

Effects of environmental variation on the composition and dynamics of an arid-adapted Australian bird community
Judy E. Smith44 Hawkins Parade, Blaxland, NSW 2774, Australia. Email: smitheco@ozemail.com.au
Pacific Conservation Biology 21(1) 74-86 https://doi.org/10.1071/PC14905
Submitted: 27 June 2014 Accepted: 14 July 2014 Published: 21 April 2015
Abstract
Environmental variables influence the dynamics of bird communities. Australian arid-adapted bird communities must cope with particularly high levels of spatial and temporal variability, including inevitable but unpredictable periods of drought. Over four years, which included a severe drought and a period of above-average rainfall, I quantified the responses of a bird community in arid north-western New South Wales to regular seasonal and irregular climatic variation, especially rainfall, as well as spatial variation. I found pronounced changes in the abundance and composition of the avifauna that related to drought and, to a lesser extent, seasonal variability. Overall bird abundance and species richness declined during the drought but the magnitude and direction of population fluctuations of resident, nomadic and migratory species, different feeding groups, and individual species were not consistent. Avian densities and species richness in the study area were higher in mesic habitats associated with drainage lines than in run-off areas. The study demonstrated the importance of local habitat heterogeneity. The asynchronous species responses to a fluctuating environment indicated that drought is likely to have a differential effect on resources and that individual species respond differently to environmental variability. Effective land management and conservation of Australian arid-adapted bird communities requires an understanding of their spatial and temporal variability and dynamics at both local and regional level. A proper understanding of the variability and dynamics of the avifauna is especially important as climate change is predicted to exacerbate the climatic variability and unpredictability of the arid zone in future years.
Additional keywords: arid zone, environmental variability, population dynamics.
Introduction
In arid regions rainfall is low and generally erratic. Birds in these regions must cope with inevitable but unpredictable periods of drought, representing ‘ecological crunches’ (Wiens 1977, 1989), with severe reductions in resource levels. At other times, resources may be superabundant. Birds respond to such environmental variability by employing three broad life-history strategies: residency, nomadism and migration (Ford 1989).
The Australian arid zone is characterised by greater spatial and temporal variability than in areas of comparable aridity elsewhere in the world (Leeper 1970; Morton et al. 2011) and is popularly considered a region of ‘boom and bust’ (Robin et al. 2009). Droughts occur at irregular, unpredictable intervals (Foley 1957; Reynolds et al. 1983) and are of an unpredictable duration and severity. There are numerous accounts of the effects of drought on the avifauna (e.g. Berney 1906, 1928; Barnard 1917, 1927; McGilp 1924; MacGillivray 1929; Robertson 1987). Droughts have been associated with decreases in both species richness and abundance, while post-drought recoveries have been associated with influxes of nomadic species (Reid et al. 1993; Paltridge and Southgate 2001; Burbidge and Fuller 2007). Population declines of birds exposed to drought have been attributed to curtailment of breeding and direct mortality (Keast 1959) as well as the departure of nomads.
Australian arid-zone birds are generally perceived to have fared well since European settlement (Burbidge and McKenzie 1989; Morton 1990). However, several authors have voiced concerns regarding the current and future status of the arid-zone avifauna (Recher and Lim 1990; Reid and Fleming 1992; Smith and Smith 1994) and some 15 species of terrestrial birds of arid mainland Australia are currently considered threatened or near threatened (Garnett et al. 2011). For arid-zone birds, major on-going threatening processes include over-grazing by domestic, feral and native herbivores, predation by feral cats and foxes, and changed fire regimes. At present, some of the impacts of grazing may be masked by the longevity of perennial shrubs and trees in the arid zone (Garnett et al. 2011).
Accurate assessment of changes in the arid-zone avifauna is hampered by the meagre historical record, a lack of detailed information on the present distribution and abundance of birds (Reid and Fleming 1992), and a poor understanding of responses of arid-zone birds to environmental variation (Burbidge and Fuller 2007). Only a few published studies provide estimates of density for Australian arid-zone bird communities: for example, Brooker et al. (1979) in the north-west Nullarbor Plain of Western Australia, Wiens (1991) in chenopod scrubland of north-western New South Wales, Reid et al. (1993) in Uluru National Park in central Australia, Cody (1994) in mulga scrub across Australia, and Burbidge and Fuller (2007) in the Gibson Desert of Western Australia. Even fewer studies include estimates of density for a given site over several years and seasons (Brooker et al. 1979; Reid et al. 1993; Burbidge and Fuller 2007).
Effective conservation of the arid-zone avifauna requires an understanding of the abundance, composition and dynamics of the avifauna. Studies of arid-zone birds need to take account of the variable and unpredictable nature of the environment (Stafford Smith and Morton 1990), particularly since climate change is likely to increase this variability and unpredictability (Hughes 2003).
In this study my aim was to quantify the responses of an arid-adapted bird community to temporal and spatial environmental variation. Here, I present data on the composition and abundance of a terrestrial bird community in a study area in arid north-western New South Wales over a four-year period that included both a prolonged drought and a time of above-average rainfall. I investigated the effects of regular seasonal variation and irregular climatic variation, especially rainfall, on avifaunal densities and species composition. I compared the responses of resident, nomadic and migratory species, different feeding groups and individual species by relating trends in the avifauna to rainfall. I also compared the avifauna of different habitats within the study area.
