

Ant mediated dispersal of spiny stick insect (Extatosoma tiaratum) eggs and Acacia longifolia seeds is ant-species dependent
Hannah R. Smart A B * , Nigel R. Andrew A and James C. O’Hanlon A C
A B * , Nigel R. Andrew A and James C. O’Hanlon A C
A Insect Ecology Lab, School of Environmental and Rural Science, University of New England, Armidale, NSW 2351, Australia.
B Hawkesbury Institute for the Environment, Western Sydney University, Locked Bag 1797, Penrith, NSW 2751, Australia.
C School of Science and Technology, University of New England, Armidale, NSW 2351, Australia.
Australian Journal of Zoology 70(4) 105-114 https://doi.org/10.1071/ZO22036
Submitted: 8 September 2022 Accepted: 16 February 2023 Published: 27 April 2023
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Myrmecochory is a widespread mutualism between angiosperm plants and ants, where seed dispersal is facilitated by a nutrient-rich appendage known as the elaiosome. Some species of phasmids (Phasmatodea) have convergently evolved an appendage, the egg capitulum, that is analogous to the elaiosome. Research into ant-mediated dispersal of phasmid eggs is limited. It is unknown whether there are species-specific interactions between ants and phasmid eggs, nor if this variation in ant species behaviour towards eggs parallels behaviour towards seeds. We observed the behaviour of five Australian ant species towards Extatosoma tiaratum eggs and Acacia longifolia seeds. We found that ant species show significant variation in the likelihood of dispersing phasmid eggs and plant seeds. Iridomyrmex purpureus removed the largest quantities of eggs and seeds. Rhytidoponera metallica also removed large amounts of seeds but less eggs. Three species did not remove any eggs but removed small amounts of seeds. We found a species-specific component to dispersal of phasmid eggs and seeds by ants, indicating that this mutualism may depend on partner identity and abundance. Although seeds and eggs have convergently evolved to exploit ant behaviours, they elicit different behaviours in certain ant species, highlighting the complex nature of this interaction.
Keywords: animal vectors, ants, coevolution, dispersal, eggs, mutualism, myrmecochory, phasmids, seeds.
Introduction
Seed dispersal facilitated by ants (Formicidae), commonly referred to as myrmecochory, is a mutualism that has evolved between angiosperm plants and numerous ant species worldwide (Andersen 1988; Beaumont et al. 2013; Bologna et al. 2017; Lengyel et al. 2010). The diaspores of these plants possess an elaiosome, a knob-like appendage, which is enriched with essential nutrients (Berg 1975; Hughes and Westoby 1990). This structure acts as an incentive that induces both omnivorous and herbivorous ants to retrieve the seed and transport it back to the nest (Hughes and Westoby 1990; Bologna et al. 2017; Rocha et al. 2018). Following dispersal, the elaiosome is typically detached, leaving the remainder of the seed intact, before being buried in nest chambers or discarded in refuse piles nearby (Beaumont et al. 2013). In some systems, seeds are also randomly discarded by ants rather than being buried or discarded in refuse piles (Canner et al. 2012). The benefits to plants associated with myrmecochory include reducing parent and sibling competition and thus increasing seed fitness (Giladi 2006), evading predators, particularly granivores and other seed predators (Giladi 2006; Christianini et al. 2007; Tanaka et al. 2015), protection from fire (Beaumont et al. 2018) and the displacement of seeds to nutrient-rich microsites (Leal et al. 2007; Gutierrez Rapalino and Domínguez Haydar 2017).
Another group that takes advantage of ants as a dispersal mechanism are stick and leaf insects, or phasmids (Phasmatodea). Known for their extraordinary camouflage, these insects resemble leaves, twigs, lichen, and moss, both phenotypically and behaviourally (Bradler and Buckley 2018; Robertson et al. 2018). Female phasmids are typically flightless and slow-moving, and thus have limited dispersal capabilities. The females of a vast number of species, however, flick eggs to the leaf litter below, while remaining immobile in the foliage, which may have evolved to facilitate egg dispersal (Hughes and Westoby 1992a; Goldberg et al. 2015; O’Hanlon et al. 2020). This oviposition technique is utilised by a majority of the 3000 species worldwide (Hughes and Westoby 1992a; Shelomi 2011; Goldberg et al. 2015; Robertson et al. 2018). In fact, a study conducted by Robertson et al. (2018), which examined the evolution of phasmid oviposition techniques of 284 species representing all major lineages, found that 168 of the 284 (59%) species exhibit egg dropping or flicking behaviours. Eggs produced by some species of phasmids that are expelled by flicking them to the leaf litter bear a capitulum, a knob-like structure attached to the operculum. These eggs bear a strong resemblance to myrmecochorous plant seeds (Fig. 1a, b), with the capitulum being equivalent to the plant seed elaiosome in both structure and function in that it induces dispersal by ants (Hughes and Westoby 1992a; Stanton et al. 2015; Robertson et al. 2018).

|
The capitulum exhibits a similar chemical profile to that of the elaiosome, with both containing large amounts of the same fatty acids (Fischer et al. 2008; Stanton et al. 2015). The main chemical components of the capitulum include oleic acid, palmitic acid, linoleic acid, linolenic acid and stearic acid (Stanton et al. 2015). Similarly, the elaiosome contains a number of lipid species, with oleic acid, a chemical thought to induce corpse-carrying behaviour, being the most abundant (Boulay et al. 2006; Fischer et al. 2008; Miller et al. 2020). The chemicals present in the capitulum are thought to facilitate the dissemination and burial of eggs by providing a food reward to ants (Hughes and Westoby 1992a; Stanton et al. 2015; Robertson et al. 2018).
