

Diet quality, liveweight change and responses to N supplements by cattle grazing Astrebla spp. (Mitchell grass) pastures in the semi-arid tropics in north-western Queensland, Australia
R. M. Dixon A * , M. T. Sullivan B , S. N. O’Connor B D and R. J. Mayer CA The University of Queensland, QAAFI, St Lucia, Qld 4072, Australia.
B Department of Agriculture and Fisheries, Rockhampton, Qld 4701, Australia.
C Department of Agriculture and Fisheries, Maroochy Research Facility, Qld 4560, Australia.
D Present address: Bostan, Tambo, Qld 4478, Australia.
The Rangeland Journal 44(2) 97-113 https://doi.org/10.1071/RJ21056
Submitted: 17 November 2021 Accepted: 7 May 2022 Published: 23 June 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing on behalf of the Australian Rangeland Society. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Experiments during 4 years examined the diets selected, growth, and responses to N supplements by Bos indicus-cross steers grazing summer-rainfall semi-arid C4 Astrebla spp. (Mitchell grass) rangelands at a site in north-western Queensland, Australia. Paddock groups of steers were not supplemented (T-NIL), or were fed a non-protein N (T-NPN) or a cottonseed meal (T-CSM) supplement. In Experiment 1, young and older steers were measured during the late dry season (LDS) and the rainy season (RS), while steers in Experiments 2–4 were measured through the annual cycle. Because of severe drought the measurements during Experiment 3 annual cycle were limited to T-NIL steers. Pasture availability and species composition were measured twice annually. Diet was measured at 1–2 week intervals using near infrared spectroscopy of faeces (F.NIRS). Annual rainfalls (1 July–30 June) were 42–68% of the long-term average (471 mm), and the seasonal break ranged from 17 December to 3 March. There was wide variation in pasture, diet (crude protein (CP), DM digestibility (DMD), the CP to metabolisable energy (CP/ME) ratio) and steer liveweight change (LWC) within and between annual cycles. High diet quality and steer liveweight (LW) gain during the RS declined progressively through the transition season (TS) and early dry season (EDS), and often the first part of the LDS. Steers commenced losing LW as the LDS progressed. In Experiments 1 and 2 where forbs comprised ≤15 g/kg of the pasture sward, steers selected strongly for forbs so that they comprised 117–236 g/kg of the diet. However, in Experiments 3 and 4 where forbs comprised substantial proportions of the pasture (173–397 g/kg), there were comparable proportions in the diet (300–396 g/kg). With appropriate stocking rates the annual steer LW gains were acceptable (121–220 kg) despite the low rainfall. The N supplements had no effect on steer LW during the TS and the EDS, but usually reduced steer LW loss by 20–30 kg during the LDS. Thus during low rainfall years in Mitchell grass pastures there were substantial LW responses by steers to N supplements towards the end of the dry season when the diet contained c. <58 g CP/kg or c. <7.0 g CP/MJ ME.
Keywords: dietary protein, digestibility, faeces, forbs, grazing, near infrared spectroscopy, protein supplements, rangelands, steers.
Introduction
Rangelands in the seasonally dry tropics are characterised by a distinct, and often brief, summer rainy season (RS) followed by an extended dry season. Tropical C4 grasses grow and mature rapidly, and when senesced are highly fibrous, with low contents of metabolisable energy (ME) and key nutrients such as nitrogen (N) and phosphorus (Winks 1984; Minson 1990; Hogan 1996). In northern Australia, C4 Mitchell grass (Astrebla spp.) rangelands comprise about 0.45 million km2 generally within the 250–550 mm rainfall zone with usually fertile alkaline cracking clay soils, and are important for cattle and sheep production (Christian et al. 1952; Orr 1975; Orr and Phelps 2013a; Cowley et al. 2018). Below-average and variable rainfall resulting in short rainy seasons with extended dry seasons, especially in years with a late seasonal break, are major limitations to cattle growth and productivity.
Cattle productivity per head in Mitchell grass rangelands is generally considered higher than in most other rangeland regions in the seasonally dry tropics of northern Australia (Weston 1988; McLennan et al. 1999; Bortolussi et al. 2005). This is associated with generally fertile soils and senesced Mitchell grass pastures in semi-arid regions tending to maintain their nutritive value more than do pastures in higher-rainfall (e.g. >600 mm) areas such as the northern speargrass (Heteropogon spp.) and Aristida–Bothriochloa rangelands (Weston and Moir 1969; McGowan et al. 2014). However, dry-season rainfall can cause major spoilage and rapid loss of the amount and nutritive value of standing Mitchell grass pastures (Blake 1938; Davies et al. 1938; Orr 1975). Cattle management to achieve high productivity accommodates high between-year variation in rainfall and pasture growth through moderate stocking rates and timely adjustments of herd numbers in response to rainfall and pasture conditions.
Mitchell grass pastures contain other perennial and annual grasses and numerous dicotyledonous herbaceous forbs, with considerable variation in their frequency and biomass among years. Where there is winter rainfall or low grass biomass owing to low summer rainfall and/or heavy grazing, forbs may comprise a substantial proportion, or even dominate, the sward (Lorimer 1978; Orr et al. 1988; Cobon and Carter 1994; Orr and Phelps 2013b). Many forb species are palatable to sheep and cattle, and are generally higher in nutritive value than are grasses, especially during winter (Hall and Lee 1980; McMeniman et al. 1986a, 1986b). Sheep often strongly select for forbs, so that forbs can comprise a substantial part of the diet even when they comprise only a small proportion of the pasture (Lorimer 1981; McMeniman et al. 1986a, 1986b, 1989; Pritchard et al. 1986; Orr et al. 1988). The general lesser ability of cattle than sheep to select higher-quality pasture components, including in semi-arid environments in southern Australia (Wilson 1976; Graetz and Wilson 1980) and Africa (Fianu 1982), suggests that forbs are likely to comprise a lower proportion of cattle diets than sheep diets. The limited data available (Coates and Dixon 2007; Dixon et al. 2007a; Cowley et al. 2018) suggest that the forbs often contribute to the diet of cattle, but more thorough knowledge of the diets of cattle grazing Mitchell grass pastures through seasonal cycles, and especially the contribution of forbs, is needed.
Nitrogen is usually the first limiting nutrient in the diet of cattle grazing senesced grass pastures in the seasonally dry tropics. Numerous pen studies with cattle fed low-quality grass hays have reported large increases in forage intake and reduced liveweight (LW) loss, or occasionally increased LW gain, in response to N supplements, including to N provided as non-protein N (NPN; Hennessy and Williamson 1990; Kennedy et al. 1992; Dixon and Mayer 2021). The CP and the CP:ME) in senesced tropical grass pastures, and in the diet of cattle grazing such pastures, are usually low (Winks 1984; Minson 1990; Hogan 1996). Non-protein N supplements have reduced the dry season LW loss by up to c. 35 kg in growing cattle and breeder cows grazing senesced pastures in the higher rainfall (>600 mm) northern speargrass and Aristida–Bothriochloa pasture regions of northern Australia (Winks et al. 1979; Holroyd et al. 1988; Dixon 1998; Coates and Dixon 2008a; Dixon and Coates 2010; Dixon et al. 2011). Also sheep fed Mitchell grass hays in pens (Stephenson and Hopkins 1978; Stephenson et al. 1983), or grazing senesced Mitchell grass pastures (Stephenson et al. 1981; McMeniman et al. 1982), have often responded to N supplements. However, there is no information based on replicated research on the responses of cattle grazing Mitchell grass pastures to N supplements.
The present study was conducted to provide quantitative information on the diet and the growth of steers, and their responses to N supplements, when grazing Mitchell grass pastures in north-western Queensland in northern Australia. Livestock production on many of these rangelands has changed in recent decades from primarily Merino wool production to beef cattle. Hence, a better understanding of the nutrition and production of cattle is needed, particularly with respect to the differences in diet selection and in the nutritional demands for cattle growth rather than wool. The study also examined the extent and timing of responses of growing steers to N supplements, and the extent to which these responses could be predicted from diet CP and diet CP:ME.
Materials and methods
Experimental site and cattle
The study was conducted on Toorak Research Station, 50 km south of Julia Creek, north-western Queensland (21°2′S, 141°48′E). The site was representative of Astrebla spp. pastures in the northern more summer-dominant rainfall pattern of this pasture system in western Queensland, Australia. The treeless pasture was dominated by Mitchell grasses (predominantly Astrebla lappacea (Lindl.) Domin) and Flinders grasses (Iseilema spp.), and a diverse range of leguminous and non-leguminous herbaceous forbs on grey, cracking, self-mulching clay soils. Long-term (1913–2000) mean annual rainfall was 471 mm, with 84% (394 mm) occurring during summer (November–March), and the median date of the seasonal break (>50 mm rain within a 3-day interval; Rainman 4.3 database, 2000) was 10 January (day of year from 1 January in the preceding year (DOY) 375; Table 1). During the 5 years (1996/1997–2000/2001) before the study commenced, annual rainfall ranged from 76 to 134% of the long-term mean, and the November–March rainfall was 80–90% of the annual rainfall. Because the experimental paddocks had been moderately grazed during these years, the pastures were in good condition.