Methods
Study area
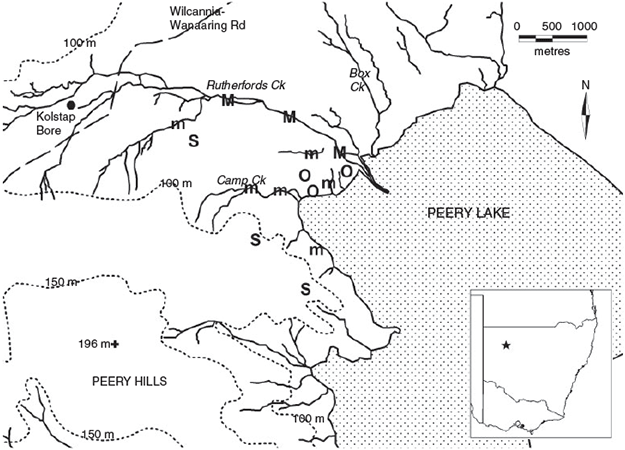
The study area (30°43′S, 143°33′E) encompasses ~1500 ha and abuts the north-west end of Peery Lake on the Paroo overflow system, some 50 km north-east of White Cliffs and 100 km north of Wilcannia (Fig. 1). At the time of the study the area was within a grazing leasehold, Peery Station, but is now part of Paroo–Darling National Park. Throughout the study the site was grazed by low numbers of livestock (sheep, Ovis aries) but the grazing pressure of macropods and feral animals, particularly goats (Capra hircus) and rabbits (Oryctolagus cuniculus), fluctuated and at times was high.
|
The study was undertaken between spring 1990 and summer 1994. Mean annual rainfall recorded at the two closest weather stations, Wilcannia and White Cliffs, was 258 mm and 245 mm respectively (Bureau of Meteorology records). Monthly records for Wilcannia and White Cliffs for the study period and preceding 12 months are highly correlated (R50 = 0.780, P < 0.001). I calculated the mean of the two sets of rainfall data to provide the best available indication of rainfall at the study site. Rainfall (Bureau of Meteorology records, G. Barlow, pers. comm.) in the year preceding the study was average. Between spring 1990 and spring 1992 the area received no effective rainfall. In December 1992 heavy local rain inundated the study area. The study thus comprised three periods: predrought with average rainfall in 1990, drought in 1991–92, and postdrought with above-average rainfall in 1993–94. There was water in Peery Lake throughout the study.
The study area included run-on (centred on drainage lines) and run-off areas, which were divided into four broad habitat types: (1) major creeklines, (2) minor creeklines (including one lake edge site with similar vegetation), (3) run-off areas with sparse perennial tall shrubs or low trees, and (4) run-off areas bereft of perennial trees and tall shrubs (open plains). Major creeklines were dominated by river red gum (Eucalyptus camaldulensis), black box (E. largiflorens) and river cooba (Acacia stenophylla) woodland. Minor creeklines were fringed by perennial shrubs and low trees, especially whitewood (Atalaya hemiglauca), prickly wattle (Acacia victoriae), mulga (A. aneura) and lignum (Muehlenbeckia florulenta). Run-off areas with perennial trees or tall shrubs included mulga scrub in the Peery Hills and harlequin eremophila (Eremophila duttonii) scrub on the plains. Vegetation on the open plains varied in response to rainfall and included short-lived grasses, herbs and subshrubs including saltbushes (Atriplex spp.) and copperburrs (Sclerolaena spp.). A detailed description of the study area is contained in Smith (1997).
Bird census
Fifteen fixed-width (50 m) census transects, divided into 100-m intervals, were established to sample the number of individuals and diversity of bird species and to compare these variables across the range of habitat variation at the site (Fig. 1). Run-on areas were sampled more intensively than run-off areas and three transects were placed in the major creekline and six in minor creeklines. In run-off areas three transects sampled areas of sparse trees and shrubs and three were on the open plains. Thirteen transects were 600 m in length, one 500 m, and one 400 m. The latter two transects were shorter due to the limited extent of the habitat type sampled. Transects within creeklines were separated by breaks of at least 200 m. The maximum distance between transects was 4000 m.
Transects were sampled each spring (October) and winter (July) between October 1990 and October 1993, and once each in autumn (April 1993) and in summer (January 1994), to give a total of nine census periods. Spring, winter and autumn censuses were restricted to a 4-h period after sunrise. I censused each transect on five different mornings in each census period. The order of counts was varied to spread counts for a given period throughout the morning and the census period. The direction of transect counts was also varied within each census period. Summer counts were made in the morning and late afternoon by two observers (JS and PS). In order to minimise observer variability (Kepler and Scott 1981; Kavanagh and Recher 1983), methods were standardised and two or three of the five counts for each transect were allotted to each observer.
In each census I took approximately 5 min to search each 100-m transect interval and record birds heard or seen. Flying birds were not counted unless obviously foraging within the census area. Waterbirds associated with Peery Lake were occasionally encountered but were not included in counts.
Habitat assessment
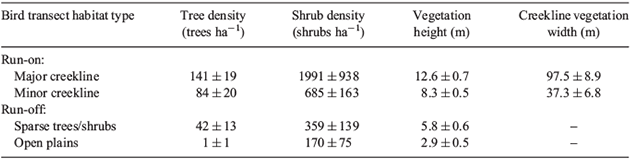
In each 100-m interval of the 15 bird census transects the following variables were measured to assess the range of habitats surveyed: (1) tree/shrub species composition (10 trees and 10 shrubs closest to centre point of interval identified to species), (2) tree/shrub density (density of all trees, all shrubs and each tree and shrub species, within the limits determined by the furthest of the 10 trees and shrubs recorded), (3) maximum height of vegetation, and (4) width of riparian vegetation at centre point of interval. Mean values of tree and shrub density, vegetation height and width of riparian habitat for each transect were calculated from the values of their 100-m intervals (Table 1).
|
Classification of birds
Birds were classified according to their movement patterns within the study area as residents, spring–summer or winter migrants, or nomads. Classification took account of observations over the study period and a banding study at the site (Smith 1997). Residents were present in all seasons and bred at the site. Spring–summer and winter migrants occurred regularly in the warmer or colder months, respectively, and included breeding and non-breeding species. They were absent at other times of the year, although occasional winter migrants were encountered at the start of spring census periods. Nomads occurred at irregular and unpredictable intervals. Birds were classified according to their use of feeding substrates into eight feeding groups: ground, ground/aerial, ground/bark, ground/fruit, foliage, flower/foliage, aerial and live vertebrate/carrion, based on an analysis of foraging behaviour at the site (Smith 1997).
