion
The need to consider the complexity of the effects of multiple fires through time on species and their communities in fire management was raised early in the 21st century (Bradstock et al. 2005). Since that time, much research has been conducted and knowledge gained about responses of birds to the effects of separate or combined components of fire regimes at a range of temporal and spatial scales (e.g. Kelly et al. 2017). Fine-scale mapping of fire characteristics, remote sensing of habitat structure, acoustic recorders for rare species and landscape genetics show considerable promise as tools in this research domain. Some forest bird species may respond positively to increased levels of one or more components of a fire regime, and negatively to greater levels of others (e.g. Franklin et al. 2021a). Therefore, attempting to characterise species as either fire-loving or fire-averse is an over-simplification that does not capture the spectrum of species responses to fire regimes. There is much that is still unknown about the implications of changing fire regimes for birds (Clarke 2020), and more options for resource provision are available to management with long unburnt forest than there are with recently-burnt habitat (Gosper et al. 2019a).
Data availability
Data sharing is not applicable as no new data were generated or analysed during this study.
Declaration of funding
MF was supported by an Australian Government Research Training Program (RTP) PhD Scholarship through the University of Wollongong.
References
Abram NJ, Henley BJ, Sen Gupta A, Lippmann TJR, Clarke H, Dowdy AJ, Sharples JJ, Nolan RH, Zhang T, Wooster MJ, Wurtzel JB, Meissner KJ, Pitman AJ, Ukkola AM, Murphy BP, Tapper NJ, Boer MM (2021) Connections of climate change and variability to large and extreme forest fires in southeast Australia. Communications Earth & Environment 2, 8.
| Crossref | Google Scholar |
Adeney JM, Ginsberg JR, Russell GJ, Kinnaird MF (2006) Effects of an ENSO-related fire on birds of a lowland tropical forest in Sumatra. Animal Conservation 9, 292-301.
| Crossref | Google Scholar |
Bain DW, Baker JR, French KO, Whelan RJ (2008) Post-fire recovery of eastern bristlebirds (Dasyornis brachypterus) is context-dependent. Wildlife Research 35, 44-49.
| Crossref | Google Scholar |
Banks SC, McBurney L, Blair D, Davies ID, Lindenmayer DB (2017) Where do animals come from during post-fire population recovery? Implications for ecological and genetic patterns in post-fire landscapes. Ecography 40, 1325-1338.
| Crossref | Google Scholar |
Barlow J, Peres CA (2004) Avifaunal responses to single and recurrent wildfires in Amazonian forests. Ecological Applications 14, 1358-1373.
| Crossref | Google Scholar |
Bennett LT, Bruce MJ, MacHunter J, Kohout M, Tanase MA, Aponte C (2016) Mortality and recruitment of fire-tolerant eucalypts as influenced by wildfire severity and recent prescribed fire. Forest Ecology and Management 380, 107-117.
| Crossref | Google Scholar |
Berry LE, Lindenmayer DB, Driscoll DA (2015) Large unburnt areas, not small unburnt patches, are needed to conserve avian diversity in fire-prone landscapes. Journal of Applied Ecology 52, 486-495.
| Crossref | Google Scholar |
Bond WJ, Keeley JE (2005) Fire as a global ‘herbivore’: the ecology and evolution of flammable ecosystems. Trends in Ecology & Evolution 20, 387-394.
| Crossref | Google Scholar | PubMed |
Bradstock RA (2008) Effects of large fires on biodiversity in south-eastern Australia: disaster or template for diversity? International Journal of Wildland Fire 17, 809-822.
| Crossref | Google Scholar |
Bradstock RA (2010) A biogeographic model of fire regimes in Australia: current and future implications. Global Ecology and Biogeography 19, 145-158.
| Crossref | Google Scholar |
Bradstock RA, Kenny BJ (2003) An application of plant functional types to fire management in a conservation reserve in southeastern Australia. Journal of Vegetation Science 14, 345-354.
| Crossref | Google Scholar |
Bradstock RA, Bedward M, Gill AM, Cohn JS (2005) Which mosaic? A landscape ecological approach for evaluating interactions between fire regimes, habitat and animals. Wildlife Research 32, 409-423.