The evolution of ant-mediated dispersal in plants has been well studied (Giladi 2006). In comparison, this dispersal strategy in phasmid species has received little attention (see Compton and Ware 1991; Hughes and Westoby 1992a; Windsor et al. 1996; Stanton et al. 2015; O’Hanlon et al. 2020; Yamada et al. 2021). The dispersal of phasmid eggs by ants has been documented in only a handful of species (Compton and Ware 1991; Hughes and Westoby 1992a; Windsor et al. 1996; Stanton et al. 2015; Yamada et al. 2021); however, it is believed that this interaction is relatively common among phasmid species worldwide (Hughes and Westoby 1992a; Stanton et al. 2015). Like myrmecochorous plant seeds, capitulate phasmid eggs reap some benefits from ant-mediated dispersal. Hughes and Westoby (1992a) found that eggs buried in ant nests were less likely to suffer from parasitism, whereas phasmid nymphs were still able to emerge even when buried under 6 cm of soil. Capitulum removal in a laboratory setting has been found to have no impact on egg viability and nymph emergence rates (Compton and Ware 1991). Similarly, phasmid eggs that were placed within an ant colony, had the capitula chewed off by ants, with the eggs and subsequent first instar nymphs being left undisturbed (Compton and Ware 1991).
Although there are some individual studies, as mentioned above, that have interactions between individual ant species and phasmid eggs, there is still much to be discovered about how phasmid eggs benefit from ant-mediated dispersal, as well as the fate of the eggs once they have been dispersed to the ant nest. Furthermore, research on plants highlights the diversity of ants that disperse seeds and suggests that species-specific interactions are limited (Giladi 2006). However, there has been no research into this aspect of the ant–phasmid interaction.
Ants show great variability in terms of both morphological and ecological characteristics, and thus not all species of ant are going to have the ability or proclivity to disperse seeds, with some species acting as ‘keystone’ dispersers (Gove et al. 2007; Ness et al. 2009; Lubertazzi et al. 2010). Ant morphological traits such as body size as well as mandible size and shape, have the potential to determine whether a particular ant species is physically capable of dispersing both seeds and phasmid eggs (Kaspari 1996; Gómez and Espadaler 1998; Ness et al. 2004; Gómez et al. 2005). Likewise, the ecology of a particular species, such as distribution and abundance of the ant species relative to myrmecochorous plant species, as well as feeding guild and foraging strategy and range, can also play a role in determining whether that species can disperse seeds or eggs (Hughes et al. 1994; Ness et al. 2009; Lubertazzi et al. 2010).
Here, we investigate how different ant species vary in their likelihood to disperse the eggs of an Australian stick insect – the spiny stick insect (Extatosoma tiaratum (Macleay, 1826)). We observed the behaviour of five ant species towards phasmid eggs in seminatural conditions and recorded the rate at which they collect phasmid eggs. We also quantified the behaviour of the same ant species towards the seeds of a native, myrmecochorous plant (Acacia longifolia). Ants differ greatly in foraging and dietary habits, and as such we predicted that the studied ant species will vary in the way they interact with both phasmid eggs and plant seeds, which will have an impact on removal rates. Furthermore, we predicted that species that disperse myrmecochorous plant seeds will display a greater propensity to disperse E. tiaratum eggs compared to species that do not collect myrmecochorous seeds.
Given that the capitula of phasmid eggs are thought to have converged on the same chemical pathways as the myrmecochorous plant seed elaiosome, we hypothesise that the presence of the capitulum on phasmid eggs should facilitate collection and dispersal in the same manner as myrmecochorous plant seeds. Thus, the phasmid eggs used in this study should be removed by ants at the same rate as that of plant seeds.
Materials and methods
Study sites
This study was conducted in Armidale, New South Wales, Australia (see Supplementary Material Appendix 1 and 2). A visual assessment of the area was conducted to locate and identify ant nests. Sites were selected based on the presence of one or more ant nests associated with the chosen ant study species. Although the distributions of the ant species selected for this study overlap with E. tiaratum distributions, the sites where we conducted our trials are not locations where this species of stick insect are found, and as such, we can assume that the ants in this study are naïve in their experience with E. tiaratum eggs.
Study species
E. tiaratum is a large arboreal stick insect native to coastal forests of eastern Australia (Brock 2001; Brock and Hasenpusch 2009). This species is a popular culture stock, not only in Australia, but also abroad, particularly in Europe, with spiny stick insects making great pets. As a result of their popularity, this species is one of the most studied phasmid species (see Brock et al. 2022 for a list of studies on E. tiaratum). Females are large (100–160 mm), heavy-bodied insects with a spikey appearance and non-functional wings (Fig. 2a; Clark 1976; Brock and Hasenpusch 2009; Schneider and Elgar 2010). Females feeding on eucalypts can produce, on average, 574 eggs during their lifetime – when feeding on a more nutritious plant (e.g. bramble) individuals have laid over 900 eggs in a lifetime (Hill et al. 2020). Males, in comparison, are smaller (75–115 mm), with thin, slender bodies and well-developed wings (Fig. 2b). The eggs produced by females possess a capitulum (Fig. 1a), which facilitates rapid removal by ants on the forest floor (Brock 2001; Brock and Hasenpusch 2009). Phasmid eggs do not vary greatly in size and shape, with all eggs within a species being generally the same size, although this is yet to be quantified. The newly hatched nymphs of this species are believed to mimic ants in the genus Leptomyrmex, allowing them to survive among the ant colony if they emerge within the nest after being collected by the ants (Key 1991).

|
Eggs of E. tiaratum were collected from a captive population housed at the University of New England. Insects were kept in indoor enclosures, at ambient room temperatures (~20–25°C). Eggs were collected regularly and stored in plastic containers at room temperature until they were used for the current experiments. Eggs were used within six months of their collection date. Due to phasmid egg dispersal being an emerging field in its infancy, it is unknown whether the way in which the eggs were stored or the time from collection to use in trials had an impact on the expression of chemical signals.
Acacia longifolia is a species of tree or shrub that grows up to 8 m in height. This species from the family Fabaceae is native to south-eastern Australia. The seeds of this species are ovoid in shape, smooth and brown or black in colour, with a large, orange elaiosome (Fig. 1b; Kodela 2005). The Acacia seeds used in this experiment were devitalised using Gamma radiation as per O’Hanlon et al. (2019) so as not to inadvertently introduce new plants to the area. Seeds were obtained from suppliers and kept at room temperature in plastic bags until they were used for the experiments. Like the phasmid eggs used in this study, seeds were stored for less than six months prior to their use in this study, but it is unknown whether the chemical signals of the elaiosome are influenced by age or storage method.
A. longifolia was an ideal species to use for this study given that it is a common model species used in myrmecochory research and thus provides a useful comparison to other studies involving myrmecochory. Although eggs and seeds are similar in size and shape (Fig. 1a, b), eggs do not have a close morphological resemblance to myrmecochorous plant seeds. Rather, chemical signals are thought to be the important part of ant–phasmid egg interactions (O’Hanlon et al. 2020).
