

Temporal changes in habitat use by dugongs in a spatially restricted coral reef environment
Christophe Cleguer A B C * , Claire Garrigue B C , Mariana M. P. B. Fuentes A D , Mark Hamann A , Claude Payri B and Helene Marsh A
A B C * , Claire Garrigue B C , Mariana M. P. B. Fuentes A D , Mark Hamann A , Claude Payri B and Helene Marsh A
A
B
C
D
Abstract
The dynamic habitat use by dugongs (Dugong dugon) in small coral reef lagoon systems spatially limited by tides is not well understood and has hampered adaptive management.
We investigated how dugongs locally used a high conservation value coral-reef lagoon system during different seasons and tides to support local management.
We conducted local-scale aerial surveys to document the seasonal and tidal changes in the distribution and number of dugongs over seagrass and non-seagrass coral reef habitats in a small lagoon in New Caledonia. The surveys were conducted fortnightly over 18 months in different seasons and at different tides. Temperature loggers and existing local footage of dugong herding behaviour were used to study the habitat use and behaviour of the animals in the area.
More dugongs were sighted in Cap Goulvain during the cool season than during the warm season. As tides restricted access to the intertidal seagrass meadows and during the cool season, more dugongs were sighted outside the lagoon on the fore reef shelf outside the lagoon. Dugongs were resting in large aggregations during their use of this non-seagrass habitat.
Our study emphasises the importance of non-seagrass habitats for dugongs in spatially restricted coral reef environments as well as the importance of considering outer lagoon habitats as key dugong management areas. This study also adds evidence of behavioural thermoregulation in dugongs.
The development of dynamic management strategies is essential, especially where different habitats should be protected at different times of the year.
Keywords: aerial survey, conservation, coral reefs, dugong, ecology, habitat use, marine mammal, thermoregulation.
Introduction
Understanding changes in habitat use by wildlife species and the mechanisms that affect these changes is a primary goal of ecologists and resource managers (Jefferson et al. 1993). Changes in habitat use by wildlife species generally reflect requirements for feeding and reproduction, the need to avoid predation, and response to changes in the physical environment such as climatic variations (Sih 1980; Bjørge 2002; Hamel and Côté 2008). These requirements often result in trade-offs and changes in distribution and abundance, as it is unusual for a single region to provide all of the resources needed for a species. For example, seals shift to solid substrate to escape predators, for thermoregulatory purposes, and/or to rest (Norris and Kunz 2012).
Changes in habitat use may take place at a range of spatial and temporal scales, from the daily requirements for adjacent habitats to seasonal use of geographically separated environments (Krausman 1999). Wildlife species inhabiting expansive geographical regions may exhibit divergent habitat preferences or utilise similar habitats for distinct needs, owing to the diverse geographical features present within their range. For example, in the western South Atlantic some juvenile green turtles (Chelonia mydas) make seasonal shifts between neritic and oceanic habitats (González Carman et al. 2014). One of the key challenges to ecologists and resource managers is to understand wildlife species distribution and habitat use at several spatial and temporal scales. Specifically, the careful examination of habitat use by coastal marine mammals on a fine spatial and temporal level is crucial to effectively manage and reduce conflicts arising from human activities along coastlines and their impact on wildlife.
The range of the dugong (Dugong dugon) spans the tropical and subtropical coastal and island waters of the Indo-West Pacific, extending from East Africa to Vanuatu. Their distribution is primarily coastal and closely associated with the presence of seagrasses, which constitute their main food source (Marsh et al. 2012). Due to their coastal lifestyle, dugongs are susceptible to interactions with human activities, and their seagrass habitats are experiencing global declines as a result of cumulative coastal pressures caused by human activities (Marsh et al. 2012). Therefore, it is crucial to comprehend the dynamics of dugong habitat utilisation in order to effectively guide conservation and management efforts for this species.
The dynamics of dugong habitat use have mainly been studied in Australian coastal waters using tracking devices (e.g. Sheppard et al. 2006), aerial surveys for abundance and distribution assessments (see Cleguer and Marsh (2023) for an inventory and review), cameras mounted onto air balloons (Hodgson 2004) and more recently drones (Cleguer et al. 2021). Put together, these studies show that the way dugongs interact with their habitat will vary with location, time of the year, and the size and heterogeneity of the habitats in which they are found (see Deutsch et al. (2022) for review). For example, the dugongs’ habitat use is largely influenced by spatial heterogeneity and temporal variations in the abundance and species composition and nutritional value of their foraging habitats, as well as tides (e.g. Sheppard et al. 2009; Derville et al. 2022), water temperature (e.g. Preen 1992; Holley et al. 2006; Zeh et al. 2018), sea state (Marsh et al. 2012) and predation (e.g. Wirsing et al. 2007a, 2007b).
Although certain dugong populations are known or suspected to reside in specific areas throughout the year, this does not exclude the occurrence of seasonal changes in distribution at a smaller scale. In locations at the higher latitudinal boundaries of the dugong’s range, such as Saudi Arabia (Preen 2004), south-western Australia (Anderson 1986; Holley 2006; Holley et al. 2006), and south-eastern Australia (Preen 1992; Lanyon 2003; Sheppard et al. 2006; Zeh et al. 2018), seasonal temperature variations lead to corresponding changes in dugong distribution, movements, and habitat utilisation. In regions where suitable habitats differ across seasons and are geographically distant from each other, dugongs may undertake seasonal migrations (e.g. Holley et al. 2006). In other locations, critical resources that cannot be substituted may be located in close proximity to each other, and thus most dugongs may then preferentially utilise those resources by making relatively short movements between habitats, demonstrating a form of landscape complementation (Dunning et al. 1992).
The geomorphological characteristics of the regions of Australia where dugong habitat use have been studied may be different from other parts of the dugong’s range. Australian studies on dugongs have been conducted where seagrass meadows are distributed over vast intertidal and subtidal areas (i.e. up to several thousand km), often associated with coral reefs (Coles et al. 2003; Taylor and Rasheed 2010; Marsh et al. 2012). In contrast in Okinawa (Japan), Palau, Mayotte, Madagascar, and the Pacific Islands such as New Caledonia, dugongs are observed in locations where the continental shelf is narrow (i.e. tens of kilometres) or nonexistent (Cleguer et al. 2015). In these regions, seagrasses are found on coral reef flats in lagoons protected by tropical barrier reefs (Short et al. 2007). The width of these lagoons ranges from a few kilometres to tens of kilometres (Pinet 2011). In these spatially restricted environments seagrass is often limited to small intertidal areas, which restricts dugongs’ access to their food resources during high tides. Despite previous work on dugong habitat use, little is known about how seasonal and tidal changes affect habitat use by dugongs in these environments.
To address this knowledge gap, this study explored how seasonal and tidal changes affect dugong habitat use in a spatially restricted coral reef lagoon of Cap Goulvain, an important dugong and seagrass coral reef lagoon area located on the central-west coast of the main island of New Caledonia (Cleguer et al. 2015). The New Caledonian dugong population is geographically and genetically isolated from other subpopulations within the dugong’s large range (Garrigue et al. 2022). The number of mature individuals in the subpopulation is estimated to range between 149 and 896 individuals based on aerial surveys of New Caledonia from 2003 to 2012 (Garrigue et al. 2008; Cleguer et al. 2017) and the population has recently been classified as ‘Endangered’ under the IUCN Red List of Threatened Species (Hamel et al. 2022).
We used a combination of local-scale line transect aerial surveys conducted from an ultra-light plane over an 18 month period at both low and high tide to document the seasonal and tidal changes in the distribution and number of dugongs found in seagrass and non-seagrass coral reef habitats. Opportunistic aerial and underwater footage of dugongs, documenting animals’ behaviour in non-seagrass habitats, completed the survey.
Methods
Study area
The lagoons around the main island of New Caledonia encompass some of the world’s most diverse reef structures, especially on the west coast of the main island where a mosaic of habitats from shallow inshore seagrasses to deep channels and a wide range of reef forms occur (Andréfouët et al. 2004).
Cap Goulvain (Fig. 1) has been identified as one of the most important dugong and seagrass habitats in New Caledonia (Payri et al. 2005; Cleguer et al. 2015). The region is also known as a fishing and illegal dugong take area. Dugongs and seagrasses in Cap Goulvain are facing increased human activities and coastal urbanisation, including the completion in 2013 of a large beach resort that attracts a large number of tourists both on land and on the water. Thus, Cap Goulvain is a high priority region for the conservation and management of dugongs and seagrasses to local resource managers.