| Crossref | Google Scholar |
Brooker LC, Brooker MG (1994) A model for the effects of fire and fragmentation on the population viability of the splendid fairy-wren. Pacific Conservation Biology 1, 344-358.
| Crossref | Google Scholar |
Brooker MG, Rowley I (1991) Impact of wildfire on the nesting behaviour of birds in heathland. Wildlife Research 18, 249-263.
| Crossref | Google Scholar |
Brown S, Clarke M, Clarke R (2009) Fire is a key element in the landscape-scale habitat requirements and global population status of a threatened bird: the Mallee Emu-wren (Stipiturus mallee). Biological Conservation 142, 432-445.
| Crossref | Google Scholar |
Brown SM, Harrisson KA, Clarke RH, Bennett AF, Sunnucks P (2013) Limited population structure, genetic drift and bottlenecks characterise an endangered bird species in a dynamic, fire-prone ecosystem. PLoS ONE 8, e59732.
| Crossref | Google Scholar | PubMed |
Burgess EE, Maron M (2016) Does the response of bird assemblages to fire mosaic properties vary among spatial scales and foraging guilds? Landscape Ecology 31, 687-699.
| Crossref | Google Scholar |
Clarke MF (2008) Catering for the needs of fauna in fire management: science or just wishful thinking? Wildlife Research 35, 385-394.
| Crossref | Google Scholar |
Clarke MF (2020) Our birds will be ok, they’ve evolved to cope with fire…haven’t they? Emu - Austral Ornithology 120, 184-186.
| Crossref | Google Scholar |
Collins L, Bradstock RA, Clarke H, Clarke MF, Nolan RH, Penman TD (2021) The 2019/2020 mega-fires exposed Australian ecosystems to an unprecedented extent of high-severity fire. Environmental Research Letters 16, 044029.
| Crossref | Google Scholar |
Connell J, Watson SJ, Taylor RS, Avitabile SC, Clarke RH, Bennett AF, Clarke MF (2017) Testing the effects of a century of fires: requirements for post-fire succession predict the distribution of threatened bird species. Diversity and Distributions 23, 1078-1089.
| Crossref | Google Scholar |
Connell J, Hall MA, Nimmo DG, Watson SJ, Clarke MF (2022) Fire, drought and flooding rains: the effect of climatic extremes on bird species’ responses to time since fire. Diversity and Distributions 28, 417-438.
| Crossref | Google Scholar |
Darras K, Batáry P, Furnas BJ, Grass I, Mulyani YA, Tscharntke T (2019) Autonomous sound recording outperforms human observation for sampling birds: a systematic map and user guide. Ecological Applications 29, e01954.
| Crossref | Google Scholar | PubMed |
Davis RA, Doherty TS, van Etten EJB, Radford JQ, Holmes F, Knuckey C, Davis BJ (2016) Conserving long unburnt vegetation is important for bird species, guilds and diversity. Biodiversity and Conservation 25, 2709-2722.
| Crossref | Google Scholar |
de Groot WJ, Flannigan MD, Cantin AS (2013) Climate change impacts on future boreal fire regimes. Forest Ecology and Management 294, 35-44.
| Crossref | Google Scholar |
Doherty TS, Geary WL, Jolly CJ, Macdonald KJ, Miritis V, Watchorn DJ, Cherry MJ, Conner LM, González TM, Legge SM, Ritchie EG, Stawski C, Dickman CR (2022) Fire as a driver and mediator of predator–prey interactions. Biological Reviews 97, 1539-1558.
| Crossref | Google Scholar | PubMed |
Doty A, Stawski C, Nowack J, Bondarenco A, Geiser F (2015) Increased lyrebird presence in a post-fire landscape. Australian Journal of Zoology 63, 9-11.
| Crossref | Google Scholar |
Dowdy AJ, Ye H, Pepler A, Thatcher M, Osbrough SL, Evans JP, Di Virgilio G, McCarthy N (2019) Future changes in extreme weather and pyroconvection risk factors for Australian wildfires. Scientific Reports 9, 10073.