Five focal ant species were selected. These species differed in size, morphology and behaviour as well as having varied ecological traits.
Iridomyrmex purpureus Smith, F, 1858 (meat ant)
I. purpureus is an ecologically dominant ant species typically found in Savannah woodlands and grasslands across Australia, with hundreds to thousands of individuals making up colonies (Shattuck 1999; van Wilgenburg and Elgar 2007; Middleton et al. 2018). The meat ant has a varied diet, often feeding on seeds, small invertebrates and honeydew that is secreted by sap-sucking insects. Although this species is known to feed on seeds, it is not known whether it is a seed predator or disperser. Additionally, whether it disperses phasmid eggs or eats the entire egg remains unclear (Shattuck 1999; van Wilgenburg and Elgar 2007; Middleton et al. 2018). For this study, the specimens used (n = 10) had a mean Weber’s length of 3.0 (±0.49) mm.
Iridomyrmex rufoniger Lowne, 1865
I. rufoniger is aggressive and common in urban areas and forages either diurnally or nocturnally. Its diet consists primarily of plant secretions such as honeydew; however, it may also scavenge for dead insects (Shattuck 1999; Heterick and Shattuck 2011). It is not known whether this species disperses myrmecochorous plant seeds or phasmid eggs. The specimens used in this study (n = 9) had a mean Weber’s length of 1.2 (±0.068) mm.
Rhytidoponera metallica Smith, F, 1858 (green-headed ant)
R. metallica is a common species of ant found in all habitats across the Australian landscape (Shattuck 1999; Pettit and Latty 2016). It is typically found in colonies of 30–1000 individuals, and is an opportunistic scavenger (Thomas and Framenau 2005; Pettit and Latty 2016). It is a solitary forager with a diet consisting of small invertebrates, seeds, and hemipteran secretions (Middleton et al. 2018). Within Australia, members of the genus Rhytidoponera act as keystone dispersers of myrmecochorous plant seeds and are associated with the highest dispersal rates compared to other local ant species (Gove et al. 2007; Lubertazzi et al. 2010). Although this species is a known seed collector, it is unclear whether it will disperse phasmid eggs. The specimens used in this study (n = 10) had a mean Weber’s length of 2.1 (±0.29) mm.
Camponotus aeneopilosus Mayr, 1862 (golden-tail sugar ant)
C. aeneopilosus is a relatively small species of ant, endemic to eastern Australia (Shattuck 1999; McArthur 2007). Species of Camponotus are mostly generalist scavengers, feeding on honeydew and plant secretions as well as small arthropods on occasion (Shattuck 1999). It is not known whether this species disperses myrmecochorous plant seeds or phasmid eggs. The specimens used in this study (n = 10) had a mean Weber’s length of 2.9 (±0.37) mm.
Myrmecia tarsata Smith, F, 1858 (bull ant)
M. tarsata is a large species of bull ant distributed mostly throughout eastern New South Wales and the Australian Capital Territory. M. tarsata is a diurnal forager, with a diet consisting of plant secretions and dead animal prey (Shattuck 1999). It is not known whether this species disperses myrmecochorous plant seeds or phasmid eggs. The specimens used in this study (n = 8) had a mean Weber’s length of 6.8 (±0.43) mm.
Weber’s length measurements
These basic morphometric measurements were obtained by measuring one worker/forager specimen of each species that were collected following each behavioural trial. Stacked images were created using a Leica MZ16A Auto-montage microscope and Leica Application Suite ver. 4.5.0 software (Leica Microsystems Limited), with measurements also being made using this software.
Ant response to E. tiaratum eggs
Trials were carried out during autumn (March–early May) and early spring (September–October) 2019. Ten E. tiaratum eggs were placed in the centre of a 10 × 10 cm marked arena, a minimum of 30 cm from the nest entrance. In the case of multiple nest entrances, which is common with I. purpureus nests, eggs were placed on the outer perimeter of the nest. The eggs were left in the marked arena for 1 h, and observations of ant behaviour towards the egg and the times that behaviours were performed were recorded. We noted when an ant removed the egg (farther than 5 cm) from the marked arena. Trials were filmed with a Panasonic HC-V270 High-Definition video camera, with data being recorded in real time and from playback of the collected footage, particularly when nest activity was high and discerning individual removals was difficult to ascertain in real time. This process was repeated at up to 10 nests per species. Worker/forager ants were collected from each site after each trial. Species identification was confirmed (S. Tremont) and compared with reference specimens from the University of New England Natural History Museum.
Ant response to A. longifolia seeds
Seed removal trials were carried out during spring (September–October) 2019. Ten A. longifolia seeds were placed in the centre of a 10 cm × 10 cm marked arena similar to that of the experimental set-up used for egg dispersal trials. Removals of seeds and the time of removal were recorded over a period of 1 h. A removal was defined as ‘transportation of a seed by an ant farther than 5 cm from the marked arena’. Each trial was repeated at eight nests per species. The same nests, in most cases, were used for both egg and seed trials; however, this was not always possible due to nests no longer being found in the same location when some egg trials were conducted months earlier, or nests were destroyed by human disturbances. Egg and seed trials were conducted separately on different days for each nest.
Statistical analyses
Differences between ant species in their behaviour towards phasmid eggs were investigated using a generalised linear model (GLM) assuming a binomial distribution to test whether there were differences in removal rates between ant species. Species was used as a fixed effect and the number of phasmid egg removals per hour was set as the response variable. A Chi-squared test was performed on the model to obtain significance levels (P-value).
Wilcoxon signed-rank post hoc tests were then performed on the data to compare differences in the number of removals per hour between ant species. Where appropriate, results are given as the mean ± standard deviation. An alpha level of 0.005 was used as the criterion for significance in all statistical tests, to account for all 10 pairwise interactions. This is based on a Bonferroni correction in which the alpha value of 0.05 was divided by 10.
To investigate how the removal rate of seeds differed between ant species, we used the same statistical procedures that were used to analyse egg removals. All statistical tests were carried out using RStudio ver. 1.1.442 (RStudio Team 2020) with R ver. 4.0.3, with figures produced using the R package ggplot2 ver. 3.3.3 (Wickham 2016).