Data analyses
In each census period the mean density (birds ha–1) of all birds in each transect was calculated. An overall index of site density was derived from the mean density of all transects in that census period. Multivariate methods (PATN Pattern Analysis Package) were used to compare bird species composition over the study area as a whole in different census periods. Data (mean density of each species in each census period) were transformed with log10(x + 1). Transformation meant that less emphasis was placed on bird abundance and more on species composition of census periods. Dissimilarity between census periods was calculated using the Bray–Curtis measure (Belbin 1990). An agglomerative hierarchical clustering strategy, flexible UPGMA (Unweighted Pair Group ArithMetic Averaging) with β = –0.1, and an ordination procedure, Semi-strong-hybrid Multidimensional Scaling (Belbin 1990), were used to elucidate patterns in the data.
Bird census results were compared with rainfall in the previous month and with the cumulative totals of the previous 2–12 months to allow for a time lag in response to rainfall. The overall density of birds and the numbers of species of all birds, resident birds, spring–summer migrants, winter migrants and nomads recorded in each census period, were related to the 12 sets of rainfall data using a correlation analysis.
Bird density and species richness differences between habitat types were tested by one-way analysis of variance (ANOVA), with post hoc testing of each pair of means by Tukey’s Method, using the SPC for Excel ver. 4 statistics package (BPI Consulting LLC, Cypress, Texas). Five habitat types were considered, consisting of the four types described above, with the minor creeklines subdivided for analysis into those associated with hilly country and those on the plains. Each habitat type was represented by three transects and the data from all census periods were pooled for analysis. The variables tested were, first, the mean number of birds per hectare recorded per census in each transect and, second, the mean number of bird species recorded per census period in each transect (i.e. the cumulative total over five individual censuses). The data were tested for normality (Anderson–Darling Test) and for homogeneity of variance (Bartlett’s Test), and the bird density data were log10-transformed before analysis in order to achieve normality.
Results
Avifauna
The study area supported a rich avifauna, with a total of 95 native land bird species recorded (Appendix 1). Well represented families (Christidis and Boles 2008) included honeyeaters and chats, woodswallows and allies, hawks and eagles, thornbills and allies, parrots, and pigeons and doves. In total, 81 species were recorded in censuses: 42 residents, 28 nomads, 6 spring–summer migrants, and 5 winter migrants. They included 29 ground feeders, 12 foliage feeders, 11 aerial feeders, 11 flower/foliage feeders, 9 live vertebrate/carrion feeders, 5 ground/fruit feeders, 2 ground/aerial feeders and 2 ground/bark feeders. Species not recorded in censuses included wide-ranging birds of prey, nocturnal birds and rare species.
Avifauna of the nine census periods
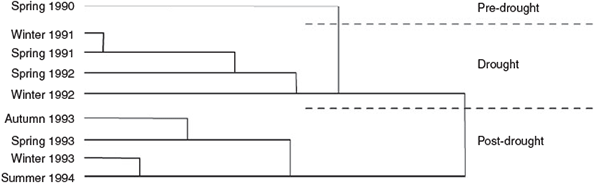
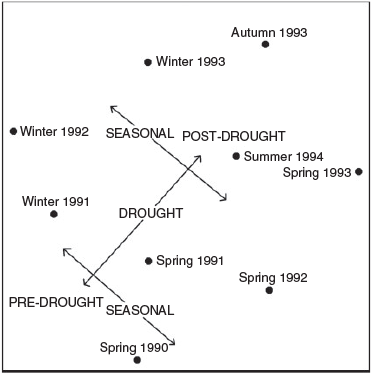
The composition and abundance of the avifauna changed constantly in the study period. UPGMA analysis indicated major differences between the predrought (1990), drought (1991–92) and postdrought (1993–94) periods of the study (Fig. 2). The greatest change in birds coincided with the breaking of the drought, between spring 1992 and autumn 1993. The ordination of results (Fig. 3) revealed a seasonal, as well as an annual, component of variation in the avifauna, with spring censuses generally separating from winter censuses.
|
|
Temporal patterns in abundance
Overall numbers of birds at the site declined quickly with the onset of drought, dropping from a mean transect density of 19.5 (±4.7, s.e.) birds ha–1 in spring 1990 to 9.6 (±2.2, s.e.) birds ha–1 the following winter. Bird numbers remained low throughout the drought and reached a minimum of 8.7 (±2.2, s.e.) birds ha–1 in spring 1992 (Fig. 4a). Following the breaking of the drought, bird numbers were high in autumn and spring 1993, but low in winter and summer. Variation between transects in terms of bird density was greatest in spring 1990, before the drought, and in autumn and spring 1993, after the drought.