| Crossref | Google Scholar |
Driscoll DA, Lindenmayer DB, Bennett AF, Bode M, Bradstock RA, Cary GJ, Clarke MF, Dexter N, Fensham R, Friend G, Gill M, James S, Kay G, Keith DA, MacGregor C, Russell-Smith J, Salt D, Watson JEM, Williams RJ, York A (2010) Fire management for biodiversity conservation: key research questions and our capacity to answer them. Biological Conservation 143, 1928-1939.
| Crossref | Google Scholar |
Dupuy J-L, Fargeon H, Martin-StPaul N, Pimont F, Ruffault J, Guijarro M, Hernando C, Madrigal J, Fernandes P (2020) Climate change impact on future wildfire danger and activity in southern Europe: a review. Annals of Forest Science 77, 35.
| Crossref | Google Scholar |
Fairman TA, Nitschke CR, Bennett LT (2016) Too much, too soon? A review of the effects of increasing wildfire frequency on tree mortality and regeneration in temperate eucalypt forests. International Journal of Wildland Fire 25, 831-848.
| Crossref | Google Scholar |
Franklin MJM, Major RE, Bedward M, Bradstock RA (2021a) Relative avian mobility linked to use of fire-affected resources in forested landscapes. Forest Ecology and Management 497, 119484.
| Crossref | Google Scholar |
Franklin MJM, Major RE, Bradstock RA (2021b) How much survey effort is required to assess bird assemblages in fire-prone eucalypt forests using acoustic recorders? Wildlife Research 48, 414-421.
| Crossref | Google Scholar |
Franklin MJM, Major RE, Bedward M, Price OF, Bradstock RA (2022) Forest avifauna exhibit enduring responses to historical high-severity wildfires. Biological Conservation 269, 109545.
| Crossref | Google Scholar |
Franklin MJM, Major RE, Bradstock RA (2023) Canopy cover mediates the effects of a decadal increase in time since fire on arboreal birds. Biological Conservation 277, 109871.
| Crossref | Google Scholar |
Gibson R, Danaher T, Hehir W, Collins L (2020) A remote sensing approach to mapping fire severity in south-eastern Australia using sentinel 2 and random forest. Remote Sensing of Environment 240, 111702.
| Crossref | Google Scholar |
Gill AM (1975) Fire and the Australian flora: a review. Australian Forestry 38, 4-25.
| Crossref | Google Scholar |
Gosper CR, Fox E, Burbidge AH, Craig MD, Douglas TK, Fitzsimons JA, McNee S, Nicholls AO, O’Connor J, Prober SM, Watson DM, Watson SJ, Yates CJ (2019a) Multi-century periods since fire in an intact woodland landscape favour bird species declining in an adjacent agricultural region. Biological Conservation 230, 82-90.
| Crossref | Google Scholar |
Gosper CR, Watson SJ, Fox E, Burbidge AH, Craig MD, Douglas TK, Fitzsimons JA, McNee S, Nicholls AO, O’Connor J, Prober SM, Watson DM, Yates CJ (2019b) Fire-mediated habitat change regulates woodland bird species and functional group occurrence. Ecological Applications 29, e01997.
| Crossref | Google Scholar |
Green K (2013) The effect of fire on the avifauna of subalpine woodland in the snowy mountains. The Victorian Naturalist 130, 240-248.
| Google Scholar |
Haslem A, Avitabile SC, Taylor RS, Kelly LT, Watson SJ, Nimmo DG, Kenny SA, Callister KE, Spence-Bailey LM, Bennett AF, Clarke MF (2012) Time-since-fire and inter-fire interval influence hollow availability for fauna in a fire-prone system. Biological Conservation 152, 212-221.
| Crossref | Google Scholar |
Hutto RL, Patterson DA (2016) Positive effects of fire on birds may appear only under narrow combinations of fire severity and time-since-fire. International Journal of Wildland Fire 25, 1074-1085.
| Crossref | Google Scholar |
Jeliazkov A, Gavish Y, Marsh CJ, Geschke J, Brummitt N, Rocchini D, Haase P, Kunin WE, Henle K (2022) Sampling and modelling rare species: conceptual guidelines for the neglected majority. Global Change Biology 28, 3754-3777.