Results
There was a significant difference between the ant species and the number of seeds (GLM: χ2(4) = 21.627, P < 0.001) and eggs they removed (GLM: χ2(4) = 21.245, P < 0.001). I. purpureus was responsible for the highest number of phasmid egg removals (mean = 7.2 ± 3.46 eggs per hour), compared with R. metallica (mean = 1 ± 1.89 eggs per hour), the only other species to remove phasmid eggs (Fig. 3). In comparison, all ant species were observed collecting at least some seeds (Fig. 4). I. purpureus removed the largest number of seeds (mean = 7.88 ± 3.09 seeds per hour) (Fig. 4). R. metallica removed a greater number of seeds (mean = 5.63 ± 4.69 seeds per hour) than eggs. Both I. rufoniger and M. tarsata removed a similar number of seeds (mean = 0.25 ± 0.46 seeds per hour: I. rufoniger, M. tarsata) over the study period and C. aeneopilosus removed slightly more seeds (mean = 0.63 ± 1.06 seeds per hour: C. aeneopilosus) (Fig. 4). Even though removal rates of seeds are low for these three species, they only removed seeds – no eggs were removed.

|

|
Iridomyrmex purpureus removed a significantly higher proportion of eggs compared to C. aeneopilosus (P < 0.001), I. rufoniger (P < 0.001) and M. tarsata (P < 0.001). Although R. metallica removed some eggs, the differences in removals compared I. purpureus, C. aeneopilosus, I. rufoniger and M. tarsata were insignificant (P < 0.01, P < 0.08, P < 0.08 and P < 0.12 respectively). Differences in egg removals by other species are null in the case of species where no removals were recorded (Table 1).
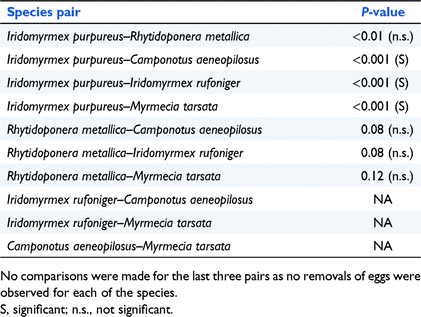
|
For seeds, I. purpureus removed significantly more seeds than did the other species (P < 0.001), with the exception of R. metallica, which removed seeds at a rate not significantly different from that of I. purpureus (P = 0.38) (Table 2). Additionally, the rate of seed removal by R. metallica was insignificant when compared with those of the other ant species (P > 0.005).
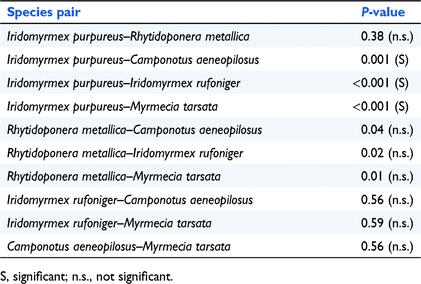
|
Discussion
The main aim of this study was to determine whether different ant species vary in how they behave towards phasmid eggs and how this compares to their behaviour towards plant seeds. Ant species show significant variation in their ability to collect and remove phasmid eggs, with I. purpureus performing the highest number of removals of E. tiaratum eggs and R. metallica removing only a small proportion of eggs. I. rufoniger, M. tarsata and C. aeneopilosus did not move any phasmid eggs.
We found that the different ant species did not remove phasmid eggs at the same rates, nor did they remove plant seeds at similar rates when presented with them. The different ant species also responded differently to both eggs and seeds, with R. metallica removing a larger number of seeds compared to eggs when presented with the options in separate trials. C. aeneopilosus, M. tarsata and I. rufoniger also removed a small proportion of seeds, while not removing any eggs. Our finding that R. metallica did not disperse large numbers of E. tiaratum eggs, contrasts with the findings of Stanton et al. (2015), who observed R. metallica frequently removing Clemacantha goliath (Goliath stick insect) eggs back to the nest. This suggests that differences in egg dispersal rates are not just due to ant species differences, but also species differences in phasmid eggs.
As a result of using a very stringent alpha value to account for multiple comparisons, however, our interpretation of the P-value shows very coarse patterns in the data, and thus doesn’t allow for a more nuanced interpretation of the values shown. This can be seen in Fig. 3, where there are definite differences in egg removal between I. purpureus and R. metallica, but this is not well reflected by the P-value. We can see general patterns between egg and seed removal between species from the figures; however, from the statistical analyses, we cannot make definite conclusions.
These results, however, do correspond with our hypothesis, in that ant species will react differently to the presence of phasmid eggs and myrmecochorous plant seeds. This can potentially be attributed to several factors, such as egg and seed characteristics, attractiveness to ants, and foraging preferences associated with different ant species.
Several characteristics of eggs and seeds may influence what species of ant are able to effectively disperse them. Diaspore and elaiosome size are major contributing factors affecting the probability that seeds will be transported towards the nest, with elaiosome size influencing whether ants can effectively carry seeds back to the nest (Hughes and Westoby 1992b; Bas et al. 2009; Leal et al. 2014). Additionally, the presence of the elaiosome acts as a handle to allow for easier manoeuvrability of the diaspore (Pfeiffer et al. 2010). Ants were frequently observed using the seed elaiosome as a handle during this study, allowing for rapid removal. The same can be said for phasmid eggs. Size and shape of eggs and their capitula are likely to affect the behaviour of ants towards phasmid eggs, with differences in shape, size, and texture of the capitula determining which species of ant can effectively disperse the eggs. Additionally, appearance of eggs may differ within a species depending on the region in which the female phasmids are found (Brock and Hasenpusch 2009) and, consequently, this may have an influence on collection and dispersal by ants. Although this is yet to be empirically tested, it provides an interesting path for future research.
