

Risk-based surveillance of avian influenza in Australia’s wild birds
John P. TraceyInvasive Animals Cooperative Research Centre, Vertebrate Pest Research Unit, Industry & Investment NSW, Forest Road, Orange, New South Wales 2800, Australia. Email: john.tracey@industry.nsw.gov.au
Wildlife Research 37(2) 134-144 https://doi.org/10.1071/WR09152
Submitted: 4 November 2009 Accepted: 8 February 2010 Published: 16 April 2010
Abstract
Context. The epidemiology of avian influenza and the ecology of wild birds are inextricably linked. An understanding of both is essential in assessing and managing the risks of highly pathogenic avian influenza (HPAI).
Aims. This project investigates the abundance, movements and breeding ecology of Australia’s Anseriformes in relation to the prevalence of low-pathogenicity avian influenza (LPAI) and provides risk profiles to improve the efficiency and relevance of wild-bird surveillance.
Methods. Generalised linear models and analysis of variance were used to examine the determinants of Anseriformes abundance and movements in Australia, and the observed prevalence of LPAI in Australia (n = 33 139) and overseas (n = 93 344). Risk profiles were developed using poultry density, estimated LPAI prevalence, the abundance of Anseriformes, and the probability of Anseriformes moving from areas of HPAI epizootics.
Key results. Analysis of Australian wild-bird surveillance data strongly supports other studies that have found the prevalence of LPAI in wild birds to be much lower (1%) in Australia than that in other countries (4.7%). LPAI prevalence was highly variable among sampling periods and locations and significantly higher in dabbling ducks than in other functional groups. Trends in Anseriformes movements, abundance and breeding are also variable, and correlated with rainfall, which could explain low prevalence and the failure to detect seasonal differences in LPAI in wild birds. Virus prevalence of faecal samples was significantly lower, whereas collecting faecal samples was 3–5 times less expensive and logistically simpler, than that of cloacal samples. Overall priority areas for on-going surveillance are provided for Australia.
Conclusions. Previous surveillance has occurred in high-priority areas, with the exception of Mareeba (North Queensland), Brisbane and Darwin, and has provided valuable information on the role of wild birds in maintaining avian influenza viruses. However, several practical considerations need to be addressed for future surveillance.
Implications. Long-term surveillance studies in wild birds in priority areas are required, which incorporate information on bird abundance, age, behaviour, breeding and movements, particularly for dabbling ducks. This is important to validate trends of LPAI prevalence, in understanding the main determinants for virus spread and persistence, and in predicting and managing future epizootics of HPAI in Australia.
Introduction
Highly pathogenic avian influenza (HPAI) has caused international concern during the past decade, particularly HPAI H5N1, because of its ability to infect and cause death in humans, its ability to cause significant mortality in wild birds (Liu et al. 2005), the longevity of HPAI circulation, the failure to isolate closely related precursor strains of low-pathogenicity avian influenza (LPAI) in wild birds (Mukhtar et al. 2007), and subsequent evolving viruses remaining highly pathogenic for poultry (Sims et al. 2005; Sims and Narrod 2009).
Between 1997 and 2004, mutations in HPAI H5N1 were progressively becoming more lethal to birds and mammals and hardier in the environment (Chen et al. 2004). Although there is still debate as to whether an independent cycle of infection of HPAI H5N1 is present in wild birds (Feare 2007; Wang et al. 2008), some evidence following wild-bird deaths suggests that wild birds, particularly anatids, can excrete virus without becoming ill (Hulse-Post et al. 2005; Gaidet et al. 2008; Keawcharoen et al. 2008) and transmission appears to occur even if the virus is difficult to detect (Stallknecht and Brown 2008).
HPAI H5N1 has now been circulating in close proximity to Australia for over 11 years and the likelihood of achieving eradication is considered low (Sims and Narrod 2009). With the exception of Antarctica, Australia remains the only continent that has not had a reported occurrence of HPAI since 1997 (Sims and Turner 2008). There has been several possible explanations for this (McCallum et al. 2008) including (1) enhanced biosecurity for the trade of live wild and domestic birds (2) low prevalence of LPAI H5 or H7 in Australia’s wild birds (Haynes et al. 2009), (3) limited interchange of anatids between Australia and Asia (Tracey et al. 2004; McCallum et al. 2008), (4) few major waterbird breeding events and low waterbird abundance (Nebel et al. 2008), (5) low poultry-farm density (Westbury 1998; cf. Hamilton et al. 2009), and (6) high biosecurity of the commercial poultry industry.
An understanding of avian influenza epidemiology in wild birds is important in assessing and managing the risks of HPAI of any type. Many H and N subtypes of LPAI have been reported in Australia’s wild birds, including H5 and H7 (Downie and Laver 1973; Downie et al. 1977; Mackenzie et al. 1984, 1985; Nestorowicz et al. 1987; Röhm et al. 1996; Peroulis and O’Riley 2004; Hurt et al. 2006; Haynes et al. 2009). Anseriformes are the primary reservoir of LPAI (Stallknecht and Brown 2008; Haynes et al. 2009), with high prevalence associated with foraging behaviour (Anas species), age, breeding and movements (Hinshaw et al. 1985; Olsen et al. 2006; Haynes et al. 2009; Munster and Fouchier 2009). Charadriiformes are also considered potentially important (Hurt et al. 2006) and regularly travel through infected areas (Tracey et al. 2004), although unique lineages of influenza viruses in Australia compared with viruses in Europe and the Americas (Banks and Alexander 1997) suggest limited virus interchange via these species.
