

Reproductive and vegetative responses of different accessions of Microlaena stipoides (Labill.) R.Br. to nitrogen applications and supplementary irrigation in southern Australia
I. H. Chivers A C and D. E. Aldous BA Native Seeds Pty Ltd, PO Box 133, Sandringham, Vic. 3191, Australia.
B School of Resource Management, The University of Melbourne, Burnley Campus, Richmond, Vic. 3121, Australia.
C Corresponding author. Email: ian@nativeseeds.com.au
The Rangeland Journal 27(2) 151-157 https://doi.org/10.1071/RJ05014
Submitted: 25 August 2000 Accepted: 22 June 2005 Published: 21 November 2005
Abstract
Native grasses such as Microlaena stipoides (Labill.) R.Br. have the potential to play an important role in pastures, revegetation and as turfgrasses, but their use to date has been limited by seed supply. The purpose of this study was to investigate, (i) the effect of nitrogen fertilisation on M. stipoides seed production, (ii) the effect of irrigation on seed production, and (iii) the variation among accessions in response to these 2 factors.
Microlaena stipoides accessions responded to added nitrogen with an increase in shoot growth, total amount of dry matter produced per plant and in reproductive characters at the end of the harvest season. Higher yields were recorded for high (split) nitrogen treatments in comparison to nil (control) or low nitrogen treatments, with added nitrogen extending seed production. Higher soil moisture regimes produced significantly more reproductive culms, more spikelets per culm, and more seeds per plant when compared to low soil moisture treatments. The frequency of irrigation had more effect on seed yield than the amount of water applied.
Microlaena stipoides appears to have a 2–step mechanism for regulating the yield of its summer inflorescences namely; the plant initially responds to a high frequency of rainfall or irrigation events by initiating culms. Numbers of culms and florets per culm that reach maturity are dependent on the volume of water applied. Such a mechanism would assist in the adaptation of this grass to a southern Australian climate, which often exhibits an erratic summer rainfall.
While there were differences among the accessions for both reproductive and vegetative characters, they gave a uniform response to the different nitrogen treatments. The responses of 3 accessions to different irrigation treatments were somewhat different but with respect to total number of spikelets produced per plant, they all responded in the same way. Different irrigation management schedules may be required for different cultivars for commercial seed production.
Additional keywords: seed production, split applications, weeping grass.
Introduction
Australia’s native grasses are highly adapted to the harsh Australian environment but their application in agricultural or other purposes has been very limited, with most emphasis being on introduced grasses. However, recent information has shown that many native grasses have advantages compared with introduced grasses for use in agriculture, revegetation and as amenity grasslands (Johnston et al. 1999).
The native year-long green perennial grass M. stipoides var. stipoides1 (weeping grass) has been evaluated as an alternative grass for pastures, revegetation and turfgrass use and has many traits that make it desirable for use in place of introduced grasses. It is more drought hardy, requires less fertiliser, tolerates higher temperatures, is more shade tolerant and tolerant of soils of higher acidity, and equally nutritious and palatable to grazing animals when compared with an introduced grass such as Lolium perenne (perennial ryegrass) (Turner 1891; Munnich et al. 1991; Whalley and Jones 1998). Given these desirable attributes there is a need to make seed of this grass more widely available commercially, rather than simply relying on naturally occurring stands as seed sources.
Domestication of a native grass involves several steps and one of the first is to investigate aspects of seed production (Lodge and Groves 1990; Smith and Whalley 2002). The ultimate aim of the domestication of this species is to produce substantial quantities of seed of selected cultivars under cropping conditions (Woods et al. 1991; Whalley and Huxtable 1993).
Work at the Botany Department at the University of New England, NSW, has shown that sufficient variability exists among accessions of this species found on the Northern Tablelands of NSW to permit selection of suitable accessions for specific purposes including pasture, turfgrass or amenity plantings (Whalley and Huxtable 1993; Whalley and Jones 1998). Similar differences have been found in many other grass species (Lodder and Groves 1989; Lodge 1993; Mitchell et al. 2001). The differences reported in these and other papers have concentrated on vegetative characters such as leaf blade width, colour and growth habit, but have also included reproductive characters such as seed retention and seed yield (Lodge 1993; Earl et al. 1994).