The presence of the elaiosome food reward represents the biggest influence on short-term seed fate (Bas et al. 2009; Reifenrath et al. 2012). Ants may be attracted to the food reward depending on whether the nutrient content of the reward meets their nutritional requirements (Hughes et al. 1994; Reifenrath et al. 2012); however, misleading chemical cues from the plant seed, which may imply a greater nutritional reward than is present, may also be responsible for attracting ants to the plant seed (Turner and Frederickson 2013). Therefore, differences in seed removal rates may reflect differing responses to the quality of the nutritional reward offered by the elaiosome or dishonest signalling from the seed. Upon initial contact with seeds, ants may be able to perceive information about the suitability of seeds and, as such, determine whether it is worth removing (Reifenrath et al. 2012). Although a food reward is an important aspect of myrmecochory, time and energetic constraints, along with chemical signalling may also influence whether an ant is initially attracted to the plant seed, and if successful collection and subsequent removal of the seed is successful.
The phasmid egg capitula has converged upon the same chemical signalling pathways as myrmecochorous plants and contains similar chemical components to that of the myrmecochorous plant seed elaiosome (Stanton et al. 2015; O’Hanlon et al. 2020). Oleic acid, the primary fatty acid in both seeds and eggs, induces a food-carrying response in ants (Fischer et al. 2008). As such, it is likely that the capitulum of phasmid eggs, and the potential food reward it represents, contributes to attracting ants and facilitating dispersal (Fischer et al. 2008; Stanton et al. 2015). However, the phenomenon of the egg capitulum and the role that it plays in attracting ants has rarely been investigated, with only five phasmid species to date being studied (O’Hanlon et al. 2020). Given that many phasmid species possess egg capitula, it is likely that the interaction of ants with phasmid eggs has had an important role in driving the evolution and distribution of phasmids. Investigating the role that the capitulum plays as an ant attractant is still in its infancy and provides an interesting pathway for exploring the evolutionary history and complex signalling interactions between ants and phasmids.
As the plant seed elaiosome contains chemical components, primarily fatty acids, similar to the haemolymph of invertebrate prey, it is assumed that omnivorous and carnivorous guilds of ants would show a greater level of attraction to seeds (Hughes et al. 1994; Fischer et al. 2008; Stanton et al. 2015). Both I. purpureus and R. metallica are opportunistic scavengers (Lubertazzi et al. 2010). C. aeneopilosus, on the other hand, is a nectar-feeding omnivore, that is less likely to encounter and disperse seeds while foraging due to specific dietary requirements (Hughes et al. 1994). Furthermore, species that have an extensive foraging range, like I. purpureus, are more likely to encounter myrmecochorous plant seeds (Lubertazzi et al. 2010), providing a potential explanation for the high removal rate of A. longifolia by I. purpureus and R. metallica observed here, compared to other species with a restricted foraging range.
Given that the capitula exhibits the same chemical composition of the plant seed elaiosome, it is also likely that carnivorous and omnivorous guilds of ants would exhibit a greater attraction to phasmid eggs, compared with those that exhibit different foraging strategies (Hughes et al. 1994; Fischer et al. 2008; Stanton et al. 2015). This, however, has yet to be quantified, given that research on ant–phasmid interactions is scarce.
Through behavioural assessments, we have shown that, across species, ants show different behaviours in response to phasmid eggs, and consequently differ in their ability to remove eggs a sufficient distance. I. purpureus was identified as being the most effective at removing E. tiaratum eggs compared with all other species studied. Furthermore, we were able to show that ant species respond differently to both myrmecochorous plant seeds and phasmid eggs, with removals differing for both treatment types. Most notably, we observed R. metallica behaving differently towards eggs and seeds.
Ant–phasmid interactions have likely played a major evolutionary role in shaping the diversity and distribution that we see in phasmids today. As such, by understanding the behavioural and ecological mechanisms associated with ant-mediated phasmid egg dispersal, we may be able to begin to unravel this complex interaction and gain insights into not only phasmid ecology and evolution, but also the mutualistic relationship with ant partners. Ant–plant interactions have been studied broadly; however, studies quantifying visual similarities between myrmecochorous plant seeds and phasmid eggs have yet to be completed. Similarly, our understanding of ant–phasmid interactions is in its infancy.
Several factors may have an impact on dispersal of phasmid eggs by ants that remain unexplored and provide avenues for future research, including seasonality and whether time of year impacts collection of eggs by ants, egg size and appearance within and between species, the role of the capitula in facilitating dispersal by ants as well as the functional benefits of ant-mediated egg dispersal. Furthermore, the phasmid eggs are assumed to be a novel stimulus for the ants used in this study, in that they were unlikely to have encountered these eggs before; however, it is not yet known whether previous exposure to eggs would affect ant behaviour. As such, further studies investigating how experience affects ant behaviour are needed.
Given that newly hatched nymphs of E. tiaratum resemble ants in the genus Leptomyrmex, and that nymphs may differ in appearance depending on location (Brock 2016), it would be beneficial to conduct future studies that utilise different culture stocks of eggs as well as conducting egg removal trials with various species of Leptomyrmex ants. Further investigation is needed into how age of eggs and seeds influences the expression of chemical signals and, consequently, if there is an impact on dispersal by ants. Finally, the fate of phasmid eggs following collection by ants and whether eggs can remain viable within an ant nest are understudied aspects of ant–phasmid interactions and should be explored in future studies.
Supplementary material
Supplementary material is available online.
Data availability
Datasets analysed during the current study are openly available on Github at https://github.com/hsmart59/Smart_etal_2022_phasmideggdispersal.
Conflicts of interest
The authors declare no conflicts of interests.
Declaration of funding
This study was supported by funding from the Australia Pacific Science Foundation to J. C. O’Hanlon (A17/2086).
Acknowledgements
We thank Steve Tremont and Dr Sarah Hill for assistance with ant species identification.