Although the importance of wild birds in avian influenza epidemiology is now widely accepted by the international animal health community (OIE, FAO), broad-scale surveillance is logistically difficult and costly because of the natural low prevalence. Initial surveillance in Australia followed a targeted approach to improve sampling efficiency (Tracey 2005; Warner et al. 2006). East et al. (2008a , 2008b ) and Hamilton et al. (2009) have also applied useful approaches to classify risks of avian influenza in Australia. Since initial surveillance, there have been significant advancements in avian influenza epidemiology, particularly for HPAI H5N1; improved information on the abundance and movements of Australian waterfowl, although many gaps in knowledge remain (McCallum et al. 2008); and a substantial increase in wild-bird surveillance for avian influenza in Australia and internationally. This information can be used to maximise the efficiency and relevance of avian influenza wild-bird surveillance in Australia. The present project investigates trends of avian influenza in wild birds and waterfowl abundance and movements, identifies high-risk areas for avian influenza in Australia and provides recommendations for surveillance.
Materials and methods
Analysis of Australian surveillance data
Published sources of avian influenza wild-bird surveillance data were collated for Australia (n = 33 139 wild birds: Downie and Laver 1973; Downie et al. 1977; Mackenzie et al. 1984, 1985; Peroulis and O’Riley 2004; Hurt et al. 2006; Haynes et al. 2009) and overseas (n = 93 344 after Olsen et al. (2006)). Generalised linear models and analysis of variance were conducted using Asreml in R (Gilmour et al. 2002) to investigate the effects of the functional group (after Roshier et al. 2002), the availability of permanent water (km2 per 1/4° grid from Geoscience Australia and National Water Commission data on rivers, dams and inland water), season and seasonal rainfall classification (a Bureau of Meteorology classification, identifying summer-dominant, summer, uniform, winter, winter-dominant, or arid rainfall) on Anseriformes abundance (log-transformed to remove heterogeneity of variance), movements (using bird banding data) and LPAI prevalence. Differences between terms for discrete variables are presented with 95% confidence intervals.
Comparison of sample methods: field trial
The estimates of prevalence of LPAI from cloacal, oropharyngeal and faecal samples were compared with quantitative real-time reverse transcriptase PCR (qRT–PCR) in a field trial in New South Wales where all three samples were collected from the same species at the same locations and sampling periods (n = 3242 samples from 2683 wild birds). These data are part of a larger surveillance dataset for eastern Australia (P. Hansbro et al., unpubl. data), which were not included in the overall analysis of Australian surveillance data described above, but were used only to compare the three methods of sample collection. Transport media, storage, transport, operators, testing preparation and testing procedures were identical for all samples.
Details of the data-collection methods, sampling techniques and testing procedures are described elsewhere (Tracey 2005; Kirkland and Tracey 2006; see also Rose et al. 2006). Briefly, swabs were taken from live-captured or recently shot birds by inserting a swab deeply into the vent (cloacal) or oropharynx and swabbing the mucosa. The tip of the plastic-shafted swab was placed into a vial containing phosphate-buffered gelatin saline (PBGS) transport media (8 g of NaCl, 0.2 g of KCl, 1.44 g of Na2HPO4, 0.24 g of KH2PO4 dissolved in 800 mL of distilled H2O). The viral transport medium was stored frozen, or at 4°C before use. Samples were maintained cold (4°C) throughout the transport process, and transported to the laboratory within 48 h of collection. Samples were either tested on delivery, or if not able to be completed within 48 h, were stored in a −80°C freezer (or −20°C for serum samples). Testing was conducted at Elizabeth McArthur Agricultural Institute with qRT–PCR (cloacal, oropharyngeal and faecal) and the Influenza A group reactive competitive enzyme-linked immunosorbent assay (cELISA, serum), based on the method and reagents supplied by the Australian Animal Health Laboratory, Geelong (www.csiro.au/places/AAHL.html).
For faecal sampling, only freshly deposited moist samples were collected, the species or group of species were identified wherever possible, and a score given for the level of confidence in determining the species or group, as follows: Highly likely (sample collected immediately after a bird was observed defaecating), Likely (bird observed in the area immediately before collecting samples), Possible (bird observed in the area within 1 h of sampling), Unknown (birds known to occur in the area). The abundance of birds was estimated with point counts (Bibby et al. 2000) each morning before collecting samples at each site, which aided species identification. Size and shape of the faeces was distinguishable for different groups of species (ducks, large waders, small waders). The swab was lightly coated with faeces. Only samples where the species was identified as Likely or Highly likely were included when comparing sample techniques.