The view that native grasses are unresponsive to added fertilisers, particularly nitrogen, (Donald 1970) has since been shown to be incorrect for a range of species and soil types. Robinson (1976) and Lodge (1979) showed that a range of native grasses responded significantly to applied nitrogen by way of increased dry matter production as did Whalley and Jones (1998) for several M. stipoides ecotypes. Seed production of introduced grasses is known to be responsive to nitrogen applications (Rolston et al. 1997) but little is known about M. stipoides in this regard.
Irrigation is often an essential part of the process of seed production for many introduced species such as Agrostis stolonifera (creeping bentgrass) (Burr 2003) and Lolium perenne (Najda and Kruger 2003). Microlaena stipoides is an indeterminate seeder when grown in the high rainfall zones of New Zealand (Woods et al. 1991) and in northern New South Wales (Magcale-Macandog and Whalley 1994). In addition, summer seed head production often follows soon after a significant rainfall event in Victoria (I. H. Chivers, unpublished data). It appears that supplementary irrigation may be an important means of increasing seed production of M. stipoides.
This study was undertaken to investigate the effects of the application of nitrogen and irrigation on some aspects of seed production of a range of M. stipoides accessions found within a 50 km radius of Melbourne, Victoria.
Materials and methods
Two separate experiments were conducted. The first was located in the field and investigated the effect of nitrogen nutrition, while the second was in pots in a polyhouse and investigated the effects of irrigation regime. Both of these experiments included a range of accessions of Microlaena stipoides (Labill.) R.Br., so that comparisons of their responses to the various inputs could be made among them.
Experiment 1 – effect of nitrogen
Several accessions of M. stipoides had been previously collected from within a 50 km radius of Melbourne, and 6 were chosen for the first experiment. The selection of these 6 was based on leaf density, colour, texture and seed yield relative to other accessions in a pilot trial conducted in 1992.
Single plants of these six accessions were grown from seed in a nursery and transplanted as small plants (fewer than 10 leaves) into holes cut in black polythene weed mat at The University of Melbourne – Burnley Field Experimental Station on 15 June 1993. The trial had 5 fertiliser treatments (main plots) with 6 accessions nested within each fertiliser plot and four replications (blocks), resulting in a randomised split plot design. Twelve plants of each accession were planted in 2 adjacent rows in each fertiliser plot in a triangular pattern that allowed for 300 mm between adjacent plants. Plants were irrigated when transplanted and the only further water was applied at the times of fertiliser application. Plants were not mowed or sprayed for pests and diseases at any stage during the experiment. Hand weeding ensured that the experiment was kept weed free.
The nitrogen treatments involved a no N control plus a high and a low single application (27 October) 8 weeks before the anticipated harvest date as well as a high and low split application eight and (27 November) 4 weeks before harvest (Table 1). Aqueous urea (9 L of 46% N) solution (1.39 or 0.69 g of urea/L) was watered on to each fertilised plot using a watering can to give the desired concentrations on each plot at each time of application. Unfertilised plots were also watered with the 9 L of water at each time of application.

|
Soil samples were collected from the plot area (0–10 cm) before the experiment being set up and the N, P, K and total salts analysed (AOAC 1990). The soil was an alluvial fine textured clay-loam and was slightly acidic (pH of 6.2 in CaCl2) and of moderate fertility (N = 0.14%, P = 190 mg/L, K = 170 mg/L and total salts = 330 mg/L).
Ripe seed was harvested continually by hand over a 6 week period from late November 1993 to January 1994 by manually stripping off mature seeds that are held loosely on the culms using the method outlined by Waters et al. (2000). The seed collected each week was weighed. At the end of the harvest period, the plants were cut off at 50 mm above ground level, separated into vegetative and reproductive tillers, dried at 80°C for 24 h, then weighed.
Experiment 2 – effect of irrigation regime
Three accessions, EMR, QRY and BK9, were selected on the basis of their uniformity of phenotype and their seed production during Experiment 1.
One hundred plugs of each of the three accessions were removed from the field station and placed into 11-cm plastic pots using soil from the same location. The pots were placed in a polyhouse roofed with clear plastic with removable sides for ventilation. Light levels were similar to those outside. The plants were acclimatised for 10 weeks and watered using a semi-automatic sprinkler system. On 12 February 1995 the plants were trimmed to 40 mm above soil level and the experimental watering regime installed a day later.
