

Re-introducing fire in sagebrush steppe experiencing decreased fire frequency: does burning promote spatial and temporal heterogeneity?
Kirk W. Davies A B and Jon D. Bates AA USDA Agricultural Research Service, 67826-A Highway 205, Burns, OR 97720, USA.
B Corresponding author. Email: kirk.davies@oregonstate.edu
International Journal of Wildland Fire 29(8) 686-695 https://doi.org/10.1071/WF20018
Submitted: 4 February 2020 Accepted: 18 April 2020 Published: 7 May 2020
Journal Compilation © IAWF 2020 Open Access CC BY-NC-ND
Abstract
Fire frequency has decreased in many shrub-steppe communities. Re-introducing fire may be needed to increase spatial and temporal variability in vegetation, but is often hindered by concerns of undesired vegetation shifts. These concerns arise, in part, because long-term effects of fire re-introduction in these communities after prolonged fire exclusion and other departures from historical conditions are unknown. To better understand the effects of re-introducing fire, we evaluated plant community response to re-introducing fire for 12 years post fire in six mountain big sagebrush communities. Herbaceous biomass production was 1.7-fold greater in burned compared with unburned areas at the conclusion of the study. Exotic annual grasses appeared to be problematic in the first 8 years post fire, but became inconsequential (~1% cover) by the end of the study. Re-introducing fire promoted other shrubs (excluding sagebrush) that were probably inhibited by competition from sagebrush. Sagebrush cover and density remained low in burned areas for the duration of the study, because of limited recruitment in the years immediately post fire and competition from herbaceous vegetation. Re-introducing fire appears to increase temporal and spatial heterogeneity in shrub-steppe communities experiencing prolonged fire exclusion and, therefore, may be needed to maintain a diversity of plant communities.
Additional keywords: Artemisia tridentata, disturbance, fire management, mountain big sagebrush, prescribed burning.
Introduction
Historical disturbance regimes created a diversity of habitats spatially and temporally and influenced the formation of major land cover types (Turner 2010). These disturbance regimes shaped plant communities and, subsequently, native fauna communities. Humans have vastly altered disturbance regimes around the world, including the exclusion of historical disturbances (Turner 2010). Re-introducing historical disturbances may be needed to maintain specific native plant communities or, alternatively, may create novel plant communities because invasive species, climate change and elevated atmospheric CO2 concentrations have altered how plant communities respond to disturbances (Williams and Jackson 2007; Davies et al. 2009; Archer et al. 2011). Managers of ecological systems face uncertainties when re-introducing historical disturbances in today’s environment.
Fire is one of the most common disturbances considered for re-introduction. Historical fire regimes have been a key driver in shaping plant community composition and dominance in many arid and semiarid rangelands (Miller and Rose 1999; Davis et al. 2000; Fuhlendorf et al. 2002). Anthropogenic activity has directly and indirectly altered many fire regimes around the world. Where fire has been excluded, re-introducing fire is often needed to conserve specific fire-dependent plant community types and associated fauna, as well as create spatial and temporal heterogeneity (Baker 1994; Brockett et al. 2001). Prolonged exclusion of fire and deviations from historical conditions, however, may have altered the response of plant communities to fire (Varner et al. 2005; Davies et al. 2009; Thorpe and Stanley 2011). Therefore, it is imperative to investigate effects, especially over longer time frames, of re-introducing fire.
In sagebrush-steppe communities of North America, fire was a periodic disturbance that shifted dominance from woody to herbaceous vegetation (Wright and Bailey 1982; Miller and Rose 1999). These periodic fires likely created spatial and temporal heterogeneity in vegetation composition and structure. Fire regimes have been substantially altered across the sagebrush-steppe. In the lower elevations, typified by Wyoming big sagebrush (Artemisia tridentata Nutt. ssp. wyomingensis Beetle & Young) communities, fire frequency often increased, especially following exotic annual grass invasion (Balch et al. 2013). In higher-elevation sagebrush communities, commonly mountain big sagebrush (A. tridentata Nutt. ssp. vaseyana (Rydb.) Beetle) communities, fire frequency has often decreased (Miller and Wigand 1994; Miller and Rose 1995, 1999). Prolonging the interval between fires in mountain big sagebrush communities has resulted in increased dominance of woody vegetation and decreases in herbaceous vegetation (West 1983; Miller and Rose 1999).
Though periodic fires were common in mountain big sagebrush communities in the Great Basin and Intermountain West, there is a current aversion to re-introducing fire. In fact, fire exclusion is actively promoted across the entire sagebrush biome (e.g. US Fish and Wildlife Service (FWS) 2013). Wide-spread loss of sagebrush habitat, especially for species such as sage grouse, is one of the driving forces behind fire-averse management policies. The rationale for fire exclusion is that sagebrush is fire-intolerant and consequently, fire, at a minimum, temporarily removes sagebrush from the community. Another significant driving force for fire exclusion is the concern that fire promotes exotic annual grasses (D’Antonio and Vitousek 1992; Chambers et al. 2007). Exotic annual grasses frequently increase after fire and other disturbances, particularly in hotter and drier sagebrush communities (Stewart and Hull 1949; Davies et al. 2009; Chambers et al. 2014). Exotic annual grasses, however, are also a risk in cooler, wetter mountain big sagebrush communities (Bates et al. 2005; Bates et al. 2014; Davies et al. 2019). Exotic annual grass dominance would homogenise vegetation and decrease diversity (Davies 2011). Understandably, ecosystem managers are hesitant to embrace fire reintroduction in these communities because of a heightened risk of substantial increases in exotic annual grasses.
