

Do sulphur-crested cockatoos (Cacatua galerita) call the weather?
Marissa L. Gillies A and Culum Brown B *
B *
A
B
Abstract
Storms can have significant impacts on wildlife and many animals, including birds, can predict a storm’s arrival and change their behaviour accordingly. Storms are often associated with changes in environmental variables, particularly with a fall in barometric pressure. Social animals may collectively detect and advertise the potential onset of inclement weather to facilitate group responses. The present study tested the hypothesis that the sulphur-crested cockatoo uses environmental cues to predict the onset of storms and communicates this to conspecifics by emitting a specific ‘rain call’. Field observations were made over a four-month period at four locations in greater Sydney. Cockatoo calls were recorded, along with multiple environmental variables: barometric pressure, relative humidity, rainfall and temperature. We also noted the time of day relative to sunrise and sunset. We found that rain calls occurred throughout the day but were most prominent at dawn and were far more common at two of the four study sites. Rain calls were more likely to occur at slightly lower temperatures, during periods of high humidity and if it was currently raining. We found no evidence that the calls were prompted by current atmospheric pressure, but the observation period did not contain many storms.
Keywords: animal communication, barometric pressure, cockatoo, dawn, humidity, storms, sociality, weather.
Introduction
Extreme weather events, such as drought, heatwaves and heavy rainfall, can negatively impact birds’ abundance (Haslem et al. 2015), food supply (Cameron 2009), migration (Lindenmayer et al. 2019), reproduction (McDonald et al. 2004) and mortality rate (Saunders et al. 2011; McKechnie et al. 2012; Conradie et al. 2020). Severe storms can be life-threatening, thus it is beneficial for birds to predict the weather and adjust their behaviour to suit (Cohen et al. 2021). Lower temperatures, rainfall and wind increase metabolic expenditure, which negatively affects their body condition and ability to incubate eggs (Breuner et al. 2013; Boyer and MacDougall-Shackleton 2020). Inclement weather coping responses include increasing foraging activity (Metcalfe et al. 2013; Boyer and MacDougall-Shackleton 2020), nest-maintenance behaviour (Laux et al. 2016), migration (Boyle et al. 2010), and abandonment of territories (Streby et al. 2015). Many of these behaviours occur just prior to an extreme weather event, which suggests that birds can predict the coming weather and prepare for it.
Birds have the unique ability to detect small changes in barometric pressure (von Bartheld and Giannessi 2011). The paratympanic organ (also known as vitali organ) is a sensory receptor found in the middle ear (Bender and Hartman 2015) which functions as an internal barometer and altimeter (von Bartheld and Giannessi 2011). As storms are associated with a decrease in barometric pressure, the paratympanic organ allows birds to predict their onset (von Bartheld and Giannessi 2011; Breuner et al. 2013; Metcalfe et al. 2013). Evidence from correlational field studies shows birds use barometric pressure as an environmental cue to prepare for storms. When barometric pressure decreased before a storm, the Canadian ferruginous hawk (Buteo regalis) increased its nest maintenance behaviours 12–24 h beforehand (Laux et al. 2016). Temperature, wind speed and rain were not associated with increased nest-maintenance behaviour (Laux et al. 2016). Experimental studies that manipulated barometric pressure in a laboratory confirmed that birds alter their behaviour when barometric pressure decreases. White-crowned sparrows (Zonotrichia leucophrys), for instance, increased food intake and foraging when barometric pressure was lowered regardless of temperature changes (Breuner et al. 2013; Metcalfe et al. 2013). Taken together, these results support the theory that birds use barometric pressure, independent of other storm-related environmental cues, to forecast and prepare for imminent storms.
Social animals face situations where they must make adaptive responses to environmental challenges that balance the costs and benefits of group living; this may be achieved through collective decision making (Ward and Webster 2016). Collective decisions help maintain group cohesion and enhance the fitness of group members, such as deciding where and when to forage, nest or take shelter (Silk et al. 2014). Social groups share information which can help generate group consensus and synchronise behaviour. Sharing information about impending storms may allow group members to take appropriate action such as taking shelter or increasing food intake. It is possible that group living birds may collectively detect declining barometric pressure and communicate this among group members using a specific call. Birds are known to use acoustic functionally referential signals most commonly for the discovery of food or predators (Ausmus and Clarke 2014; Grieves et al. 2014; Farrow et al. 2017). Golden Sebright bantams (Gallus gallus domesticus), for example, have different alarm calls for two types of predators, flying raptors and racoons (Procyon lotor) (Evans et al. 1993). Birds can communicate specific details through alarm calls, such as predator behaviour. Siberian jays (Perisoreus infaustus) use different alarm calls for hawks who were perched, attacking, or searching for prey, each representing a different category of threat, which, in turn, elicited appropriate escape responses in conspecifics (Griesser 2008).
Parrots (order Psittaciformes), like humans and passerines, are vocal learners and have complex vocal behaviour (Marler 1970). Research shows that the social environment is a key driver for such vocal plasticity (Ali et al. 1993). Experimental studies found that hand-reared, captive parrots can use functionally referential vocalisations (Bottoni et al. 2009; Giret et al. 2012), and some species are capable of learning referential human speech (Pepperberg 1999; Giret et al. 2010). In the wild, parrots use acoustic communication to signal danger and food availability, to enhance flight and flock cohesion through contact calls, to advertise social status, and to facilitate individual recognition (Saunders 1983; Ali et al. 1993; Bradbury 2003; Marler 2004). Contact calls can also have geographic variation between populations of the same species (Wright and Dahlin 2018). Multiple studies have shown that some calls are used in specific contexts. For example, a study of the lilac-crowned amazon (Amazona finschi) found that its vocalisations had context specificity across nine contexts (Montes-Medina et al. 2016) and the palm cockatoo (Probosciger aterrimus) has 27 vocalisations used across five behavioural contexts (Zdenek et al. 2015).