|
The experimental cattle comprised four annual drafts of Droughtmaster type steers (c. 5/8 Bos indicus × 3/8 Bos taurus cross) from the Swans Lagoon Research Station herd near Ayr, northern Queensland, in the northern speargrass region. Steers were relocated to the research site 2–4 months before introduction to the experiments to allow adaptation.
Experimental design and treatments
The study was designed as a randomised block to examine three nutritional treatments, each with two paddock replicates for three annual cycles. The six 128 ha paddocks were, on the basis of soil type and topography, considered as two blocks, and the three paddocks within each block were allocated to one of three treatments during each annual cycle. It was planned to graze two age groups of steers in each paddock, with introduction of a new draft of the young steers and disposal of the older age-group steers at the end of each RS. The number of steers per paddock in each new draft was calculated at the end of the RS based on pasture on offer, with assumptions that steers consumed 20 g pasture DM/kg liveweight (LW) per day, and that 600 kg/ha of residual pasture was required 10 months after assessment. The study commenced in August 2001. Rainfall below the long-term average during the three RS in the 2001/2002, 2002/2003 and 2003/2004 summers (Table 1) led to moderate to low pasture production. Thus, a reduced number of steers only of the young age group were introduced in the replacement drafts commencing at the end of the RS in Experiments 2, 3 and 4. Furthermore, from June 2003 to March 2004 during Experiment 3, the experimental paddocks were destocked because of very low pasture availability. Given the differences among annual cycles in rainfall and numbers of cattle, and the need to destock during 8 months of one annual cycle, the study was regarded as a series of separate experiments (Experiments 1–4).
The commencement and termination dates, numbers of steers, and their initial LW and body condition scores for each experiment are given in Table A1. The experimental paddocks were not grazed during the RS, rainy-to-dry transition season (TS) and early dry season (EDS) before Experiment 1 commenced on 3 August 2001 and continued until 23 March 2002. Cattle in each paddock comprised 12 steers initially c. 1.6 years old (YO) and six steers initially c. 2.6 YO. In subsequent years, only younger steers initially c. 1.2 YO were used. Experiment 2 commenced on 23 March 2002 with 12 steers and continued until 1 April 2003. Experiment 3 commenced on 2 April 2003 with six steers per paddock, but with only 200 mm summer rainfall in 2002/2003 and no further rain, the paddocks were destocked on 24 June 2003. Then, until 22 March 2004, steers given the T-NIL treatment and five additional similar steers that had also not been fed supplements (n = 17 in total) grazed an adjacent 256 ha paddock with similar soil and pasture species, but a higher pasture availability. Experiment 4 commenced on 23 March 2004, with 18 steers per paddock, and continued until 11 April 2005. The three paddocks within each block were initially randomly allocated to treatments, and each treatment was imposed in each paddock during Experiments 1, 2 and 4.
The three treatments consisted of no supplement (T-NIL), or supplements of loose mineral mix, to provide principally NPN offered ad libitum (T-NPN), or a restricted amount of cottonseed meal (T-CSM). Supplements were fed through the TS, EDS and late dry (LDS) seasons until the seasonal break. The T-NPN supplement comprised (g/kg air-dry) 350 cottonseed meal, 300 urea, 290 salt (sodium chloride) and 60 ammonium sulfate, and contained 165 g N/kg DM with 87 of the N as NPN. This supplement contained (g/kg DM) 111 sodium, 14.5 sulfur, 4.0 phosphorus and 1.6 calcium. The NPN supplement was offered twice weekly, and the voluntary intake was measured from the amounts of supplement DM offered and refused. The DM contents were determined by drying at 60°C to constant weight. The CSM supplement was offered twice weekly at intervals of 3 and 4 days in open troughs (>0.25 m trough space per steer) in amounts equivalent to 400 g air-dry per steer per day (g/steer day), and 28 g N/steer day, and was always consumed promptly. On average this supplement contained 914 g DM/kg as‐fed CSM, and (g/kg DM) 77.2 N, 4.8 sulfur, 1.9 sodium, 12.7 phosphorus and 2.6 calcium, and an estimated 11.5 MJ ME/kg DM. Supplements were fed from shortly after the commencement of each new draft of cattle through to the seasonal break.
Pasture and animal measurements
Pasture yield and species composition were estimated in May in the TS and October in the LDS of each annual cycle using Botanal, with 400–500 quadrat (0.5 × 0.5 m) estimates in a grid pattern per paddock on each occasion (Tothill et al. 1992). Herbaceous forbs were classified as likely palatable or unpalatable to cattle according to Cunningham et al. (1981) and Milson (2000), and as C3 or C4 plants according to Cobon and Carter (1994) and Sage (2016). Each 4–5 weeks, steers were mustered to yards in the late afternoon, fasted overnight with access to water, and weighed the following day. Faecal samples were obtained by rectal sampling when the cattle were weighed, and also from fresh dung pats in each paddock at 2 week intervals between musters in Experiments 1–3, and at weekly intervals in Experiment 4. To ensure that faecal samples from steers in the T-CSM treatment were representative of the diet forage, faeces were sampled on Days 3 or 4 after feeding the CSM supplement when the faeces would have comprised principally undigested residues of the forage ingested and little undigested CSM residue (Dixon and Coates 2005). Faecal samples from individual steers were bulked within paddocks, and in Experiment 1 also within steer age groups, and stored frozen.
The annual cycle was considered as the following four seasonal intervals:
the TS from 1 April until 31 May (DOY 91–151) for 60 days,
the EDS from 1 June until 31 August (DOY 152–243) for 90 days,
the LDS from 1 September (DOY 244) until the seasonal break, and was for 107, 183, 130 and 129 days in Experiments 1–4 respectively,
the RS from the seasonal break until 31 March (DOY 90), and was for 104, 28, 81 and 80 days in Experiments 1–4 respectively.
Laboratory analyses and calculations
Faecal samples were oven dried (65°C) and then ground (1 mm screen, Model 1093 Cyclotec mill, Foss Tecator AB, Hoganas, Sweden). For NIR analyses, samples were re-dried (65°C) and scanned (400–2500 nm range) using a monochromator fitted with a spinning cup module (Foss 6500, NIRSystems, Inc., Silver Spring, MD, USA). Chemometric analysis used ISI software (Infrasoft International, Port Matilda, PA, USA), with global H values calculated as (Mahalanobis distance)2/f), where f is the number of factors in the model (Shenk and Westerhaus 1991). Calibration equations developed for northern Australian pastures (Coates 2004; Coates and Dixon 2008b, 2011; Dixon and Coates 2009; R. M. Dixon, unpubl. data) were used to measure the diet CP concentration, diet DM digestibility (DMD), the proportion of non-grass in the diet and the N concentration of faeces. Because no browse was available in the experimental paddocks, the component measured by near-infrared spectroscopy of faeces (F.NIRS) as non-grass must have comprised forbs, and hence this term is used throughout this paper. The diet ME content (M/D) was calculated as: M/D = (0.172 × DMD%) − 1.707 (CSIRO 2007). The daily LW change (LWC) of the steers between successive measurements of steer weight was calculated as the change since the previous measurement. The voluntary intake of pasture DM was calculated from the ME requirement by steers to achieve the measured LWC on each occasion the steers were weighed, and the contents of ME and forbs in the diets following CSIRO (2007) equations.
Statistical analyses
The effect of steer age and supplement treatments on the diet attributes in Experiment 1 was examined as a 2 × 3 factorial by ANOVA on each of the 3 days within the LDS and RS when faecal samples were obtained separately from the age groups. Similarly, the effects of the supplement treatments on the diet attributes in Experiments 2 and 4 were examined by ANOVA for each of the seasons in each annual cycle on each of the days when faecal samples were obtained. The effects of each of the seasons (RS, TS, EDS and LDS) and year (Experiments 1–4) on diet attributes, pasture intake and steer LWC were analysed in the steers not fed supplements (T-NIL treatments) by a one-way ANOVA, using paddocks as replicates. For all analyses, the paddock treatment groups were considered as independent replicate experimental units. Analyses were performed using GENSTAT (release 18.1, VSN International, Hemel Hempstead, UK). Means were compared when appropriate using Fisher’s protected l.s.d. test. Linear regression was used to examine the changes with time in each of the diet attributes, and of DM intake and LWC, first within seasons and, when appropriate, combined across seasons.