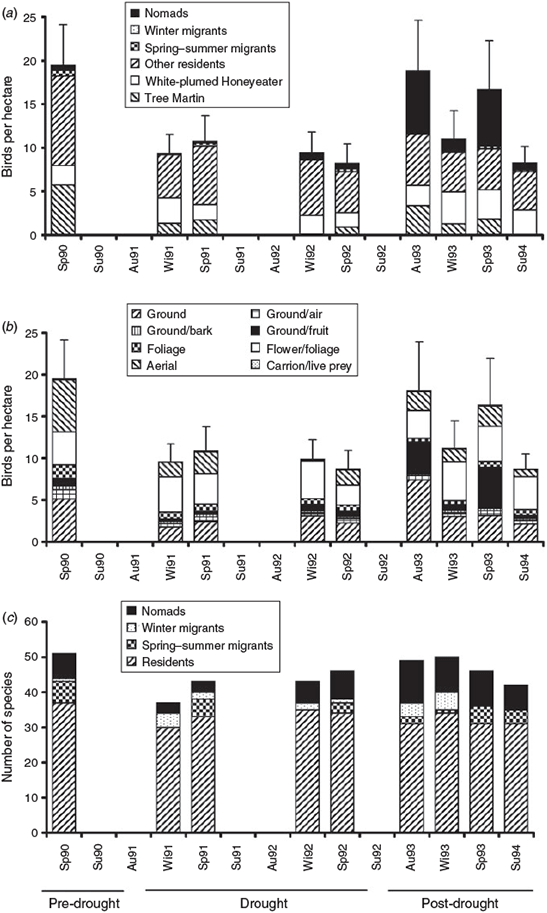
|
In the spring 1990 predrought survey, the avifauna was dominated by resident birds, with the tree martin (Petrochelidon nigricans) and white-plumed honeyeater (Lichenostomus penicillatus) easily the most numerous species. The fall in overall numbers between spring 1990 and winter 1991 was largely attributable to a decline in numbers of tree martins and other residents, but not the white-plumed honeyeater, whose numbers remained fairly constant (Fig. 4a). In contrast, the postdrought increase in numbers resulted mainly from an influx of nomadic species, such as the zebra finch (Taeniopygia guttata), budgerigar (Melopsittacus undulatus), crimson chat (Ephthianura tricolor), diamond dove (Geopelia cuneata) and rufous songlark (Cinclorhampus mathewsi). Numbers of spring–summer and winter migrants were low throughout the study.
Before the drought (spring 1990), aerial feeders, flower/foliage feeders and ground feeders dominated the avifauna (Fig. 4b). Numbers of flower/foliage feeders remained fairly constant throughout the study, while numbers of aerial and ground feeders declined when drought set in. Numbers of ground feeders peaked in autumn 1993 after the drought and ground/fruit feeders were most numerous in the autumn and spring after the drought.
Temporal patterns in species richness
Fluctuations in numbers of species were less pronounced than density changes. The numbers of species recorded in transects ranged from 51 in spring 1990 to 37 in winter 1991 (Fig. 4c). Differences in numbers of species recorded in winter and spring were not consistent between years. Residents accounted for 63% (autumn 1993) to 81% (winter 1991) of all species in each census. The number of nomadic species was lowest in 1991, the first year of the drought, and highest in 1993, the year after the drought. There were only modest changes in species richness of all foraging groups through the study period. The ground-feeding group was most rich in species, with ground feeders accounting for at least 35% (spring 1992) and up to 45% (summer 1994) of species in transects.
Bird/rainfall patterns
Total bird density and the density of residents, spring–summer migrants and winter migrants in each census period were not correlated with total rainfall in any of the previous 1–12-month periods (all P > 0.05). However, the density of nomads was significantly correlated with rainfall in the previous three-, four-, five-, 10-, 11- and 12-month periods (R7 = 0.698–0.793, P < 0.05). Similarly, the total number of species and number of resident, spring–summer migrant and winter migrant species were not correlated with rainfall. The number of nomadic species recorded was significantly correlated with rainfall in the previous 6–12-month periods (R7 = 0.721–0.867, P < 0.05).
Differences between habitat types
One-way ANOVA indicated significant differences between habitat types in both the mean bird density per census per transect (F4,10 = 28.461, P < 0.001) and the mean number of bird species per census period per transect (F4,10 = 9.421, P = 0.002). Post hoc comparison of means for each pair of habitats using Tukey’s Method showed no significant differences between the two minor creekline categories (hills and plains) and these have been combined below.
Mean bird density in major creekline transects (34.0 ± 2.1 birds ha–1) was significantly higher than in all other habitat types. Mean bird density in minor creekline transects (10.5 ± 1.4 birds ha–1) was significantly higher than in run-off transects without trees/tall shrubs (1.8 ± 0.5 birds ha–1), while mean bird density in run-off transects with trees/tall shrubs was intermediate between the two and not significantly different from either (4.2 ± 0.9 birds ha–1).
Mean species richness showed a similar increase from run-off transects without trees/tall shrubs (5.3 ± 1.5 species per census period), through run-off transects with trees/tall shrubs (9.7 ± 1.2 species per census period) and minor creekline transects (14.4 ± 1.6 species per census period) to major creekline transects (19.2 ± 0.9 species per census period), but the differences were significant only for the comparison of the major creeklines with the two run-off categories, and the comparison of the minor creeklines of the hills with the run-off areas without trees/tall shrubs.
Individual species
The magnitude and direction of population fluctuations of individual species were not consistent across transects over the study period. The mean density of each species in each census period is given in Appendix 1, together with the number of census transects (maximum 15) in which the species was recorded. Of the 22 most common residents (mean density over all transects greater than 1 bird per 10 ha) in spring 1990, the densities of all but the white-plumed honeyeater and grey shrike-thrush (Colluricincla harmonica) were lower in spring 1993. Numbers of white-plumed honeyeaters increased by over 50% between spring 1990 and spring 1993, but those of the grey shrike-thrush increased only slightly. In the same period, numbers of tree martins declined by two thirds. The small, similar-sized chestnut-rumped thornbill (Acanthiza uropygialis), yellow-rumped thornbill (A. chrysorrhoa), variegated fairy-wren (Malurus lamberti) and white-winged fairy-wren (M. leucopterus) were present in roughly equivalent numbers in spring 1990. Population declines were greatest in the chestnut-rumped and yellow-rumped thornbills, whose densities declined by factors of 11 and 8 respectively. In contrast, densities of the variegated and white-winged fairy-wrens were only about halved by spring 1993.