Sulphur-crested cockatoos (Cacatua galerita) originally lived in forests but have adapted to the changing environment and now thrive in urban landscapes. They are arguably one of the most conspicuous cockatoo species in Australia. Cockatoos are adept at extractive foraging and problem solving, which facilitates dietary flexibility (Juniper and Parr 2003; Magat and Brown 2009; O’Hara et al. 2019). In Sydney, they open household bins to retrieve food, a skill that has spread through social learning (Klump et al. 2021). Like other long-lived parrots with advanced cognition, their social structures require a complex communication system (Freeberg et al. 2012; Kaplan 2014; Wirthlin et al. 2018). Sulphur-crested cockatoos live in a fission–fusion social system, where they forage with different individuals during the day and regroup at the roosting site at night using contact calls (Lindenmayer et al. 1996; Aplin et al. 2021). Communal roosting facilitates information sharing about foraging effectiveness and social dynamics (Juniper and Parr 2003). There is evidence of sentinel behaviour when flocks of eight or more forage on the ground (Higgins 1999; Moore 2021). Surprisingly, there have been few studies on their vocal repertoire. They are most vocal when flocks gather at roosting sites at dusk and dawn, and quieter when feeding or resting during the day and have a series of distinct vocalisations (Noske 1980). Sulphur-crested cockatoos’ unmistakable contact call is often used in flight with an upward inflection at the end. They use variations of their alarm call when threatened or startled, or for antipredator behaviour; alarm calls are abrupt guttural screeches and change with intensity depending on the threat level. Some of their calls occur in the context of sharing location, distress, change of mood, begging for food (by juveniles), and eating (by fledglings); however, the purpose of many calls remains unknown (Noske 1980).
Here we examined if sulphur-crested cockatoos have a context-specific vocalisation they use for approaching storms. Casual observations suggested that they emit a specific ‘rain call’ just prior to the onset of inclement weather. We predicted that if sulphur-crested cockatoos’ sensitivity to barometric pressure permits them to detect the onset of storms, then a specific ‘rain call’ vocalisation should increase in frequency when barometric pressure is declining. To address this question, wild sulphur-crested cockatoos’ vocalisations were recorded at four locations in Sydney during various weather conditions, over a 4-month period. Call types were identified and catalogued to enable us to determine if rain calls were associated with weather variables, such as barometric pressure, temperature, humidity, and precipitation.
Method
Procedure
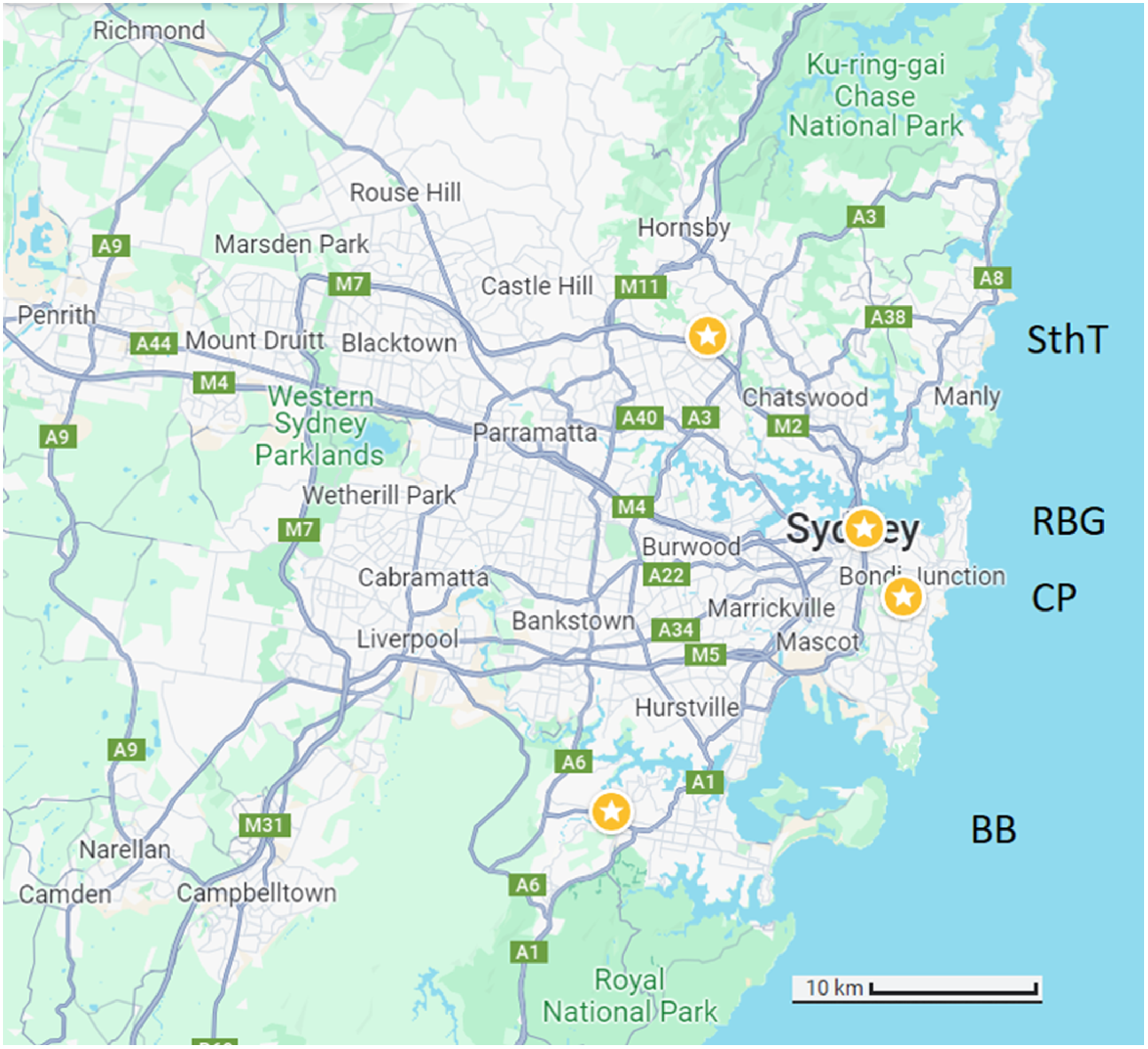
The sulphur-crested cockatoo (C. galerita) is a large (up to 1 kg), stocky, white parrot with a yellow crest (Saunders 2009). It occurs in eastern and northern Australia, Indonesia and Papua New Guinea, across a wide range of habitats (Juniper and Parr 2003). Field observations were conducted over 84 days from April to November in 2021, including part of their breeding season (Heinsohn et al. 2003). Most recordings were made between 0600 and 1100 hours and between 1600 and 1900 hours when the birds are most vocal (Noske 1980), but representative recordings were attained over the full 24-h period. Observations were made at four locations in Sydney, Australia, that included suburban bushland and urban environments. These sites were selected as sulphur-crested cockatoos are frequently observed foraging, resting and roosting at each location: South Turramurra (33.75°S, 