Results
Rainfall and pasture measurements
During the 4 years of the study, the annual rainfall was only 50, 42, 65 and 68% of the long-term rainfall, whereas seasonal breaks ranged from 17 December 2001 to 3 March 2003 (Table 1).
Pasture available varied throughout the study (Table 2). In the TS, forbs comprised only 13–15 g/kg of the pasture in Experiments 1 and 2, but 324 and 173 g/kg in Experiments 3 and 4, respectively. These high forb proportions were associated with a low availability of grasses (296 and 1062 kg/ha in Experiments 3 and 4 respectively), and followed only a moderate pasture availability in May 2002. In Experiments 1 and 2, the most frequent forb species present in quadrat measurements were Rynchosia minima (Rynchosia) and Sida spp. (Table 3). In Experiments 3 and 4, when forbs comprised a higher proportion of the pasture, there was a greater variety in forb species, the most frequent being Boerhavia spp. (Tar vines), Cleome viscose (Tick weed), Ipomoea and Polymeria spp., Sida spp., Streptoglossa adscendens (Mint bush), Tribulus terrestris (Caltrop) and Alysicarpus rugosus (Chain pea).

|

|
Supplement intake
The T-CSM-treatment steers consumed their supplement within a few hours of being fed each 3rd or 4th day, and thus consumed the equivalent of 366 g DM and 28 g N of CSM per day. In Experiment 1, the voluntary intake of the loose mineral mix supplement (T-NPN) averaged 95 g DM/steer.day and 16 g N/steer.day (Fig. 1a) during the LDS; thus the intake of NPN-N was 55% of CSM-N. In Experiments 2 and 4 the NPN supplement intakes increased progressively through the TS, EDS and LDS seasons. During the LDS the T-NPN steers consumed on average 46 and 38 g N/steer.day in Experiments 2 and 4 respectively (Fig. 1b, c); thus, the intake of supplementary N in the T-NPN steers was 164% and 136% that in the T-CSM steers.
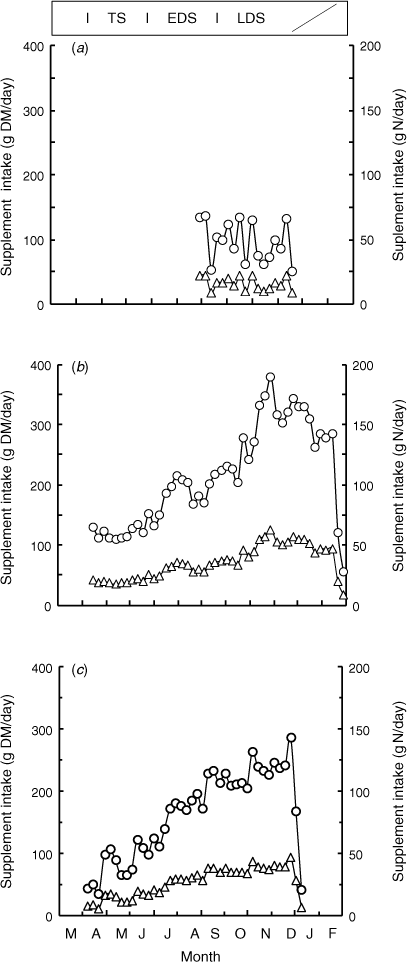
|
Diets selected by steers
During the LDS in Experiment 1, steer age had no significant (P > 0.05) effect on any of the diet attributes (Table A2). In contrast, during the RS when no supplements were fed, the diet CP, DMD and CP:ME, and the faecal N concentration were all higher (P < 0.05 or P < 0.01) in younger steers, indicating that the older steers selected diets of lower nutritional quality. However, despite these statistical significances, the differences were likely to be too small to have had any important nutritional consequences on these high-quality RS diets.
There were effects of supplementation on the F.NIRS-measured diet attributes during the LDS of Experiment 1. Averaged across both age groups, both supplements increased (P < 0.05 or P < 0.01) diet CP (from 42.7 to 49.8 and 55.0 g/kg), and diet CP:ME (from 5.8 to 6.7 and 7.5 g CP/MJ ME), for the T-NIL, T-NPN and T-CSM diets respectively (Table A2). The diet DMD and forbs contents were not significantly affected (P > 0.05). There were no significant (P > 0.05) effects of the supplementation on the diet attributes measured with F.NIRS in Experiments 2 and 4. Nevertheless, given the differences in Experiment 1, the F.NIRS measurements made in the T-NIL-treatment steers were used throughout as the most reliable estimates of the attributes of the diets selected by the steers in all three treatments.
During each annual cycle, the values for the diet attributes of the T-NIL steers in each experiment followed the profile in Experiment 4 (Fig. 2). The diet CP and DMD increased abruptly to maxima 2–3 weeks after the seasonal break, and then progressively declined. Across all experiments, the diet CP averaged 114 (range 91–152) g/kg, DMD 603 (range 575–630) g/kg and diet CP:ME 13.0 (range 11.1–16.6) g/MJ during the RS. These diet attributes were always high during the RS and then declined through the following seasons (Table 4). During the RS, these diet attributes decreased linearly within the paddock groups (P < 0.05 or P < 0.01) in Experiments 3 and 4 (18–21 g/kg.month for CP, 23–32 g/kg.month for DMD and 1.3–1.9 g/MJ.month for CP/ME). However, no such relationships (P > 0.10) were observed during the RS in Experiment 1 where there was high variability, or in Experiment 2 where the RS to the end of March was very brief, owing to the late seasonal break. The changes in diet quality from the RS to the TS were confounded by the introduction of the new draft of steers at the beginning of each TS. Nevertheless, there were consistent and often significant (P < 0.05) decreases in the CP, DMD and CP:ME from the RS to the TS, and, from the TS through the EDS. There were generally smaller differences in these diet attributes, or no differences, between the EDS and the LDS. During the LDS, the diet CP averaged 55 (range 48–62) g/kg, DMD 539 (range 511–574) g/kg, and diet CP:ME 7.3 (range 6.3–8.7) g CP/MJ ME across all experiments. These diet attributes were generally consistent through the LDS, except in Experiment 1 where the diet CP and DMD from 27 September to 31 October 2001 (44.7 and 527 g/kg, n = 3) appeared to increase from 19 November to 17 December (53.0 g CP/kg and 550 g/kg DMD, n = 3) in association with rain between 28 October and 26 November that was not sufficient to constitute a seasonal break but led to some pasture ‘green pick’.
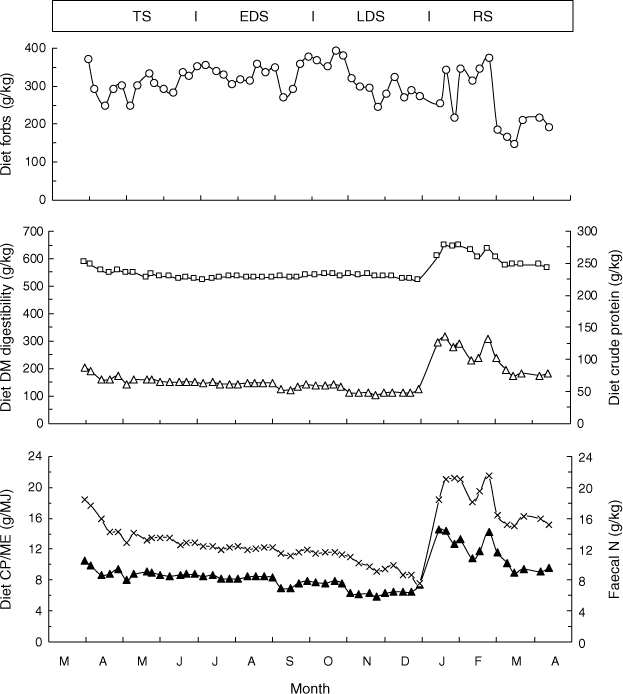
|

|
There was strong selection by the steers for forbs in Experiments 1 and 2 where forbs comprised only 13 and 15 g/kg of the pasture in May 2001 and 2002 respectively (Table 4). In the LDS of Experiment 1, forbs in the diet averaged 146 g/kg (Table 3), ranging from 258 g/kg in September 2001 to 94 g/kg in late November 2001; this decrease was presumably due to removal of forbs by grazing during the interim. Similarly, during the TS in Experiment 2, forbs comprised only 15 g/kg of the pasture, but averaged 236 g/kg in the diet during the TS, decreasing to 118 g/kg in the LDS (Table 4). In contrast, in Experiments 3 and 4, there were much higher proportions of forbs in the pasture in May during the TS (324 and 173 g/kg), and the average diet forbs through the TS, EDS and LDS seasons ranged from 300 to 396 g/kg; thus, in these latter experiments, there was little selection of the forbs by the steers. Diet forb content was high during each of the RS (238–447 g/kg in Experiments 1–4). In all the seasons, there was a high variability in forb content within experiments, with the coefficient of variation ranging from 9 to 77%.