An evaporative pan was installed adjacent to the pots to measure background evaporation in the polyhouse. This allowed an assessment of the amount of water required to recharge the soil to 50 or 100% of field capacity using the volume of soil in each pot, the volumetric soil water content at field capacity of 3 mm per cm depth, and the evaporative losses during the acclimatisation period. All pots started the experiment at field capacity.
There were 6 irrigation treatments (Table 2) and each treatment was applied to 12 pots of each of the three accessions using a completely randomised design. The 72 pots of each accession used in the experiment were chosen from the 100 prepared using a random number generator. Each pot was provided with a dripper that was positioned such that all water was retained in the pot. No fertiliser, herbicide, fungicide or any soil treatment other than irrigation was provided to the plants at any stage of this experiment.
|
Reproductive culm numbers in each pot were counted weekly from weeks 1–7, and at weeks 9 and 11. New and old culms were included in each assessment and the number of culms that had grown in the intervening periods between assessments was determined. Following the counting at week 11, the reproductive culms were removed by clipping above the level of the leaves of the vegetative tillers, and then all leaves and stems were clipped at 40 mm above ground level and referred to as the vegetative culms. The number of spikelets on each culm was counted and the numbers of spikelets per plant were derived by multiplying the number of culms by the number of spikelets per culm. All samples were weighed following oven drying at 80°C for 24 h.
Statistical analyses
Data were subjected to analysis of variance using the Minitab statistical package (Ryan and Joiner 1994) and means were separated using least significant differences (P < 0.05). Normality of data was confirmed using the Ryan–Joiner test.
Results
Experiment 1 – effect of nitrogen
There were no significant interactions between accessions and nitrogen treatments for the observed attributes (P > 0.05). In other words, all accessions responded in a similar way to the different nitrogen applications.
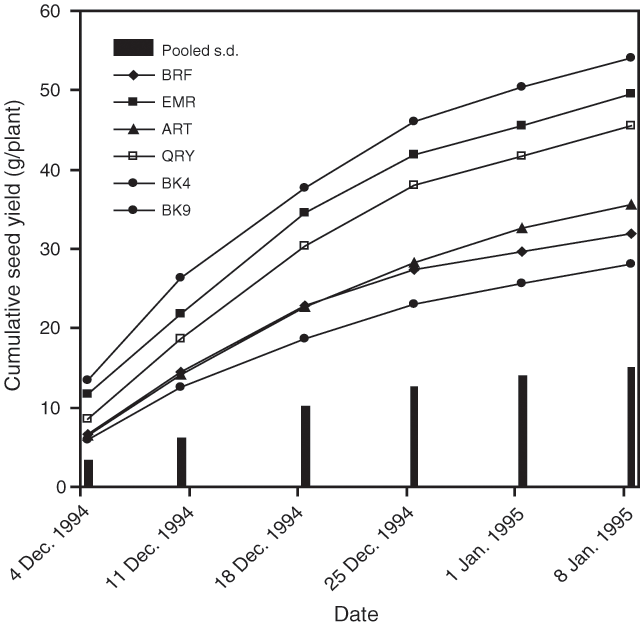
Significant differences existed for each assessment date and for total seed production among accessions (P < 0.05). Cumulative seed production showed that accessions BK9, EMR and QRY yielded significantly more seed than the other three at most dates and in total production averaged over all nitrogen treatments (Fig. 1).
|
There was a strong correlation (R2 = 0.761) between increased dry matter production and seed yield for all accessions averaged over all nitrogen treatments, i.e. increased dry matter accumulation was correlated with higher seed yield. For example for accession QRY the regression equation was total seed yield = 72.5 g + 13.4 × shoot dry weight (g) (R2 adj. = 0.69, P < 0.001).