151.10°E), which borders Lane Cove National Park; Bonnet Bay (34.02°S, 151.05°E), on the eastern bank of the Woronora River surrounded by bushland; Centennial Parklands (33.89°S, 151.24°E), a 360 ha park; and Royal Botanic Gardens (33.86°S, 151.22°E), a 30 ha park with a well-studied population of sulphur-crested cockatoos (Aplin et al. 2021; Fig. 1). It is worth noting that the number of observation periods varied between locations: Bonnet Bay (n = 43), Royal Botanic Garden (n = 16), South Turramurra (n = 208), and Centennial Park (n = 29). Most observations were made in South Turramurra because it is adjacent to the university campus. We opportunistically sampled the other locations.
Temperature (°C), relative humidity and time of sunrise and sunset were obtained from the Bureau of Meteorology (BoM 2021). Barometric pressure (hPa), and its activity (rising, steady or falling) were recorded using a smartphone application that measures air pressure using the device’s pressure sensor. Here we used the iPhone12 whose sensor has a standard deviation of 0.0093 hPa. We also noted if it was raining or not at the time of observation. It was unseasonably dry at the time of the study and rain was recorded in only 32 of the observation periods.
For each session, the current time and times of sunset and sunrise were recorded. Time of observation was then categorised relative to sunset and sunrise: dawn (30 min before or after sunrise), morning (30 min after sunrise to 1200 hours), afternoon (1200 hours to 30 min before sunset), dusk (30 min before or after sunset), and night (30 min after sunset to 30 min before sunrise).
Vocalisations of free-living sulphur-crested cockatoos were recorded in 10-min sessions at random times throughout the day, in a range of behavioural contexts (resting, foraging, roosting, agonistic, or human interaction) and group sizes. Behaviour was sometimes noted if the sulphur-crested cockatoos were visible, but most often their calls were heard from a distance and behaviour could not be noted. Audio recordings were made on an iPhone 12 with the Voice application. We collected recordings from the same position at each location, no attempt to follow flocks was made and, although birds were nearly always present at each location, they may not have been vocal. In total, 50 h of recordings were made and reviewed by ear and visual inspection of spectrograms. Rain or croak calls were heard in 127 of the 297 10-min sampling intervals. Calls were then categorised, guided by previous studies (Noske 1980; Bradbury 2003), and a description and spectrogram was made for each call. From this call library, two calls were selected for further analysis, the rain call (Supplementary Fig. S1) and croak call (Fig. S2). The frequency of the rain call and croak call was recorded for each session. As the rain call was the focus of the study, the croak call was selected for comparison (control), as it sounds notably different from more common flight or contact calls.
We hypothesised that the rain call would be associated with weather variables. Specifically, we expected that if barometric pressure was declining, then the frequency of the rain call would increase. We analysed two dependent variables: the frequency of rain calls and croak calls within the 10-min observation windows. Given the large variation in frequency scores and zero inflated data set, we transformed frequency into a binomial score (present or absent). There were six independent variables: location, precipitation (present or absent), temperature (°C), relative humidity (%), time of day (night, dawn, morning, afternoon and dusk), and barometric pressure (hPa) activity (rising, falling and steady).