Short-term studies may not be adequate to determine exotic annual grass response to fire in intact mountain big sagebrush communities. Exotic annual grass response to disturbances in these communities may not be readily apparent in the first few years post disturbance. In juniper-encroached big sagebrush communities, exotic annual grasses were not a substantial component of the plant community for the first three growing seasons after disturbance (juniper treatment), then co-dominated the understorey by the sixth year after disturbance, but then declined to a minor component of the understorey in subsequent years (Bates et al. 2005). In contrast, exotic annual grasses did not respond to disturbance (fire) in a before-and-after comparison in Utah (Riginos et al. 2019). Exotic annual grass response varies by severity of fire, whether or not perennial grasses recover (Bates et al. 2014), and the abundance of perennial grasses pre-fire (Ellsworth and Kauffman 2017). Clearly a long-term investigation of fire effects on exotic annual grasses in mountain big sagebrush communities with untreated controls and without conifer encroachment is needed.
Fire alters plant competition dynamics in sagebrush communities. However, significant knowledge gaps exist, particularly regarding fire effects in intact mountain big sagebrush communities. High sagebrush cover can competitively suppress herbaceous vegetation; thus, burning sagebrush communities often results in a 2- to 3-fold increase in herbaceous vegetation (Harniss and Murray 1973; Davies et al. 2012; Riginos et al. 2019). Perennial grass production and cover generally increase with reductions in big sagebrush cover (Davies et al. 2007, 2012; Riginos et al. 2019). However, other functional groups responses are less clear. Perennial forb response has frequently been grouped with the annual forb response (e.g. Mueggler and Blaisdell 1958; Dahlgren et al. 2006), making it difficult to tease out fire effects on perennial forbs. The few studies assessing perennial forb response to fire in mountain big sagebrush communities have contrasting results. Some studies found perennial forbs increased (Pyle and Crawford 1996; Riginos et al. 2019) and others found they decreased with burning (Davies et al. 2012). Limited information also exists for evaluating fire reintroduction effects on shrubs other than sagebrush. Determining the long-term response of different plant groups to fire in mountain big sagebrush communities is needed to guide management, especially with current efforts to limit fire in these communities. Better long-term data on vegetation responses will allow managers to assess potential consequences and benefits of re-introducing fire in mountain big sagebrush communities.
The purpose of the present project was to determine the long-term effects of fire reintroduction in mountain big sagebrush communities experiencing prolonged fire exclusion. We speculated that: (1) perennial herbaceous functional groups would increase with burning for the duration of the study; (2) exotic annual grasses would initially increase with burning but decrease as other vegetation increased; and (3) sagebrush would be removed with burning, but would be recovering within a decade. Therefore, we expected that burned areas would be dissimilar from unburned areas for the duration of the study, creating spatial heterogeneity, and that vegetation community characteristics would vary over time in burned areas (post-disturbance succession), creating temporal heterogeneity.
Methods
Study area
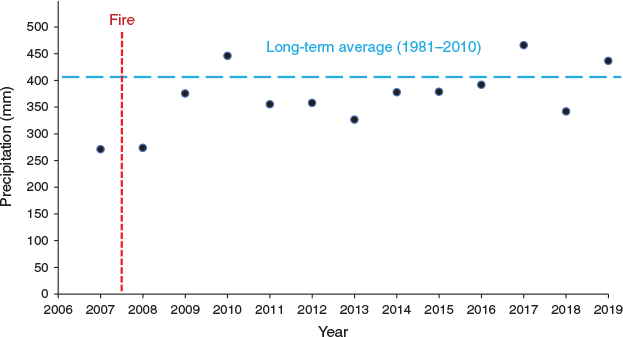
Our study was conducted in south-eastern Oregon on the 112 503-ha Hart Mountain National Antelope Refuge (42°21′16″N 119°22′54″W). Study sites ranged in elevation from 2013 to 2166 m above sea level. Slopes were from 0 to 7° with aspects ranging from north to south. Long-term average annual precipitation was 401 mm (PRISM 2020). Most precipitation occurs in the winter and early spring and summers are hot and dry. Annual precipitation the year of the burn (2007) was 68% of the long-term average and then was 68 and 94% of the long-term average the 2 years following burning (PRISM 2020; Fig. 1). The study area was classified as having high ecosystem resilience to disturbance and resistance to cheatgrass (Bromus tectorum L.), an exotic annual grass, invasion (Sage Grouse Initiative 2020). Mountain big sagebrush communities in this region historically experienced fire-return intervals between 10 and 25 years (Miller and Rose 1995, 1999; Miller et al. 2000). Our study area had no record of fire since the formation of the Hart Mountain National Antelope Refuge in 1936, but we speculated that fire exclusion had occurred for >100 years because wildfires became rare with European settlement across the region (Miller and Rose 1995, 1999). Prior to burning, the study site was intact mountain big sagebrush steppe (Davies and Bates 2010). The understorey was dominated by perennial grasses and perennial forbs. Prior to treatment, mountain big sagebrush, native perennial grasses (excluding Sandberg bluegrass (Poa secunda J. Presl)) and native perennial forb cover averaged 30, 20 and 12% respectively (Davies et al. 2014). Common perennial grasses included Columbia needlegrass (Achnatherum nelsonii (Scribn.) Barkworth), bottlebrush squirreltail (Elymus elymoides (Raf.) Swezey), prairie Junegrass (Koeleria macrantha (Ledeb.) Schult.), Idaho fescue (Festuca idahoensis Elmer), bluebunch wheatgrass (Pseduoroegneria spicata (Pursh) A. Löve), mountain brome (Bromus marginatus Nees ex Steud.), Thurber’s needlegrass (Achnatherum thurberianum (Piper) Barkworth), Sandberg bluegrass and other bluegrass species (Poa L.). Common perennial forbs included lupines (Lupinus L.), milkvetches (Astragalus L.), yarrow (Achillea millefolium L.), paintbrushes (Castilleja Mutis ex L. f.), hawksbeard (Crepis L.), biscuitroots (Lomatium Raf.), ragwort (Senecio integerrimus Nutt.) and fleabanes (Erigeron L.). Wildlife were allowed full access to our study sites, but low ungulate numbers in the area and limited observations of wildlife (other than sage grouse) suggest that wildlife use was quite low. Domestic livestock were removed from the Refuge in the mid-1990s.