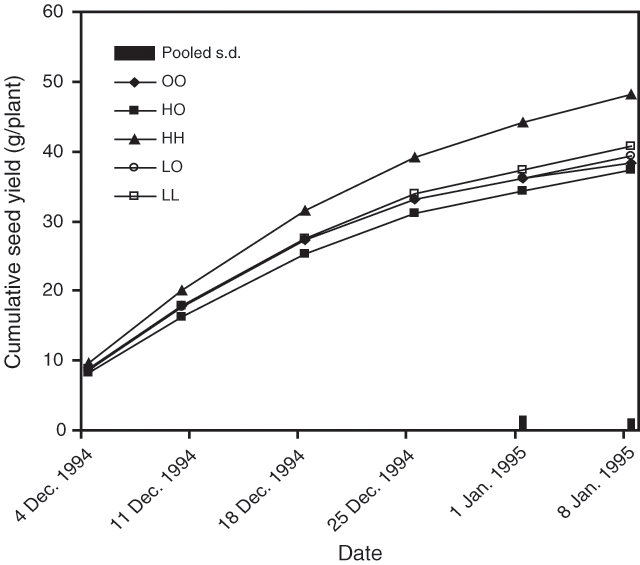
Cumulative seed production, averaged over all accessions for the early harvests, was consistently higher with the split nitrogen applications but the increase was not significant (P > 0.05). However, for the last 2 harvests of the season, the split (HH) nitrogen treatments produced significantly higher cumulative seed yields (P < 0.05) compared with the plots that received no nitrogen (OO) or only 1 nitrogen application (LO and HO). The plots that received the 2 applications of nitrogen, but at a lower rate (LL), were not significantly different from the higher split rate (HH) treatment. Final cumulative seed yield was 47.5 g/plant in the HH treatment compared with the other N treatments where the yield ranged from 37.3 to 40.7 g/plant. Nitrogen application increased vegetative culm dry weight and total dry weight of reproductive and vegetative culms. Results from this study demonstrated that split applications of nitrogen (HH) can result in significantly higher dry weight per plant (Fig. 2).
|
Experiment 2 – effect of irrigation
There were no mortalities under any of the experimental treatments, nor did any of the plants show wilting. Although the soil in the 33, 25 and 16.5% treatments (treatments 4,5 and 6, respectively) was dry and consolidated, the plants were not water stressed beyond wilting point.
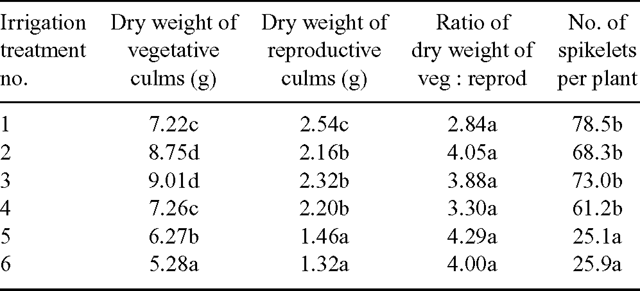
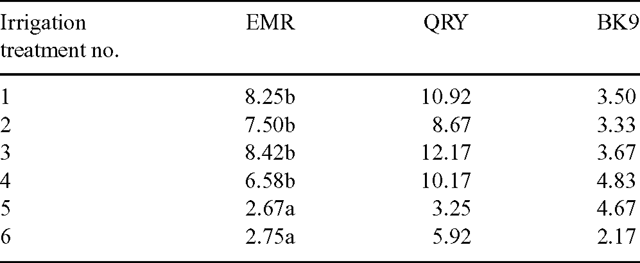
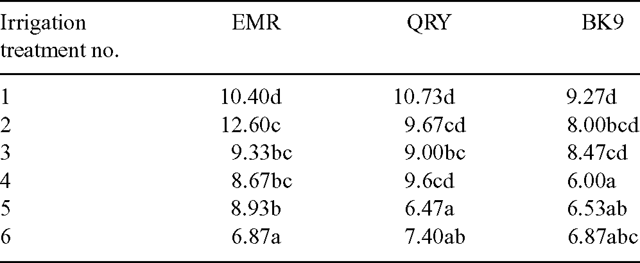
The accession × irrigation treatment interactions were not significant (P > 0.05) for the dry weight of vegetative culms, dry weight of reproductive culms and the number of spikelets produced per plant (Table 3). In other words, the 3 accessions gave the same response to the irrigation treatments for the above attributes. However, the accession × irrigation interactions were significant (P < 0.05) for the number of new reproductive culms per plant produced over the experimental period and the mean number of spikelets produced per culm (Tables 4 and 5).
|
|
|
Vegetative culm dry weight was significantly higher (P < 0.05) when pots were brought up to 50% capacity daily and 100% every second day (treatments 2 and 3) than when pots were brought up to 100% capacity every day (treatment 1) (Table 3). There was a further decline in vegetative culm dry matter production at the lowest frequency of irrigation (every third day) with the lowest value at 16.5% of the maximum amount of water applied.