We employed a generalised linear model with a binomial error distribution to analyse the call data. The model included call frequency (binomial) as the dependent variable, with temperature and humidity as covariates, and time of day, location, and barometric pressure activity as independent variables. The initial model included 2-way interactions, but none of these interactions were significant and were subsequently removed. For the rain call data, temperature had no influence in the model and its removal improved model fit as indicated by BIC. For the croak call data, the best model retained temperature but location was removed.
Results
Rain calls
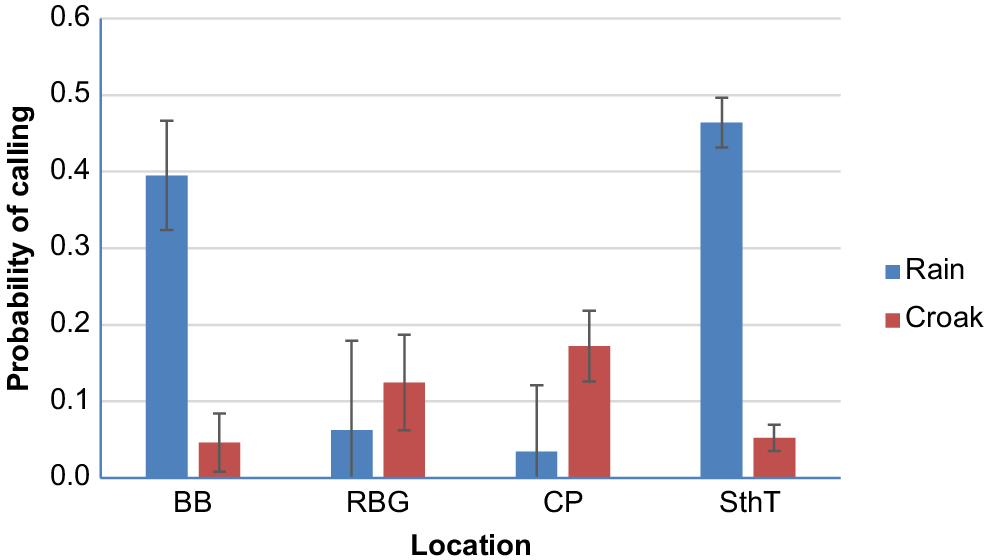
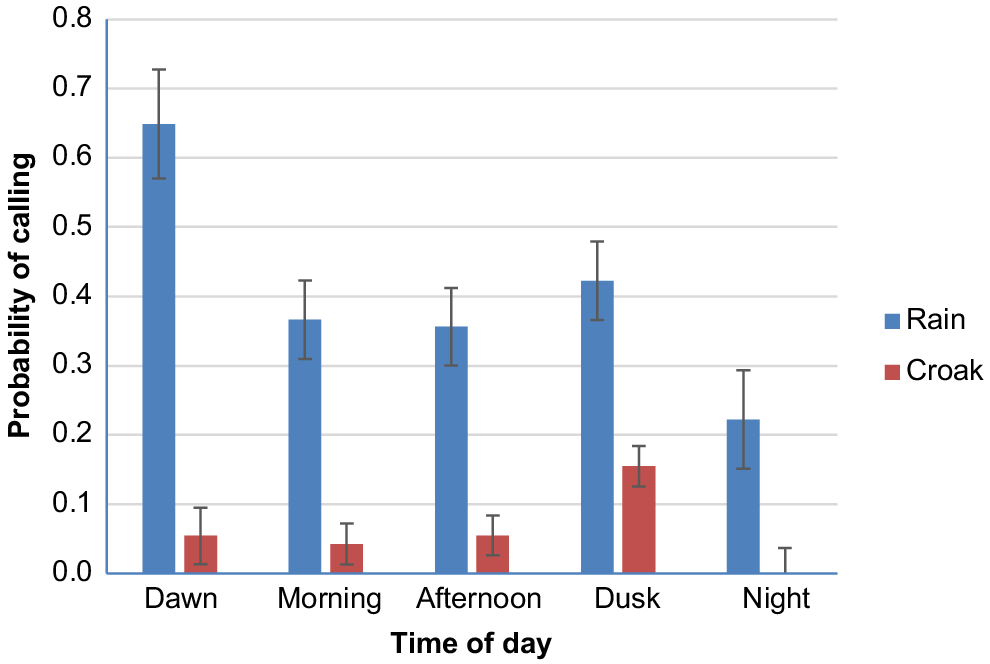
The model for rain calls suggests that probability that the cockatoos made calls significantly varied with time of day and between locations (Table 1). Rain calls were most likely to occur at South Turramurra followed by Bonnet Bay but they were reasonably rare at Centennial Park and the Botanic Gardens (Fig. 2) (post hoc pair-wise comparisons show BB and SthT were significantly higher than BG and CP, P < 0.003 in all cases). Rain calls were most likely to occur at dawn and least likely to occur at night (Fig. 3). By comparison, the probability of cockatoos making croak calls did not vary significantly between locations (location did not feature in the best fit model) nor time of day, although they were slightly more likely to occur at dusk (Fig. 3) (W = 8.067, d.f. = 4, P = 0.089).
| Wald Chi-Square | d.f. | P | ||
|---|---|---|---|---|
| (Intercept) | 12.107 | 1 | <0.001 | |
| ToDay | 22.996 | 4 | <0.001 | |
| Precipitation | 4.782 | 1 | 0.029 | |
| Humidity | 6.509 | 1 | 0.011 | |
| hPaActivity | 1.525 | 2 | 0.467 | |
| Location | 15.550 | 3 | 0.001 |
ToDay, time of day; hPaActivity, rising, falling or steady air pressure.