Aerial survey design, and associated habitat and location of water temperature sampling in Cap Goulvain. Side figures show (a) the location of New Caledonia in the Pacific, and (b) the location of Cap Goulvain on the central-west coast of the main Island of New Caledonia. The lagoon in Cap Gouvlain (c) is 4 km wide on average.
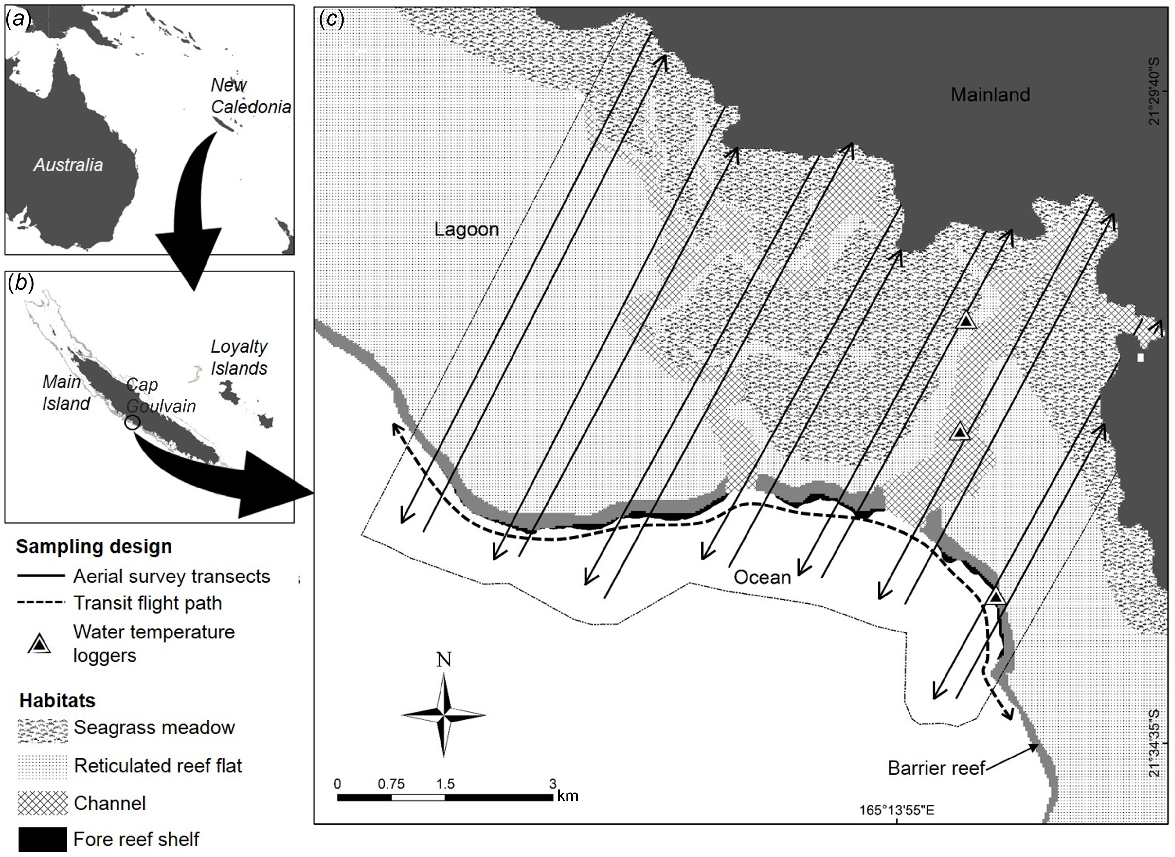
The lagoon in Cap Goulvain is approximately 4 km wide, much narrower than the lagoons on the northern and southern part of New Caledonia’s west coast, which reach 15–30 km width respectively (Bonvallot et al. 2012) (Fig. 1).
Habitat identification was based on the Atlas of the New Caledonian Coral Reefs (Andréfouët et al. 2004) and a generic map of shallow (<5 m) seagrass produced from Landsat images, the best available dataset on seagrass at the scale of the study (Andréfouët et al. 2010). We identified four main habitats, three of which are located inside the lagoon: (1) intertidal seagrass meadows, (2) reticulated reef flats, and (3) deep water channels; and one located outside the lagoon: (4) the fore reef shelf on the seaward edge of the barrier reef, between the fore reef crest and the reef slope leading to the deep open ocean waters (Fig. 1, Table 1; Andréfouët et al. 2004; Payri et al. 2005). Seagrasses are absent from the fore reef shelf and the reticulated reef flats, and are likely to be extremely scarce or nonexistent in the channels where the water is deep (i.e. up to 86 m) and turbid (Hily et al. 2010).
| Habitat type | Size (km2) | Depth (m) A | Presence/absence of seagrass B | |||
|---|---|---|---|---|---|---|
| Mean ± s.d. | Median | Max | ||||
| Seagrass meadows | 17.4 | 0.98 ± 1.99 | 0.3 | 4.7 | Six seagrass species represented, dominated by Cymodocea serrulata | |
| Reticulated reef flats | 20.5 | 1.8 ± 2.57 | 1.5 | 20.3 | Absent | |
| Channels | 6.8 | 27.4 ± 15.8 | 29.9 | 86.2 | Likely scarce | |
| Fore reef shelf | 14.2 | 29.17 ± 14.75 | 29.2 | 97.3 | Absent | |
Habitat area (km2) was calculated by importing the habitat layers into ArcGIS® 10.2 (Esri 2013) and summing the area of each habitat.
Environmental variables
In New Caledonia the cool season extends from April to September and the warm season extends from October to March (Météo France 2014). To examine the seasonal changes in water temperature in the different habitat types in Cap Goulvain, a water temperature logger (Sea-Bird SBE56 V0.96, accuracy ± 0.002°C) was deployed at each of the following sites: (1) the southern edge of the main seagrass meadow at a depth of 2 m; (2) the southern channel at 17 m and (3) over the fore reef shelf in the southern part of the study area at 20 m. The temperature loggers were deployed in mid-August 2012 and recorded temperature every 10 min for a year.
Tide information was retrieved from the local weather forecast website (Météo France 2014). Tidal dynamics of the lagoons of New Caledonia are generally dominated by semi-diurnal tides (e.g. two high tides each day). The tidal range does not vary substantially diurnally or seasonally (Bonvallot et al. 2012).
Dugong sightings
Sixty-two dedicated aerial transect surveys were conducted over Cap Goulvain, during 18 months from mid-February 2012, to record the number of dugongs and seasonal changes in their habitat use. Twenty-nine surveys were conducted during the warm season as defined by Météo France (2014), including 15 surveys at high tide and 14 surveys at low tide (Supplementary Table S1). Thirty-three surveys were conducted during the cool season (Météo France 2014); 16 at high tide and 17 at low tide.
The surveys were conducted using a two-seater Fly Synthesis Storch ultra-light aircraft. A trained observer sat on the starboard side next to the pilot. To limit potential observer bias, the same observer conducted most of the surveys (i.e. 60 of the 62 surveys).
Surveys were conducted at a constant ground speed of 110 km/h. In order to meet New Caledonia safety requirements of a single-engine aircraft over water, flights were conducted at 274 m. Flying at this altitude also enabled the observer to identify the limits of and count large groups of dugongs (Marsh and Sinclair 1989a, 1989b). These ultra-light surveys were flown at approximately two-week intervals (mean interval between surveys = 16.2 days, ±s.e. = 1.4) and took an average of 46.6 min each (±s.e. = 0.68) excluding aircraft transit times to and from the airport. If possible, surveys were conducted at slack low and high tides on the same survey day and in calm weather with wind speeds of less than 20 km/h; and cloud cover less than 6 oktas to minimise sighting bias caused by poor visibility in bad weather.