|
Experimental design
A randomised complete block design was used to evaluate the response of mountain big sagebrush communities to re-introducing fire. Blocks were selected that were large enough for both treatments, were accessible with fire equipment, and did not interfere with other research or management objectives on the refuge. Six blocks (sites) with varying site (elevation, topography, soil) and vegetation characteristics were included in this experiment. Treatments were randomly assigned to two 60 × 90-m plots within each block. Treatment plots within a block had similar vegetation and site characteristics and were adjacent to one another with a 5-m buffer between them. Treatments were a fall (autumn) prescribed burn and an unburned control. Burned treatments were applied between mid-October and early November 2007 as strip head fires ignited with drip-torches. Fine fuel loads ranged from 327 to 977 kg ha−1 and shrub cover averaged >30%. During the burns, wind speed varied from 2 to 10 km h−1, air temperatures ranged from 6 to 11°C, and relative humidity was between 33 and 43%. Burns were complete across the plots assigned the burn treatment.
Measurements
Response variables were measured in late June or early July 2008, 2009, 2010, 2013, 2015, 2017 and 2019. Vegetation cover and density measurements were along four parallel 50-m transects spaced at 10-m intervals in each treatment plot. Herbaceous vegetation, bare ground, litter and biological soil crust cover was visually estimated in sixty 0.2-m2 quadrats spaced at 3-m intervals along the 50-m transects (starting at 3 m and ending at 45 m). Shrub cover was estimated using the line-intercept method along the 50-m transects. Herbaceous vegetation density was also measured in the 0.2-m2 quadrats by counting all plants rooted inside the quadrat. Shrub density was measured by counting all shrubs rooted inside 2 × 50-m belt transects, positioned on each of the 50-m transects. Above-ground herbaceous biomass was measured by clipping by plant group 15 randomly located 1-m2 quadrats per treatment plot. Harvested biomass was oven-dried for 72 h at 50°C, separated into current and prior years’ growth and weighed.
Statistical analyses
Repeated-measures analysis of variance (ANOVA) using the mixed-models procedure (Proc Mixed) in SAS ver.9.4 (SAS Institute Inc., Cary, NC) was used to determine the influence of re-introducing fire on response variables across years. Fixed variables were treatment and time since treatment (year) and their interactions. Random variables were blocks (sites) and block by treatment interactions. Covariance structures were selected using the Akaike’s Information Criterion (Littell et al. 1996). Data that violated assumptions of ANOVA were square-root-transformed. All figures present original data (i.e. non-transformed). Response variable means were reported with standard errors. Differences between means were considered significant at P ≤ 0.05. For analyses, herbaceous cover, density and biomass were separated into plant groups: perennial grass excluding Sandberg bluegrass, Sandberg bluegrass, exotic annual grass, perennial forb and annual forb. Sandberg bluegrass was treated as a separate functional group from the other perennial grasses because it is smaller in stature, develops phenologically earlier, and responds differently to management and disturbances than other perennial grasses (Robertson 1971; Davies et al. 2007; James et al. 2008). The exotic annual grass group primary consisted of cheatgrass. For analyses, shrubs were separated into sagebrush and other shrubs. The other shrubs group consisted of wax currant (Ribes cereum Dougl.), Wood’s rose (Rosa woodsia Lindl.), horsebrush species (Tetradymia DC.), snowberry (Symphoricarpos oreophilus Gray), green rabbitbrush (Chrysothamnus viscidiflorus (Hook) Nutt.), grey rabbitbrush (Ericameria nauseosa (Pall. ex Pursh) G.L. Nesom & Baird) and bitterbrush (Purshia tridentata (Pursh) DC.).