| Crossref | Google Scholar | PubMed |
Jolly CJ, Dickman CR, Doherty TS, van Eeden LM, Geary WL, Legge SM, Woinarski JCZ, Nimmo DG (2022) Animal mortality during fire. Global Change Biology 28, 2053-2065.
| Crossref | Google Scholar | PubMed |
Karna YK, Penman TD, Aponte C, Hinko-Najera N, Bennett LT (2020) Persistent changes in the horizontal and vertical canopy structure of fire-tolerant forests after severe fire as quantified using multi-temporal airborne lidar data. Forest Ecology and Management 472, 118255.
| Crossref | Google Scholar |
Kelly LT, Bennett AF, Clarke MF, McCarthy MA (2015) Optimal fire histories for biodiversity conservation. Conservation Biology 29, 473-481.
| Crossref | Google Scholar | PubMed |
Kelly LT, Haslem A, Holland GJ, Leonard SWJ, Machunter J, Bassett M, Bennett AF, Bruce MJ, Chia EK, Christie FJ, Clarke MF, Di Stefano J, Loyn R, McCarthy MA, Pung A, Robinson N, Sitters H, Swan M, York A (2017) Fire regimes and environmental gradients shape vertebrate and plant distributions in temperate eucalypt forests. Ecosphere 8, e01781.
| Crossref | Google Scholar |
Knaggs M, Haché S, Nielsen SE, Pankratz RF, Bayne E (2020) Avian response to wildfire severity in a northern boreal region. Forests 11, 1330.
| Crossref | Google Scholar |
Le Breton TD, Lyons MB, Nolan RH, Penman T, Williamson GJ, Ooi MKJ (2022) Megafire-induced interval squeeze threatens vegetation at landscape scales. Frontiers in Ecology and the Environment 20, 327-334.
| Crossref | Google Scholar |
Legge S, Rumpff L, Woinarski JCZ, Whiterod NS, Ward M, Southwell DG, Scheele BC, Nimmo DG, Lintermans M, Geyle HM, et al. (2022) The conservation impacts of ecological disturbance: time-bound estimates of population loss and recovery for fauna affected by the 2019–2020 Australian megafires. Global Ecology and Biogeography 31, 2085-2104.
| Crossref | Google Scholar |
Lindenmayer DB, Wood JT, Cunningham RB, Macgregor C, Crane M, Michael D, Montague-Drake R, Brown D, Muntz R, Gill AM (2008) Testing hypotheses associated with bird responses to wildfire. Ecological Applications 18, 1967-1983.
| Crossref | Google Scholar | PubMed |
Lindenmayer DB, Blanchard W, McBurney L, Blair D, Banks SC, Driscoll DA, Smith AL, Gill A (2014) Complex responses of birds to landscape-level fire extent, fire severity and environmental drivers. Diversity and Distributions 20, 467-477.
| Crossref | Google Scholar |
Lindenmayer DB, Candy SG, MacGregor CI, Banks SC, Westgate M, Ikin K, Pierson J, Tulloch A, Barton P (2016) Do temporal changes in vegetation structure additional to time since fire predict changes in bird occurrence? Ecological Applications 26, 2267-2279.
| Crossref | Google Scholar | PubMed |
Loyn RH (1997) Effects of an extensive wildfire on birds in far eastern Victoria. Pacific Conservation Biology 3, 221-234.
| Crossref | Google Scholar |
Loyn RH, McNabb EG (2015) Bird population responses to wildfire and planned burns in foothill forests of Victoria, Australia. Journal of Ornithology 156, 263-273.
| Crossref | Google Scholar |
Neuwald JL, Templeton AR (2013) Genetic restoration in the eastern collared lizard under prescribed woodland burning. Molecular Ecology 22, 3666-3679.
| Crossref | Google Scholar | PubMed |
Nimmo DG, Avitabile S, Banks SC, Bliege Bird R, Callister K, Clarke MF, Dickman CR, Doherty TS, Driscoll DA, Greenville AC, Haslem A, Kelly LT, Kenny SA, Lahoz-Monfort JJ, Lee C, Leonard S, Moore H, Newsome TM, Parr CL, Ritchie EG, Schneider K, Turner JM, Watson S, Westbrooke M, Wouters M, White M, Bennett AF (2019) Animal movements in fire-prone landscapes. Biological Reviews 94, 981-998.