Costs of collection methods were estimated and included labour ($15 h–1), costs of consumables (feed for traps, ammunition), and the average number of samples collected per hour. To allow for direct comparison of collection methods, cost of travel (vehicle, fuel, labour) to sites was not included.
Risk profiles
Ecological and epidemiological information has been used to assign risks of exposure by wild-bird species and location according to a range of variables to achieve the following two main aims:
-
to assess the risk of endemic LPAI viruses in wild birds becoming highly pathogenic through interactions with poultry (Surveillance Aim 1) and
-
to assess the risk of wild birds introducing foreign subtypes of avian influenza (Surveillance Aim 2).
Risk profiles are consistent with ‘exposure assessment’ under the OIE risk analysis framework (Murray 2002) and were developed for Surveillance Aim 1 by using (in order of importance): (a) log of poultry density (Robinson et al. 2007), (b) the estimated prevalence of LPAI, and (c) the abundance of Anseriformes. Risk profiles developed for Surveillance Aim 2 used (in order of importance): (a) the probability of moving from areas where HPAI epizootics have occurred in 2003–09 (FAO 2009; OIE 2009; WHO 2009), (b) the abundance of Anseriformes known to move into South-east Asia and (c) the estimated prevalence of LPAI.
To classify the risks to poultry, the risk of incursion was assumed to be dependent on poultry density (Robinson et al. 2007), using the natural log of the number of birds (Snow et al. 2007). The abundance of Anseriformes was estimated by using reporting rate and bird-count data from Birds Australia (Barrett et al. 2003). Atlas data were collected during the ‘New Altas of Australian Birds’ project 1998–2002 from 279 000 bird surveys by 7000 observers. Australian Bird Count Data were collected during 79 000 surveys involving repeated counts of birds by 952 observers at 1681 sites between 1989 and 1995. Surveys for both datasets followed the standard methods for Birds Australia’s 20-min, 2-ha search (Barrett et al. 2003), with the Australian Bird Count Data including complete counts of all individual birds observed, as well as the number of species observed. The relationship between the number of birds and number of species per observation was examined to test the use of reporting rate (number of surveys a bird species was present divided by the total number of surveys for each 1/4° map grid) as an index of abundance. For each 1/4° grid cell, abundance and prevalence was estimated separately for functional groups, which was found to be important in predicting LPAI prevalence (see Results).
Distances moved and movement probabilities for Anseriformes were estimated using banding data from the Australian Bird and Bat Banding Scheme (www.environment.gov.au/biodiversity/science/abbbs/, accessed 1 January 2009). Spatial analyses were conducted in Arcview 3.2 (ESRI, Redlands, CA) and Manifold® (Carson City, NV). In addressing Surveillance Aim 2, movement probabilities were estimated for species identified as conducting regular or occasion movements in South-east Asia (after Tracey et al. 2004; Delaney and Scott 2006; Table 1), using a movement probability model (see Results; y = 22 928 x –2.2541, where x is the distance to the current distribution of HPAI epizootics).

|
Prevalence of LPAI for each grid cell (p total) was estimated by

where pf is the prevalence of LPAI according to the functional group, using Australian surveillance data (Table 1) and af is the abundance index for Anseriformes in each functional group in each 1/4° grid cell.
The final scores were calculated with a normalised weight (wi ), by using a rank sum (2) (Malczewski 1999),

where wi was the normalised weight for the jth criterion, n was the number of criteria under consideration (k = 1, 2, …., n), and rj was the rank position of the criterion. Each criterion was weighted (n − rj + 1) and then normalised by the sum of all weights, i.e. ∑(n − rk + 1).
The value for each criterion for each grid cell was normalised by using (3) before applying weights, as follows:

where δ is the normalised value and d is the original value.
Results
Analysis of Anseriformes movements and abundance
The number of species recorded during Australian Bird Counts was found to be sufficient in predicting the log of the number of birds per observation (y = 0.6913x + 1.4456, P < 0.001). Movement probabilities were estimated for Anseriformes by using distance moved from recapture data (n = 8095), with a power model showing a good fit to the data (r 2 = 0.89, y = 22 928x –2.2541). From banding and recovery data, within 14 days of capture, 75% of birds remained within 5 km of capture, 80% within 10 km, 90% within 35 km, and 95% within 100 km; the maximum distance moved from the capture location was 2305 km (n = 1314). Greater movements of Anseriformes occurred in winter (mean = 247 km ± 79, n = 200) than in other seasons (summer: mean = 92 km ± 12, n = 1013; autumn: mean = 63 km ± 10, n = 643; spring: mean = 39 km ± 13, n = 440).
The abundance of Anseriformes was correlated with the availability of permanent water (P = 0.00025, y = 28.248x + 0.448, r 2 = 0.814) and the seasonal rainfall zone (P < 0.0001), as follows (in a decreasing order of abundance – mean reporting rate): uniform (0.57 ± 0.05, n = 1 291), summer (0.47 ± 0.03, n = 1 840), winter-dominant (0.46 ± 0.05, n = 496), winter (0.42 ± 0.02, n = 2 631), summer-dominant (0.29 ± 0.03, n = 2625) and arid (0.19 ± 0.02, n = 4 548) rainfall zone.