Results
Cover
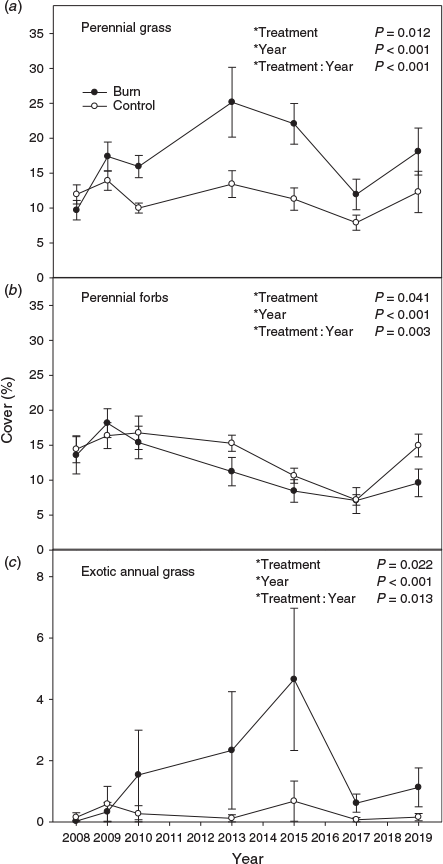
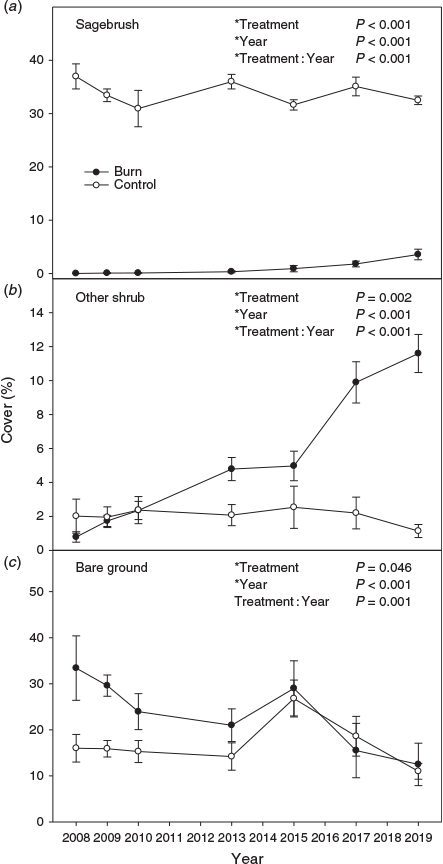
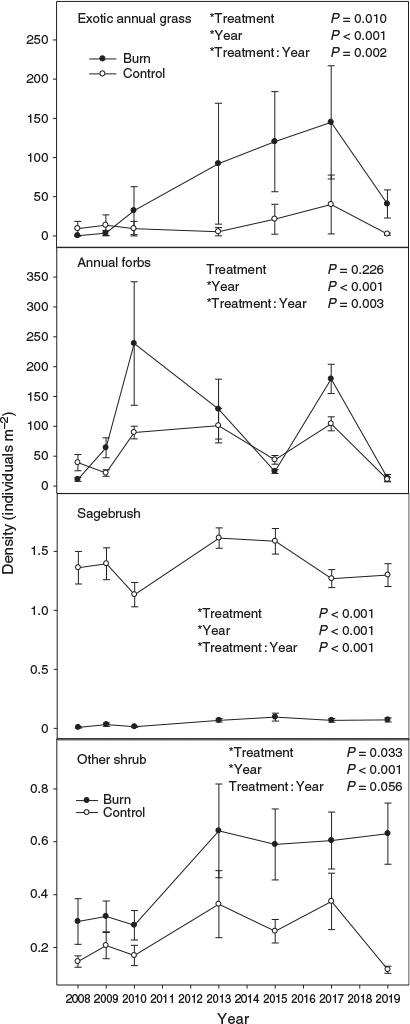
Perennial grass cover response to treatment varied with year (i.e. treatment by year interaction) (Fig. 2a; P < 0.001). Perennial grass cover in the burned treatment was less than the untreated control in the first year after fire, then became almost 2 times greater by the sixth year post treatment. By 10 years post treatment, the difference in perennial grass cover had decreased to ~1.5 times. Sandberg bluegrass cover did not differ among treatments (data not shown; P = 0.168), but varied by year (P < 0.001). Perennial forb cover response to treatment varied by year (Fig. 2b; P = 0.003), with it generally being greater in the control than the burn after 2010. Exotic annual grass cover response to treatment varied by year (Fig. 2c; P = 0.013). Exotic annual grass cover was generally greater in the burned than untreated control, except for the first 2 years post fire, but by a decade post treatment, exotic annual grass cover was ~1% or less in the burn treatment. Annual forb and total herbaceous cover responses to treatment varied with year (data not shown; P < 0.001) and both followed a pattern similar to perennial grass cover. Sagebrush and other shrub cover responses to treatment varied by year (P < 0.001). Sagebrush cover was less in the burn than the control, but the difference became smaller over time (Fig. 3a). Other shrub cover was initially less in the burn, but became 4.5 and 10.2 times greater in the burn compared with the control in the 10th and 12th year post fire (Fig. 3b). Bare ground and litter responses to treatment varied with year (P = 0.001 and <0.001). Bare ground was generally greater in the burn compared with the control early in the study, but converged by the eighth year post fire (Fig. 3c). Litter response (data not shown) was the inverse of bare ground early in the study, but similarly converged between treatments in later years. Biological soil crust cover was low in both treatments (burn 0.14 ± 0.04% and control 0.47 ± 0.11%), but was less in the burn compared with the control (P = 0.023) and varied by year (P = 0.026).
|
|
Density
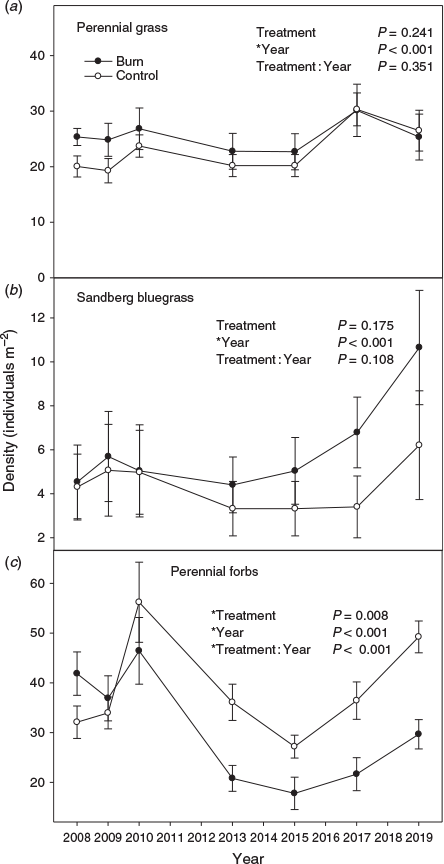
Perennial grass and Sandberg bluegrass density did not vary by treatment (Fig. 4a and 4b; P = 0.241 and 0.175 respectively), but differed among years (P < 0.001). Perennial forb density response to treatment was influenced by year (Fig. 4c; P < 0.001). Perennial forb density was initially similar or greater in the burn compared with the unburned control, but approximately half way through the study, it became greater in the control. The response of exotic annual grass density to burning varied with time (Fig. 5a; P = 0.002). The first 2 years post fire, annual grass density was generally greater in the control than the burn. For the next 10 years, exotic annual grass density was greater in the burn compared with the control. The response of annual forb density to treatment varied by year (Fig. 5b; P = 0.003). The first year post fire, annual forb density was greater in the control than burn treatment, but after that, it was generally greater in the burn than the control except in the final year of the study where the treatments appear to have converged. Sagebrush density was greater in the control than the burn, but the magnitude of difference decreased with time (Fig. 5c; P < 0.001). Other shrub density was greater in the burn compared with the control and varied among years (Fig. 5d; P = 0.033 and < 0.001, respectively). By the end of the study, other shrub density was >5 times greater in the burn compared with the control.