The survey design and procedures were a variant of the protocols of Marsh and Saalfeld (1989) and Marsh and Sinclair (1989a); coloured tape was attached to the starboard wing strut of the aircraft to delineate a 400 m wide strip on the water surface on the starboard side of the aircraft. This transect width delimited the zone in which dugongs were counted. Habitats inside the lagoon and the fore reef shelf (total of 60 km2) were covered by 14 non-overlapping transects (Fig. 1). Transects were paired because observer coverage was limited to the starboard side of the aircraft. Within pairs, transects were spaced at intervals of 350 m; the inter-pair transect distance was 1 km. Transect distance varied between 4.4 and 7.6 km and cumulatively totalled 86.7 km of survey distance (mean = 6.2 km, s.d. = 0.95). This design allowed the survey to intensively cover the study area with a 59% sampling intensity while limiting the risks of double counting dugongs. All dugong and large shark (>2 m) sightings and their associated habitats were recorded onto an audio-recorder. In all habitats other than the channels the bottom was visible from the aircraft. Dugong counts were recorded as minimum uncorrected counts because perception bias could not be estimated (as only one observer was present in the aircraft), and availability corrections would have only applied to the deep channels (since the sea bottom was visible in all other habitats). We did not have enough information about dugong diving behaviour and the time spent in the detection zone to apply a correction at the time of the study.
Dugong aggregations were observed over the fore reef shelf outside the lagoon during the ultra-light transect surveys. To add to our ultra-light surveys and investigate the spatial and temporal patterns of dugong aggregations in the region we examined the data collected on observations of dugong aggregations made during 41 separate aerial passes made over the fore reef shelf as part of independent dugong surveys conducted north of our study area. For each pass of this dugong survey crew over the fore reef shelf, the flight altitude was purposely adjusted to 274 m to count the dugongs present outside the lagoon. Passes heading north saw the starboard observers counting dugongs over the fore reef shelf, whereas the port observers undertook the count when the aircraft was heading south. There were two observers seated on each side of the plane, which helped verifying counts between observers.
Some aerial video footage of dugong herds (distinct groups of >10 animals separated by approximately no more than an adult dugong body length) over the fore reef shelf was available for analysis. The videos were taken using a Cineflex camera mounted onto a helicopter of the Province Sud Resource Department (Martial Dosdane pers. comm.). Seven videos were taken on 7 July 2011 and four on 16 July 2012, in optimal weather conditions (no overcast and low wind, Table S2). An underwater video, taken by local sea rangers snorkelling over the fore reef shelf on 18 April 2011, was also available for examination (Romain Laigle pers. comm.; Table S2).
Data analysis
We explored how the numbers of dugongs observed from the transect surveys were influenced seasonally (monthly) and by tidal variation. The response was the total number of dugongs observed in each month-tide combination divided by the corresponding number of surveys.
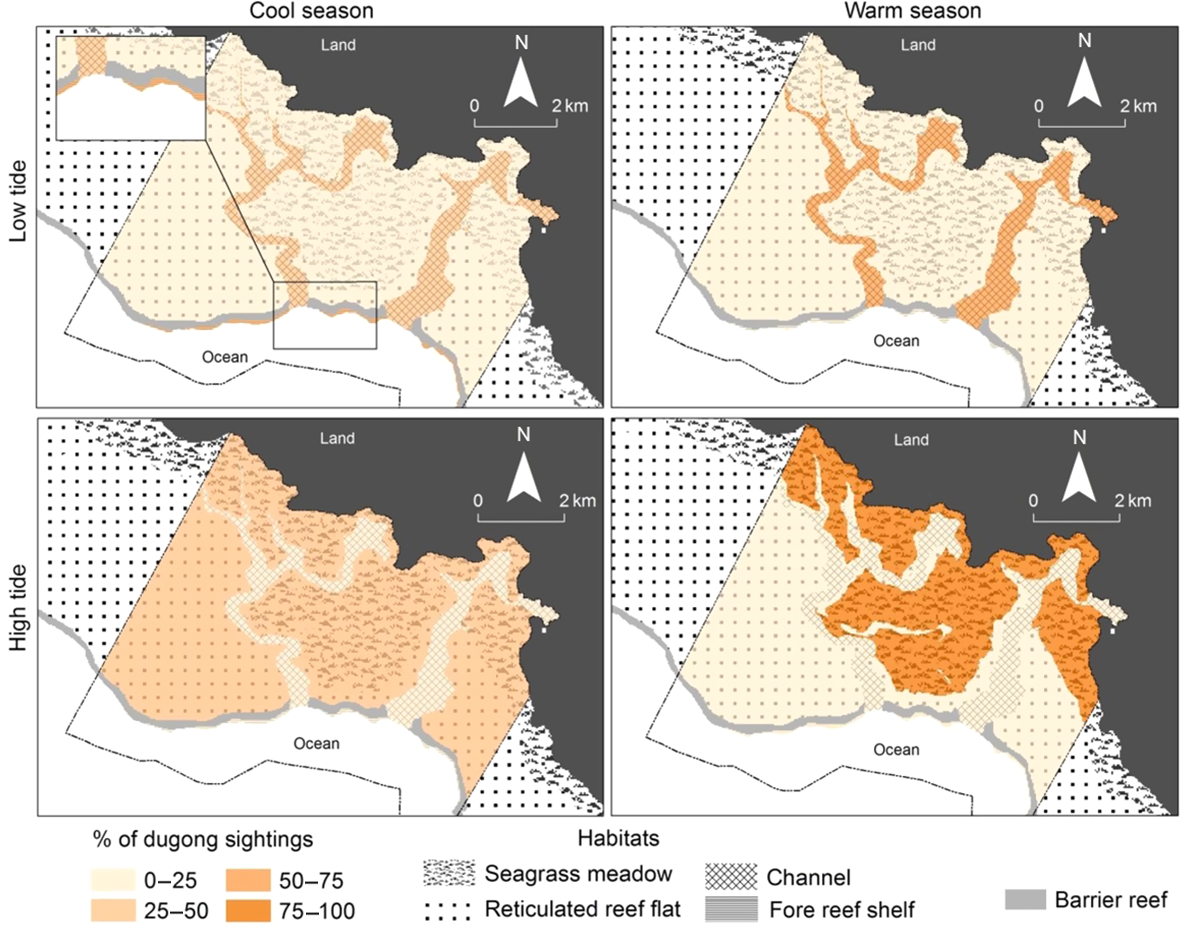
The association between dugong sightings and habitat was made by extracting each sighting shapefile according to the overlaying habitat shapefiles in GIS (ARCGIS® 10.2, Esri 2013). The interaction between the total counts of dugongs for each of the four habitat types (seagrass meadows, channels, reticulated reef flats, fore reef shelf), season and tide was assessed using log-linear regression via generalised linear models (GLM) with a Poisson distribution (Myers and Montgomery 1997) using R (3.1.3, https://cran-archive.r-project.org/bin/windows/base/old/3.1.3/, R Core Team 2020). The size of each habitat was accounted for using the logarithm to the base of the habitat area as an offset in the models. The null hypothesis tested was that there was no three-way interaction between habitat, tide and season. The dataset was subsequently divided into season-tide categories (i.e. cool season-low tide, cool season-high tide, warm season-low tide, warm season-high tide) and the proportion of dugongs sighted across habitats in each season-tide category was calculated in GIS. The resulting values were binned into four classes of dugong sightings (i.e. 0–25, 25–50, 50–75, 75–100% of dugong counts present in a habitat). These classes were coloured and assigned to their relevant habitat layer within each season-tide category in ArcGIS 10.2 (Esri 2013) to enable better visual representation of the results.
We explored the occurrence of dugong aggregations: (1) spatially – inside or outside the lagoon; and (2) temporally – by month. Dedicated transect surveys and transiting flight data were combined for the analysis of the dugong herd observations outside the lagoon, but not for the analysis of the dugong herd observation inside the lagoon because the transiting flights were conducted only outside the lagoon. We then examined the correlation between water temperature changes and the spatial and temporal patterns of dugong herding. We calculated the mean minimum monthly water temperature difference between the fore reef shelf – outside the lagoon, and the seagrass meadow – inside the lagoon during the transect surveys. A Spearman rank correlation test (Zar 1998) was conducted to assess the correlation between the probability of seeing at least one dugong herd and the mean minimum water temperature difference between outside and inside the lagoon. We chose to focus on the minimum water temperatures, which are more likely to trigger a thermoregulatory behaviour response from dugongs. The association between the proportions of dugong herd observations by tides was tested using a Fisher’s exact test (Upton 1992). We examined whether sharks were observed during the same dedicated transect survey as the dugong herd observations. The behavioural protocols developed by Hodgson (2004) were used for the analysis of the video footage in order to identify the behaviour of dugongs in the herds located on the fore reef shelf (Table S3).