Analysis of Australian surveillance data
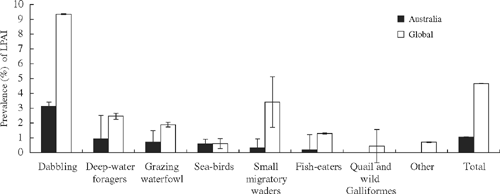
LPAI prevalence was significantly greater in dabbling ducks than in all other functional groups for Australia and overseas (Fig. 1). Prevalence of LPAI was significantly lower in Australia (1.04% ± 0.06, n = 29 167) than that in other countries (4.67% ± 0.02, n = 95 441), with the prevalence 2.6–4 times less for all functional groups, with the exception of seabirds, where there was no significant difference between Australia and overseas, and small migratory waders, where the prevalence was 11 times less in Australia than in overseas (Fig. 1). However, sampling is unlikely to be representative across all species and locations, particularly for dabbling ducks in North America where many samples are regularly taken in areas of previous high virus activity. In Australia and overseas, LPAI was either not detected or was of low prevalence for other functional groups (quail and wild Galliformes: 0/27 (Australia), 4/899 (overseas); large waders: 0/58 (Australia), 0/87 (overseas); small resident waders: 0/260 (Australia), 1/58 (overseas); birds of prey: 0/6 (Australia), 2/192 (overseas); pigeons and doves: 0/1 (Australia), 1/166 (overseas); or bush birds: 0/34 (Australia), 0/92 (overseas)).
|
Comparison of sample methods: field trial
The cost of collecting faecal samples ($1.95 per sample) was less than the cost of collecting samples by shooting ($6.12 per bird) and trapping ($9.10 per bird). When compared on the same populations during the same time periods, detection of antibodies with cELISA from serum (18.45% ± 4.38, 95% confidence interval, n = 374) was much more likely than detection of the virus (1.07% ± 2.16, n = 2 868). Also, the prevalence of LPAI viruses detected with qRT–PCR was similar for cloacal (2.27% ± 0.97, n = 948) and oropharyngeal (2.17% ± 2.3, n = 185) samples, whereas it was significantly lower from faecal samples (0.29% ± 0.28, n = 1 735).
Risk profiles
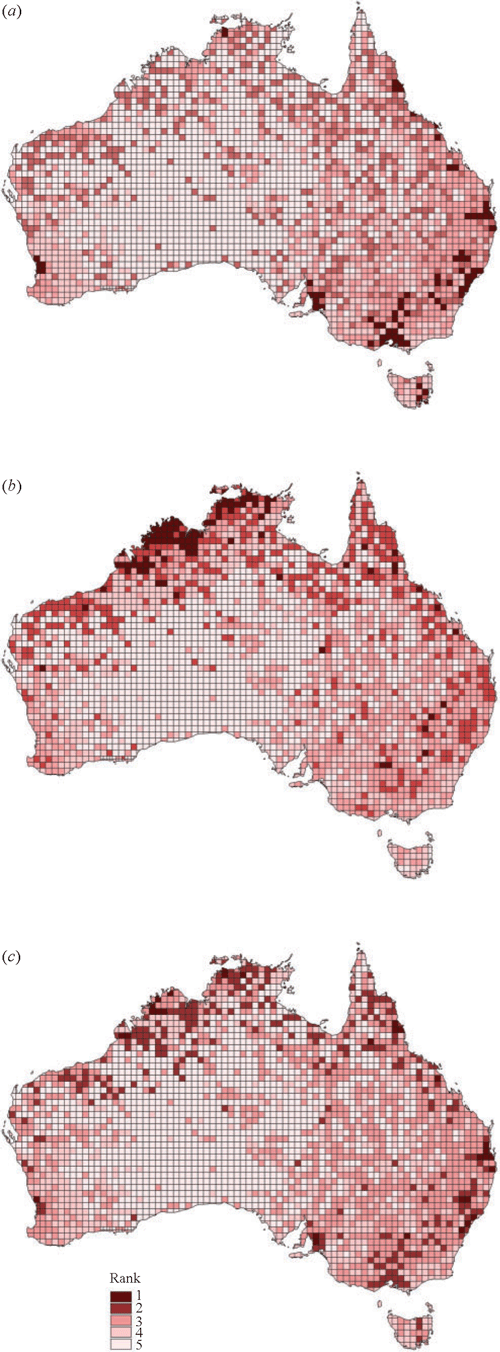
On the basis of risk profiles, highest priorities to assess risks of endemic viruses becoming highly pathogenic are in the region of state capitals, i.e. Melbourne, Sydney, Brisbane, Adelaide, Perth, Darwin and Hobart, and in the Mareeba area near Cairns, Queensland (Fig. 2a ). Current poultry densities for areas where previous HPAI epizootics occurred are 266 km–2 for Keysborough, Victoria (1976, Turner 1976), 464 km–2 for Bendigo, Victoria (1985, Barr et al. 1986; 1992, Selleck et al. 1997), 222 km–2 for Lowood, Queensland (1994, Westbury 1998), and 900 km–2 for Tamworth, New South Wales (1997, Selleck et al. 2003). Highest priorities to assess risks of wild birds introducing foreign viruses are the regions of north-western Australia from Broome through to Arnhem Land, particularly in the Kimberley, Western Australia (Fig. 2b ). Combined ranks to address both surveillance aims include all these high-priority locations (Fig. 2c ). Australian surveillance (1971–2007) has generally occurred in these priority areas, with the exception of Brisbane, Darwin and Mareeba, where surveillance is currently underway.