|
|
Production
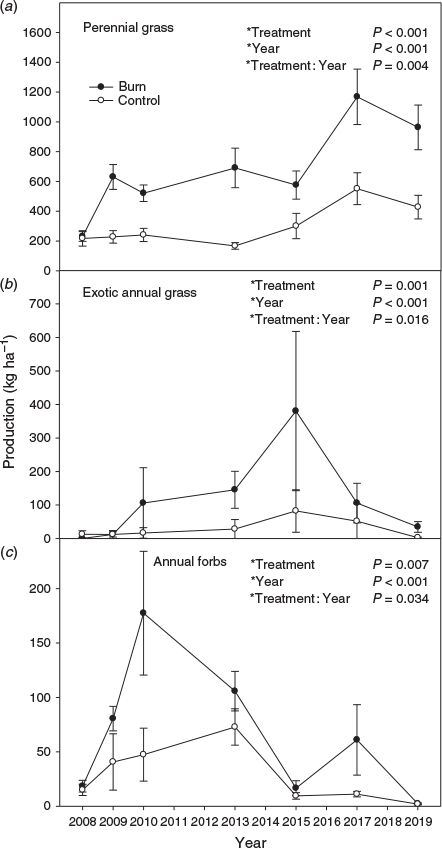
The response of perennial grass production to treatment varied with year (Fig. 6a; P = 0.004). The first-year post-fire perennial grass production was similar between treatments, but after that, it was 2 to 4 times greater in the burn compared with the control treatment. Sandberg bluegrass production was greater in the burn compared with the control treatment (data not shown; P = 0.029) and varied among years (P = 0.025). At the end of the study, Sandberg bluegrass production was 19.4 ± 10.7 and 47.8 ± 13.1 kg ha−1 in control and burned treatments respectively. Perennial forb production was similar between treatments (data not shown; P = 0.959), but varied among years (P = 0.003). Exotic annual grass production response to treatment varied with year (Fig. 6b; P = 0.016). The first year post-fire, annual grass production was less in the burn than the control treatment. In the second year, annual grass production was similar between treatments, and then it was greater in the burn compared with the control treatment for the rest of the study, but had substantially declined in the burn treatment after the eighth year post fire. Annual forb production was generally greater in the burn compared with the control treatment, but by the end of the study, it was similar between treatments (Fig. 6c; P = 0.034). Total herbaceous production response to treatment varied by year (data not shown; P = 0.039) and followed a similar pattern to perennial grass production. At the end of the study, total herbaceous production was 765.8 ± 74.6 and 1287.6 ± 167.5 kg ha−1 in control and burned treatment respectively.
|
Discussion
Re-introducing fire in shrub-steppe communities experiencing decreased fire frequency can increase vegetation heterogeneity without promoting long-term undesirable shifts in vegetation composition. Burning in mountain big sagebrush communities had substantial effects on vegetation that were still evident 12 years post fire. Total herbaceous and perennial grass production was consistently greater in the burned compared with the unburned control. However, not all plant groups responded to burning and those that responded to burning had varying responses. Some responses also varied across time, creating temporal heterogeneity, and others were more consistent. Re-introducing fire in mountain big sagebrush and other shrub-steppe communities to create a mosaic of burns of different ages and unburned areas can increase spatial and temporal heterogeneity. Similarly, prescribed burning increased heterogeneity in savannas in South Africa (Van Wilgen et al. 2004) and Australia (Price et al. 2005).
Anthropogenic activities, such as fire exclusion, have resulted in plant communities becoming homogeneous, resulting in decreased biodiversity and reduced ecosystem function (Derner et al. 2009; Fuhlendorf et al. 2009). Re-introducing fire within large unburned mountain big sagebrush landscapes increased heterogeneity and similar results would be expected in other ecosystems experiencing prolonged fire exclusion, but that have adequate ecological memory for recovery. These results are important, because increased heterogeneity promotes greater diversity in higher trophic levels (Adler et al. 2001; Hovick et al. 2015). Greater spatiotemporal heterogeneity also provides a wider breadth of habitats, increased ecosystem services and greater community stability (Fuhlendorf et al. 2006; Engle et al. 2008; Hovick et al. 2015). Re-introducing disturbances that promote vegetation heterogeneity undoubtedly provides ecological value and should be considered by ecosystem managers.
Fire removed sagebrush and generally favoured herbaceous vegetation, most evident in the doubling to quadrupling of perennial grass production. This is reflective of how wildfires historically shifted dominance from sagebrush to herbaceous vegetation, primarily perennial grasses (Wright and Bailey 1982; Miller and Rose 1999). Shorter-term studies have also found that burning mountain big sagebrush communities increases herbaceous vegetation (Davies et al. 2012; Riginos et al. 2019). In general, decreases in woody vegetation result in increases in herbaceous vegetation (Archer et al. 2011). Total herbaceous vegetation and perennial grasses show limited evidence of converging between unburned and burned areas. The lack of substantial increases in mountain big sagebrush cover in the 12 years post fire was likely the reason herbaceous vegetation was generally not converging between treatments. Sagebrush and herbaceous vegetation, in particular perennial grasses, compete for resources in sagebrush communities (Robertson 1947; Cook and Lewis 1963; Rittenhouse and Sneva 1976). Increases in mountain big sagebrush cover result in decreases in herbaceous vegetation (Davies and Bates 2019). Differences in herbaceous vegetation between burned and unburned areas will likely continue until sagebrush cover returns to pre-burn levels.