References
Andersen, AN (1988). Dispersal distance as a benefit of myrmecochory. Oecologia 75, 507–511.| Dispersal distance as a benefit of myrmecochory.Crossref | GoogleScholarGoogle Scholar |
Bas, JM, Oliveras, J, and Gómez, C (2009). Myrmecochory and short-term seed fate in Rhamnus alaternus: ant species and seed characteristics. Acta Oecologica 35, 380–384.
| Myrmecochory and short-term seed fate in Rhamnus alaternus: ant species and seed characteristics.Crossref | GoogleScholarGoogle Scholar |
Beaumont, KP, Mackay, DA, and Whalen, MA (2013). Multiphase myrmecochory: the roles of different ant species and effects of fire. Oecologia 172, 791–803.
| Multiphase myrmecochory: the roles of different ant species and effects of fire.Crossref | GoogleScholarGoogle Scholar |
Beaumont, KP, Mackay, DA, and Whalen, MA (2018). The role of Rhytidoponera metallica (Hymenoptera, Formicidae) in facilitating post-fire seed germination of three ant-dispersed legume species. Austral Ecology 43, 128–138.
| The role of Rhytidoponera metallica (Hymenoptera, Formicidae) in facilitating post-fire seed germination of three ant-dispersed legume species.Crossref | GoogleScholarGoogle Scholar |
Berg, RY (1975). Myrmecochorous plants in Australia and their dispersal by ants. Australian Journal of Botany 23, 475–508.
| Myrmecochorous plants in Australia and their dispersal by ants.Crossref | GoogleScholarGoogle Scholar |
Bologna, A, Toffin, E, Detrain, C, and Campo, A (2017). An automated method for large-scale monitoring of seed dispersal by ants. Scientific Reports 7, 40143.
| An automated method for large-scale monitoring of seed dispersal by ants.Crossref | GoogleScholarGoogle Scholar |
Boulay, R, Coll-Toledano, J, Manzaneda, AJ, and Cerdá, X (2006). Geographic variations in seed dispersal by ants: are plant and seed traits decisive? Naturwissenschaften 94, 242–246.
| Geographic variations in seed dispersal by ants: are plant and seed traits decisive?Crossref | GoogleScholarGoogle Scholar |
Bradler S, Buckley TR (2018) Biodiversity of Phasmatodea. In ‘Insect biodiversity: science and society’. (Eds PH Adler, RG Foottit) (pp. 281–313). (John Wiley & Sons, Ltd) Available at https://onlinelibrary.wiley.com/doi/abs/10.1002/9781118945582.ch11
Brock, PD (2001). Studies on the Australasian stick-insect genus Extatosoma Gray (Phasmida: Phasmatidae: Tropoderinae: Extatosomatini). Journal of Orthoptera Research 10, 303–313.
| Studies on the Australasian stick-insect genus Extatosoma Gray (Phasmida: Phasmatidae: Tropoderinae: Extatosomatini).Crossref | GoogleScholarGoogle Scholar |
Brock, PD (2016). Extatosoma and ants. The Phasmid Study Group Newsletter 136, .
Brock PD, Hasenpusch JW (2009) ‘The complete field guide to stick and leaf insects of Australia.’ (CSIRO Publishing) Available at https://www.publish.csiro.au/book/6012 [Retrieved 11 August 2021]
Brock PD, Büscher T, Baker E (2022) Phasmida species file online. Version 5.0/5.0. Available at http://Phasmida.SpeciesFile.org [Retrieved 12 December 2022]
Canner, J, Dunn, R, Giladi, I, and Gross, K (2012). Redispersal of seeds by keystone ant augments the spread of common wildflowers. Acta Oecologica 40, 31–39.
| Redispersal of seeds by keystone ant augments the spread of common wildflowers.Crossref | GoogleScholarGoogle Scholar |
Christianini, AV, Mayhé-Nunes, AJ, and Oliveira, PS (2007). The role of ants in the removal of non-myrmecochorous diaspores and seed germination in a neotropical savanna. Journal of Tropical Ecology 23, 343–351.
| The role of ants in the removal of non-myrmecochorous diaspores and seed germination in a neotropical savanna.Crossref | GoogleScholarGoogle Scholar |
Clark, JT (1976). The capitulum of phasmid eggs (Insecta: Phasmida). Zoological Journal of the Linnean Society 59, 365–375.
| The capitulum of phasmid eggs (Insecta: Phasmida).Crossref | GoogleScholarGoogle Scholar |
Compton, SG, and Ware, AB (1991). Ants disperse the elaiosome-bearing eggs of an African stick insect. Psyche: A Journal of Entomology 98, 207–213.
| Ants disperse the elaiosome-bearing eggs of an African stick insect.Crossref | GoogleScholarGoogle Scholar |
Fischer, RC, Richter, A, Hadacek, F, and Mayer, V (2008). Chemical differences between seeds and elaiosomes indicate an adaptation to nutritional needs of ants. Oecologia 155, 539–547.
| Chemical differences between seeds and elaiosomes indicate an adaptation to nutritional needs of ants.Crossref | GoogleScholarGoogle Scholar |
Giladi, I (2006). Choosing benefits or partners: a review of the evidence for the evolution of myrmecochory. Oikos 112, 481–492.
| Choosing benefits or partners: a review of the evidence for the evolution of myrmecochory.Crossref | GoogleScholarGoogle Scholar |
Goldberg, J, Bresseel, J, Constant, J, Kneubühler, B, Leubner, F, Michalik, P, and Bradler, S (2015). Extreme convergence in egg-laying strategy across insect orders. Scientific Reports 5, 7825.
| Extreme convergence in egg-laying strategy across insect orders.Crossref | GoogleScholarGoogle Scholar |
Gove, AD, Majer, JD, and Dunn, RR (2007). A keystone ant species promotes seed dispersal in a “diffuse” mutualism. Oecologia 153, 687–697.
| A keystone ant species promotes seed dispersal in a “diffuse” mutualism.Crossref | GoogleScholarGoogle Scholar |
Gutierrez Rapalino, BP, and Domínguez Haydar, YDC (2017). Contribution of Pheidole fallax and Ectatomma ruidum (Hymenoptera: Formicidae) to the dispersion and germination of seeds in rehabilitated areas of the Cerrejón coal mine, Colombia. Revista de Biología Tropical 65, 575–587.
| Contribution of Pheidole fallax and Ectatomma ruidum (Hymenoptera: Formicidae) to the dispersion and germination of seeds in rehabilitated areas of the Cerrejón coal mine, Colombia.Crossref | GoogleScholarGoogle Scholar |
Gómez, C, and Espadaler, X (1998). Seed dispersal curve of a Mediterranean myrmecochore: influence of ant size and the distance to nests. Ecological Research 13, 347–354.
| Seed dispersal curve of a Mediterranean myrmecochore: influence of ant size and the distance to nests.Crossref | GoogleScholarGoogle Scholar |
Gómez, C, Espadaler, X, and Bas, JM (2005). Ant behaviour and seed morphology: a missing link of myrmecochory. Oecologia 146, 244–246.