|
Discussion
Anseriformes and avian influenza in Australia
Abundance and movement patterns for Anseriformes are found to be irregular, varying with the availability of permanent water and seasonal rainfall, which is commonly reported for Australian anatids, with flood events and temporary rainfall particularly important (Roshier et al. 2001). Anatids are often more dispersive in arid areas, and more sedentary and abundant on permanent water (Frith 1982; Woodall 1985). The greater Anseriformes movements being evident in winter is consistent with historical studies in southern Australia, where anatids were previously thought to be more concentrated (high local abundance) in summer and dispersive in winter (low local abundance) (Ford 1958). However, cues for and patterns of anatid movements are complex, determined by individual behavioural strategies (Roshier et al. 2008) and surface water over large spatial scales (Roshier et al. 2001), rather than season. Anatid movements and abundance in Northern Australia, however, may fluctuate seasonally, where large numbers can congregate during the dry (May–October, includes southern winter) period and disperse very widely during the wet (November–April) period (Morton et al. 1990).
The abundance and movements of waterbirds have implications for understanding persistence of LPAI in natural reservoirs and for managing HPAI epizootics. In Australia, LPAI would be expected to peak in Anseriformes during ‘boom’ breeding periods where thousands of birds congregate on major wetlands or floodplains. This occurs infrequently, with the largest breeding events occurring perhaps once every 10 years. In tropical Australia, LPAI may be more seasonal with peaks expected in winter, and greater potential for dispersal of LPAI in summer. This is consistent with HPAI H5N1 in tropical South-east Asia, where prevalence was significantly higher in winter, when large flocks of Anseriformes congregate during the winter (= dry season) (Siengsanan et al. 2009).
The lower prevalence of AI found in Australia than overseas (Olsen et al. 2006; Haynes et al. 2009) is likely to be a result of differences in the behaviour and movements of Australian Anseriformes from those overseas (Tracey et al. 2004; McCallum et al. 2008). Australia is dry with irregular rainfall and as a consequence breeding and movements of waterbirds are irregular. During the past 20 years, breeding has been infrequent and waterbird abundance has declined markedly in some areas (Kingsford and Porter 2006), by up to 80% for some species (Nebel et al. 2008). Loss of wetlands because of dams, water extractions and levee banks, particularly in south-eastern Australia, is likely to have contributed to these declines (Kingsford 2000; Nebel et al. 2008).
The persistence of avian influenza viruses is likely to be affected by the regularity of breeding, as well as movement patterns, both being correlated with water availability (Frith 1982). Hence, breeding occurs in southern Australia in spring and in northern Australia at the end of the wet season (April–May, southern autumn). During severe drought, most Australian anatids do not breed (Frith 1982), which is likely to limit LPAI prevalence. Increased virus prevalence following breeding is often observed or assumed for animal pathogens, including avian influenza virus (Hinshaw et al. 1985; Alfonso et al. 1995), as a result of the boost in immunologically naïve individuals (juveniles) (Clark and Hall 2006; Munster and Fouchier 2009).
LPAI in Australian wild birds was highly variable among sampling periods and locations and no seasonal trends were apparent. This is likely to be a consequence of a lack of long-term studies (low sample sizes over time) coupled with a high variability in rainfall and Anseriformes movements and abundance between climatic zones (northern: wet season – summer-dominant rainfall v. southern: uniform or winter-dominant rainfall).
Functional group was clearly important in predicting LPAI prevalence, with dabbling ducks identified as the main reservoir for Australia and overseas. The propensity of dabbling ducks to skim surface water is a likely explanation (Olsen et al. 2006). Avian influenza viruses are known to persist in water (Webster et al. 1978; Stallknecht et al. 1990a , 1990b ; Brown et al. 2009; Roche et al. 2009) and high levels of faecal material may occur on the surface (Lang et al. 2008).
The substantial difference in prevalence (11 times) for small migratory waders between Australia and overseas may suggest that these species are unlikely to be responsible for transferring viruses into Australia. Rather, these species may act as a sentinel for endemic viruses maintained by dabbling ducks. In contrast, seabirds, whereas also conducting regular global travel, have similar prevalence between Australia and overseas. This may support the view that seabirds maintain viruses that are unique from viruses on mainland Australia, which is consistent with their behaviour and movements and phylogenetic differences among virus groups (Munster and Fouchier 2009). Future investigations of genetic differences between Australian and Eurasian and American subtypes may confirm these trends.