The estimated voluntary pasture intake calculated from steer LWC and diet ME (Table 4) averaged 36.9 (range 35.6–39.2) g DM/kg LW during the RS (excluding Experiment 2), and 21.0 (range 19.5–24.0) g DM/kg LW during the LDS (Table 4). However, the estimated DMI was much lower than were these mean values when steers were rapidly losing LW towards the end of the LDS.
Liveweight changes of steers not fed supplements (T-NIL treatment)
Steer LW through the annual cycles are shown in Fig. 3, and the average LWC in Table 4. There tended (P = 0.067) to be differences among seasons in the average LWC. During the RS, steers always gained LW rapidly (0.78–1.10 kg/day for 4–13 weeks). Liveweight gains during the TS were 0.55 and 0.34 kg/day in Experiments 2 and 4, but were much higher in Experiment 3 (1.09 kg/day) in association with the preceding late seasonal break. Across all experiments, steers continued to gain LW, but more slowly during the EDS (0.16–0.66 kg/day). During the LDS, the T-NIL-treatment steers in Experiment 1 were losing LW from September. However, in Experiments 2, 3 and 4, steers did not begin to lose LW until late in the LDS (Fig. 3). The LW maxima in Experiments 2, 3 and 4 during the dry season were on 7 January 2003, 28 October 2003 and 16 November 2004, before the steers lost LW for 8–12 weeks through to the seasonal break. Calculated over the entire LDS, steers were at about LW maintenance in Experiments 1, 2 and 4 and gained LW slowly in Experiment 3 (0.21 kg/day; Table 4). Annual LW gains of steers were 121, 220 and 172 kg during Experiments 2, 3 and 4 (Table A1).
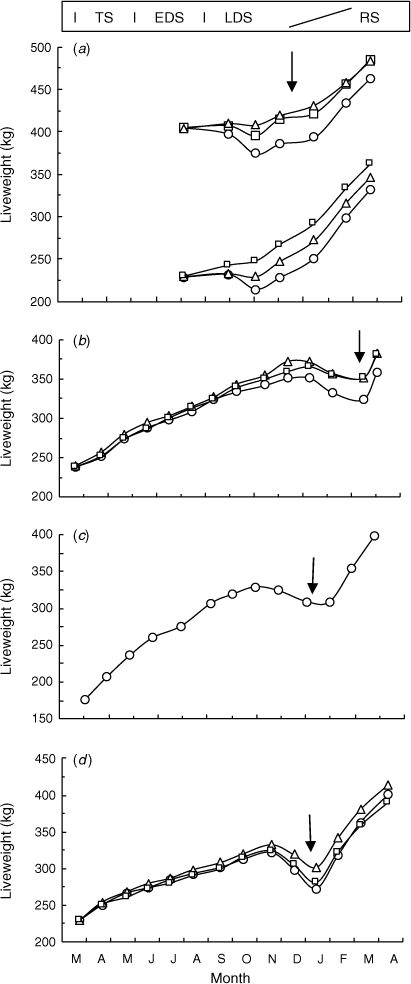
|
The effects of the steer maturity on LWC could be examined only in Experiment 1. At the commencement of the LDS, older steers were heavier (405 versus 229 kg), and through to the seasonal break lost 19 kg LW, compared with negligible LW loss in younger steers (Fig. 3). The LW gain during the late LDS in Experiment 1 was apparently due to rain being sufficient to increase diet quality as described above. The older steers gained less LW from the seasonal break through to the end of the RS (70 versus 81 kg), so that during the 8 months of the LDS and the RS, the older steers gained only 59 kg, versus 103 kg for younger steers.
Liveweight responses of the steers to the N supplements
In Experiment 1, both supplements tended (P = 0.061) to reduce the LW loss in younger steers, with the T-NPN and T-CSM treatments 15 and 32 kg heavier respectively, shortly before the seasonal break, and still 14 and 30 kg heavier at the end of the RS (Fig. 3a). The supplemented steers were heavier (P < 0.05 or P < 0.01) than the T-NIL steers at each LW measurement from late October 2001 through to the end of the RS. The effects of the supplement were less marked in older steers, and the measured differences in the LW between the T-NIL and supplemented steers were comparable in magnitude but were not significantly (P > 0.05) different. In Experiment 2, both the T-NPN- and T-CSM-treatment steers were 22–24 kg heavier at the seasonal break on the 3 March 2003 (P < 0.05) (Fig. 3), and these LW increases were retained through to the end of the nominal RS only 4 weeks later (P < 0.05). In Experiment 4, similar trends were observed for the T-NIL and T-NPN treatments, but there were no significant (P > 0.05) differences. In summary, N supplements increased steer LW only in the LDS, from September in Experiment 1, and from late October or November in Experiments 2 and 4. The measured differences in LW of young steers fed the NPN supplement were 20, 24 and 29 kg in Experiments 1, 2 and 4 respectively, at the seasonal break.
Discussion
Diet quality and cattle growth though the annual cycles
Despite low annual rainfalls and only moderate pasture availability, with appropriate stocking rates the steers were able to select diets that provided sufficient ME and CP to support LW gains during the RS that approached or exceeded 1 kg LW/day for 11–15 weeks. These LW gains on these Mitchell grass pastures are high compared with those often observed in cattle grazing RS pastures in the seasonally dry tropics of northern Australia. In a review of 140 experiments, LW gain during the ‘peak’ period of growth in cattle grazing grass pastures exceeded 1 kg/day in only 18 of 160 measurements (Winter et al. 1991).
Although diet quality declined as expected following the RS, it was still sufficient through the TS and EDS of Experiments 2 and 4 to provide for moderate LW gains (ranging from 0.16 to 0.55 kg/day) from April to August. Higher steer LW gains occurred during the TS and EDS of Experiment 3 (1.09 and 0.66 kg/day respectively). This was presumably due to the late seasonal break (3 March 2003) in this experiment, leading to a late commencement of pasture growth and pasture in less mature stages and higher nutritive value during the TS and EDS. The continuation of slow steer LW gain for c. 18, 8 and 11 weeks after the commencement of the LDS in Experiments 2, 3 and 4 respectively, followed from an average diet DMD of 530–582 g/kg, diet CP ≥ 59.5 g/kg DM, and CP:ME ≥ 7.2 g CP/MJ ME, and LW gains, during the EDS. The annual LW gains of 121–220 kg in the present study are in accord with the few research reports available, indicating that in reasonable to good seasonal conditions, the annual LW gain of young cattle grazing Mitchell grass pastures is likely to be in the range of 110–190 kg per annum (Smith and Cheffins 1986; McLennan et al. 1999; Bortolussi et al. 2005). These LW gains are also in accord with the expectations of experienced cattle managers in the region, and with observations that at very low stocking rates, cattle can often maintain or gain LW during low-rainfall years. The diet quality and LW gains of steers in the present study are in contrast to those reported for many other pasture systems in northern Australia where the diet quality during the TS, EDS and LDS was lower, and steers are usually losing LW by August (Winks 1984; McLennan et al. 1988; Dixon et al. 2007b, 2011; Dixon 2008; Coates and Dixon 2008a; Dixon and Coates 2010).