Mean (s.e.) probability that cockatoos were calling at four different locations. BB, Bonnet Bay; RBG, Royal Botanic Gardens; CP, Centennial Park; SthT, South Turramurra.
The mean (s.e.) probability of cockatoos making rain or croak calls at different times of the day.
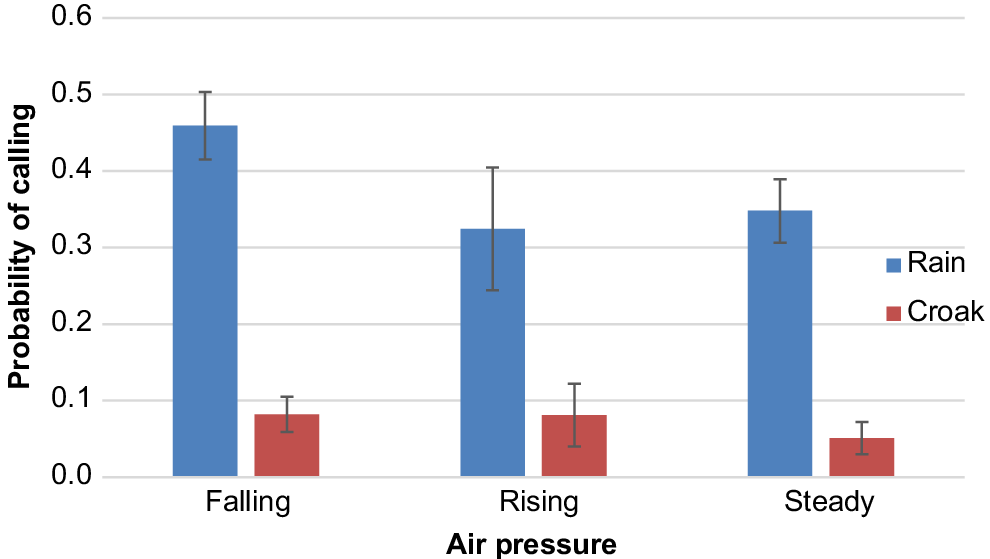
Cockatoos were more likely to make rain calls if it was raining at the time of the observation (59% versus 37%: Table 1) but this was not the case with croak calls (W = 0.528, d.f. = 1, P = 0.467). There was no significant effect of atmospheric pressure activity, with both calls equally likely to occur irrespective of whether atmospheric pressure was rising, falling or steady. However, there was a slight tendency for rain calls to be more likely if barometric pressure was falling (Fig. 4).
Mean probability (±s.e.) of making calls when air pressure was falling, rising or steady at the time of observation.
The probability of cockatoos making rain calls was significantly positively correlated with relative humidity (Table 1); relative humidity was higher (~75%) when the cockatoos were making rain calls than when they were not calling (~64%). The probability of making croak calls, in contrast, was not affected by humidity (W = 2.59, d.f. = 2, P = 0.108).
For the rain calls model, temperature was not included in the best model, but for croak calls temperature was a positive predictor of the probability of cockatoos making calls. The temperature was higher (16.3°C) when cockatoos were making croak calls than when they were not (14.3°C) (W = 4.544, d.f. = 1, P = 0.033).
Discussion
Despite the ubiquity of sulphur-crested cockatoos and their distinct calls, there has been limited research on their vocalisations. The aim of this research was to investigate if sulphur-crested cockatoos use a context-specific call (the rain call) to signal oncoming storms. A drop in barometric pressure can occur up to 24 h before a storm and many animals, including birds, use environmental cues to predict the weather and change their behaviour accordingly (Saunders and Derebeira 1986; Breuner et al. 2013; Metcalfe et al. 2013). Here we hypothesised that sulphur-crested cockatoos could detect decreases in barometric pressure and communicate this to the rest of the flock with a rain call. For comparison, we also analysed call frequency data from a call that did not appear to be related to storms a priori – the croak call. In addition to examining the effects of shifts in barometric pressure, we also analysed other key environmental cues including temperature, humidity and precipitation. We found that the probability of making rain calls was higher if it was raining at the time of observation, and rain calls were more likely at higher humidity. Contrary to our hypothesis, however, atmospheric pressure seemed to have little predictive value, although rain calls were slightly more likely to occur if pressure was falling. In contrast, croak calls were only influenced by temperature. The probability of making rain calls also varied between the four study sites and with the time of day, whereas the croak call was not significantly influenced by time of day or location.
The catalogue of sulphur-crested cockatoo vocalisations created by this study predominantly matched common call types found in wild parrots from previous studies. The present study identified 17 calls with variations whereas Noske (1980) documented 15 calls plus variations, and there is considerable overlap between the two studies. Here we identified and focused on a new call which we labelled the ‘rain call’. This call was most often heard in quick succession when the sulphur-crested cockatoos were in flocks. The rain call was associated with higher relative humidity and was more likely to occur if it was raining at the time of the observation; thus, rain call may be an appropriate name. There was also a hint that this call may be more likely to occur with declining atmospheric pressure, but more work needs to be done to clarify this.