| Ant behaviour and seed morphology: a missing link of myrmecochory.Crossref | GoogleScholarGoogle Scholar |
Heterick, BE, and Shattuck, S (2011). Revision of the ant genus Iridomyrmex (Hymenoptera: Formicidae). Zootaxa 2845, 1–175.
| Revision of the ant genus Iridomyrmex (Hymenoptera: Formicidae).Crossref | GoogleScholarGoogle Scholar |
Hill, SJ, Silcocks, SC, and Andrew, NR (2020). Impacts of temperature on metabolic rates of adult Extatosoma tiaratum reared on different host plant species. Physiological Entomology 45, 7–15.
| Impacts of temperature on metabolic rates of adult Extatosoma tiaratum reared on different host plant species.Crossref | GoogleScholarGoogle Scholar |
Hughes, L, and Westoby, M (1990). Removal rates of seeds adapted for dispersal by ants. Ecology 71, 138–148.
| Removal rates of seeds adapted for dispersal by ants.Crossref | GoogleScholarGoogle Scholar |
Hughes, L, and Westoby, M (1992a). Capitula on stick insect eggs and elaiosomes on seeds: convergent adaptations for burial by ants. Functional Ecology 6, 642–648.
| Capitula on stick insect eggs and elaiosomes on seeds: convergent adaptations for burial by ants.Crossref | GoogleScholarGoogle Scholar |
Hughes, L, and Westoby, M (1992b). Effect of diaspore characteristics on removal of seeds adapted for dispersal by ants. Ecology 73, 1300–1312.
| Effect of diaspore characteristics on removal of seeds adapted for dispersal by ants.Crossref | GoogleScholarGoogle Scholar |
Hughes, L, Westoby, M, and Jurado, E (1994). Convergence of elaiosomes and insect prey: evidence from ant foraging behaviour and fatty acid composition. Functional Ecology 8, 358–365.
| Convergence of elaiosomes and insect prey: evidence from ant foraging behaviour and fatty acid composition.Crossref | GoogleScholarGoogle Scholar |
Kaspari, M (1996). Worker size and seed size selection by harvester ants in a neotropical forest. Oecologia 105, 397–404.
| Worker size and seed size selection by harvester ants in a neotropical forest.Crossref | GoogleScholarGoogle Scholar |
Key KHL (1991) Phasmatodea (stick-insects). In ‘The insects of Australia: a textbook for students and research workers, Vol. 1, Issue v. 2’. 2nd edn. (Ed. CSIRO) pp. 394–404. (Melbourne University Press). Available at https://books.google.com.au/books?id=xQyJnQAACAAJ
Kodela PG (2005) PlantNET – FloraOnline– Acacia longifolia. Available at https://plantnet.rbgsyd.nsw.gov.au/cgi-bin/NSWfl.pl?page=nswfl&lvl=sp&name=Acacia~longifolia
Leal, IR, Wirth, R, and Tabarelli, M (2007). Seed dispersal by ants in the semi-arid Caatinga of north-east Brazil. Annals of Botany 99, 885–894.
| Seed dispersal by ants in the semi-arid Caatinga of north-east Brazil.Crossref | GoogleScholarGoogle Scholar |
Leal, LC, Neto, MCL, de Oliveira, AFM, Andersen, AN, and Leal, IR (2014). Myrmecochores can target high-quality disperser ants: variation in elaiosome traits and ant preferences for myrmecochorous Euphorbiaceae n Brazilian Caatinga. Oecologia 174, 493–500.
| Myrmecochores can target high-quality disperser ants: variation in elaiosome traits and ant preferences for myrmecochorous Euphorbiaceae n Brazilian Caatinga.Crossref | GoogleScholarGoogle Scholar |
Lengyel, S, Gove, AD, Latimer, AM, Majer, JD, and Dunn, RR (2010). Convergent evolution of seed dispersal by ants, and phylogeny and biogeography in flowering plants: a global survey. Perspectives in Plant Ecology, Evolution and Systematics 12, 43–55.
| Convergent evolution of seed dispersal by ants, and phylogeny and biogeography in flowering plants: a global survey.Crossref | GoogleScholarGoogle Scholar |
Lubertazzi, D, Aliberti Lubertazzi, MA, McCoy, N, Gove, AD, Majer, JD, and Dunn, RR (2010). The ecology of a keystone seed disperser, the ant Rhytidoponera violacea. Journal of Insect Science 10, 158.
| The ecology of a keystone seed disperser, the ant Rhytidoponera violacea.Crossref | GoogleScholarGoogle Scholar |
McArthur AJ (2007) A key to Camponotus Mayr of Australia. In ‘Advances in ant systematics (Hymenoptera: Formicidae): homage to E. O. Wilson – 50 years of contributions’. (Eds RR Snelling, BL Fisher, PS Ward) pp. 290–351. (Memoirs of the American Entomological Institute 80)
Middleton, EJT, Reid, CR, Mann, RP, and Latty, T (2018). Social and private information influence the decision making of Australian meat ants (Iridomyrmex purpureus). Insectes Sociaux 65, 649–656.
| Social and private information influence the decision making of Australian meat ants (Iridomyrmex purpureus).Crossref | GoogleScholarGoogle Scholar |
Miller, CN, Whitehead, SR, and Kwit, C (2020). Effects of seed morphology and elaiosome chemical composition on attractiveness of five Trillium species to seed-dispersing ants. Ecology and Evolution 10, 2860–2873.
| Effects of seed morphology and elaiosome chemical composition on attractiveness of five Trillium species to seed-dispersing ants.Crossref | GoogleScholarGoogle Scholar |
Ness, JH, Bronstein, JL, Andersen, AN, and Holland, JN (2004). Ant body size predicts dispersal distance of ant-adapted seeds: implications of small-ant invasions. Ecology 85, 1244–1250.
| Ant body size predicts dispersal distance of ant-adapted seeds: implications of small-ant invasions.Crossref | GoogleScholarGoogle Scholar |
Ness, JH, Morin, DF, and Giladi, I (2009). Uncommon specialization in a mutualism between a temperate herbaceous plant guild and an ant: are Aphaenogaster ants keystone mutualists? Oikos 118, 1793–1804.