That the quality of the diet selected by the steers declined only slowly through the TS and dry seasons was most likely because steers were able to maintain substantial intakes of forbs of higher nutritive value than that of the senesced grasses (Table 4). Numerous studies have reported that the forbs present in Mitchell grass pastures are usually much higher in CP and DMD than are the grasses (Weston and Moir 1969; Hall and Lee 1980; Holm and Eliot 1980; Roe and Allen 1993). Sheep strongly select for many of the forb species, and forbs are important to their diet quality and productivity (Lorimer 1978, 1981; Pritchard et al. 1986; McMeniman et al. 1986a, 1986b; Orr et al. 1988). The present study is in accord with the few other reports available of substantial selection by cattle grazing Mitchell grass pastures of non-grass species during the dry season at other sites, presumably including forbs when present (Coates and Dixon 2007; Dixon et al. 2007a; Cowley et al. 2018). Non-grass species (forbs and browse) are present in many other seasonally dry native pasture rangelands both in northern Australia (Siebert et al. 1968; Newman 1969; Hall 1981; Squires and Siebert 1983) and elsewhere such as tropical western Africa (Innes and Mabey 1964; Fianu 1982), and often make an important contribution to dry-season diet quality. However, these non-grass forages typically comprise only 50–150 g/kg of the diet during the dry season in the northern and southern speargrass, Aristida–Bothriochloa and semi-arid pasture systems (Squires and Siebert 1983; Coates and Dixon 2007; Dixon et al. 2007b, 2011; Hall et al. 2016). Moreover, because many browse species contain antinutritional factors (e.g. condensed tannins) that severely reduce the availability of the N and the digestibility of browse for ruminants, their contribution to diet CP and diet DMD will usually be much lower than is the contribution of the forbs such as in the present study. There are other additional characteristics of Mitchell grass pastures in semi-arid environments that may contribute to the diet quality of both sheep and cattle through the dry season. The generally fertile soils and growth of only moderate pasture biomass, particularly during low-rainfall years, would likely be associated with a lesser dilution of soil N and other plant nutrients during grass growth than in higher-rainfall regions. This results in senesced grasses having a higher CP than in pastures growing on infertile soils. In addition, during the present studies, the CP and DMD of the senesced grasses were presumably maintained with an absence of dry season spoiling rainfall.
Response of the steers to the N supplements
During the dry season, mature tropical grass pastures are often deficient in N relative to ME, and N supplements providing rumen-degradable N are often effective in increasing rumen microbial digestion, voluntary intakes of ME, and animal LW in rangelands such as the northern speargrass and Aristida–Bothriochloa pasture regions of northern Australia (Winks 1984; Holroyd et al. 1988; Dixon 1998; Coates and Dixon 2008a; Dixon and Coates 2010; Dixon et al. 2011). In these latter pasture systems, the responses to NPN supplement often commenced during the TS or EDS, and continued through to the seasonal break. This contrasts with the present study where LW loss and a response to N supplements did not occur until the LDS. The F.NIRS measurements showed that T-NIL steers were selecting diets with sufficient ME content to provide for slow LW gain, and diets adequate in CP and CP:ME. As discussed above, this was likely to be associated primarily with forbs in the diet, but possibly also with higher N and ME in these senesced Mitchell grasses than what occurs in senesced grasses in most other native pasture systems in northern Australia. The N supplements reduced LW loss, and the supplemented steers were about 20–30 kg heavier shortly before the seasonal break. The magnitude of this reduction in dry-season LW loss is in accord with reported studies of NPN supplementation of cattle grazing senesced dry-season speargrass, Aristida–Bothriochloa and Cenchrus ciliaris pastures in northern Australia in the studies cited above.
The responses to NPN supplements in cattle consuming low-quality forages are due to provision of rumen-degradable N as a substrate for rumen microbial digestion. Because the requirement for rumen degradable protein is directly related to the fermentable ME available for microbial activity, the ratio of dietary CP to fermentable ME (for forage diets simply diet CP:ME) should provide a reliable measure of the adequacy of the supply of rumen-degradable N. In Experiments 2 and 4, during the EDS, and during the LDS until c. 4 weeks before the steers achieved the maximum dry-season LW, the CP:ME ratios were ≥7.0 g CP/MJ ME (paddock means ranged from 7.0 to 7.9). However, during the 4 weeks before the maximum dry-season LW, and when the LW responses to the NPN supplement became apparent (Fig. 3), the CP:ME averaged 6.6, 6.7 and 6.3 in Experiments 1, 2 and 4 respectively. This suggests that when cattle are grazing Mitchell grass pastures with some dietary forbs, the supply of rumen-degradable N for rumen digestion becomes limiting when the g CP/MJ ME is about <7.0. This CP:ME ratio is equivalent to a DMD:CP ratio of >10 when Bos indicus cattle grazing other tropical grass pastures have responded to NPN supplements (Dixon and Coates 2005, 2010; Coates and Dixon 2008).
For several reasons, the CP:ME threshold when cattle respond to NPN supplements is likely to vary with the diet and the pasture system. A major uncertainty will be the proportion of forage N present as rumen-degradable N and this may be low in senesced C4 grasses (Hogan et al. 1989; Hogan 1996). There is a lack of information on the availability of N in forbs at various stages of growth and maturity. In addition, the diet CP:ME threshold will be affected by the transfer of endogenous urea N to the rumen that also potentially provides rumen-degradable N. For comparison, the ratio of microbial CP to digestible organic matter (or ME is often used as a measure of the efficiency of rumen microbial protein synthesis and the supply of microbial protein to the animal, but the CP:ME ratios required for optimal microbial protein synthesis (about 8.3 g microbial CP/MJ ME at animal maintenance; CSIRO 2007) and the thresholds required for responses to NPN supplements or increased rumen digestion may well differ. Where F.NIRS can be used, the diet CP:ME should provide the best estimate of when cattle are likely to respond to N supplements. The results here suggest that N supplementation responses will not occur until October–November in the LDS in Mitchell grass pastures where cattle are able to select forbs. This contrasts with pastures such as the northern speargrass and Aristida–Bothriochloa regions, where diet CP and CP:ME are often much lower during EDS, and sometimes the TS, and when N supplementation responses often occur (Winks 1984; Holroyd et al. 1988; Dixon 1998, 2008; Coates and Dixon 2008a; Dixon and Coates 2010; Dixon et al. 2011).
Supplementary N was provided as either NPN or as cottonseed meal to compare the responses to the two forms on N. Greater responses to protein meal supplements occur in some circumstances because of an increased supply of absorbed amino acids from protein meal escaping rumen digestion, as well as the increased supply and form of rumen-degradable N (Dixon and Egan 2000; McLennan et al. 2016). However, in the present experiments there was no evidence that the CSM supplement led to higher pasture intakes or LW than did the NPN supplement. However, during the LDS when steer LW responses occurred the intake of supplementary N by steers in Experiments 2 and 4 was substantially greater in steers fed the NPN supplement than in those fed the CSM supplement, possibly because of the high voluntary intake of the NPN supplement. Also, the NPN supplement contained 350 g CSM/kg. The responses to N supplementation could be attributed entirely to increased supply of rumen-degradable N as ammonia.
The reliability of the F.NIRS measurements of diet quality and diet attributes
The measured changes in diet quality through the annual cycles and the conclusion that the steers responded to N supplements when the diet contained less than about 7.0 g CP/MJ ME obviously depend on the accuracy and precision of the F.NIRS measurements. The robustness and expected errors in the F.NIRS measurement of diet CP and DMD using the northern Australian calibration equations have been described comprehensively (Coates 2004; Dixon and Coates 2009; Coates and Dixon 2011). Because only a small proportion of the reference samples used to construct these calibrations were from Mitchell grass pastures, the appropriateness of the calibrations to measure attributes of Mitchell grass pasture diets was evaluated using two approaches. First, because the northern Australian calibration sample sets (n = 1223 for diet CP and n = 1052 for diet DMD) included some Astrebla spp. forages (n = 39; mean 37.8 g CP/kg and 478 g/kg DMD), the cross-validation prediction errors for these forages could be examined (Table 5). Second, data not included in the calibration equations were available from other pen experiments with cattle fed Astrebla or Iseilema spp. forages in pens (n = 31) at the Toorak Research Station and elsewhere, and could be examined as an external validation sample set. In both sets of samples, the diet CP and DMD were measured satisfactorily, with errors being comparable to those for other common pasture grasses in the northern Australian seasonally dry tropics. There was no evidence of bias error causing the CP or DMD measurements of Astrebla spp. pasture diets to be consistently over- or underestimated (Table 5). Furthermore, these Astrebla or Iseilema spp. forages represented low-quality senesced pastures that are the most relevant for estimating the diet CP:ME at which cattle will respond to NPN supplements.
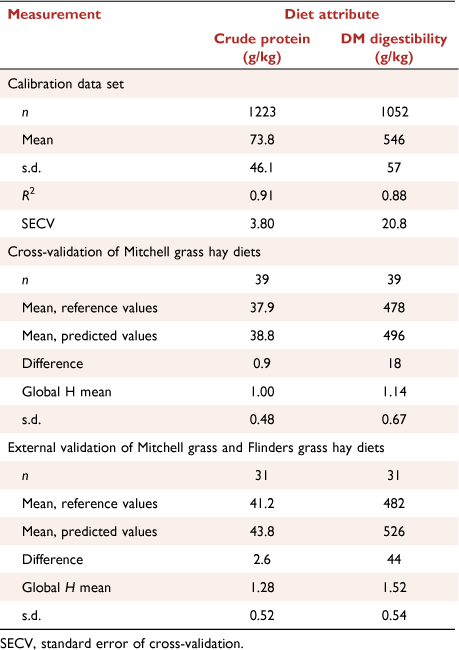
|
The observation that diet CP as measured with F.NIRS was increased by NPN supplementation in Experiment 1 (Table A2), but not in Experiments 2 or 4, was in accord with previous reports that mineral-mix supplements containing urea have only small and inconsistent effects on F.NIRS measurements of the diet CP or DMD (Coates and Dixon 2008a; Dixon and Coates 2005, 2009, 2010). The conservative approach adopted in the present experiments to use the F.NIRS results from the non-supplemented (T-NIL) steers as measurements of the diet forage, and to calculate the total N concentration of the diet from the N concentrations and intakes of both the forage and the supplement (Dixon and Coates 2005), would have avoided error from this source. These calculations indicated that the NPN supplement would have increased the CP concentration of the entire diet by 30–35 g CP/kg DM during the EDS in Experiments 2 and 4, respectively, and by c. 20, 40 and 50 g CP/kg DM during the LDS in Experiments 1, 2 and 4 respectively.