Total numbers of nomadic birds peaked after the drought in autumn and spring 1993 but numbers of individual nomadic species peaked at different times (Fig. 5). The autumn peak was dominated by zebra finches and, to a lesser extent, budgerigars. In spring 1993, budgerigars were over nine times as numerous as zebra finches. Diamond doves occurred in similar numbers in autumn and spring 1993. Numbers of white-fronted honeyeaters (Purnella albifrons) and white-browed woodswallows (Artamus superciliosus) peaked during the drought, the white-fronted honeyeaters in winter 1992 and the white-browed woodswallows in spring 1992. The peaks in numbers of these species were, however, much lower than those recorded for the zebra finch and budgerigar.
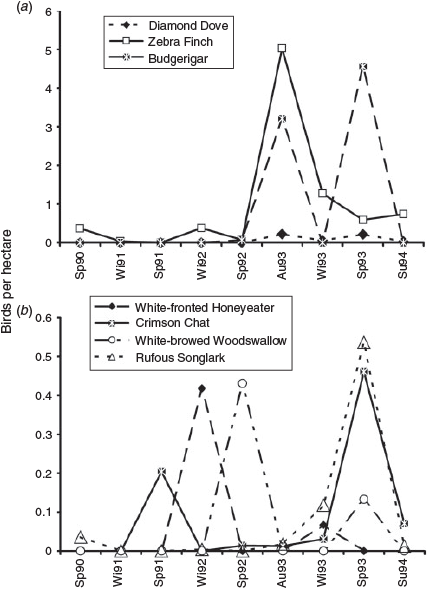
|
Discussion
This study has documented the dynamic and variable nature of a terrestrial bird community in a study area in arid north-western New South Wales. Over four years, which included periods of both drought and above-average rainfall, I found pronounced changes in the abundance and species composition of the avifauna. While overall bird abundance and species richness declined during the drought, the magnitude and direction of population fluctuations of individual species were not consistent. Within the study area, temporal variability was coupled with the spatial variability of the avifauna in different habitats.
The avifauna of the study area was dominated by resident species. Overall, 52% of birds recorded in censuses were classified as residents, 35% as nomads and the remainder (13%) as regular migrants. A predominance of resident species has also been recorded in land bird communities elsewhere in the arid zone (Pianka and Pianka 1970; Wyndham 1978; Brooker et al. 1979; Henle 1989; Pavey and Nano 2009). The proportion of nomads at Peery is only slightly higher than the figure of 26% estimated by Keast (1959) for the entire Australian avifauna. Though many birds in arid Australia are nomadic, nomadism is not, as once thought (e.g. Keast 1959; Immelmann 1963; Serventy 1971), the most common strategy adopted by birds in response to environmental unpredictability.
My classification of species as residents, nomads or migrants, based on movement patterns within the study area, differs from other classifications in the Australian arid zone (e.g. Schmidt 1978; Wyndham 1978; Burbidge and Fuller 2007; Pavey and Nano 2009). Differences in classifications may reflect differences in habitat quality for particular species in different localities and times as well as the scale of studies. Further, the distinction between movement classes may not always be clear cut. In my study area, the tree martin, for example, was classified as a resident. It was present in all census periods and nested each spring. One banded pair nested in the same hollow in consecutive springs (Smith 1997). However, huge fluctuations in tree martin numbers (some sixty-fold overall) suggested a mix of nomadic and sedentary individuals.
Changes in the avifauna over the study period could be related to both drought and, to a lesser extent, seasonal (winter–spring) variability. The onset of drought was accompanied by a marked decline in numbers of resident birds, especially in transects closest to Peery Lake. The proximity of fresh water in the lake, which was full before the drought and receded during the drought, may have contributed to the particularly high predrought densities in transects near the lake margin. Residents were slow to recover when conditions improved after the drought and the lake refilled, presumably because they had to breed and rear young rather than just move into the area.
Numbers of all feeding groups apart from flower/foliage feeders declined in the drought. Flower/foliage feeders included the nomadic white-fronted honeyeater and resident white-plumed, spiny-cheeked and singing honeyeaters (Lichenostomus virescens) and yellow-throated miner (Manorina flavigula). Perennial plant species, which flowered regularly in the study period despite the drought, were the chief food source for this group. During the period of this study white-fronted honeyeaters fed chiefly at the flowers of the perennial, winter-flowering harlequin eremophila (Smith 1997). Numbers of white-fronted honeyeaters peaked in winter 1992, at the height of the drought. They were absent from the site in winter 1991 and occurred in only very low numbers in winter 1993, despite there being no obvious differences in the abundance of harlequin eremophila flowers in the three winter periods. Resident flower/foliage feeders used a broad range of feeding substrates. Eucalypts in major creeklines, which supplied carbohydrates in the form of lerps and galls, as well as insects, were their main site for foliage gleaning (Smith 1997). These eucalypts tap supplies of subterranean water and may, to some extent, be buffered from the effects of drought. The resident flower/foliage feeders probably suffered less than other residents in the drought due to the relative stability and variety of their food supply. Measures of flowering intensities and invertebrate abundances, availabilities and depletion rates in major creeklines, in both drought and non-drought periods, would be needed to confirm this.
The relative stability of the food supply of some resident species in a period of short-term drought may, in part, explain why numbers of residents were not correlated with rainfall in the previous 12 months. Additionally, residents were slow to respond to improved conditions. Given their depleted numbers and their generally regular, rather than opportunistic, breeding patterns (Smith 1997), they may have been incapable of rapid population increases. Rates of population change will also be affected by factors not directly attributable to rainfall, for example, predation pressure. Predators of birds at the site included feral cats, foxes, snakes, goannas and avian predators including the collared sparrowhawk (Accipiter cirrocephalus) and Australian hobby (Falco longipennis).
Migrants provided an element of seasonal variation but their low numbers made it difficult to discern trends. The lack of correlation between rainfall and numbers of migratory birds was not surprising, considering the regularity of their movements and the additional influences on their populations of environmental conditions outside the study area.