In the present study, a number of climatic variables were associated with call probability. Unexpectedly, barometric pressure activity was not significantly associated with the rain call. Previous studies found that birds use a decrease in barometric pressure as an environmental cue to predict storms. When barometric pressure decreases, hawk nest maintenance behaviours increase (Laux et al. 2016), as does sparrow foraging behaviour (Breuner et al. 2013; Metcalfe et al. 2013). These changes in behaviour help birds prepare for challenging weather and clearly have fitness value. One of the possible explanations for the lack of an overall effect in the present study is that the period of observation was unusually dry and few storm events occurred. It rained during only 11% of our observation periods and barometric pressure remained reasonably constant (mean 1008, range 985–1028 hPa). East coast lows are usually reasonably common in south-east Australia and usually occur several times each year, bringing heavy rainfall and strong winds but none occurred during the study period. Future studies should ensure more severe storms are captured in the data set.
Interestingly, precipitation and humidity were both positively associated with the rain call, but not the croak call, whereas croak calls were more likely at higher temperatures. These associations are not overly surprising based on previous research (Metcalfe et al. 2013; Laux et al. 2016), but the fact that the rain call was so closely linked with rain and humidity gives further credence to its association with poor weather. During light rain events, the sulphur-crested cockatoos were observed sitting quietly in trees, continuing to forage, or bathing while hanging upside down and flapping their wings; this behaviour has been reported in previous studies (Noske 1980). For cockatoos, light rainfall may be a pleasant event, as rain-bathing is important for feather maintenance (Murphy et al. 2011). In contrast, big storms can present serious challenges to birds and, collectively, the results of this study suggest it is probable that the cockatoos can use environmental cues to predict poor weather but further research is needed.
The sulphur-crested cockatoos were most vocal at dusk and dawn at their communal roosts, as observed in previous studies (Noske 1980). Specifically, rain calls were most common at dawn whereas croak calls were distributed across all times of the day. The cockatoos disperse into smaller flocks to forage during the day then regroup at dusk. Flock size is larger at dusk and dawn, and communal roosts play an important part in communication (Juniper and Parr 2003). Given the hypothesised role of the rain call as a social warning that poor weather is imminent, it makes sense that it would be more prominent when flock sizes are larger. The rain call may be used for collective decision making, where social animals make adaptive responses to environmental changes (Ward and Webster 2016). Parrots are known to make collective decisions in other contexts, such as where the group will forage, nest, or take shelter (Silk et al. 2014).
It is interesting to note that the rain call occurred most frequently at South Turramurra (SthT) and Bonnet Bay (BB), with very few calls at Royal Botanic Garden (RBG) and Centennial Park (CP). The rain call occurred in only about 5% of observations at the RBG and CP, both inner city locations, whereas the calls occurred in more than 40% of observations at the other two locations. There are several possible explanations for this observation. One possibility is that these locations vary in important climatic variables; the locations where the rain call was recorded most frequently are both suburban with surrounding bush land, compared to inner city parks surrounded by built-up environments. Parrot abundance can vary across urban landscapes and studies in Sydney suggest that urban parrots may be more abundant in locations with remnant vegetation, particularly during the breeding season (Davis et al. 2012). South Turramurra also has the highest elevation of all study sites (184 m) compared to Bonnet Bay (50 m), Centennial Park (38 m) and Royal Botanic Gardens (15 m) (BoM 2021). Moreover, SthT generally has one of the highest rainfall totals for Sydney, although this was not the case during the study period; nevertheless, humidity was significantly higher at SthT than at all other locations. However, the remaining three locations did not differ in humidity, so clearly climatic variables alone do not explain the variation in rain calls we observed.
Finally, it is interesting to note that although rain calls were more likely to occur while it was raining (and correspondingly high humidity), they still occurred during observation periods when it was not raining, albeit at only 37% of the time. Nonetheless, this does suggest that the rain call is not exclusively used during rainy periods and may be used in a slightly broader context perhaps to signal discomfort or dissatisfaction with their current state. Although we made some notes on the behavioural context of the calls, this was reasonably rare as most of the birds were out of sight while calling. Future studies should attempt to identify the social context in which the calls are made and use targeted individual follows to pay more attention to the behaviour of individuals while calling.
In conclusion, the present study built upon previous research on the vocal repertoire of the sulphur-crested cockatoo by generating a vocalisation library and documenting a new call – the rain call. Our data showed that this call increased with relative humidity and was more common if it was raining at the time of the observation. While not statistically significant, we also found a trend that rain calls were slightly more likely to occur during declining barometric pressure although more research is needed during strong storms. The rain call was observed frequently at some locations but not others, though it is unclear why. Taken together, these findings suggest that sulphur-crested cockatoos may have population specific calls, that are used in certain contexts or prompted by environmental cues – poor weather. Further research is needed into functionally referential communication capability in sulphur-crested cockatoos. Parrots are highly social species, they predominantly use acoustic communication and are vocal learners, which makes them ideal subjects for studying animal communication.
Data availability
All data used to generate the results in the paper is available on reasonable request to the corresponding author.