Practical considerations of sample-collection methods and testing procedures are important to consider when interpreting results of surveillance (Munster et al. 2009) and in preparation for future HPAI epizootics. Although variable, faecal sampling was three and five times less expensive than sampling involving shooting and trapping respectively. However, the significantly lower prevalence from faecal samples than from cloacal samples highlights the need for reporting results separately. Possible reasons include degradation of samples (low volumes of RNA), or contamination as a result of excess faecal material or other substances from the environment. In comparison, Pannwitz et al. (2009) reported similar recovery rates from faecal and cloacal samples for some species (geese and swan, but not ducks). Pannwitz et al. (2009), however, compared recovery rates from different locations and time periods, which is problematic because of low prevalence and considerable variation in prevalence commonly reported between locations and over time. Improved collection procedures for faecal samples may increase the rate of detection, for example, by minimising the amount of faecal material, or collecting samples from hardened or more sterile surfaces (e.g. concrete, bitumen, compacted soil or gravel, sand and decks). The prevalence from cloacal swabs was not significantly different from that from oropharyngeal swabs, which is consistent with Peroulis and O’Riley (2004). However, Ellström et al. (2008) and Munster et al. (2009) reported significantly higher LPAI prevalence from cloacal samples. In comparison, for HPAI H5N1, virus recovery was significantly higher from the respiratory tract than from the cloaca (Sturm-Ramirez et al. 2005; Keawcharoen et al. 2008). For on-going surveillance, faecal (environmental) samples may be collected as a rapid and cost-effective means of investigating virus presence. However, to verify virus prevalence, the collection of oropharyngeal and cloacal samples from hunted or captured birds is recommended.
Risk profiles
There is some uncertainty as to the role of poultry density in initiating HPAI in Australia. Although a shift in pathogenicity for avian influenza can occur rapidly (Brugh and Beck 1992, one or two passages; Arzey 2005), population size or density is likely to be important in determining the levels of prevalence, transmissibility and mutation rates for many viruses (e.g. rabbit haemorrhagic disease virus in rabbits (Calvete and Estrada 2000; Henzell et al. 2002), brucellosis in bison (Dobson and Meagher 1996), Mycoplasma gallisepticum in house sparrows (Hochachka and Dhondt 2000)), including avian influenza (Bunn 2004; Turner 2004; Pfeiffer et al. 2007; Snow et al. 2007). Westbury (1998) suggested that poultry-farm density was low in the first four HPAI epizootics in Australia. However, the current study indicates that both poultry-farm density and poultry density are highest in the areas where previous epizootics occurred; areas where all five HPAI epizootics took place are ranked highest by using poultry population per 1/4° grid. Hamilton et al. (2009) identified the density of poultry farms as a risk factor for HPAI in Australia, listing five regions (the Sydney region, Central Coast NSW, Tamworth, Mornington Peninsula and Bendigo) that had poultry-farm density equal to or greater than regions of Canada and Italy affected by large epizootics of HPAI (>0.05 farms km–2), which is consistent with the current study. Hamilton et al. (2009) also emphasised the importance of biosecurity measures to prevent the spread of the virus from infected farms in the event of an epizootic, resulting from service providers regularly contacting multiple farms.
Although currently unavailable, future risk profiles could incorporate additional variables, including housing (caged, floor, free range, barn, deep litter; Pfeiffer 2006; Fossum et al. 2009) and the type of operation (pullets, breeders, broilers, layers; Snow et al. 2007). Poultry-farm density may also be more appropriate than poultry density in predicting spread, once an epizootic occurs (Truscott et al. 2007; Hamilton et al. 2009).
There has been considerable debate on the ability of wild birds to spread HPAI virus over large distances while infectious (up to 14 days, Kida et al. 1980), with several recent studies suggesting that this is likely to have occurred (Sabirovic et al. 2006; Stallknecht and Brown 2008). However, the persistence of virus in the environment, the connectivity of the landscape relevant to wild birds (particularly dabbling ducks, Roshier et al. 2001) and the frequency of movements for multiple species (McCallum et al. 2008) are likely to be more important than individual bird movements within short periods. Bird populations can maintain avian influenza viruses despite low prevalence (Stallknecht and Brown 2008) and viruses can remain infective in freshwater lakes for 4 days at 22°C, more than 30 days at 0°C (Webster et al. 1978), or up to 200 days at 17°C, when virus concentrations are higher (Stallknecht et al. 1990b ).
When estimating the distance moved from banding data, there are several biases that should be considered when interpreting risk profiles. In particular, individual ducks are more likely to be recaptured at the same location than elsewhere when consecutive trapping periods occur at the same location. This would create an underestimate of HPAI risk for these criteria. Recoveries may also be more likely where damage mitigation permits are issued to protect rice. Satellite transmitters have demonstrated that large movements of grey teal can occur within hours (up to 345 km) (Roshier et al. 2006), and within days (up to 1268 km) (Roshier et al. 2008), with some birds returning to their point of origin. These individual movements would have been difficult to detect with banding studies. However, movement probabilities estimated in the current study (y = 22 928x –2.2541, see Results) are consistent with overall patterns of movement reported using satellite transmitters (Roshier et al. 2006, 2008). For example, Roshier et al. (2006) found that 78% and 83% of grey teal movements occurred within 5 km in the Riverina and Lake Eyre Basin respectively.