Perennial forbs did not increase with re-introduction of fire in mountain big sagebrush communities in our study. In contrast with our results, other authors have reported increases in perennial forbs after fire in sagebrush communities (Pyle and Crawford 1996; Riginos et al. 2019) or increased reproductive efforts that they speculated would result in increased perennial forb abundance (Wrobleski and Kauffman 2003). A few years after burning, perennial forb density was less in burned than unburned areas in our study. We speculate the limited negative impact of fire in our study on perennial grasses probably precluded any increase in perennial forbs and as grasses increased, perennial forbs experienced more competition. Though burning appeared to slightly negatively impact perennial forbs, year appeared to be the primary factor determining perennial forb abundance, cover and production. Similarly, other authors have reported annual weather conditions were the main determinant of perennial forbs response in sagebrush ecosystems (Sneva 1982; Bates et al. 2005; Bates and Davies 2019).
Sagebrush recovery was slow, in part owing to low establishment the first couple of years post fire and competition from herbaceous vegetation. Mountain big sagebrush establishment after fire is often limited if precipitation is below average in the year following the fire (Nelson et al. 2014). Precipitation at our study sites was lower than average before and the year after burning. If mountain big sagebrush does not establish well in the first couple of years after fire, sagebrush recovery is slow (Ziegenhagen and Miller 2009). Sagebrush also establishes better in microsites that are largely devoid of herbaceous vegetation (Davies and Bates 2019; Davies et al. 2020). In our study, limited-to-no fire-induced mortality of perennial grasses resulted in few open spaces for establishment of sagebrush seedlings. This limited mortality of perennial grasses and subsequent rapid increase in herbaceous vegetation were likely the result of wet and cool conditions during prescribed fires. Competition from herbaceous vegetation has been repeatedly shown to limit sagebrush and other shrub recovery (Schuman et al. 1998; Hall et al. 1999; Rinella et al. 2015, 2016). The slow recovery of sagebrush in our study sites was expected considering annual precipitation was low following burning and fire effects on perennial herbaceous vegetation were limited because of cool, wet conditions during burns.
The removal of sagebrush presumably allowed increases in other shrubs that were subdominant. Similarly, fire increased subdominant shrubs in subtropical shrublands in Florida (Horn 2018). Removal of dominant species allows species that were suppressed by the dominant species to increase, often increasing diversity (Kunte 2008; McCain et al. 2010). Our results were similar to other studies that found burning sagebrush communities resulted in increases in other shrubs (Harniss and Murray 1973; Beck et al. 2009; Bates et al. 2014). In contrast, other shrubs did not increase when mountain big sagebrush communities were burned in eastern Idaho (Seefeldt et al. 2007). This may have been because other shrubs were not abundant at these sites or because their response was measured for only 2 to 3 years post fire (Seefeldt et al. 2007). In our study, there was a distinct increase in the density of other shrubs between the third and sixth sampling year that may have been missed if we had employed a short-term study. Increases in other shrub species may increase biodiversity, especially if burned areas are intermixed within an unburned landscape as this creates a mosaic of different habitats. In a manner similar to our results, we expect that removing dominant woody vegetation in other ecosystems would favour subdominant woody species. In agreement with this expectation, subdominant plant functional groups were found to be able to exploit lower levels of competition after fire in woodlands in south-western Australia (Gosper et al. 2013).
The steady increase in exotic annual grasses in the burn treatment for 8 years post fire was an alarming trajectory. Increases in exotic grasses can result in a permanent shift in plant communities as they can induce a grass–fire cycle that prevents reestablishment of native species (D’Antonio and Vitousek 1992; Rossiter et al. 2003). Exotic annual grasses in the burn treatment, however, peaked in 2015 and subsequently became a much more minor component of the plant community for the last 4 years of the study. This corresponds to a substantial increase in perennial grass production and other shrub cover after 2015. We speculate that the increase in perennial grass production and other shrub cover caused the decrease in annual grasses because competition from established perennial vegetation, especially perennial grasses, greatly limits exotic annual grasses (Chambers et al. 2007; Davies 2008; Davies and Johnson 2017). Annual grasses are also less competitive with native perennials in cooler and wetter sagebrush habitats compared with hotter and drier sagebrush communities where exotic annual grasses are more problematic (Chambers et al. 2007, 2014). Therefore, sagebrush communities with lower resilience to disturbance and resistance to exotic annual grass invasion may not respond favourably to burning. At the conclusion of the study, exotic annual grasses were not a concern in the burn treatment as they had decreased to the point of being largely innocuous (~1% cover).
Our study highlights the importance of long-term research. If we had only sampled the first 8 years post fire, we would have incorrectly presumed that burning had resulted in a substantial and increasing exotic annual grass problem. This may have led to an incorrect assumption that re-introducing the historical disturbance regime in these communities creates a novel and undesirable plant community. Similarly, short-term research would not have identified the substantial increases in abundance and cover of other shrubs with the re-introduction of fire. Our results provide another example demonstrating that short-term results need to be verified with long-term research and caution against making land or wildlife management decisions based on short-term research.