The study emphasised the greater mobility and flexibility of nomads compared with residents and migrants. During the drought nomads were largely absent. The breaking of the drought in summer 1992–93 was followed by the appearance of large numbers of nomads, especially zebra finch and budgerigars. However, not all nomads fluctuated on the time scale of local rainfall. The population peaks of the various nomads were asynchronous and relatively short-lived. The autumn and spring 1993 peaks in numbers of nomads were of similar size, but differed in species composition. In autumn 1993, nomads consisted largely of small granivorous zebra finches, budgerigars and diamond doves. In spring 1993, budgerigars were common and numbers of zebra finches and diamond doves were reduced. The insectivorous crimson chat and rufous songlark were more common in spring than autumn. In contrast to these general trends in nomad numbers, populations of some nomads peaked during the drought. The white-fronted honeyeater, as noted previously, was most common at the height of the drought. Numbers of white-browed woodswallows peaked during the drought in spring 1992, when they also nested and fledged young. Observations of white-browed woodswallows have suggested that, in times of inland drought, they are nomadic invaders of coastal areas in which conditions are relatively more suitable (Recher and Schulz 1983; Wood 1994). On this occasion, an isolated thunderstorm in the study area several weeks before the spring 1992 census may have induced them to settle and breed at the site, despite the widespread and severe drought. At the study site, the white-browed woodswallow fed almost exclusively on aerial invertebrates (Smith 1997), but without measures of abundance of aerial invertebrates it is not possible to say whether white-browed woodswallows had been attracted by an increased availability of food. Numbers of all nomadic species, including the white-browed woodswallow, must, however, be determined to some extent by the relative availability of food outside compared with inside the study area.
Overall patterns discerned in this study are indicative of the generally deleterious effects of drought on birds. As found elsewhere in the arid zone (Reid et al. 1993; Paltridge and Southgate 2001; Burbidge and Fuller 2007), both species richness and overall abundance declined during the drought. The varied responses of different species suggest that drought has a differential effect on resources. Different species, each with unique habitat and food requirements and differing ability to migrate, respond accordingly. Studies in arid areas outside Australia have also shown that species vary greatly in abundance from year to year and place to place, apparently to some degree independently of each other (Wiens and Rotenberry 1980, 1981; Rosenberg et al. 1982; Rice et al. 1983a, 1983b).
The spatial variability found in the avifauna of the study area reflects the spatial heterogeneity of arid areas at a local level. In the study area, the relatively mesic creekline habitats supported significantly more birds and contained more species than habitats of run-off areas. The importance of creekline habitats to birds of arid areas is well supported by other studies (Pianka and Pianka 1970; Wyndham 1978; Brooker et al. 1979; Shurcliff 1980; Badman 1989; Henle 1989). Creekline habitats are structurally more complex and more productive than run-off habitats. They provide diverse feeding opportunities for birds and are a source of nesting, roosting and watering sites. They also offer protection from predators for birds that feed in and outside of creeklines. Thirteen species were recorded only in creekline transects. Only one species, the white-fronted chat, was recorded only in run-off transects. The avifauna of run-off areas, however, was not merely a depauperate sample of the avifauna of creekline habitats. Several species, including the southern whiteface (Aphelocephala leucopsis), red-capped robin (Petroica goodenovii), Australasian pipit (Anthus novaeseelandiae) and crimson chat, were largely confined to run-off areas and only ventured into the upper reaches of creeklines that graded into run-off habitat.
The findings of this study are in contrast to the findings of Cody (1994) and Pavey and Nano (2009), who concluded that assemblage patterns of arid birds are predictable and are not driven by temporal and spatial variability in food and water resources. The different findings may reflect the differing spatial and temporal scales of the studies. Cody (1994) and Pavey and Nano (2009) investigated avian assemblages at a landscape level (in chenopod associations, mulga shrubland and riverine woodland) whereas this study investigated variability in the avifauna in relation to local habitat heterogeneity. In addition, neither Cody (1994) nor Pavey and Nano (2009) investigated temporal variability in avian community structure which, in this study, was found to be pronounced.
Birds were censused to provide a relative rather than an absolute measure of abundance. Comparison of results of this study with results of other studies using different counting methods needs to be made with great caution – density estimates derived using different methods and observers are rarely comparable. Further, the temporal and spatial heterogeneity of the arid-zone avifauna, as revealed in this study, needs to be considered. Within the arid zone, Cody (1994), in mulga scrub/open woodland between late winter and early summer, estimated that the density of birds was 7.0–17.0 birds ha–1, while Wiens (1991), in chenopod scrubland in summer, estimated 0.9–2.1 birds ha–1. These figures are towards the lower end of densities recorded in the various habitats in my study area. In eucalypt forest and woodland in more mesic regions, abundances vary considerably, but are generally higher than abundances estimated in arid areas. Most densities reported in south-eastern Australian woodland, low open-forest and tall open-forest range from 12 to 24 birds ha–1, with the greatest densities (30–35 birds ha–1) recorded in tall open-forest (Recher 1985). Densities recorded in tall open-forest are comparable to densities estimated in the most mesic habitat in the study area, riparian eucalypt woodland of major creeklines.
Effective land management and conservation of the arid zone avifauna requires an understanding of the variability and dynamics of bird communities at both the local and regional level. Discernment of trends in the avifauna requires sound and comprehensive baseline data against which change can be measured. Studies such as this can detect patterns and lead to hypotheses. However, given the great variability from year to year and place to place within the Australian arid zone, it will be extremely difficult to design and implement meaningful experiments to test these hypotheses. More could be gained, however, by implementing concurrent and standardised bird community studies at a range of sites in the arid zone over long periods. Such studies would need to take account of the spatial heterogeneity of arid areas at both local and regional level. A proper understanding of the variability and dynamics of the avifauna is especially important as climate change is predicted to exacerbate the climatic variability and unpredictability of the arid zone in future years.