To develop risk profiles for avian influenza in wild birds, a range of simple seasonal and climatic variables have been explored to explain the abundance of anatids and the likelihood of their movement over a large area. However, these ignore the finer-scale processes of wetland quality, the temporary availability of wetlands and flood events. These are known to be important in predicting anatid movements and abundance, particularly in arid Australia; however, they are difficult to incorporate when presenting spatial data that can be interpreted over time.
Although highest priorities have been assigned to dabbling ducks, because they represent the major reservoir of LPAI in Australia, surveillance of other species should not be excluded. Migratory and resident Charadriiformes, seabirds (including pelagic gulls and terns and Procellariiformes), quail, ratites and other functional groups may also play a role in maintaining avian influenza viruses, including those with unique lineages (e.g. gulls and terns, Munster and Fouchier 2009).
There are many uncertainties that affect the risks of an incursion of HPAI. Risk profiles developed here are not for predicting future epizootics, but rather, are a tool to maximise the efficiency and relevance of wild-bird surveillance, and to provide insights into patterns of LPAI occurrence. Hence, poultry producers should continue to maintain high biosecurity (including limiting contact with wild birds, regular treatment of water, rapid reporting of unusual mortalities), regardless of whether they are located in high- or low-priority areas. The major risks for poultry operations are likely to be Anseriformes in the vicinity, a failure in biosecurity (e.g. water quality or entry of contaminated personnel) and confined poultry of sufficient density to allow development and dissemination of a pathogenic virus (Bunn 2004). However, there are other potential sources of LPAI, including live-bird markets and movements of domestic birds (poultry, turkeys, ducks, emus, quails) (Arzey 2004), and more important modes of transmission following outbreaks of HPAI (Sims et al. 2005; Feare 2007; Gilbert et al. 2008; Hamilton et al. 2009). In Australia, service providers regularly contact multiple farms (Hamilton et al. 2009) and are a direct potential source of secondary spread.
Previous surveillance for avian influenza in Australia has generally occurred in areas identified as highest priority, with the exception of Mareeba (northern Queensland), Brisbane and Darwin, with the current Avian Influenza Wild Bird Surveillance Program addressing these gaps. This surveillance has provided valuable information on the role of wild birds in maintaining LPAI viruses, and provides the basis for future insights into global patterns of avian influenza, in particular in the investigation of genetic similarities of subtypes between continents. However, surveillance has been sporadic, with a limited number of samples collected (35 000 samples in 1970–2007 in Australia v. 300 000 samples per year in other countries, Munster and Fouchier 2009) and with information on bird abundance, age, behaviour, breeding and movements rarely being collected during surveillance activities. This limits our ability to offer explanations for the spatial and temporal variability of virus prevalence. Enhanced surveillance in priority areas that incorporates ecological information over a longer time frame is important to validate trends of LPAI prevalence, in understanding the main determinants for virus spread and persistence, and in predicting and managing future epizootics of HPAI in Australia.
Acknowledgements
I am indebted to Glen Saunders, Peter Fleming (Industry and Investment NSW), Rupert Woods, Leesa Haynes, Tiggy Grillo (Australian Wildlife Health Network), Chris Bunn (Department of Agriculture Fisheries and Forestry), Tony Peacock (IACRC) and Brian Boyle (Game Council NSW) for their on-going support and for collaborating on aspects of this project, and Camilla Myers for suggesting this paper. Thanks also go to Brian Lukins, Franz Zikesch, Megan Moppett, Brent Waldron, Shannon Slade and Game Council NSW for assistance with bird sampling; Peter Kirkland and Edla Arzey for testing samples; Remy van de Ven and Steven McLeod for valuable advice on data analysis; the staff of the Vertebrate Pest Research Unit for advice and technical support, and Simone Warner, David Roshier, Phil Hansbro and the Avian Influenza Wild Bird Steering Group for useful discussions. Thank you to David Drynan and Andrew Silcocks and many participants of Birds Australia Atlas, the Australian Bird Count Data and the Australian Bird and Bat Banding Scheme for collecting and collating data on anatids, in particular Greg Hocking, Clive Minton, Roz Jessop, Victorian Wader Study Group, Australasian Wader Study Group, Raoul Mulder and Patrick-Jean Guay. Thank you to Iain East and Graeme Garner (Department of Agriculture Fisheries and Forestry) for advice on Australian commercial poultry operations. Funding was provided by the Wildlife and Exotic Disease Preparedness Program, Industry and Investment NSW and the Invasive Animals Cooperative Research Centre.