Conclusions
Re-introducing fire in shrub-steppe communities experiencing elongated fire-free periods can increase spatial and temporal heterogeneity. Subdominant shrubs benefited from re-introducing fire, with their cover increasing to more than 10-fold that of unburned areas by the end of the study. This likely promotes diversity in higher trophic levels when burns are applied within large unburned landscapes because of increased variability in vegetation structure and composition. Burning mountain big sagebrush communities also substantially increased herbaceous vegetation, resulting in more forage for herbivores. These effects were still evident 12 years post fire and will likely remain until sagebrush returns to pre-burn cover levels. There was a temporary increase in exotic annual grasses, but, unlike lower-elevation sagebrush communities, this was a transient response. This response does caution against applying treatments that substantially reduce the abundance of perennial grasses or in areas with less resistance to annual grass invasion as recovery may be slower or not as successful. The results of our study suggest that excluding fire from communities that evolved with periodic fire may not be warranted. This policy may lead to declines in fire-adapted species and plant groups that are important contributors to biodiversity in shrub-steppe ecosystems and a general homogenisation of vegetation characteristics across landscapes. Re-introducing fire in intact mountain big sagebrush communities will likely be challenging with the current emphasis on maintaining sagebrush-dominated landscapes for sagebrush-associated wildlife. This highlights the need to reverse the decline of mountain big sagebrush communities from conifer encroachment to allow more use of fire in these communities. This research highlights that re-introducing fire to create spatial and temporal variability could be valuable in mountain big sagebrush and other shrub-steppe communities experiencing prolonged fire absence.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgments
The authors thank the Hart Mountain National Antelope Refuge for allowing this research to be conducted on lands they administer and for applying the prescribed fires. Gail Collins was especially helpful in locating study sites and providing logistical support. The authors thank Woody Strachan and Aleta Nafus for leading the field crews and data management. The authors also appreciate all the summer technicians for field sampling and sample processing. We also are grateful to Dustin Johnson and an anonymous reviewer for reviewing earlier versions of this manuscript. We appreciate the thoughtful and constructive reviews by the anonymous reviewers and Associate Editor. This research did not receive any specific funding. USDA is an equal opportunity provider and employer. Mention of a proprietary product does not constitute a guarantee or warranty of the product by USDA or the authors and does not imply its approval to the exclusion of other products.
References
Adler PB, Raff DA, Lauenroth WK (2001) The effects of grazing on spatial heterogeneity of vegetation. Oecologia 128, 465–479.| The effects of grazing on spatial heterogeneity of vegetation.Crossref | GoogleScholarGoogle Scholar |
Archer S, Davies KW, Fulbright TE, McDaniel KC, Wilcox BP, Predick KI (2011) Brush management as a rangeland conservation strategy: a critical evaluation. In ‘Conservation benefits of rangeland practices’. (Ed. DD Briske) pp. 105–170. (USDA Natural Resources Conservation Service: Washington, DC, USA)
Baker WL (1994) Restoration of landscape structure altered by fire suppression. Conservation Biology 8, 763–769.
| Restoration of landscape structure altered by fire suppression.Crossref | GoogleScholarGoogle Scholar |
Balch JK, Bradley BA, D’Antonio CM, Gómez-Dans J (2013) Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Global Change Biology 19, 173–183.
| Introduced annual grass increases regional fire activity across the arid western USA (1980–2009).Crossref | GoogleScholarGoogle Scholar | 23504729PubMed |
Bates JD, Davies KW (2019) Characteristics of intact Wyoming big sagebrush associations in south-eastern Oregon. Rangeland Ecology and Management 72, 36–46.
| Characteristics of intact Wyoming big sagebrush associations in south-eastern Oregon.Crossref | GoogleScholarGoogle Scholar |
Bates JD, Miller RF, Svejcar T (2005) Long-term successional trends following western juniper cutting. Rangeland Ecology and Management 58, 533–541.
| Long-term successional trends following western juniper cutting.Crossref | GoogleScholarGoogle Scholar |
Bates JD, Davies KW, Sharp RN (2014) Sagebrush steppe recovery after fire varies by development phase of Juniperus occidentalis woodland. International Journal of Wildland Fire 23, 117–130.
| Sagebrush steppe recovery after fire varies by development phase of Juniperus occidentalis woodland.Crossref | GoogleScholarGoogle Scholar |
Beck JL, Connelly JW, Reese KP (2009) Recovery of greater sage-grouse habitat features in Wyoming big sagebrush following prescribed fire. Restoration Ecology 17, 393–403.
| Recovery of greater sage-grouse habitat features in Wyoming big sagebrush following prescribed fire.Crossref | GoogleScholarGoogle Scholar |
Brockett BH, Biggs HC, Van Wilgen BW (2001) A patch mosaic burning system for conservation areas in southern African savannas. International Journal of Wildland Fire 10, 169–183.
| A patch mosaic burning system for conservation areas in southern African savannas.Crossref | GoogleScholarGoogle Scholar |
Chambers JC, Roundy BA, Blank RR, Meyer SE, Whittaker A (2007) What makes Great Basin sagebrush ecosystems invasible by Bromus tectorum? Ecological Monographs 77, 117–145.
| What makes Great Basin sagebrush ecosystems invasible by Bromus tectorum?Crossref | GoogleScholarGoogle Scholar |
Chambers JC, Bradly BA, Brown CS, Germino MJ, Grace JB, Hardegree SP, Miller RF, Pyke DA (2014) Resilience to stress and disturbance and resistance to Bromus tectorum L. invasion in cold desert shrublands of western North America. Ecosystems 17, 360–375.
| Resilience to stress and disturbance and resistance to Bromus tectorum L. invasion in cold desert shrublands of western North America.Crossref | GoogleScholarGoogle Scholar |
Cook CW, Lewis CE (1963) Competition between big sagebrush and seeded grasses on foothill ranges in Utah. Journal of Range Management 16, 245–250.
| Competition between big sagebrush and seeded grasses on foothill ranges in Utah.Crossref | GoogleScholarGoogle Scholar |
D’Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annual Review of Ecology and Systematics 23, 63–87.
| Biological invasions by exotic grasses, the grass/fire cycle, and global change.Crossref | GoogleScholarGoogle Scholar |
Dahlgren DK, Chi R, Messmer TA (2006) Greater sage-grouse response to sagebrush management in Utah. Wildlife Society Bulletin 34, 975–985.
| Greater sage-grouse response to sagebrush management in Utah.Crossref | GoogleScholarGoogle Scholar |
Davies KW (2008) Medusahead dispersal and establishment in sagebrush steppe plant communities. Rangeland Ecology and Management 61, 110–115.