| Uncommon specialization in a mutualism between a temperate herbaceous plant guild and an ant: are Aphaenogaster ants keystone mutualists?Crossref | GoogleScholarGoogle Scholar |
O’Hanlon, JC, Hill, SJ, and Andrew, NR (2019). Using devitalised seeds in myrmecological research. Austral Entomology 58, 805–809.
| Using devitalised seeds in myrmecological research.Crossref | GoogleScholarGoogle Scholar |
O’Hanlon, JC, Jones, BR, and Bulbert, MW (2020). The dynamic eggs of the Phasmatodea and their apparent convergence with plants. The Science of Nature 107, 34.
| The dynamic eggs of the Phasmatodea and their apparent convergence with plants.Crossref | GoogleScholarGoogle Scholar |
Pettit, L, and Latty, T (2016). Greenhead ants Rhytidoponera metallica make trade-offs between food temperature and food concentration. Ecological Entomology 41, .
| Greenhead ants Rhytidoponera metallica make trade-offs between food temperature and food concentration.Crossref | GoogleScholarGoogle Scholar |
Pfeiffer, M, Huttenlocher, H, and Ayasse, M (2010). Myrmecochorous plants use chemical mimicry to cheat seed-dispersing ants. Functional Ecology 24, 545–555.
| Myrmecochorous plants use chemical mimicry to cheat seed-dispersing ants.Crossref | GoogleScholarGoogle Scholar |
Reifenrath, K, Becker, C, and Poethke, HJ (2012). Diaspore trait preferences of dispersing ants. Journal of Chemical Ecology 38, 1093–1104.
| Diaspore trait preferences of dispersing ants.Crossref | GoogleScholarGoogle Scholar |
Robertson, JA, Bradler, S, and Whiting, MF (2018). Evolution of oviposition techniques in stick and leaf insects (Phasmatodea). Frontiers in Ecology and Evolution 6, 216.
| Evolution of oviposition techniques in stick and leaf insects (Phasmatodea).Crossref | GoogleScholarGoogle Scholar |
Rocha, MLC, Cristaldo, PF, Cruz, JS, Sacramento, JJM, Ferreira, DV, and Araújo, APA (2018). Ants associated with Turnera subulata (Turneraceae): elaiosome attraction, seed dispersion and germination. Neotropical Entomology 47, 750–756.
| Ants associated with Turnera subulata (Turneraceae): elaiosome attraction, seed dispersion and germination.Crossref | GoogleScholarGoogle Scholar |
RStudio Team (2020) RStudio: Integrated Development for R. RStudio, PBC, Boston, MA. Available at http://www.rstudio.com/
Schneider, A, and Elgar, MA (2010). Facultative sex and reproductive strategies in response to male availability in the spiny stick insect, Extatosoma tiaratum. Australian Journal of Zoology 58, 228–233.
| Facultative sex and reproductive strategies in response to male availability in the spiny stick insect, Extatosoma tiaratum.Crossref | GoogleScholarGoogle Scholar |
Shattuck S (1999) ‘Australian ants.’ (CSIRO Publishing) Available at https://www.publish.csiro.au/book/503
Shelomi, M (2011). Phasmid eggs do not survive digestion by quails and chickens. Journal of Orthoptera Research 20, 159–162.
| Phasmid eggs do not survive digestion by quails and chickens.Crossref | GoogleScholarGoogle Scholar |
Stanton, AO, Dias, DA, and O’Hanlon, JC (2015). Egg dispersal in the Phasmatodea: convergence in chemical signaling strategies between plants and animals? Journal of Chemical Ecology 41, 689–695.
| Egg dispersal in the Phasmatodea: convergence in chemical signaling strategies between plants and animals?Crossref | GoogleScholarGoogle Scholar |
Tanaka, K, Ogata, K, Mukai, H, Yamawo, A, and Tokuda, M (2015). Adaptive advantage of myrmecochory in the ant-dispersed herb Lamium amplexicaule (Lamiaceae): predation avoidance through the deterrence of post-dispersal seed predators. PLoS ONE 10, e0133677.
| Adaptive advantage of myrmecochory in the ant-dispersed herb Lamium amplexicaule (Lamiaceae): predation avoidance through the deterrence of post-dispersal seed predators.Crossref | GoogleScholarGoogle Scholar |
Thomas, ML, and Framenau, VW (2005). Foraging decisions of individual workers vary with colony size in the greenhead ant Rhytidoponera metallica (Formicidae, Ectatomminae). Insectes Sociaux 52, 26–30.
| Foraging decisions of individual workers vary with colony size in the greenhead ant Rhytidoponera metallica (Formicidae, Ectatomminae).Crossref | GoogleScholarGoogle Scholar |
Turner, KM, and Frederickson, ME (2013). Signals can trump rewards in attracting seed-dispersing ants. PLOS ONE 8, e71871.
| Signals can trump rewards in attracting seed-dispersing ants.Crossref | GoogleScholarGoogle Scholar |
van Wilgenburg, EV, and Elgar, MA (2007). Colony structure and spatial distribution of food resources in the polydomous meat ant Iridomyrmex purpureus. Insectes Sociaux 54, 5–10.
| Colony structure and spatial distribution of food resources in the polydomous meat ant Iridomyrmex purpureus.Crossref | GoogleScholarGoogle Scholar |
Wickham H (2016) ‘Ggplot2.’ 2nd edn. (Springer)
Windsor, DM, Trapnell, DW, and Amat, G (1996). The egg capitulum of a neotropical walkingstick, Calynda bicuspis, induces aboveground egg dispersal by the ponerine ant, Ectatomma ruidum. Journal of Insect Behavior 9, 353–367.
| The egg capitulum of a neotropical walkingstick, Calynda bicuspis, induces aboveground egg dispersal by the ponerine ant, Ectatomma ruidum.Crossref | GoogleScholarGoogle Scholar |
Yamada, A, Bresseel, J, Chen, Z, Nguyen, AD, and Eguchi, K (2021). Deposition of phasmid eggs (Phasmatodea) in the nests of Acanthomyrmex glabfemoralis Zhou & Zheng, 1997 (Hymenoptera: Formicidae: Myrmicinae). Taiwania 66, 267–272.
| Deposition of phasmid eggs (Phasmatodea) in the nests of Acanthomyrmex glabfemoralis Zhou & Zheng, 1997 (Hymenoptera: Formicidae: Myrmicinae).Crossref | GoogleScholarGoogle Scholar |