Results
Environmental variables: water temperature
During the cool season the mean minimum monthly water temperature was significantly higher over the fore reef shelf outside of the lagoon, than in the channel (i.e. up to 1.9°C difference in May) or over the seagrass meadows (i.e. up to 1.1°C difference in May) (Tukey multiple comparisons tests, P < 0.0001, Table 2). These seasonal changes in water temperature in Cap Goulvain are concordant with the definition of seasons used by Météo France (2014). The difference between the mean minimum monthly water temperature over the fore reef shelf and in the southern channel became positive in April and peaked in May at 1.9°C (Table 2). A similar pattern was observed between the fore reef shelf and the seagrass meadow with the mean minimum monthly water temperature being warmer over the fore reef shelf from May through October (water temperature difference ranging 0.1–1.1°C). During the warm season, the spatial differences in water temperature reversed: the mean minimum monthly water temperature was significantly higher in the channel (up to 1.1°C difference; Tukey multiple comparisons tests, P < 0.0001) and over the seagrass meadow (up to 0.8°C Tukey multiple comparisons tests, P < 0.0001) compared with the fore reef shelf (Table 2).
| Survey month | Fore reef shelf | Southern channel | Seagrass meadow | Mean minimum monthly water temperature difference | ||
|---|---|---|---|---|---|---|
| Mean min | Mean min | Mean min | Fore reef shelf – Seagrass meadow | Fore reef shelf – Southern channel | ||
| January | 24.5 ± 0.9 | 25.6 ± 1.2 | 25.3 ± 0.9 | −0.8 | −1.1 | |
| February | 26.0 ± 0.4 | 26.5 ± 0.9 | 26.2 ± 0.4 | −0.2 | −0.5 | |
| March | 25.6 ± 0.7 | 26.0 ± 1.0 | 25.8 ± 0.7 | −0.2 | −0.4 | |
| April | 25.0 ± 0.8 | 25.0 ± 1.0 | 25.2 ± 0.7 | −0.2 | 0.0 | |
| May | 24.8 ± 0.6 | 22.9 ± 1.0 | 23.7 ± 0.8 | 1.1 | 1.9 | |
| June | 23.6 ± 0.2 | 22.1 ± 0.6 | 22.7 ± 0.4 | 0.9 | 1.5 | |
| July | 22.9 ± 0.3 | 21.2 ± 1.4 | 21.9 ± 1.2 | 1.0 | 1.7 | |
| August | 22.6 ± 0.2 | 21.1 ± 1.0 | 22.0 ± 0.5 | 0.6 | 1.5 | |
| September | 22.4 ± 0.1 | 21.3 ± 1.0 | 22.0 ± 0.5 | 0.4 | 1.0 | |
| October | 22.8 ± 0.4 | 22.7 ± 1.2 | 22.7 ± 0.7 | 0.1 | 0.1 | |
| November | 23.7 ± 0.5 | 24.2 ± 1.2 | 23.8 ± 0.6 | −0.1 | −0.5 | |
| December | 24.6 ± 0.6 | 25.5 ± 0.9 | 24.6 ± 0.7 | 0.0 | −0.9 | |
Seasonal and tidal patterns in the number of dugongs observed per survey
Dugong were observed in Cap Goulvain throughout the entire year. Nonetheless, the number of dugongs observed per survey was three times higher during the cool season, between April and September ( = 18.4 dugongs per survey) than during the warm season, between October and March ( = 5.6 dugong per survey) (Fig. 2). The lowest numbers of dugongs observed per survey was in February ( = 5 dugongs per survey) and December ( = 7 dugongs per survey). Dugongs were observed in large numbers during the cool months of June ( = 50 dugongs per survey), July ( = 41 dugongs per survey) and August ( = 40 dugongs per survey).
Number of dugongs observed per aerial survey in each survey month at high and low tide in Cap Goulvain in 2012–2013.
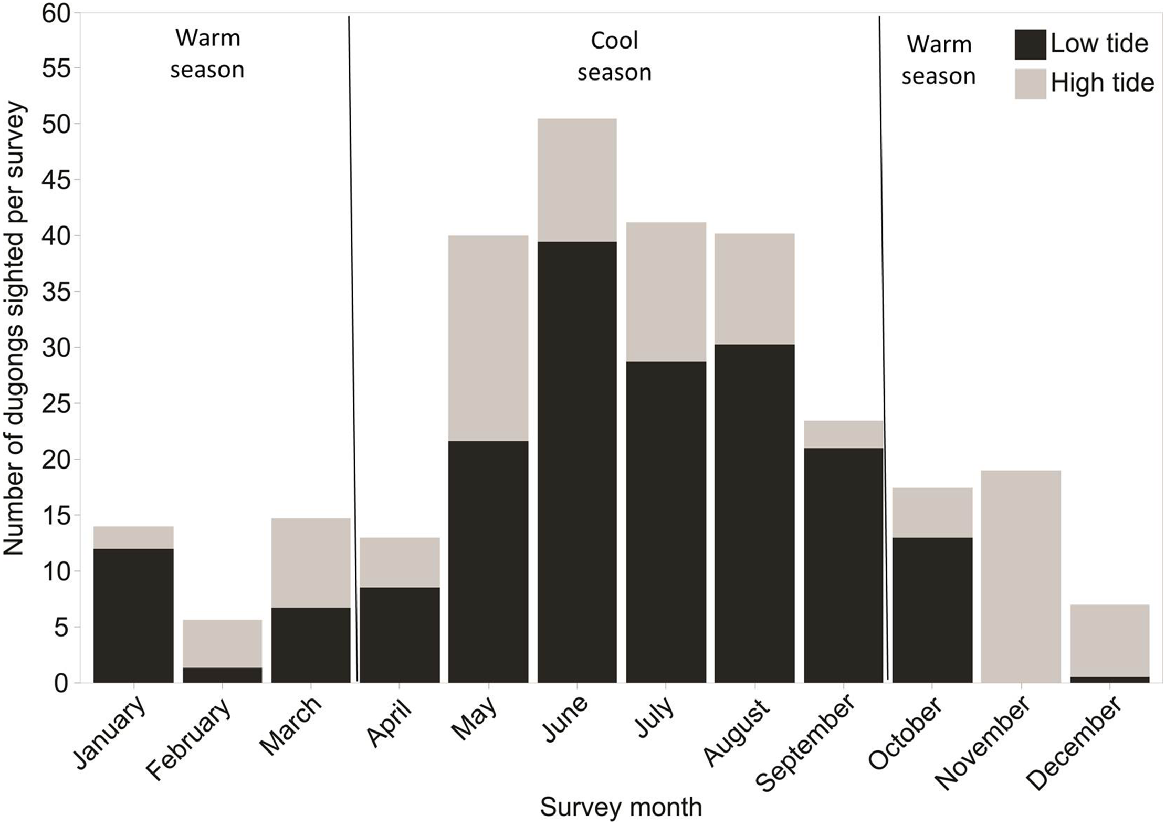
More dugongs were observed per survey at low tide than at high tide in all months of the cool season (Fig. 2). During the warm season there was no apparent influence of tidal pattern in the number of dugongs observed per survey: dugongs were mostly seen at high tide in November, December, February and March, whereas in January and April most of the dugongs were observed at low tide.
The distribution of dugongs across the four habitats varied with season and tides. The log-linear model revealed a significant three-way interaction in the counts of dugongs between habitat, season and tide (G2 = 68.5, d.f. = 4, P < 0.0001; Table S4) and significant two-way tide-habitat (G2 = 372.68, d.f. = 4, P < 0.0001), tide-season (G2 = 35.17, d.f. = 1, P < 0.0001) and season-habitat interactions (G2 = 107.70, d.f. = 4, P < 0.0001). To compare habitat use by dugongs in each season-tide category, we split the dataset into season-tide categories to interpret the interaction.