| Crossref | Google Scholar | PubMed |
Nolan RH, Collins L, Leigh A, Ooi MKJ, Curran TJ, Fairman TA, Resco de Dios V, Bradstock R (2021) Limits to post-fire vegetation recovery under climate change. Plant, Cell & Environment 44, 3471-3489.
| Crossref | Google Scholar | PubMed |
Noske RA (1992) The status and ecology of the white-throated grasswren Amytornis woodwardi. Emu - Austral Ornithology 92, 39-51.
| Crossref | Google Scholar |
Pereoglou F, Lindenmayer DB, MacGregor C, Ford F, Wood J, Banks SC (2013) Landscape genetics of an early successional specialist in a disturbance-prone environment. Molecular Ecology 22, 1267-1281.
| Crossref | Google Scholar | PubMed |
Potvin DA, Parris KM, Smith Date KL, Keely CC, Bray RD, Hale J, Hunjan S, Austin JJ, Melville J (2017) Genetic erosion and escalating extinction risk in frogs with increasing wildfire frequency. Journal of Applied Ecology 54, 945-954.
| Crossref | Google Scholar |
Prior LD, Williamson GJ, Bowman DMJS (2016) Impact of high-severity fire in a Tasmanian dry eucalypt forest. Australian Journal of Botany 64, 193-205.
| Crossref | Google Scholar |
Prowse TAA, Collard SJ, Blackwood A, O’Connor PJ, Delean S, Barnes M, Cassey P, Possingham HP (2017) Prescribed burning impacts avian diversity and disadvantages woodland-specialist birds unless long-unburnt habitat is retained. Biological Conservation 215, 268-276.
| Crossref | Google Scholar |
Rainsford FW, Kelly LT, Leonard SWJ, Bennett AF (2021) Post-fire habitat relationships for birds differ among ecosystems. Biological Conservation 260, 109218.
| Crossref | Google Scholar |
Reside AE, Vanderwal J, Kutt A, Watson I, Williams S (2012) Fire regime shifts affect bird species distributions. Diversity and Distributions 18, 213-225.
| Crossref | Google Scholar |
Robinson NM, Leonard SWJ, Bennett AF, Clarke MF (2014) Refuges for birds in fire-prone landscapes: the influence of fire severity and fire history on the distribution of forest birds. Forest Ecology and Management 318, 110-121.
| Crossref | Google Scholar |
Russell EM, Rowley I (1993) Demography of the cooperatively breeding Splendid Fairy-wren, malurus-splendens (Maluridae). Australian Journal of Zoology 41, 475-505.
| Crossref | Google Scholar |
Sitters H, Di Stefano J (2020) Integrating functional connectivity and fire management for better conservation outcomes. Conservation Biology 34, 550-560.
| Crossref | Google Scholar | PubMed |
Sitters H, Christie F, Di Stefano J, Swan M, Collins P, York A (2014) Associations between occupancy and habitat structure can predict avian responses to disturbance: implications for conservation management. Forest Ecology and Management 331, 227-236.
| Crossref | Google Scholar |
Smith AL, Bull CM, Gardner MG, Driscoll DA (2014) Life history influences how fire affects genetic diversity in two lizard species. Molecular Ecology 23, 2428-2441.
| Crossref | Google Scholar | PubMed |
Steel ZL, Fogg AM, Burnett R, Roberts LJ, Safford HD (2022) When bigger isn’t better—implications of large high-severity wildfire patches for avian diversity and community composition. Diversity and Distributions 28, 439-453.
| Crossref | Google Scholar |
Stokes T (1975) The effect of a bushfire on the banding of Flame Robins in the Brindabella Ranges. Australian Bird Bander 13, 75-76.
| Google Scholar |
Taylor RS, Watson SJ, Nimmo DG, Kelly LT, Bennett AF, Clarke MF (2012) Landscape-scale effects of fire on bird assemblages: does pyrodiversity beget biodiversity? Diversity and Distributions 18, 519-529.