References
Ali NJ, Farabaugh S, Dooling R (1993) Recognition of contact calls by the budgerigar (Melopsittacus undulatus). Bulletin of the Psychonomic Society 31(5), 468-470.
| Crossref | Google Scholar |
Aplin LM, Major RE, Davis A, Martin JM (2021) A citizen science approach reveals long-term social network structure in an urban parrot, Cacatua galerita. Journal of Animal Ecology 90(1), 222-232.
| Crossref | Google Scholar | PubMed |
Ausmus DM, Clarke JA (2014) Mother knows best: functionally referential alarm calling in white-tailed ptarmigan. Animal Cognition 17(3), 671-679.
| Crossref | Google Scholar | PubMed |
Bender MJ, Hartman GD (2015) Bat activity increases with barometric pressure and temperature during autumn in central Georgia. Southeastern Naturalist 14(2), 231-242.
| Crossref | Google Scholar |
BoM (2021) Bonnet Bay weather. Available at http://www.bom.gov.au/places/nsw/bonnet-bay/
Bottoni L, Masin S, Lenti-Boero D (2009) Vowel-like sound structure in an African grey parrot (Psittacus erithacus) vocal production. The Open Behavioral Science Journal 3(1), 1-16.
| Crossref | Google Scholar |
Boyer AC, MacDougall-Shackleton SA (2020) High rates of exposure to simulated winter storm cues negatively affect white-throated sparrow (Zonotrichia albicollis) energy reserves. Frontiers in Ecology and Evolution 8, 222.
| Crossref | Google Scholar |
Boyle WA, Norris DR, Guglielmo CG (2010) Storms drive altitudinal migration in a tropical bird. Proceedings of the Royal Society B: Biological Sciences 277(1693), 2511-2519.
| Crossref | Google Scholar |
Breuner CW, Sprague RS, Patterson SH, Woods HA (2013) Environment, behavior and physiology: do birds use barometric pressure to predict storms? Journal of Experimental Biology 216(11), 1982-1990.
| Crossref | Google Scholar |
Cameron M (2009) The influence of climate on glossy black-cockatoo reproduction. Pacific Conservation Biology 15(1), 65-71.
| Crossref | Google Scholar |
Cohen JM, Fink D, Zuckerberg B (2021) Extreme winter weather disrupts bird occurrence and abundance patterns at geographic scales. Ecography 44, 1143-1155.
| Crossref | Google Scholar |
Conradie SR, Woodborne SM, Wolf BO, Pessato A, Mariette MM, McKechnie AE (2020) Avian mortality risk during heat waves will increase greatly in arid Australia during the 21st century. Conservation Physiology 8(1), coaa048.
| Crossref | Google Scholar |
Davis A, Taylor CE, Major RE (2012) Seasonal abundance and habitat use of Australian parrots in an urbanised landscape. Landscape and Urban Planning 106(2), 191-198.
| Crossref | Google Scholar |
Evans CS, Macedonia JM, Marler P (1993) Effects of apparent size and speed on the response of chickens, Gallus gallus, to computer-generated simulations of aerial predators. Animal Behaviour 46(1), 1-11.
| Crossref | Google Scholar |
Farrow LF, Doohan SJ, McDonald PG (2017) Alarm calls of a cooperative bird are referential and elicit context-specific antipredator behavior. Behavioral Ecology 28(3), 724-731.
| Crossref | Google Scholar |
Freeberg TM, Dunbar RIM, Ord TJ (2012) Social complexity as a proximate and ultimate factor in communicative complexity. Philosophical Transactions of the Royal Society B: Biological Sciences 367(1597), 1785-1801.
| Crossref | Google Scholar |
Giret N, Péron F, Lindová J, Tichotová L, Nagle L, Kreutzer M, Tymr F, Bovet D (2010) Referential learning of French and Czech labels in African grey parrots (Psittacus erithacus): different methods yield contrasting results. Behavioural Processes 85(2), 90-98.
| Crossref | Google Scholar |
Giret N, Albert A, Nagle L, Kreutzer M, Bovet D (2012) Context-related vocalizations in African grey parrots (Psittacus erithacus). Acta Ethologica 15(1), 39-46.
| Crossref | Google Scholar |
Griesser M (2008) Referential calls signal predator behavior in a group-living bird species. Current Biology 18(1), 69-73.
| Crossref | Google Scholar | PubMed |
Grieves LA, Logue DM, Quinn JS (2014) Joint-nesting smooth-billed anis, Crotophaga ani, use a functionally referential alarm call system. Animal Behaviour 89, 215-221.
| Crossref | Google Scholar |
Haslem A, Nimmo DG, Radford JQ, Bennett AF (2015) Landscape properties mediate the homogenization of bird assemblages during climatic extremes. Ecology 96(12), 3165-3174.
| Crossref | Google Scholar | PubMed |
Heinsohn R, Murphy S, Legge S (2003) Overlap and competition for nest holes among eclectus parrots, palm cockatoos and sulphur-crested cockatoos. Australian Journal of Zoology 51(1), 81.
| Crossref | Google Scholar |
Kaplan G (2014) Animal communication. WIREs Cognitive Science 5(6), 661-677.
| Crossref | Google Scholar | PubMed |
Klump BC, Martin JM, Wild S, Hörsch JK, Major RE, Aplin LM (2021) Innovation and geographic spread of a complex foraging culture in an urban parrot. Science 373, 456-460.
| Crossref | Google Scholar | PubMed |
Laux CM, Nordell CJ, Fisher RJ, Ng JW, Wellicome TI, Bayne EM (2016) Ferruginous hawks Buteo regalis alter parental behaviours in response to approaching storms. Journal of Ornithology 157(1), 355-362.
| Crossref | Google Scholar |
Lindenmayer DB, Pope MP, Cunningham RB, Donnelly CF, Nix HA (1996) Roosting of the sulphur-crested cockatoo Cacatua galerita. Emu - Austral Ornithology 96(3), 209-212.