At high tide, the highest percentage of dugongs was sighted over the seagrass meadow during both the cool and the warm seasons (Fig. 3). However, this percentage changed seasonally and was more than twice as high in the warm season than in the cold season. The seagrass meadow that was used by dugongs was mostly exposed and not available to dugongs at low tide. Use of the fore reef shelf was virtually limited to the cool season and more than twice as frequent at low tide than at high tide. Dugongs were mostly sighted over the reticulated reef flat at low tide during the warm season and at high tide during the cool season and used the channels mostly at low tide, especially in the warm season.
Dugong herds
Dugong herds (Fig. 4) were observed in Cap Goulvain on 27 of 103 flights, including both transect surveys and transiting flights. Six observations of dugong herds were made inside the lagoon and 21 outside the lagoon. There was a noticeable seasonal difference in the location of the dugong herds: herds were sighted inside the lagoon from the end of the cool season to the end of the warm season between August and March, whereas herds were sighted outside the lagoon during the cool season between April and September (transect and transiting flights; Fig. 5).
A herd of 69 dugongs over the fore reef shelf outside of the lagoon in Cap Goulvain on 22 June 2011. A high proportion of animals were apparently basking at the surface.

Proportion of dugong herd sightings per month outside the lagoon (grey columns) and inside the lagoon (white columns) in Cap Goulvain in relation to the mean minimum monthly water temperature difference between the fore reef shelf, outside the lagoon and the southern channel, inside the lagoon. The dotted black line is a reference point to enable the reader to see when the water temperature becomes warmer (positive temperatures) or cooler (negative temperatures) on the fore reef shelf compared to the southern channel.
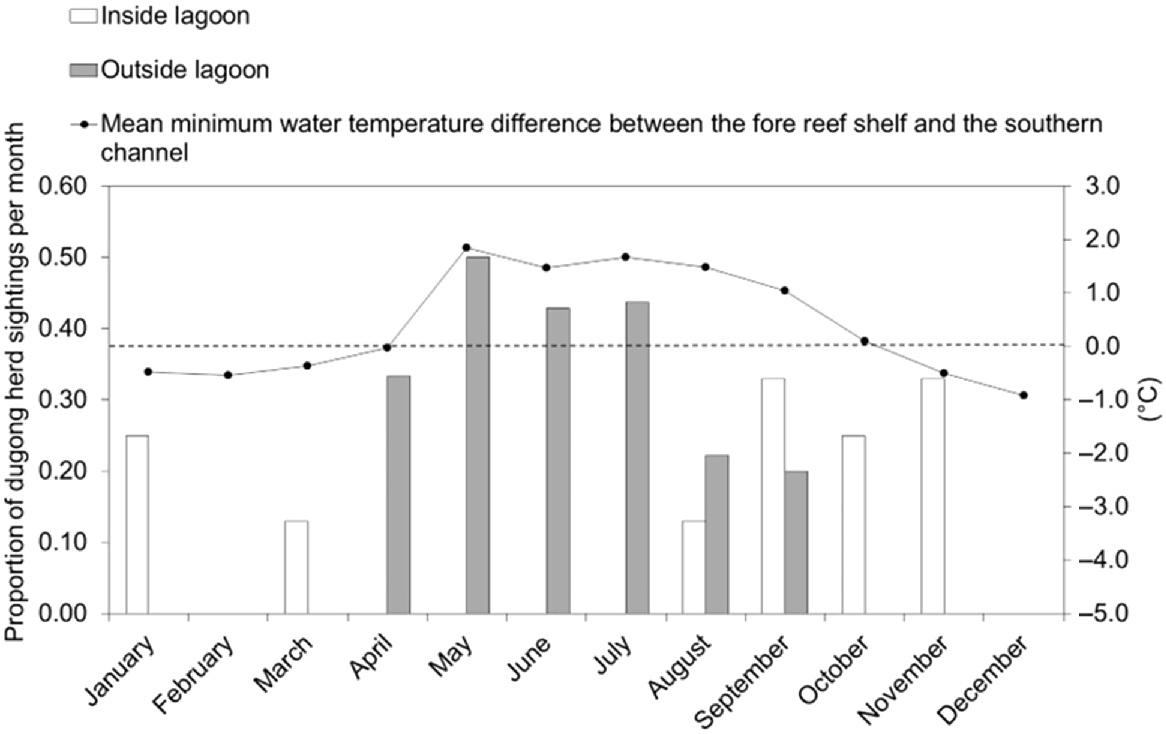
The probability of seeing at least one dugong herd outside the lagoon ranged between 0.12 at high tide and 0.38 at low tide and inside the lagoon between 0.03 at high tide and 0.17 at low tide (Table S5). These variations did not differ significantly across tides, neither outside the lagoon (Fisher’s exact test, P = 0.29) nor inside the lagoon (Fisher’s exact test, P = 0.29). On four occasions, dugong herds were sighted at the same location on the same day but at different tidal regimes (Table S6). For example, a herd of 57 dugongs was sighted at the peak of the low tide on the fore reef shelf during a transect survey on 14 May 2012. A smaller herd of 20 dugongs was sighted at the same location during the transect survey on the following high tide peak 6 h later.
There was a strong positive correlation between the proportion of dugong herd sightings in a month on the fore reef shelf and the difference in the mean minimum monthly water temperatures on the fore reef shelf and in the southern channel (Spearman rank correlation coefficient, n = 12, r = 0.858, P < 0.01).
Sharks were never observed in the vicinity of a dugong herd (Table S6). However, sharks were occasionally observed on the same flight as a dugong herds but in a different habitat. For example, a shark was observed over the seagrass meadow at low tide on 27 June 2013 when two dugong herds were approximately 2.5 km away (straight line distance) on the fore reef shelf.
Behaviour of individuals within herds
In the videos of the dugong herds located over the fore reef shelf, we examined if any of the behaviours listed by Hodgson (2004) were displayed by the individuals forming the herds. We identified the behavioural states of resting, rolling, surfacing, socialising and feeding. The videos were too short (i.e. the longest video lasted for 24 min) and moving too much across the herds to conduct comprehensive focal follow examinations (Supplementary video1). Travelling was not observed. Feeding behaviour was limited to calves suckling from their mother. Dugongs rested at the surface and in the mid-water column. Calves rested on their mother’s back or side. The social events (sensuHodgson 2004) displayed were ‘close pass by’, ‘join’ and ‘swim away’. Dugongs were also observed swimming at very slow speed with changes in direction or immobile basking at the water surface. The mother and calf pairs were swimming together with the calf on the side or over the back of their mother. There was no evidence of pre-copulatory behaviours or mating.
Discussion
This study comprehensively analyses the temporal changes in habitat use by dugongs within a geographically constrained coral reef setting. Our findings show that the inherent behavioural propensities and/or evolutionary adaptations of dugongs enable them to effectively utilise small coral reef ecosystems, using unexpected habitats, notably the fore reef shelf situated outside the lagoon. Strong seasonal variations in the number of dugongs sighted in Cap Goulvain as well as seasonal and tidal changes in their habitat use were found. We focus our discussion on the potential drivers of these changes.
Seasonal and tidal changes in dugong numbers and their use of habitats in Cap Goulvain
Expectedly, dugongs were sighted in higher numbers over the intertidal seagrass meadows at high tide than in any other habitat regardless of the season, suggesting that, conditional on availability, dugongs select seagrass foraging habitats over non-seagrass habitats. When tides restricted access to the seagrass meadows there was a seasonal variability in the way dugongs were distributed over the non-seagrass coral reef habitats, suggesting that some external factors may have influenced the trade-offs between the habitats available to the animals. For example, during the cool season at low tide a higher percentage of dugongs was sighted on the fore reef shelf, outside the lagoon, than in any other non-seagrass habitat inside the lagoon. Contrastingly, during the warm season at low tide dugongs appeared to remain in the lagoon (as is most commonly reported in dugong aerial surveys conducted at the scale of the main island of New Caledonia; Cleguer et al. 2017), particularly in the channels close to the seagrass habitats. During the cool season the water temperature on the fore reef shelf is overall higher than the water temperature inside the lagoon. This difference reversed during the warm season. A similar difference in water temperature between habitats located inside the lagoon and oceanic waters has been documented in other regions on the west coast of New Caledonia where the lagoon is wider than in Cap Goulvain including the south-west lagoon in the Nouméa region (i.e. up to 35 km from coastline to barrier reef).