Alfonso, C. P. , Cowen, B. S. , and van Campen, H. (1995). Influenza A viruses isolated from waterfowl in two wildlife management areas of Pennsylvania. Journal of Wildlife Diseases 31, 179–185.
| PubMed |
Barr, D. A. , Kelly, A. P. , Badman, R. T. , and Campey, A. R. (1986). Avian influenza on a multi-age chicken farm. Australian Veterinary Journal 63, 195–196.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Brown, J. D. , Goekjian, G. , Poulson, R. , Valeika, S. , and Stallknecht, D. E. (2009). Avian influenza virus in water: infectivity is dependent on pH, salinity and temperature. Veterinary Microbiology 136, 20–26.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Bunn, C. (2004). Correspondence regarding the role of wild aquatic birds in the epidemiology of avian influenza in Australia. Australian Veterinary Journal 82, 4.
| Crossref | GoogleScholarGoogle Scholar |
Chen, H. , Deng, G. , Li, Z. , Tian, G. , Li, Y. , Jiao, P. , Zhang, L. , Liu, Z. , Webster, R. G. , and Yu, K. (2004). The evolution of H5N1 influenza viruses in ducks in southern China. Proceedings of the National Academy of Sciences, USA 101, 10 452–10 457.
| Crossref | GoogleScholarGoogle Scholar |
Clark, L. , and Hall, J. (2006). Avian influenza in wild birds: status as reservoirs, and risks to humans and agriculture. Ornithological Monographs 60, 3–29.
| Crossref | GoogleScholarGoogle Scholar |
Dobson, A. , and Meagher, M. (1996). The population dynamics of Brucellosis in the Yellowstone National Park. Ecology 77, 1026–1036.
| Crossref | GoogleScholarGoogle Scholar |
Ford, J. R. (1958). Seasonal variation in populations of Anatidae at the Bibra Lake District, Western Australia. Emu 58, 31–41.
Gaidet, N. , Cattoli, G. , Hammoumi, S. , Newman, S. H. , and Hagemeijer, W. , et al. (2008). Evidence of infection by H5N2 highly pathogenic avian influenza viruses in healthy wild waterfowl. PLoS Pathogens 4, e1 000 127.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Hamilton, S. A. , East, I. J. , Toribio, J.-A. , and Garner, M. G. (2009). Are the Australian poultry industries vulnerable to large outbreaks of highly pathogenic avian influenza? Australian Veterinary Journal 87, 165–174.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Lang, A. S. , Kelly, A. , and Runstadler, J. A. (2008). Prevalence and diversity of avian influenza viruses in environmental reservoirs. Journal of General Virology 89, 509–519.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Morton, S. R. , Brennan, K. G. , and Armstrong, M. D. (1990). Distribution and abundance of ducks in the Alligator Rivers region, Northern Territory. Wildlife Research 17, 573–590.
| Crossref | GoogleScholarGoogle Scholar |
Nebel, S. , Porter, J. L. , and Kingsford, R. T. (2008). Long-term trends of shorebird populations in eastern Australia and impacts of freshwater extraction. Biological Conservation 141, 971–980.
| Crossref | GoogleScholarGoogle Scholar |
Olsen, B. , Munster, V. J. , Wallensten, A. , Waldernstrom, J. , Osterhaus, A. D. M. E. , and Fouchier, R. A. M. (2006). Global patterns of influenza A virus in wild birds. Science 312, 384–388.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Roshier, D. A. , Robertson, A. I. , Kingsford, R. T. , and Green, D. G. (2001). Continental-scale interactions with temporary resources may explain the paradox of large populations of desert waterbirds in Australia. Landscape Ecology 16, 547–556.
| Crossref | GoogleScholarGoogle Scholar |
Selleck, P. W. , Gleeson, L. J. , Hooper, P. T. , Westbury, H. A. , and Hansson, E. (1997). Identification and characterisation of an H7N3 influenza A virus from an outbreak of virulent avian influenza in Victoria. Australian Veterinary Journal 75, 289.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Sims, L. D. , DomèNech, J. , Benigno, C. , Kahn, S. , Kamata, A. , Lubroth, J. , Martin, V. , and Roeder, P. (2005). Origin and evolution of highly pathogenic H5N1 avian influenza in Asia. The Veterinary Record 157, 159–164.
| PubMed |
Stallknecht, D. E. , Kearney, M. T. , Shane, S. M. , and Zwank, P. J. (1990a). Effects of pH, temperature, and salinity on persistence of avian influenza viruses in water. Avian Diseases 34, 412–418.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Tracey, J. P. , Woods, R. , Roshier, D. , West, P. , and Saunders, G. R. (2004). The role of wild birds in the transmission of avian influenza for Australia: an ecological perspective. Emu 104, 109–124.
| Crossref | GoogleScholarGoogle Scholar |
Webster, R. G. , Yakhno, M. , Hinshaw, V. S. , Bean, W. J. , and Murti, K. G. (1978). Intestinal influenza: Replication and characterization of influenza virus in ducks. Virology 84, 268–278.
| Crossref | GoogleScholarGoogle Scholar |
Woodall, P. F. (1985). Waterbird populations in the Brisbane region, 1972–83, and correlates with rainfall and water heights. Australian Wildlife Research 12, 495.
| Crossref | GoogleScholarGoogle Scholar |
{kind=link}