| Crossref | Google Scholar |
Taylor RS, Watson SJ, Bennett AF, Clarke MF (2013) Which fire management strategies benefit biodiversity? A landscape-perspective case study using birds in mallee ecosystems of south-eastern Australia. Biological Conservation 159, 248-256.
| Crossref | Google Scholar |
Tingley MW, Ruiz-Gutiérrez V, Wilkerson RL, Howell CA, Siegel RB (2016) Pyrodiversity promotes avian diversity over the decade following forest fire. Proceedings of the Royal Society B: Biological Sciences 283, 20161703.
| Crossref | Google Scholar |
Tingley MW, Stillman AN, Wilkerson RL, Howell CA, Sawyer SC, Siegel RB (2018) Cross-scale occupancy dynamics of a postfire specialist in response to variation across a fire regime. Journal of Animal Ecology 87, 1484-1496.
| Crossref | Google Scholar | PubMed |
Turner R (1992) Effect of wildfire on birds at Weddin Mountain, New South Wales. Corella 16, 65-74.
| Google Scholar |
Valentine LE, Schwarzkopf L, Johnson CN (2012) Effects of a short fire-return interval on resources and assemblage structure of birds in a tropical savanna. Austral Ecology 37, 23-34.
| Crossref | Google Scholar |
Valentine LE, Fisher R, Wilson BA, Sonneman T, Stock WD, Fleming PA, Hobbs RJ (2014) Time since fire influences food resources for an endangered species, Carnaby’s cockatoo, in a fire-prone landscape. Biological Conservation 175, 1-9.
| Crossref | Google Scholar |
Ward M, Tulloch AIT, Radford JQ, Williams BA, Reside AE, Macdonald SL, Mayfield HJ, Maron M, Possingham HP, Vine SJ, O’Connor JL, Massingham EJ, Greenville AC, Woinarski JCZ, Garnett ST, Lintermans M, Scheele BC, Carwardine J, Nimmo DG, Lindenmayer DB, Kooyman RM, Simmonds JS, Sonter LJ, Watson JEM (2020) Impact of 2019–2020 mega-fires on Australian fauna habitat. Nature Ecology & Evolution 4, 1321-1326.
| Crossref | Google Scholar | PubMed |
Watson SJ, Taylor RS, Nimmo DG, Kelly LT, Haslem A, Clarke MF, Bennett AF (2012a) Effects of time since fire on birds: how informative are generalized fire response curves for conservation management? Ecological Applications 22, 685-696.
| Crossref | Google Scholar | PubMed |
Watson SJ, Taylor RS, Nimmo DG, Kelly LT, Clarke MF, Bennett AF (2012b) The influence of unburnt patches and distance from refuges on post-fire bird communities. Animal Conservation 15, 499-507.
| Crossref | Google Scholar |
Whelan RJ, Rodgerson L, Dickman CR, Sutherland EF (2002) Critical life cycles of plants and animals: Developing a process-based understanding of population changes in fire-prone landscapes. In ‘Flammable Australia: the fire regimes and biodiversity of a continent’. (Eds RA Bradstock, JE Williams, AM Gill) pp. 94–124. (Cambridge University press: Cambridge, UK)
Williamson GJ, Prior LD, Grose MR, Harris RMB, Bowman DMJS (2014) Projecting canopy cover change in Tasmanian eucalypt forests using dynamically downscaled regional climate models. Regional Environmental Change 14, 1373-1386.
| Crossref | Google Scholar |
Woinarski JCZ, Legge S (2013) The impacts of fire on birds in Australia’s tropical savannas. Emu - Austral Ornithology 113, 319-352.
| Crossref | Google Scholar |
Woinarski JCZ, Recher HF (1997) Impact and response: a review of the effects of fire on the Australian avifauna. Pacific Conservation Biology 3, 183-205.
| Crossref | Google Scholar |
Yates CP, Edwards AC, Russell-Smith J (2008) Big fires and their ecological impacts in Australian savannas: size and frequency matters. International Journal of Wildland Fire 17, 768-781.
| Crossref | Google Scholar |