Collectively, these results suggest that behavioural thermoregulation may have played a key role in the seasonal changes in number and behaviour of dugongs in Cap Goulvain as well as in the way animals used their habitats in the region. Below we evaluate the evidence supporting this hypothesis while considering predation risk as another possible, not mutually exclusive explanatory factor.
Behavioural thermoregulation
Thermoregulation is a particular problem for marine mammals because they spend all or a greater part of their lives in water, a medium with conductivity 25 times greater than air and with temperatures generally lower than their body temperature (Pabst et al. 1999; Rosen et al. 2007; Satinoff 2011). Each species of marine mammals have evolved their own biological (i.e. including morphological and physiological) and behavioural adaptations to regulate their body temperature and endure changes in water temperature (White and Odell 1971; Odell 1974; Pabst et al. 1999; Newell 2013).
However, several lines of evidence suggest biological thermal sensitivity in sirenians (Marshall et al. 2022). Dugongs and manatees have a poor thermoregulatory capacity (Worthy 2001) even though they have a well-developed dermis (Horgan et al. 2014). The manatees’ basal body temperature is considered near the lower end of the normal mammalian range (Costa and Williams 1999). They have a very narrow thermoneutral zone (i.e. range of environmental temperatures where metabolic heat production is minimal for the animal) (Gallivan et al. 1983). This information is not available for dugongs but given their biology it is likely to be similar to that of manatees. Finally, dugongs lack an effective heat conservation mechanism (Bryden et al. 1978; Pabst et al. 1999).
The seasonal changes in the distribution and number of dugongs at low tide in non-seagrass coral reef habitats of Cap Goulvain were correlated with the differences in water temperature between the outer and the inner-lagoon areas. Using Newton’s law of cooling (Davidson et al. 2012), we calculated that the ~2°C temperature difference between the dugongs spending low tide on the fore reef shelf rather than in the subtidal sections of the lagoon in the cool season could reduce the heat transfer to the dugong’s body core to the skin by 24% (the equation of the variables influencing the rate of conductive heat loss from a body is provided in Pabst et al. (1999). These energy savings would be reversed in the warm season when it is warmer in the lagoon than on the fore reef shelf.
Our findings contribute to the existing body of evidence indicating that dugongs employ behavioural thermoregulation as a response to fluctuations in water temperature within their habitat. Behavioural thermoregulation, including movements into warmer waters, has been extensively studied on manatees in Florida (USA, See Laist and Reynolds (2005a) for review). For example, manatees in Florida respond to cold water temperatures by seasonal migrations and local movements between warm-water springs and industrial warm-water sources during winter (Laist and Reynolds 2005b; Deutsch et al. 2006). Several evidences, including our study, show that dugongs undertake, to some extent, a similar behaviour. For example, Preen (1992) reported movements in and out of Moreton Bay (eastern Australia) over a temperature gradient of less than 2°C between offshore and inshore areas (i.e. 17.7°C inside the bay vs 19.4°C recorded outside the bay). Zeh et al. (2018) further investigated the movements of dugongs satellite-tracked in Moreton Bay and found that dugongs were undertaking behavioural thermoregulation by regularly swimming a relatively short distance to thermoregulate in warmer offshore waters during the cooler winter season. A dugong radio-tagged in Cleveland Bay, eastern Australia, undertook a large-scale movement nearing 100 km that coincided with a drop in the sea surface temperature, although the temperature (>27°C) was still well within the known range of the thermal tolerance of dugongs (Marsh and Rathbun 1990). Six other dugongs tracked in Hervey Bay, eastern Australia, undertook up to 80 km movements across the bay in winter from the 19–20°C coastal waters to the 21–22°C waters off Sandy Cape, despite the apparent lack of seagrass meadows in the Sandy Cape area (Sheppard et al. 2006). Finally, sighting records and aerial surveys also indicate that most dugongs avoid waters with temperatures lower than approximately 19°C (Anderson 1986, 1994; Marsh and Sinclair 1989b; Preen 1989, 1992; Marsh et al. 1994; Preen et al. 1997; Gales et al. 2004; Holley et al. 2006).
Dugongs were apparently basking at the surface on the fore reef shelf. Basking is a frequently observed behaviour in aquatic animals. Two forms of basking exist: aquatic basking, where animals float in warm surface water (observed in our study); and atmospheric basking, where animals haul out onto land and are exposed to the sun. Both are thermoregulatory behaviour used by animals to rise their body temperature. Most basking behaviour is documented for seals (e.g. Butler and Udvardy 1966), freshwater turtles (e.g. Crawford et al. 1983) and sea turtles (e.g. Whittow and Balazs 1982). The effect of basking on the dugong body temperature has never been studied and should be further investigated.
The number of dugongs observed per survey was three times higher during the cool season than during the warm season. The increase in the number of dugongs in Cap Goulvain during the cool season could be a strategy to reduce the energy cost of accessing warm waters. The lagoon in Cap Goulvain is narrower than other regions on the west coast where seagrass meadows are extensive. For example, Nepoui and Poya Bays, the closest bays north of Cap Goulvain that support extensive seagrass meadows, are up to 10 km wide. Similar distances separate inshore seagrasses from oceanic waters in Moindou located south of Cap Goulvain. Thus, the energy cost of dugongs commuting from their seagrass feeding grounds to warm water at Cap Goulvain is potentially lower than in other regions where the lagoon is wider.
Reducing the risk of predation
As long-lived slow breeding marine mammals, dugongs need to maximise survival by minimising the risk of predation. The evidence of direct attacks on dugongs from sharks summarised by Marsh et al. (2012) indicates that there is a risk of dugongs being attacked by large sharks, particularly tiger sharks (Galeocerdo cuvier). Wirsing et al. (2007a, 2007b) suggested that dugongs face trade-offs in habitat use to reduce the risk of encountering a predator and to increase the likelihood of escaping from it in the case of an attack. In Wirsing et al.’s (2007a, 2007b) studies, dugongs over-used edges of seagrass meadows when sharks were abundant in the study area. This strategy presumably enables dugongs to increase their chances to move to deep waters adjacent to the seagrass meadows to escape from a shark attack.
These strategies are likely to fail if dugongs are ambushed in the reticulated reef flats and channels in Cap Goulvain. The reticulated reef flats are subtidal but shallow (<5 m) formations with the potential to limit movements of large vertebrates (Andréfouët et al. 2004). There are reports of shark attacks of live dugongs in the reticulated reef flats of the west coast of New Caledonia: one during the first large-scale dugong population survey conducted in New Caledonia (Garrigue et al. 2008) and by sea rangers during boat patrols (Bruno Manach. pers. comm.). Adjacent to the reticulated reef flats, the channels provide access to deep water to escape from predation. However, the channels in Cap Goulvain are deep and turbid, possibly limiting the dugongs’ capacity to detect shark approaching them from below. The fore reef shelf where we observed herds of dugongs may be a more suitable habitat than deep channels to avoid shark predation because (1) it is a shallow platform where the water is clear so predation from below may be more easily avoided than in the deep channels; and (2) the fore reef shelf provides access to deep oceanic waters to escape shark attacks as suggested by Wirsing et al. (2007a, 2007b).
Why are dugongs forming herds over the fore reef shelf in Cap Goulvain?
The function of dugong herds is poorly understood because it has been difficult to study wild dugongs’s behaviour because of their elusive behaviour, the habitats they utilise, and the lack of effective tools to easily conduct focal follows of animals (note that this study was conducted before the recent emergence of drones, which now provide a cost-effective way to study wild aquatic animals in remote areas).
Hodgson (2004) conducted the most comprehensive study on the behaviour of dugongs forming herds using a blimp-mounted video system. She concluded that dugongs were not forming herds to facilitate social interactions, rest or reduce the risk of predation. Rather their aggregations stimulated the growth of pioneer seagrass species with high quality foliage (Preen 1992, 1995; Hodgson 2004). However, these interpretations of the function of herding in dugongs in Moreton Bay may have been biased by the focus of the observations, which were mostly conducted over seagrass meadows.