| Crossref | Google Scholar |
Lindenmayer DB, Lane P, Foster CN, Westgate MJ, Sato C, Ikin K, Crane M, Michael D, Florance D, Scheele BC (2019) Do migratory and resident birds differ in their responses to interacting effects of climate, weather and vegetation? Diversity and Distributions 25, 449-461.
| Crossref | Google Scholar |
Magat M, Brown C (2009) Laterality enhances cognition in Australian parrots. Proceedings of the Royal Society B: Biological Sciences 276(1676), 4155-4162.
| Crossref | Google Scholar |
Marler P (1970) A comparative approach to vocal learning: song development in white-crowned sparrows. Journal of Comparative and Physiological Psychology 71(2, Pt.2), 1-25.
| Crossref | Google Scholar |
Marler P (2004) Bird calls: their potential for behavioral neurobiology. Annals of the New York Academy of Sciences 1016(1), 31-44.
| Crossref | Google Scholar |
McDonald PG, Olsen PD, Cockburn A (2004) Weather dictates reproductive success and survival in the Australian brown falcon Falco berigora. Journal of Animal Ecology 73(4), 683-692.
| Crossref | Google Scholar |
McKechnie AE, Hockey PAR, Wolf BO (2012) Feeling the heat: Australian landbirds and climate change. Emu - Austral Ornithology 112(2), i-vii.
| Crossref | Google Scholar |
Metcalfe J, Schmidt KL, Bezner Kerr W, Guglielmo CG, Macdougall-Shackleton SA (2013) White-throated sparrows adjust behaviour in response to manipulations of barometric pressure and temperature. Animal Behaviour 86(6), 1285-1290.
| Crossref | Google Scholar |
Montes-Medina AC, Salinas-Melgoza A, Renton K (2016) Contextual flexibility in the vocal repertoire of an Amazon parrot. Frontiers in Zoology 13(1), 40.
| Crossref | Google Scholar |
Moore GM (2021) Harvesting of onion grass, Romulea rosea L. by sulphur-crested cockatoo, Cacatua galerita Latham. Ecological Management & Restoration 22(1), 40-44.
| Crossref | Google Scholar |
Murphy SM, Braun JV, Millam JR (2011) Bathing behavior of captive orange-winged Amazon parrots (Amazona amazonica). Applied Animal Behaviour Science 132(3-4), 200-210.
| Crossref | Google Scholar |
Noske S (1980) Aspects of the behaviour and ecology of the white cockatoo (‘Cacatua galerita’) and galah (‘C. roseicapilla’) in croplands in north-east New South Wales. Available at https://hdl.handle.net/1959.11/19443
O’Hara M, Mioduszewska B, Haryoko T, Prawiradilaga DM, Huber L, Auersperg A (2019) Extraction without tooling around – the first comprehensive description of the foraging- and socio-ecology of wild Goffin’s cockatoos (Cacatua goffiniana). Behaviour 156(5–8), 661-690.
| Crossref | Google Scholar |
Saunders DA (1983) Vocal repertoire and individual vocal recognition in the short-billed form of the white-tailed black cockatoo, Calyptorhynchus funereus latirostris Carnaby. Australian Wildlife Research 10(3), 527-536.
| Crossref | Google Scholar |
Saunders DA, Derebeira CP (1986) Seasonal occurrence of members of the suborder Charadrii (waders or shorebirds) on Rottnest Island, Western-Australia. Australian Wildlife Research 13(2), 225-244.
| Crossref | Google Scholar |
Saunders DA, Mawson P, Dawson R (2011) The impact of two extreme weather events and other causes of death on Carnaby’s black cockatoo: a promise of things to come for a threatened species? Pacific Conservation Biology 17(2), 141.
| Crossref | Google Scholar |
Silk MJ, Croft DP, Tregenza T, Bearhop S (2014) The importance of fission–fusion social group dynamics in birds. Ibis 156(4), 701-715.
| Crossref | Google Scholar |
Streby HM, Kramer GR, Peterson SM, Lehman JA, Buehler DA, Andersen DE (2015) Tornadic storm avoidance behavior in breeding songbirds. Current Biology 25(1), 98-102.
| Crossref | Google Scholar | PubMed |
von Bartheld CS, Giannessi F (2011) The paratympanic organ: a barometer and altimeter in the middle ear of birds? Journal of Experimental Zoology Part B: Molecular and Developmental Evolution 316B(6), 402-408.
| Crossref | Google Scholar |
Wirthlin M, Lima NCB, Guedes RLM, Soares AER, Almeida LGP, Cavaleiro NP, Loss De Morais G, Chaves AV, Howard JT, Teixeira MdM, Schneider PN, Santos FR, Schatz MC, Felipe MS, Miyaki CY, Aleixo A, Schneider MPC, Jarvis ED, Vasconcelos ATR, Prosdocimi F, Mello CV (2018) Parrot genomes and the evolution of heightened longevity and cognition. Current Biology 28(24), 4001-4008.e7.
| Crossref | Google Scholar |
Wright TF, Dahlin CR (2018) Vocal dialects in parrots: patterns and processes of cultural evolution. Emu - Austral Ornithology 118(1), 50-66.
| Crossref | Google Scholar |
Zdenek CN, Heinsohn R, Langmore NE (2015) Vocal complexity in the palm cockatoo (Probosciger aterrimus). Bioacoustics 24(3), 253-267.
| Crossref | Google Scholar |