Acknowledgements
Hugh Ford and Harry Recher supervised this study. Peter, Stephen, Robert and Kate Smith assisted in the field. Lola Smith helped compile data. Greg and Kathy Barlow allowed me to camp and work on their property, Peery Station. I thank them all. The manuscript was much improved by the comments of editors and two anonymous referees. I am grateful for financial assistance provided by an Australian Postgraduate Research Award, Linnean Society of New South Wales (Joyce W. Vickery Scientific Research Fund), Ecological Society of Australia (Student Travel Award) and Australian Rangeland Society (Travelling Fellowship). My special thanks to Peter Smith, who encouraged me throughout.
References
Badman, F. J. (1989). ‘The Birds of Middle and Lower Cooper Creek in South Australia.’ (Nature Conservation Society of South Australia: Adelaide.)Barnard, C. (1917). Bird life as affected by drought. Emu 16, 234–236.
| Bird life as affected by drought.Crossref | GoogleScholarGoogle Scholar |
Barnard, H. G. (1927). Effects of drought on bird-life in central Queensland. Emu 27, 35–37.
| Effects of drought on bird-life in central Queensland.Crossref | GoogleScholarGoogle Scholar |
Belbin, L. (1990). ‘Technical Reference Pattern Analysis Package.’ (CSIRO Division of Wildlife and Ecology: Australia.)
Berney, F. L. (1906). Drought and flood in Queensland. Emu 6, 26.
| Drought and flood in Queensland.Crossref | GoogleScholarGoogle Scholar |
Berney, F. L. (1928). Birds and drought in central-western Queensland. Memoirs of the Queensland Museum 9, 194–195.
Brooker, M. G., Ridpath, M. G., Estbergs, A. J., Bywater, J., Hart, D. S., and Jones, M. S. (1979). Bird observations on the north-western Nullabor Plain and neighbouring regions, 1967–1978. Emu 79, 176–190.
| Bird observations on the north-western Nullabor Plain and neighbouring regions, 1967–1978.Crossref | GoogleScholarGoogle Scholar |
Burbidge, A. A., and Fuller, P. J. (2007). Gibson Desert birds: responses to drought and plenty. Emu 107, 126–134.
| Gibson Desert birds: responses to drought and plenty.Crossref | GoogleScholarGoogle Scholar |
Burbidge, A. A., and McKenzie, N. L. (1989). Patterns in the modern decline of Western Australia’s vertebrate fauna: causes and conservation implications. Biological Conservation 50, 143–198.
| Patterns in the modern decline of Western Australia’s vertebrate fauna: causes and conservation implications.Crossref | GoogleScholarGoogle Scholar |
Christidis, L., and Boles, W. E. (2008). ‘Systematics and Taxonomy of Australian Birds.’ (CSIRO Publishing: Melbourne.)
Cody, M. L. (1994). Mulga bird communities. 1. Species composition and predictability across Australia. Australian Journal of Ecology 19, 206–219.
| Mulga bird communities. 1. Species composition and predictability across Australia.Crossref | GoogleScholarGoogle Scholar |
Foley, J. C. (1957). Droughts in Australia. Review of records from earliest years of settlement to 1955. Bureau of Meteorology Bulletin 43.
Ford, H. A. (1989). ‘Ecology of Birds.’ (Surrey Beatty: Sydney.)
Garnett, S. T., Szabo, J. K., and Dutson, G. (2011). ‘The Action Plan for Australian Birds.’ (CSIRO Publishing: Melbourne.)
Henle, K. (1989). A two year avifaunistic survey in Kinchega National Park western New South Wales. Australian Birds 22, 53–68.
Hughes, L. (2003). Climate change and Australia: trends, projections and impacts. Austral Ecology 28, 423–443.
| Climate change and Australia: trends, projections and impacts.Crossref | GoogleScholarGoogle Scholar |
Immelmann, K. (1963). Drought adaptations in Australian desert birds. In ‘Proceedings of the 13th International Ornithological Congress, Ithaca 17–24 June 1962’ (Ed. C. G. Sibley) pp. 649–657. (International Ornithological Congress: Ithaca, NY)
Kavanagh, R., and Recher, H. F. (1983). Effects of observer variability on the census of birds. Corella 7, 93–100.
Keast, A. (1959). Australian birds: their zoogeography and adaptations to an arid continent. In ‘Biogeography and Ecology in Australia’. (Eds A. Keast, R. L. Crocker and C. S. Christian.) pp. 95–114. Monographie Biologicae 8. (W. Junk: The Hague.)
Kepler, C. B., and Scott, J. M. (1981). Reducing bird count variability by training. Studies in Avian Biology 6, 366–371.
Leeper, G. W. (1970). Climates. In ‘The Australian Environment’. (Ed. G. W. Leeper.) pp. 12–20. (CSIRO: Melbourne.)
MacGillivray, W. D. K. (1929). Through a drought stricken land. Emu 29, 113–129.
| Through a drought stricken land.Crossref | GoogleScholarGoogle Scholar |
McGilp, J. N. (1924). Seasonal influences on the breeding of native birds. Emu 24, 155.
| Seasonal influences on the breeding of native birds.Crossref | GoogleScholarGoogle Scholar |
Morton, S. R. (1990). The impact of European settlement on the vertebrate animals of arid Australia: a conceptual model. Proceedings of the Ecological Society of Australia 16, 201–213.