The maximum dry weight of reproductive culms was produced when water was applied to bring the pots up to field capacity every day with significantly less (P < 0.05) produced in treatment 2 (replenishment to 50% of field capacity every day) or for the less frequent watering treatments (Table 3). The ratio of vegetative to reproductive culm dry weight was not affected by the irrigation regime (Table 3).
The lowest frequency of irrigation (every third day) reduced the number of new reproductive culms produced during the course of the experiment for accession EMR, but not for QRY and BK9 (Table 4). The amount of irrigation had no effect on the number of new reproductive culms. For this reason it is suggested that the frequency of irrigation, rather than the volume of irrigation, was the more critical factor in the initiation of new reproductive culms, at least for this accession.
Significantly fewer spikelets per culm (P < 0.05) were observed on plants receiving lower than on those receiving higher volumes of water (Table 5), but the pattern of the reduction with both frequency of irrigation and the amount of water applied varied among the accessions.
Irrigation frequency affected the number of spikelets produced per plant in that the two treatments that were irrigated every third day had significantly fewer spikelets per plant than the other more-frequently irrigated treatments (Table 3) and there was no significant accession × irrigation treatment interaction (P < 0.05). Thus the frequency of irrigation was more important than the amount of water applied in determining the number of spikelets produced by each plant over the course of the experiment. There was some compensation among the different accessions concerning the production of new reproductive culms and the number of spikelets per culm so that the effect of the frequency of irrigation on the number of spikelets produced per plant was similar for all three accessions.
Discussion
All 6 accessions of Microlaena stipoides responded to nitrogen with increases in total amount of dry matter production per plant and in higher seed yields. These results are consistent with the work of Vieira (1980), Robinson (1976) and Lodge (1979) who demonstrated increased vegetative growth over a range of native grasses with increased nitrogen application. In the present study, significantly higher yields were recorded for high (split) nitrogen treatments in comparison with nil (control) or low nitrogen treatments. The effect of nitrogen on seed yield occurred at the end of the season through more prolonged seed production. In the Melbourne environment seed yield can be improved by about 30% with split applications of nitrogen towards the end of the vegetative growth phase. In other words, the second N application may extend the harvesting season for M. stipoides, if other factors, such as rainfall or temperature, are not limiting (Fig. 2). These results suggest that a single methodology can be adopted with respect to nitrogen fertilisation for all accessions of M. stipoides rather than needing to develop individual routines for particular cultivars. However, commercial experience with larger areas for seed production of different cultivars may show that this approach needs to be modified.
Some accessions produced more herbage mass and seed than others and these results are consistent with data presented for M. stipoides (Whalley and Jones 1998), Austrodanthonia spp. (wallaby grass) (Lodge 1993) and Themeda australis (kangaroo grass) (Sindel and Groves 1990) and confirm the capacity for the selection of desirable cultivars for different purposes from within natural populations of M. stipoides. There appears to be a strong relationship between the total dry matter accumulation and total seed yield and that seed production is not competitive with dry matter accumulation. Again, further commercial experience may show that this generalisation does not apply to all cultivars.
Total dry matter accumulation of the highest yielding lines (about 2.5 t/ha over a 14-week period during the summer), is consistent with the work by Robinson and Archer (1988) on the New England Tablelands of NSW which showed dry matter yields of 2.6 t/ha for the same species could be achieved over a similar period. Their work compared some native with common introduced grasses and found that the herbage mass production of the native grasses, including M stipoides, could be equal to or better than the introduced grasses.