An unusual characteristic of Mitchell grass pastures is that some common forb species are C4 rather than C3 plants (Cobon and Carter 1994; Sage 2016), and not all are palatable (Table 3). However, the F.NIRS calibration equations used to measure the proportion of forbs in the diet depend on the grasses being C4 and forbs being C3 plants, and using the 12C:13C ratio in faeces as reference values (Coates and Dixon 2008b). This has two implications for measurement of the proportion of herbaceous forbs in the diet. First, because of the potential inclusion of C4 forbs in the diet, it was not possible to use the 12C:13C in faeces measured by mass spectroscopy as reference values to validate the F.NIRS measurements of forbs for these Mitchell grass pastures, as is possible for other pasture systems (Dixon and Coates 2008; Dixon et al. 2020). Second, as discussed by Coates and Dixon (2008b), F.NIRS calibrations actually depend on the differences in anatomical structure of C3 and C4 plants rather than differences in the 12C:13C ratio. The presence of some C4 forbs in the diet would have caused the F.NIRS calibration to underestimate the proportion of dietary forbs, but likely by no more than 10% of the predicted proportion. Thus, the presence of some C4 forbs in the diet should not have affected the conclusion that forbs often contributed 20–40% of the diet in Experiments 3 and 4, and the steers strongly selected for forbs in Experiments 1 and 2.
Conclusions
This series of experiments provided accurate and detailed information on the quality of the diet selected and the LW change of tropically adapted cattle grazing Mitchell grass pastures in north-western Queensland through a series of abnormally low rainfall years. The steers strongly selected for forbs, even when they comprised only a minor proportion of the pasture, such that forbs comprised a substantial part of the diet. Diet quality and growth of the steers was highest during the RS, but then decreased only slowly. Diet quality was sufficient for slow steer LW gain through to late in the dry season. Nitrogen supplements increased steer LW near the end of the dry season when the diet CP:ME decreased to c. 7.0 g CP/MJ, and increased steer LW at the end of the dry season and at the end of the subsequent RS.
Data availability
The data that support this study cannot be publicly shared due to ethical and privacy reasons, but may be shared upon reasonable request to the corresponding author if appropriate.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
We wish to acknowledge Meat and Livestock Australia for their partial financial support.
Acknowledgements
The experimentation was conducted under the (then) Department of Primary Industries of the Queensland Government with Animal Ethics Approval number WLAEC 027-01. We thank staff of the Toorak Research Station and of the Animal Research Institute for their technical assistance, Dr David Orr and Dr David Phelps for their assistance with the pasture evaluation measurements and comments on the manuscript, and Mr David Coates for for the F.NIRS laboratory analyses and numerous discussions through the experimentation and the development of the present paper.
References
Blake, ST (1938). The plant communities of Western Queensland and their relationships, with special reference to the grazing industry. Proceedings of the Royal Society of Queensland 49, 156–204.Bortolussi, G, McIvor, JG, Hodgkinson, JJ, Coffey, SG, and Holmes, CR (2005). The northern Australian beef industry, a snapshot. 3. Annual liveweight gains from pasture based systems. Australian Journal of Experimental Agriculture 45, 1093–1108.
| The northern Australian beef industry, a snapshot. 3. Annual liveweight gains from pasture based systems.Crossref | GoogleScholarGoogle Scholar |
Christian CS, Noakes LC, Perry RA, Slatyer RO, Stewart GA, Traves DM (1952) ‘Survey of the Barkly region, Northern Territory and Queensland, 1947-48. Land Series 3.’ (CSIRO: Melbourne, Vic., Australia)
Coates DB (2004) Faecal NIRS – Technology for improving nutritional management of grazing cattle. Final Report of Project NAP3.121. Meat and Livestock Australia, Sydney, NSW, Australia.
Coates, DB, and Dixon, RM (2007). Faecal near infrared reflectance spectroscopy (F.NIRS) measurements of non-grass proportions in the diet of cattle grazing tropical rangelands. The Rangeland Journal 29, 51–63.
| Faecal near infrared reflectance spectroscopy (F.NIRS) measurements of non-grass proportions in the diet of cattle grazing tropical rangelands.Crossref | GoogleScholarGoogle Scholar |
Coates, DB, and Dixon, RM (2008a). Faecal near infrared reflectance spectroscopy measurements of diet quality and responses to N supplements by cattle grazing Bothriochloa pertusa pastures. Australian Journal of Experimental Agriculture 48, 829–834.
| Faecal near infrared reflectance spectroscopy measurements of diet quality and responses to N supplements by cattle grazing Bothriochloa pertusa pastures.Crossref | GoogleScholarGoogle Scholar |
Coates, DB, and Dixon, RM (2008b). Development of NIRS analysis of faeces to estimate non-grass proportions in the diets selected by cattle grazing tropical pastures. Journal of Near Infrared Spectroscopy 16, 471–480.
| Development of NIRS analysis of faeces to estimate non-grass proportions in the diets selected by cattle grazing tropical pastures.Crossref | GoogleScholarGoogle Scholar |
Coates, DB, and Dixon, RM (2011). Developing robust faecal NIRS calibrations to predict diet dry matter digestibility in cattle consuming tropical forages. Journal of Near Infrared Spectroscopy 19, 507–519.
| Developing robust faecal NIRS calibrations to predict diet dry matter digestibility in cattle consuming tropical forages.Crossref | GoogleScholarGoogle Scholar |
Cobon, DH, and Carter, JO (1994). Effect of herbage in Astrebla spp. pastures in north-west Queensland on lamb survival. Journal of Agricultural Science, Cambridge 122, 291–298.
| Effect of herbage in Astrebla spp. pastures in north-west Queensland on lamb survival.Crossref | GoogleScholarGoogle Scholar |
Cowley R, Scott K, Kraft P, Hearnden M (2018) Newcastle Waters grazing trial: Comparing cell grazing systems with traditional practice in the north-western Barkly region. Northern Territory Government. Technical Bulletin number 357. Darwin, NT, Australia.
CSIRO (2007) ‘Nutrient requirements of domesticated ruminants’, (CSIRO Publishing: Melbourne, Vic., Australia)
Cunningham GM, Mulham WE, Milthorpe PL, Leigh JH (1981) ‘Plants of Western New South Wales’, (Soil Conservation Service of New South Wales: Sydney, NSW, Australia)
Davies, JG, Scott, AE, and Kennedy, JF (1938). The yield and composition of Mitchell grass pasture for a period of twelve months. Journal of the CSIR 11, 127–139.
Dixon RM (1998) Improving cost-effectiveness of supplementation systems for breeder herds in northern Australia. Final Report DAQ.098. Meat and Livestock Australia, Sydney, NSW, Australia.
Dixon RM (2008) Utilizing faecal NIRS measurements to improve prediction of grower and breeder cattle performance and supplement management. Final Report of Project NBP.302. Meat and Livestock Australia, Sydney, NSW, Australia.
Dixon, RM, and Coates, DB (2005). The use of faecal NIRS to improve nutritional management of cattle in northern Australia. Recent Advances in Animal Nutrition in Australia 15, 65–75.