Morton, S. R., Stafford Smith, D. M., Dickman, C. R., Dunkerley, D. L., Friedel, M. H., McAllister, R. R. J., Reid, J. R. W., Roshier, D. A., Smith, M. A., Walsh, F. J., Wardle, G. M., Watson, I. W., and Westoby, M. (2011). A fresh framework for the ecology of arid Australia. Journal of Arid Environments 75, 313–329.
| A fresh framework for the ecology of arid Australia.Crossref | GoogleScholarGoogle Scholar |
Paltridge, R., and Southgate, R. (2001). The effect of habitat type and seasonal conditions on fauna in two areas of the Tanami Desert. Wildlife Research 28, 247–260.
| The effect of habitat type and seasonal conditions on fauna in two areas of the Tanami Desert.Crossref | GoogleScholarGoogle Scholar |
Pavey, C. R., and Nano, C. E. M. (2009). Bird assemblages of arid Australia: vegetation patterns have a greater effect than disturbance and resource pulses. Journal of Arid Environments 73, 634–642.
| Bird assemblages of arid Australia: vegetation patterns have a greater effect than disturbance and resource pulses.Crossref | GoogleScholarGoogle Scholar |
Pianka, H. D., and Pianka, E. R. (1970). Bird censuses from desert localities in Western Australia. Emu 70, 17–26.
| Bird censuses from desert localities in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Recher, H. F. (1985). Synthesis: a model of forest and woodland bird communities. In ‘Birds of Eucalypt Forests and Woodlands: Ecology, Conservation, Management’. (Eds A. Keast, H. F. Recher, H. Ford and D. Saunders.) pp. 129–135. (Surrey Beatty & Royal Australasian Ornithologists Union: Sydney.)
Recher, H. F., and Lim, L. (1990). A review of current ideas of the extinction, conservation and management of Australia’s terrestrial vertebrate fauna. Proceeding of the Ecological Society of Australia 16, 287–301.
Recher, H. F., and Schulz, M. (1983). Observations on the breeding of white-browed woodswallows. Corella 7, 1–6.
Reid, J., and Fleming, M. (1992). The conservation status of birds in arid Australia. The Rangeland Journal 14, 65–91.
| The conservation status of birds in arid Australia.Crossref | GoogleScholarGoogle Scholar |
Reid, J. R. W., Kerle, J. A., and Morton, S. R. (1993). Uluru Fauna. The distribution and abundance of vertebrate fauna of Uluru (Ayers Rock–Mount Olga) National Park, N.T. Kowari 4. Australian National Parks and Wildlife Service, Canberra.
Reynolds, R. G., Watson, W. D., and Collins, J. D. (1983). Water 2000: Consultants Report No. 13. Water Resources. Aspects of drought in Australia. Department of Resources and Energy. Australian Government Publishing Service, Canberra.
Rice, J., Ohmart, R. D., and Anderson, B. W. (1983a). Turnovers in species composition of avian communities in contiguous riparian habitats. Ecology 64, 1444–1455.
| Turnovers in species composition of avian communities in contiguous riparian habitats.Crossref | GoogleScholarGoogle Scholar |
Rice, J., Ohmart, R. D., and Anderson, B. W. (1983b). Habitat selection attributes of an avian community: a discriminant analysis investigation. Ecological Monographs 53, 263–290.
| Habitat selection attributes of an avian community: a discriminant analysis investigation.Crossref | GoogleScholarGoogle Scholar |
Robertson, G. (1987). Effects of drought on a breeding population of wedge-tailed eagles Aquila audax. Emu 87, 220–223.
| Effects of drought on a breeding population of wedge-tailed eagles Aquila audax.Crossref | GoogleScholarGoogle Scholar |
Robin, L., Heinsohn, R., and Joseph, L. (Eds) (2009). ‘Boom & Bust. Bird Stories For A Dry Country.’ (CSIRO Publishing: Melbourne.)
Rosenberg, K. V., Ohmart, R. D., and Anderson, B. W. (1982). Community organisation of riparian breeding birds: response to an annual resource peak. The Auk 99, 260–274.
Schmidt, B. L. (1978). Birds of the Cobar region, New South Wales. Australian Birds 12, 61–86.
Serventy, D. L. (1971). Biology of desert birds. In ‘Avian Biology’. (Eds D. S. Farner and J. R. King.) pp. 287–339. (Academic Press: New York.)
Shurcliff, K. S. (1980). Vegetation and bird community characteristics in an Australian arid mountain range. Journal of Arid Environments 3, 331–348.
Smith, J. E. (1997). Ecological patterns in a terrestrial bird community in arid north-western New South Wales. Ph.D. Thesis, University of New England, Armidale.
Smith, P., and Smith, J. (1994). Historical changes in the bird fauna of western New South Wales: ecological patterns and conservation implications. In ‘Future of the Fauna of Western New South Wales’. (Eds D. Lunney, S. Hand, P. Reed and D. Butcher.) pp. 123–148. (Royal Zoological Society of New South Wales: Sydney.)
Stafford Smith, D. M., and Morton, S. R. (1990). A framework for the ecology of arid Australia. Journal of Arid Environments 18, 255–278.
Wiens, J. A. (1977). On competition and variable environments. American Scientist 65, 590–597.
Wiens, J. A. (1989). ‘The Ecology of Bird Communities.’ Vols. 1 and 2. (Cambridge University Press: Cambridge.)
Wiens, J. A. (1991). Ecological similarity of shrub-desert avifaunas of Australia and North America. Ecology 72, 479–495.
| Ecological similarity of shrub-desert avifaunas of Australia and North America.Crossref | GoogleScholarGoogle Scholar |
Wiens, J. A., and Rotenberry, J. T. (1980). Patterns of morphology and ecology in grassland and shrubsteppe bird populations. Ecological Monographs 50, 287–308.
| Patterns of morphology and ecology in grassland and shrubsteppe bird populations.Crossref | GoogleScholarGoogle Scholar |
Wiens, J. A., and Rotenberry, J. T. (1981). Habitat associations and community structure of birds in shrubsteppe environments. Ecological Monographs 51, 21–41.
| Habitat associations and community structure of birds in shrubsteppe environments.Crossref | GoogleScholarGoogle Scholar |
Wood, K. A. (1994). Drought-related movements of white-browed woodswallows in New South Wales. Australian Birds 28, 42–53.
Wyndham, E. (1978). Birds of the Milparinka district and Cooper Creek Basin. Emu 78, 179–187.
| Birds of the Milparinka district and Cooper Creek Basin.Crossref | GoogleScholarGoogle Scholar |

|