Microlaena stipoides appears to have a 2-step mechanism for regulating seed yield in response to irrigation or rainfall events. It is suggested that the plant responds initially to a high frequency of rainfall or irrigation events by initiating culms. Whether the culms then become reproductive, and the number of spikelets per culm that actually reach maturity, will then depend on the volume of water applied or the rainfall received. In pots at the test site in Melbourne, it was shown that greater seed yields could be obtained by more frequent irrigation of M. stipoides, even with relatively small amounts of water, than from infrequent heavy irrigation. Such a mechanism has obvious benefits for the adaptation of this grass to a southern Australian climate with an erratic summer rainfall and for irrigation management for maximum seed production.
High frequency, low-volume irrigation should be used to stimulate summer culm production of M. stipoides in areas of southern Australia, which are unlikely to receive reliable summer rainfall. In such areas M. stipoides would not normally produce reproductive parts, but would instead remain in a vegetative state.
The decline in vegetative culm dry matter production below about 25% of the maximum amount of water applied, is reasonably consistent with results for turfgrass (Beard 1973) tropical grasses (Whiteman 1980) and temperate pasture grasses (Leeper and Uren 1993) when grown on similar soil types.
The relative allocation of dry weight to vegetative as opposed to reproductive culms was affected by fertiliser treatments under non-limiting irrigation (I. H. Chivers, unpublished data), and not by irrigation under non-limiting fertility. It is suggested that the decline in vegetative culm dry matter production below about 25% of the maximum amount of water applied could be a sign to re-apply water to enable the plant to survive. This suggests that reproductive growth for these accessions of M. stipoides is more sensitive to soil moisture status or the incidence of rainfall than it is to natural soil fertility. This is likely to be an adaptive response to the Australian climate where rainfall or moisture conditions are more variable than soil fertility conditions. However, where a seed production stand is being managed for maximum seed production, both split additions of nitrogen and frequent low-volume irrigation applications would appear to be the best strategy. The details of this strategy have yet to be worked out under commercial conditions.
Results from experiments 1 and 2 showed that the three accessions did have some different characteristics. Accession BK9 grew almost exclusively vegetatively after the spring peak of reproductive growth (early maturing), whereas accession EMR produced more reproductive culms throughout the year (late maturing). Accession QRY seemed to be highly productive throughout the summer as it produced high levels of both vegetative and reproductive culms during that time. This confirms results obtained elsewhere that there is substantial morphological variability among M. stipoides accessions in their vegetative and reproductive characters, and of the potential for successful collection and selection programmes to develop more desirable cultivars of this grass. It also means that the details of fertiliser and irrigation management for maximum seed production may have to be varied for different cultivars.
Acknowledgments
The authors would like to thank Ross Payne, The University of Melbourne, Burnley Campus, in assisting in the manufacture and installation of the evaporative pan.
AOAC
(1990)
Beard, J. B. (1973).
Burr J. B.
(2003) Production of Roundup resistant creeping bentgrass seed. : ‘Proceedings of the 5th International Herbage Seed Conference’. Gatton, Australia, November, 2003. (Ed. D. S. Loch )
pp. 38–42. (International Herbage Seed Group: Aberystwyth.)
Donald, C. M. (1970). Temperate pasture species. : ‘Australian grasslands’. pp. 303–320. (Australian National University Press: Canberra.)
Earl J. M., Whalley R. D. B., Jones C. E.
(1994) Variation in the components of seed yield and germination requirements among three accessions of Microlaena stipoides. : ‘Proceedings of the 8th Biennial Conference of the Australian Rangeland Society’
Johnston W. H.,
Clifton C. A.,
Cole I. A.,
Koen T. B.,
Mitchell M. L., Waterhouse D. B.
(1999) Low input grasses useful in limiting environments (LIGULE). Australian Journal of Agricultural Research 50, 29–53.

Leeper, G. W. ,
and
Uren, N. C. (1993).
Lodder, M. S. ,
and
Groves, R. H. (1989).
Lodge G. M.
(1979) Effect of fertility level on the yield of some native perennial grasses on the north-west slopes, New South Wales. Australian Rangeland Journal 1, 327–333.
Lodge G. M.
(1993) The domestication of the native grasses Danthonia richardsonii Cashmore and Danthonia linkii Kunth for agricultural use. 1. Selecting for inflorescence seed yield. Australian Journal of Agricultural Research 44, 59–67.
| Crossref | GoogleScholarGoogle Scholar |
Lodge G. M., Groves R. H.