Dixon, RM, and Coates, DB (2008). Diet quality and liveweight gain of steers grazing Leucaena-grass pasture estimated with near infrared reflectance spectroscopy. Australian Journal of Experimental Agriculture 48, 835–842.
| Diet quality and liveweight gain of steers grazing Leucaena-grass pasture estimated with near infrared reflectance spectroscopy.Crossref | GoogleScholarGoogle Scholar |
Dixon, RM, and Coates, DB (2009). NIR spectroscopy of faeces to evaluate the nutrition and physiology of herbivores: a review. Journal of Near Infrared Spectroscopy 17, 1–31.
| NIR spectroscopy of faeces to evaluate the nutrition and physiology of herbivores: a review.Crossref | GoogleScholarGoogle Scholar |
Dixon, RM, and Coates, DB (2010). Diet quality estimated with near infrared reflectance spectroscopy (F.NIRS) and responses to N supplementation by cattle grazing buffel grass pastures. Animal Feed Science and Technology 158, 115–125.
| Diet quality estimated with near infrared reflectance spectroscopy (F.NIRS) and responses to N supplementation by cattle grazing buffel grass pastures.Crossref | GoogleScholarGoogle Scholar |
Dixon, R M, and Egan, AR (2000). Response of lambs fed low quality roughage to supplements based on urea, cereal grain, or protein meals. Australian Journal of Agricultural Research 51, 811–821.
| Response of lambs fed low quality roughage to supplements based on urea, cereal grain, or protein meals.Crossref | GoogleScholarGoogle Scholar |
Dixon, RM, and Mayer, RJ (2021). Effects of a nitrogen-based supplement on intake, liveweight and body energy reserves in breeding Bos indicus cross cows. Tropical Grasslands – Forrajes Tropicales 9, 70–80.
| Effects of a nitrogen-based supplement on intake, liveweight and body energy reserves in breeding Bos indicus cross cows.Crossref | GoogleScholarGoogle Scholar |
Dixon RM, Hamlyn-Hill F, Cribb R, McClymont A, Coates DB (2007a) Faecal NIRS measurements of the diet quality and growth of cattle grazing Mitchell grass–forest country pastures near Richmond, N Qld. In ‘Northern Beef Research Update Conference’, Townsville, Qld, Australia, 21–22 March 2007.
Dixon, RM, Smith, DR, and Coates, DB (2007b). Using faecal NIRS to improve nutritional management of breeders in the seasonally dry tropics. Recent Advances in Animal Nutrition in Australia 16, 123–134.
Dixon, RM, Playford, C, and Coates, DB (2011). Nutrition of beef breeder cows in the dry tropics. 1. Effects of nitrogen supplementation and weaning on breeder performance. Animal Production Science 51, 515–528.
| Nutrition of beef breeder cows in the dry tropics. 1. Effects of nitrogen supplementation and weaning on breeder performance.Crossref | GoogleScholarGoogle Scholar |
Dixon, RM, Shotton, P, and Mayer, R (2020). Diets selected and growth of steers grazing buffel grass (Cenchus ciliaris cv Gayndah)–centro (Centrosema brasilianum cv Oolloo) pastures in a seasonally dry tropical environment. Animal Production Science 60, 1459–1468.
| Diets selected and growth of steers grazing buffel grass (Cenchus ciliaris cv Gayndah)–centro (Centrosema brasilianum cv Oolloo) pastures in a seasonally dry tropical environment.Crossref | GoogleScholarGoogle Scholar |
Fianu FK (1982) The role of forbs and shrubs in the nitrogen intake of grazing livestock in the Accra plains. In ‘Forage protein in Ruminant animal production. Occasional Publication No. 6 – British society of animal production 1982’. (Eds DJ Thomson, DE Beever, RG Gunn) pp. 189–191.
| Crossref |
Graetz, RD, and Wilson, AD (1980). Comparison of the diets of sheep and cattle grazing a semi-arid chenopod shrubland. Australian Rangelands Journal 2, 67–75.
| Comparison of the diets of sheep and cattle grazing a semi-arid chenopod shrubland.Crossref | GoogleScholarGoogle Scholar |
Hall, TJ (1981). The nitrogen and phosphorus concentrations of some pasture species in the Dichanthium–Eulalia grasslands of north-west Queensland. Australian Rangelands Journal 3, 67–73.
| The nitrogen and phosphorus concentrations of some pasture species in the Dichanthium–Eulalia grasslands of north-west Queensland.Crossref | GoogleScholarGoogle Scholar |
Hall, TJ, and Lee, GR (1980). Response of Astrebla spp. grassland to heavy grazing by cattle and light grazing by sheep in north-west Queensland. Australian Rangelands Journal 2, 83–93.
| Response of Astrebla spp. grassland to heavy grazing by cattle and light grazing by sheep in north-west Queensland.Crossref | GoogleScholarGoogle Scholar |
Hall, TJ, McIvor, , Jones, P, Smith, DR, and Mayer, DG (2016). Comparison of stocking methods for beef production in northern Australia: seasonal diet quality and composition. The Rangeland Journal 38, 553–567.
| Comparison of stocking methods for beef production in northern Australia: seasonal diet quality and composition.Crossref | GoogleScholarGoogle Scholar |
Hennessy, DW, and Williamson, PJ (1990). Feed intake and liveweight of cattle on subtropical native pasture hays. 1. The effect of urea. Australian Journal of Agricultural Research 41, 1169–1177.
| Feed intake and liveweight of cattle on subtropical native pasture hays. 1. The effect of urea.Crossref | GoogleScholarGoogle Scholar |
Hogan, JP (1996). Options for manipulation nutrition if feed supply is immutable. Australian Journal of Agricultural Research 47, 289–305.
| Options for manipulation nutrition if feed supply is immutable.Crossref | GoogleScholarGoogle Scholar |
Hogan, JP, Kennedy, PM, McSweeney, CS, and Schlink, AC (1989). Quantitative studies of the digestion of tropical and temperate forages by sheep. Australian Journal of Experimental Agriculture 29, 333–337.
| Quantitative studies of the digestion of tropical and temperate forages by sheep.Crossref | GoogleScholarGoogle Scholar |
Holm, AMcR, and Eliot, GJ (1980). Seasonal changes in the nutritive value of some native pasture species in north-western Australia. The Australian Rangeland Journal 2, 175–182.
| Seasonal changes in the nutritive value of some native pasture species in north-western Australia.Crossref | GoogleScholarGoogle Scholar |
Holroyd, RG, Mason, GWJ, Loxton, ID, Knights, PT, and O’Rourke, PK (1988). Effects of weaning and supplementation on performance of Brahman cross cows and their progeny. Australian Journal of Experimental Agriculture 28, 11–20.
| Effects of weaning and supplementation on performance of Brahman cross cows and their progeny.Crossref | GoogleScholarGoogle Scholar |
Innes, RR, and Mabey, GL (1964). Studies on browse plants in Ghana. III. Browse/grass ingestion ratios. (a) Determination of the free-choice Griffonia/grass ingestion ratio for West African Shorthorn cattle on the Accra plains using the ‘simulated shrub’ technique. Empire Journal of Experimental Agriculture 32, 180–192.
Kennedy, PM, Boniface, AN, Liang, ZJ, Muller, D, and Murray, RM (1992). Intake and digestion in swamp buffaloes and cattle. 2. The comparative response to urea supplements in animals fed tropical grasses. Journal of Agricultural Science, Cambridge 119, 243–254.
| Intake and digestion in swamp buffaloes and cattle. 2. The comparative response to urea supplements in animals fed tropical grasses.Crossref | GoogleScholarGoogle Scholar |
Lorimer, MS (1978). Forage selection studies. 1. The botanical composition of forage selected by sheep grazing Astrebla spp. pasture in north-west Queensland. Tropical Grasslands 12, 97–108.
Lorimer, MS (1981). Forage selection studies. 2. Diet quality, liveweight change and wool production of sheep grazing Astrebla spp. pasture in north-west Queensland. Tropical Grasslands 15, 183–192.
McGowan M, McCosker K, Fordyce G, Smith D, O’Rourke P, Perkins N, Barnes T, Marquart L, Morton J, Newsome T, Menzies D, Burns B, Jephcott S (2014) Northern Australian beef fertility project: CashCow. Final Report B.NBP.0382. Meat and Livestock Australia, Sydney, NSW, Australia.
McLennan SR, Hendricksen RE, Beale IF, Winks L, Miller CP, Quirk MF (1988) The nutritive value of native pastures. In ‘Native Pastures in Queensland. The resources and their management’. (Eds WH Burrows, JC Scanlan, MT Rutherford) pp. 21–33. (Queensland Department of Primary Industries, Q187023: Brisbane, Qld, Australia)
McLennan SR, Dixon RM, Hendricksen R, Dodt R (1999) Nutrition of cattle in western Queensland. Report to the Western Queensland Regional Beef Research Committee, Department of Primary Industries, Qld Government.
McLennan, SR, Bolam, MJ, Kidd, JF, Chandra, KA, and Poppi, DP (2016). Responses to various protein and energy supplements by steers fed low-quality hay. 1. Comparison of response surfaces for young steers. Animal Production Science 57, 473–488.
| Responses to various protein and energy supplements by steers fed low-quality hay. 1. Comparison of response surfaces for young steers.Crossref | GoogleScholarGoogle Scholar |
McMeniman, NP, O’Dempsey, N, Niven, DR, Jordan, D, and O’Brian, J (1982). The effect of cottonseed meal and urea supplementation on the reproductive performance of ewes in central and south west Queensland. Proceedings of the Australian Society of Animal Production 14, 447–450.