Our study shows that there may be other functions in herding than just a food resource maintenance strategy because there is no doubt from the videos that the dugongs sighted within the herds on the fore reef shelf in Cap Goulvain were resting and this habitat is devoid of seagrass. Our video footage were opportunistically taken and limited in time. After reviewing the available footage, we did not detect any discernible social interactions among dugongs, with the exception of females nursing their calves. Although our results concur with Hodgson’s (2004) suggestion that dugong herds may not occur to facilitate social interactions, further studies need to be conducted particularly in examining the use of vocalisation during herding in a range of habitats (O’Shea et al. 2022). Besides, the dugong aggregations observed resting and presumably basking over the fore reef shelf were mostly observed during the cool season, coinciding with the increase in dugong abundance in Cap Goulvain. We conclude that dugong herd formation over the fore reef shelf may have simply reflected an inevitable increase in density associated with the seasonal increase in abundance in Cap Goulvain. This seasonal increase in dugong numbers combined with the need to use a habitat as warm as possible (i.e. the behavioural thermoregulation theory explained above) with easy access to food resources is a plausible explanation for why dugongs aggregate on the fore reef shelf at Cap Goulvain during the cool season. However, we cannot exclude the possibility that the formation of an aggregation may also decrease the risk of predation on individuals by increasing the probability of the herd detecting and defending itself from predators as has been documented in other species of marine mammals (e.g. De Vos et al. 2015), insects (e.g. Roberge et al. 2016), and fishes (e.g. Hamilton 1971; Pitcher 1986; Ioannou et al. 2008).
Conclusion
Our study contributes to the growing body of evidence regarding the behavioural thermoregulation of dugongs at the uppermost latitudinal limit of their range. It also highlights the importance of both seagrass and non-seagrass habitats for dugongs. Even though it is a non-seagrass habitat, the fore reef shelf is a key habitat for dugongs where they can stay warm while resting, and possibly avoid predation during the cool season (on that note, further investigation is needed to comprehend the potential effect of predation on the herding behaviour we have reported in this study). The ecological importance of the fore reef shelf has also been identified for species of reef fish in the tropical Atlantic (Olavo et al. 2011) and reef sharks in the Great Barrier Reef in Australia (Rizzari et al. 2014) and therefore should be considered in management plans.
This study confirms the consistent presence of dugongs in Cap Goulvain in substantial numbers throughout the year. Furthermore, the use of diverse habitats within this local area exhibits a significant level of complexity, influenced by a multifaceted interplay of geomorphological and environmental factors such as tides and water temperature, potentially compounded by the impact of natural predation. As such, we strongly advocate for an intensified commitment from local authorities towards the management of this area. This should commence with the re-initiation of consultations and active engagement with the local communities residing in the vicinity, along with collaboration with tourism companies.
Data availability
Data is available and accessible upon request made to the lead author. The video footage is available from the Research Data JCU repository at https://doi.org/10.25903/zhfx-4847.
Declaration of funding
This research was supported by the New Caledonian Dugong Technical Committee under the 2010−2015 Dugong Action Plan in New Caledonia. At the time of this study, the Dugong Action Plan was coordinated by the Marine Protected Area Agency and included local environmental managers from the Province Nord, the Province Sud and the Province des Iles Loyauté of New Caledonia, as well as representatives from the French and New Caledonian Governments, WWF-New Caledonia and the local environmental NGO Operation Cétacés.
Acknowledgements
We thank staff from Cap ULM whom conducted the ultra-light surveys and Martial Dosdanes and Romain Laigle for the footage they provided. This paper forms part of the PhD thesis of Christophe Cleguer (first author) completed in 2015.
References
Anderson PK (1986) Dugongs of Shark Bay, Australia-Seasonal migration, water temperature, and forage. National Geographic Research 2, 473-490.
| Google Scholar |
Anderson PK (1994) Dugong distribution, the seagrass Halophila spinulosa, and thermal environment in winter indeeper waters of Eastern Shark Bay, WA. Wildlife Research 21, 381-388.
| Crossref | Google Scholar |
Andréfouët S, Payri C, Kulbicki M, Scopélitis J, Dalleau M, Mellin C, Scamps M, Dirberg G (2010) ‘Mesure, suivi et potentiel économique de la diversité de l’habitat récifo-lagonaire néo-calédonien: inventaire des herbiers, suivi des zones coralliennes et rôle des habitats dans la distribution des ressources en poissons de récifs. Biologie Marine.’ Institut de Recherche pour le Développement (IRD) Centre de Nouméa/ZoNéCo.
Bryden MM, Marsh H, MacDonald BW (1978) Skin and hair of the dugong, Dugong Dugon. Journal of Anatomy 126, 637-638.
| Google Scholar |
Butler GD, Jr., Udvardy MDF (1966) Basking behavior of the Hawaiian monk seal on Laysan Island. The Journal of Wildlife Management 30, 627-628.
| Crossref | Google Scholar |
Cleguer C, Grech A, Garrigue C, Marsh H (2015) Spatial mismatch between marine protected areas and dugongs in New Caledonia. Biological Conservation 184, 154-162.
| Crossref | Google Scholar |
Cleguer C, Garrigue C, Fuentes MMPB, Everingham Y, Hagihara R, Hamann M, Payri C, Marsh H (2017) Drivers of change in the relative abundance of dugongs in New Caledonia. Wildlife Research 44(4), 365-376.
| Crossref | Google Scholar |
Cleguer C, Kelly N, Tyne J, Wieser M, Peel D, Hodgson A (2021) A novel method for using small unoccupied aerial vehicles to survey wildlife species and model their density distribution. Frontiers in Marine Science 8, 640338.
| Crossref | Google Scholar |
Crawford KM, Spotila JR, Standora EA (1983) Operative environmental temperatures and basking behavior of the turtle Pseudemys scripta. Ecology 64, 989-999.
| Crossref | Google Scholar |
Davidson AD, Boyer AG, Kim H, Pompa-Mansilla S, Hamilton MJ, Costa DP, Ceballos G, Brown JH (2012) Drivers and hotspots of extinction risk in marine mammals. Proceedings of the National Academy of Sciences 109, 3395-3400.
| Crossref | Google Scholar |
De Vos A, Justin O’Riain M, Meÿer MA, Kotze PGH, Kock AA (2015) Behavior of Cape fur seals (Arctocephalus pusillus pusillus) in response to spatial variation in white shark (Carcharodon carcharias) predation risk. Marine Mammal Science 31, 1234-1251.
| Crossref | Google Scholar |
Derville S, Cleguer C, Garrigue C (2022) Ecoregional and temporal dynamics of dugong habitat use in a complex coral reef lagoon ecosystem. Scientific Reports 12, 552.
| Crossref | Google Scholar |
Deutsch CJ, Castelblanco-Martinez DN, Groom R, Cleguer C (2022) Movement behavior of manatees and dugongs: I. Environmental challenges drive diversity in migratory patterns and other large-scale movements. In ‘Ethology and behavioral ecology of Sirenia’. (Ed. H Marsh) pp. 155–232. (Springer: Berlin)
Dunning JB, Danielson BJ, Pulliam HR (1992) Ecological processes that affect populations in complex landscapes. Oikos 65, 169-175.
| Crossref | Google Scholar |
Esri (2013) ArcGis Network Analyst. Desktop Developer. Esri, Redlands. California. Available at www.esri.com/software/arcgis/extensions/networkanalyst/index.html
Gales N, McCauley RD, Lanyon J, Holley D (2004) Change in abundance of dugongs in Shark Bay, Ningaloo and Exmouth Gulf, Western Australia: evidence for large-scale migration. Wildlife Research 31, 283-290.
| Crossref | Google Scholar |
Gallivan GJ, Best RC, Kanwisher JW (1983) Temperature regulation in the Amazonian manatee Trichechus inunguis. Physiological Zoology 56, 255-262.
| Crossref | Google Scholar |
Garrigue C, Patenaude N, Marsh H (2008) Distribution and abundance of the dugong in New Caledonia, southwest Pacific. Marine Mammal Science 24, 81-90.
| Crossref | Google Scholar |
Garrigue C, Bonneville CD, Cleguer C, Oremus M (2022) Extremely low mtDNA diversity and high genetic differentiation reveal the precarious genetic status of dugongs in New Caledonia, South Pacific. Journal of Heredity 113(5), 516-524.
| Crossref | Google Scholar | PubMed |
González Carman V, Botto F, Gaitán E, Albareda D, Campagna C, Mianzan H (2014) A jellyfish diet for the herbivorous green turtle Chelonia mydas in the temperate SW Atlantic. Marine Biology 161, 339-349.