(1990) The domestication and agronomy of native grasses. : ‘Proceedings of the Australian Native grass Workshop, Dubbo’. (Ed. P. M. Dowling ,
D. L. Garden )
pp. 73–86. (Australian Wool Corporation: Melbourne.)
Magcale-Macandog D. B., Whalley R. D. B.
(1994) Factors affecting the distribution and abundance of Microlaena stipoides (Labill.) R.Br. on the Northern Tablelands of New South Wales. Rangeland Journal 16, 26–38.
Mitchell M. L.,
Koen T. B.,
Johnston W. H., Waterhouse D. B.
(2001) LIGULE: an evaluation of indigenous perennial grasses of dryland salinity management in south-eastern Australia 2. Field performance and the selection of promising ecotypes. Australian Journal of Agricultural Research 52, 351–365.
| Crossref |
Munnich D. J.,
Simpson P. C., Nicol H. I.
(1991) A survey of native grasses in the Goulburn district and factors influencing their abundance. Rangeland Journal 13, 118–129.
Najda H., Kruger A.
(2003) Effect of time and method of establishment on seed yield of irrigated perennial ryegrass. : ‘Proceedings of the 5th International Herbage Seed Conference’. Gatton, Australia. (Ed. D. S. Loch )
pp. 123–127. (International Herbage Seed Group: Aberystwyth.)
Robinson G. G.
(1976) Productivity and response to nitrogen fertiliser of the native grass Danthonia racemosa (Wallaby grass). Rangeland Journal 1, 49–52.
Robinson G. G., Archer K. A.
(1988) Agronomic potential of native grass species on the northern tablelands of New South Wales. 1. Growth and herbage production. Australian Journal of Agricultural Research 39, 415–423.
| Crossref | GoogleScholarGoogle Scholar |
Rolston M. P., Rowarth J. T., Young W. C., Mueller-Warrant G.W.
(1997) Grass seed crop management. : ‘Forage seed production, Vol. 1: temperate species’. (Eds D. T. Fairey, J. G. Hampton)
pp. 105–126. (CAB International: Wallingford.)
Ryan B. F., Joiner B. L.
(1994)
Sindel B. M., Groves R. H.
(1990) Seed production potential and domestication of kangaroo grass (Themeda triandra). : ‘Proceedings of the Australian Native Grass Workshop, Dubbo’. (Ed. P. M. Dowling ,
D. L. Garden )
pp. 171–172. (Australian Wool Corporation: Melbourne.)
Smith S. R., Whalley R. D. B.
(2002) Model for expanded use of native grasses. Native Plants Journal 3, 38–49.
Turner F.
(1891) The grasses of New South Wales: Microlaena stipoides, R.Br. ‘Meadow rice grass’. Agricultural Gazette of NSW 2, 22–23.
Vieira A.
(1980) Grazing value of Microlaena stipoides (Labill.) R.Br. on sheep camps on the northern tablelands of New South Wales and its response to fertiliser when growing in a glasshouse. Dip. Sci. Agric. thesis.
(University of New England:
Armidale.)
Waters, C. J. ,
Whalley, R. D. B. ,
and
Huxtable, C. H. A. (2000).
Whalley R. D. B., Huxtable C. H. A.
(1993) Domestication of the Australian perennial native grass, Microlaena stipoides. : ‘Proceedings of the XVII International Grassland Congress’. Palmerston North, NZ. (Ed. M. J. Baker ,
J. R. Crush ,
L. K. Humphreys )
pp. 214–215. (New Zealand Grassland Association: Palmerston North.)
Whalley, R. D. B. ,
and
Jones, C. M. (1998).
Wheeler, D. J. B. ,
Jacobs, S. W. L. ,
and
Whalley, R. D. B. (2002).
Whiteman, P. C. (1980).
Woods P. M., Couchman J.N., Rumball P.J., Christie A.R.J., Goodwin B.M.
(1991) Domestication of Microlaena stipoides (Labill.) R.Br. for amenity use in northern New Zealand. : Proceedings of the Australian Native Grass Workshop, Dubbo’. (Eds P. M. Dowling, D. L. Garden)
pp. 169–170. (Australian Wool Corporation: Melbourne.)
1Nomenclature according to Wheeler et al. (2002).