McMeniman, NP, Beale, IF, and Murphy, GM (1986a). Nutritional evaluation of south-west Queensland pastures. 1. The botanical and nutrient content of diets selected by sheep grazing Mitchell grass and mulga/grass associations. Australian Journal of Agricultural Research 37, 289–302.
| Nutritional evaluation of south-west Queensland pastures. 1. The botanical and nutrient content of diets selected by sheep grazing Mitchell grass and mulga/grass associations.Crossref | GoogleScholarGoogle Scholar |
McMeniman, NP, Beale, IF, and Murphy, GM (1986b). Nutritional evaluation of south-west Queensland pastures. 2. The intake and digestion of organic matter and nitrogen by sheep grazing Mitchell grass and mulga/grass associations. Australian Journal of Agricultural Research 37, 303–314.
| Nutritional evaluation of south-west Queensland pastures. 2. The intake and digestion of organic matter and nitrogen by sheep grazing Mitchell grass and mulga/grass associations.Crossref | GoogleScholarGoogle Scholar |
McMeniman, NP, Beale, IF, and Payne, G (1989). The nutrient status of sheep grazing on a dry Mitchell grass pasture association on central-western Queensland. Australian Rangeland Journal 11, 7–14.
| The nutrient status of sheep grazing on a dry Mitchell grass pasture association on central-western Queensland.Crossref | GoogleScholarGoogle Scholar |
Milson J (2000) ‘Pasture plants of north west Queensland’, (Department of Primary Industries Queensland: Brisbane, Qld, Australia)
Minson DJ (1990) ‘Forage in ruminant nutrition’, (Academic Press: London, UK)
Newman, DMR (1969). The chemical composition, digestibility and intake of some native pasture species in central Australia during winter. Australian Journal of Experimental Agriculture and Animal Husbandry 9, 599–602.
| The chemical composition, digestibility and intake of some native pasture species in central Australia during winter.Crossref | GoogleScholarGoogle Scholar |
NRBC (2016) ‘Nutrient requirements of beef cattle’, 8 revised edn. (The National Academies Press: Washington DC, USA)
Orr, DM (1975). A review of Astrebla (Mitchell grass) pastures in Australia. Tropical Grasslands 9, 21–36.
Orr, DM, and Phelps, DG (2013a). Impact of level of utilization by grazing on an Astrebla (Mitchell grass) grassland in north-western Queensland between 1984 and 2010. 1. Herbage mass and population dynamics of Astrebla spp. The Rangeland Journal 35, 1–15.
| Impact of level of utilization by grazing on an Astrebla (Mitchell grass) grassland in north-western Queensland between 1984 and 2010. 1. Herbage mass and population dynamics of Astrebla spp.Crossref | GoogleScholarGoogle Scholar |
Orr, DM, and Phelps, DG (2013b). Impact of level of utilization by grazing on an Astrebla (Mitchell grass) grassland in north-western Queensland between 1984 and 2010. 2. Plant species richness and abundance. The Rangeland Journal 35, 1–15.
| Impact of level of utilization by grazing on an Astrebla (Mitchell grass) grassland in north-western Queensland between 1984 and 2010. 2. Plant species richness and abundance.Crossref | GoogleScholarGoogle Scholar |
Orr, DM, Evenson, CJ, Jordan, DJ, Bowly, PS, Lehane, KJ, and Cowan, DC (1988). Sheep productivity in an Astrebla grassland of south-west Queensland. Australian Rangeland Journal 10, 39–47.
| Sheep productivity in an Astrebla grassland of south-west Queensland.Crossref | GoogleScholarGoogle Scholar |
Pritchard, DA, Connelly, PT, Kelly, JG, and Hopkins, PS (1986). Diet selection and wool growth of high and low producing sheep at pasture in north-west Queensland. Proceedings of the Australian Society of Animal Production 16, 307–310.
Roe, R, and Allen, GH (1993). Studies on the Mitchell grass association in South-western Queensland. 3. Pasture and wool production under different rates of stocking and continuous or rotational grazing. The Rangelands Journal 15, 302–319.
| Studies on the Mitchell grass association in South-western Queensland. 3. Pasture and wool production under different rates of stocking and continuous or rotational grazing.Crossref | GoogleScholarGoogle Scholar |
Sage, RF (2016). A portrait of the C4 photosynthetic family on the 50th anniversary of its discovery: species number, evolutionary lineages, and Hall of Fame. Journal of Experimental Botany 67, 4039–4056.
| A portrait of the C4 photosynthetic family on the 50th anniversary of its discovery: species number, evolutionary lineages, and Hall of Fame.Crossref | GoogleScholarGoogle Scholar | 27053721PubMed |
Shenk, JS, and Westerhaus, MO (1991). Population definition, sample selection, and calibration procedures for near infrared reflectance spectroscopy. Crop Science 31, 469–474.
| Population definition, sample selection, and calibration procedures for near infrared reflectance spectroscopy.Crossref | GoogleScholarGoogle Scholar |
Siebert, BD, Newman, DMR, and Nelson, DJ (1968). The chemical composition of some arid zone pasture species. Tropical Grasslands 2, 31–40.
Smith PC, Cheffins RC (1986) ‘Beef production in the Mitchell grass downs’, (Department of Primary Industries, Queensland Government: Qld, Australia)
Squires, VR, and Siebert, BD (1983). Botanical and chemical components of the diet and liveweight change in cattle on semi-desert rangeland in central Australia. Australian Rangeland Journal 1, 28–34.
| Botanical and chemical components of the diet and liveweight change in cattle on semi-desert rangeland in central Australia.Crossref | GoogleScholarGoogle Scholar |
Stephenson, RGA, and Hopkins, PS (1978). Urea supplementation of ewes in the tropics. Proceedings of the Australian Society of Animal Production 12, 178.
Stephenson, RGA, Edwards, JC, and Hopkins, PS (1981). The use of urea to improve milk yields and lamb survival of Merinos in a dry tropical environment. Australian Journal of Agricultural Research 32, 497–509.
| The use of urea to improve milk yields and lamb survival of Merinos in a dry tropical environment.Crossref | GoogleScholarGoogle Scholar |
Stephenson, RGA, Weston, RH, Margan, D E, and Hogan, JP (1983). Intake and digestion of mature Flinders grass (Iseilema spp.) by sheep and their response to the addition of urea and minerals. Australian Journal of Agricultural Research 34, 549–556.
| Intake and digestion of mature Flinders grass (Iseilema spp.) by sheep and their response to the addition of urea and minerals.Crossref | GoogleScholarGoogle Scholar |
Tothill JC, Hargreaves JNC, Jones RM, McDonald CK (1992) ‘BOTANAL – a comprehensive sampling and computing procedure for estimating pasture yield and composition. 1. Field sampling.’ Technical Memorandum No. 78. (Division of Tropical Crops and Pastures, CSIRO: Australia)
Weston EJ (1988) Native pasture communities. In ‘Native Pastures in Queensland. The resources and their management’. QI87023. (Eds WH Burrows, JC Scanlan, MT Rutherford) pp. 21–33. (Queensland Department of Primary Industries: Brisbane, Qld, Australia)
Weston, EJ, and Moir, KW (1969). Grazing preferences of sheep and nutritive value of plant components in a Mitchell grass association in north-western Queensland. Queensland Journal of Agricultural and Animal Sciences 26, 639–650.
Wilson, AD (1976). Comparison of sheep and cattle grazing on a semi-arid grassland. Australian Journal of Agricultural Research 27, 155–162.
| Comparison of sheep and cattle grazing on a semi-arid grassland.Crossref | GoogleScholarGoogle Scholar |
Winks L (1984) ‘Cattle growth in the dry tropics of Australia.’ Review number 45. (Australian Meat Research Committee: Sydney, NSW, Australia)
Winks, L, Laing, AR, O’Rourke, PK, and Wright, GS (1979). Factors affecting response top urea-molasses supplements by yearling cattle in tropical Queensland. Australian Journal of Experimental Agriculture and Animal Husbandry 19, 522–529.
| Factors affecting response top urea-molasses supplements by yearling cattle in tropical Queensland.Crossref | GoogleScholarGoogle Scholar |
Winter, WH, Winks, L, and Seebeck, RM (1991). Sustaining productive pastures in the tropics. 10. Forage and feeding systems for cattle. Tropical Grasslands 25, 145–152.
Appendix

|

|