| Crossref | Google Scholar |
Hamel S, Côté SD (2008) Trade-offs in activity budget in an alpine ungulate: contrasting lactating and nonlactating females. Animal Behaviour 75, 217-227.
| Crossref | Google Scholar |
Hamel MA, Marsh H, Cleguer C, Garrigue C, Oremus M (2022) Dugong dugon (New Caledonia subpopulation). The IUCN Red List of Threatened Species 2022: e.T218582754A218589361. Available at https://dx.doi.org/10.2305/IUCN.UK.2022-2.RLTS.T218582754A218589361.en
Hamilton WD (1971) Geometry for the selfish herd. Journal of Theoretical Biology 31(2), 295-311.
| Crossref | Google Scholar | PubMed |
Holley DK, Lawler IR, Gales NJ (2006) Summer survey of dugong distribution and abundance in Shark Bay reveals additional key habitat area. Wildlife Research 33, 243-250.
| Crossref | Google Scholar |
Horgan P, Booth D, Nichols C, Lanyon JM (2014) Insulative capacity of the integument of the dugong (Dugong dugon): thermal conductivity, conductance and resistance measured by in vitro heat flux. Marine Biology 161, 1395-1407.
| Crossref | Google Scholar |
Ioannou CC, Tosh CR, Neville L, Krause J (2008) The confusion effect—from neural networks to reduced predation risk. Behavioral Ecology 19, 126-130.
| Crossref | Google Scholar |
Laist DW, Reynolds JE, III (2005a) Florida manatees, warm-water refuges, and an uncertain future. Coastal Management 33, 279-295.
| Crossref | Google Scholar |
Laist DW, Reynolds J (2005b) Influence of power plants and other warm-water refuges on Florida manatees. Marine Mammal Science 21, 739-764.
| Crossref | Google Scholar |
Lanyon JM (2003) Distribution and abundance of dugongs in Moreton Bay, Queensland, Australia. Wildlife Research 30, 397-409.
| Crossref | Google Scholar |
Marsh H, Rathbun GB (1990) Development and application of conventional and satellite radio tracking techniques for studying dugong movements and habitat use. Australian Wildlife Research 17, 83-100.
| Crossref | Google Scholar |
Marsh H, Saalfeld WK (1989) Distribution and Abundance of Dugongs in the Northern Great Barrier-Reef Marine Park. Australian Wildlife Research 16, 429-440.
| Crossref | Google Scholar |
Marsh H, Sinclair DF (1989a) Correcting for visibility bias in strip transect aerial surveys of aquatic fauna. The Journal of Wildlife Management 53, 1017-1024.
| Crossref | Google Scholar |
Marsh H, Sinclair DF (1989b) An experimental evaluation of dugong and sea turtle aerial survey techniques. Australian Wildlife Research 16, 639-650.
| Crossref | Google Scholar |
Marsh H, Prince RIT, Saafeld WK, Shepherd R (1994) The distribution and abundance of the dugong in Shark Bay, Western Australia. Wildlife Research 21, 149-161.
| Crossref | Google Scholar |
Marshall CD, Sarko DK, Reep RL (2022) Morphological and sensory innovations for an aquatic lifestyle. In ‘Ethology and behavioral ecology of Sirenia’. (Eds H Marsh) pp. 19–65 10.1007/978-3-030-90742-6_2
Myers RH, Montgomery DC (1997) A tutorial on generalized linear models. Journal of Quality Technology 29, 274-291.
| Crossref | Google Scholar |
Météo France (2014) Données publiques de Météo France. Available at https://meteofrance.fr [accessed 9 December 2014]
Odell DK (1974) Behavioral thermoregulation in the California Sea Lion. Behavioral Biology 10, 231-237.
| Crossref | Google Scholar | PubMed |
Olavo G, Costa PAS, Martins AS, Ferreira BP (2011) Shelf-edge reefs as priority areas for conservation of reef fish diversity in the tropical Atlantic. Aquatic Conservation: Marine and Freshwater Ecosystems 21, 199-209.
| Crossref | Google Scholar |
O’Shea TJ, Beck CA, Hodgson AJ, Keith-Diagne L, Marmontel M (2022) Social and reproductive behaviors. In ‘Ethology and behavioral ecology of Sirenia’. (Ed. H Marsh) pp. 101–154. (Springer). doi:10.1007/978-3-030-90742-6_4
Preen A (1995) Diet of dugongs: are they omnivores? Journal of Mammalogy 76, 163-171.
| Crossref | Google Scholar |
Preen A (2004) Distribution, abundance and conservation status of dugongs and dolphins in the southern and western Arabian Gulf. Biological Conservation 118, 205-218.
| Crossref | Google Scholar |
Preen AR, Marsh H, Lawler IR, Prince RIT, Shepherd R (1997) Distribution and abundance of dugongs, turtles, dolphins and other megafauna in Shark Bay, Ningaloo Reef and Exmouth Gulf, Western Australia. Wildlife Research 24, 185-208.
| Crossref | Google Scholar |
Rizzari JR, Frisch AJ, Magnenat KA (2014) Diversity, abundance, and distribution of reef sharks on outer-shelf reefs of the Great Barrier Reef, Australia. Marine Biology 161, 2847-2855.
| Crossref | Google Scholar |
Roberge C, Fréchette B, Labrie G, Dumont F, Lucas E (2016) Gregarious pupation act as a defensive mechanism against cannibalism and intraguild predation. Insect Science 23, 612-620.
| Crossref | Google Scholar | PubMed |
Rosen DAS, Winship AJ, Hoopes LA (2007) Thermal and digestive constraints to foraging behaviour in marine mammals. Philosophical Transactions of the Royal Society B: Biological Sciences 362, 2151-2168.
| Crossref | Google Scholar | PubMed |
Sheppard JK, Preen AR, Marsh H, Lawler IR, Whiting SD, Jones RE (2006) Movement heterogeneity of dugongs, Dugong dugon (Müller), over large spatial scales. Journal of Experimental Marine Biology and Ecology 334, 64-83.
| Crossref | Google Scholar |
Sheppard JK, Jones RE, Marsh H, Lawler IR (2009) Effects of tidal and diel cycles on dugong habitat use. The Journal of Wildlife Management 73, 45-59.
| Crossref | Google Scholar |
Short F, Carruthers T, Dennison W, Waycott M (2007) Global seagrass distribution and diversity: a bioregional model. Journal of Experimental Marine Biology and Ecology 350, 3-20.
| Crossref | Google Scholar |
Sih A (1980) Optimal behavior: can foragers balance two conflicting demands. Science 210, 1041-1043.
| Crossref | Google Scholar | PubMed |
Upton GJG (1992) Fisher’s exact test. Journal of the Royal Statistical Society. Series A (Statistics in Society) 155, 395-402.
| Crossref | Google Scholar |
White FN, Odell DK (1971) Thermoregulatory behavior of the northern elephant seal, Mirounga angustirostris. Journal of Mammalogy 52, 758-774.
| Crossref | Google Scholar |
Whittow GC, Balazs GH (1982) Basking behavior of the Hawaiian green turtle (Chelonia mydas). Pacific Science 36, 129-139.
| Google Scholar |
Wirsing AJ, Heithaus MR, Dill LM (2007a) Living on the edge: dugongs prefer to forage in microhabitats that allow escape from rather than avoidance of predators. Animal Behaviour 74, 93-101.
| Crossref | Google Scholar |
Wirsing AJ, Heithaus MR, Dill LM (2007b) Can you dig it? Use of excavation, a risky foraging tactic, by dugongs is sensitive to predation danger. Animal Behaviour 74, 1085-1091.
| Crossref | Google Scholar |
Zar JH (1998) Spearman rank correlation. In ‘Encyclopedia of Biostatistics’, vol. 7. (Eds P Armitage, T Colton). (John Wiley and Sons). doi:10.1002/0470011815.b2a15150
Zeh DR, Heupel MR, Hamann M, Jones R, Limpus CJ, Marsh H (2018) Evidence of behavioural thermoregulation by dugongs at the high latitude limit to their range in eastern Australia. Journal of Experimental Marine Biology and Ecology 508, 27-34.
| Crossref | Google Scholar |