| Medusahead dispersal and establishment in sagebrush steppe plant communities.Crossref | GoogleScholarGoogle Scholar |
Davies KW (2011) Plant community diversity and native plant abundance decline with increasing abundance of an exotic annual grass. Oecologia 167, 481–491.
| Plant community diversity and native plant abundance decline with increasing abundance of an exotic annual grass.Crossref | GoogleScholarGoogle Scholar | 21509533PubMed |
Davies KW, Bates JD (2010) Vegetation characteristics of mountain and Wyoming big sagebrush plant communities in the northern Great Basin. Rangeland Ecology & Management 63, 461–466.
| Vegetation characteristics of mountain and Wyoming big sagebrush plant communities in the northern Great Basin.Crossref | GoogleScholarGoogle Scholar |
Davies KW, Bates JD (2019) Longer-term evaluation of sagebrush restoration after juniper control and herbaceous vegetation trade-offs. Rangeland Ecology and Management 72, 260–265.
| Longer-term evaluation of sagebrush restoration after juniper control and herbaceous vegetation trade-offs.Crossref | GoogleScholarGoogle Scholar |
Davies KW, Johnson DD (2017) Established perennial vegetation provides high resistance to reinvasion by exotic annual grasses. Rangeland Ecology and Management 70, 748–754.
| Established perennial vegetation provides high resistance to reinvasion by exotic annual grasses.Crossref | GoogleScholarGoogle Scholar |
Davies KW, Bates JD, Miller RF (2007) Short-term effects of burning Wyoming big sagebrush steppe in south-eastern Oregon. Rangeland Ecology and Management 60, 515–522.
| Short-term effects of burning Wyoming big sagebrush steppe in south-eastern Oregon.Crossref | GoogleScholarGoogle Scholar |
Davies KW, Svejcar TJ, Bates JD (2009) Interaction of historical and non-historical disturbances maintains native plant communities. Ecological Applications 19, 1536–1545.
| Interaction of historical and non-historical disturbances maintains native plant communities.Crossref | GoogleScholarGoogle Scholar | 19769101PubMed |
Davies KW, Bates JD, Nafus AM (2012) Comparing burned and mowed treatments in mountain big sagebrush steppe. Environmental Management 50, 451–461.
| Comparing burned and mowed treatments in mountain big sagebrush steppe.Crossref | GoogleScholarGoogle Scholar | 22744159PubMed |
Davies KW, Bates JD, Boyd CS, Nafus AM (2014) Is fire exclusion in mountain big sagebrush communities prudent? Soil nutrient, plant diversity, and arthropod response to burning. International Journal of Wildland Fire 23, 417–424.
| Is fire exclusion in mountain big sagebrush communities prudent? Soil nutrient, plant diversity, and arthropod response to burning.Crossref | GoogleScholarGoogle Scholar |
Davies KW, Rios RC, Bates JD, Johnson DD, Kerby J, Boyd CS (2019) To burn or not to burn: comparing re-introducing fire with cutting an encroaching conifer for conservation of an imperiled shrub-steppe. Ecology and Evolution 9, 9137–9148.
| To burn or not to burn: comparing re-introducing fire with cutting an encroaching conifer for conservation of an imperiled shrub-steppe.Crossref | GoogleScholarGoogle Scholar | 31463011PubMed |
Davies KW, Bates JD, Clenet D (2020) Improving restoration success through microsite selection: an example with planting sagebrush seedlings after wildfire. Restoration Ecology
| Improving restoration success through microsite selection: an example with planting sagebrush seedlings after wildfire.Crossref | GoogleScholarGoogle Scholar |
Davis M, Peterson DW, Reich PB, Crozier M, Query T, Mitchell E, Huntington J, Bazakas P (2000) Restoring savanna using fire: impact on the breeding bird community. Restoration Ecology 8, 30–40.
| Restoring savanna using fire: impact on the breeding bird community.Crossref | GoogleScholarGoogle Scholar |
Derner J, Lauenroth WK, Stapp P, Augustine DJ (2009) Livestock as ecosystem engineers for grassland bird habitat in the Western Great Plains of North America. Rangeland Ecology and Management 62, 111–118.
| Livestock as ecosystem engineers for grassland bird habitat in the Western Great Plains of North America.Crossref | GoogleScholarGoogle Scholar |
Ellsworth LM, Kauffman JB (2017) Plant community response to prescribed fire varies by pre-fire conditions and season of burn in mountain big sagebrush ecosystem. Journal of Arid Environments 144, 74–80.
| Plant community response to prescribed fire varies by pre-fire conditions and season of burn in mountain big sagebrush ecosystem.Crossref | GoogleScholarGoogle Scholar |
Engle DM, Fuhlendorf SD, Roper A, Leslie DM (2008) Invertebrate community response to a shifting mosaic of habitat. Rangeland Ecology and Management 61, 55–62.
| Invertebrate community response to a shifting mosaic of habitat.Crossref | GoogleScholarGoogle Scholar |
Fuhlendorf SD, Woodward AJW, Leslie DM, Shackford JS (2002) Multiscale effects of habitat loss and fragmentation on lesser prairie-chicken populations of the US Southern Great Plains. Landscape Ecology 17, 617–628.
| Multiscale effects of habitat loss and fragmentation on lesser prairie-chicken populations of the US Southern Great Plains.Crossref | GoogleScholarGoogle Scholar |
Fuhlendorf SD, Harrell WC, Engle DM, Hamilton R, Davis CA, Leslie DM (2006) Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing. Ecological Applications 16, 1706–1716.
| Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing.Crossref | GoogleScholarGoogle Scholar | 17069365PubMed |
Fuhlendorf SD, Engle DM, Kerby J, Hamilton R (2009) Pyric herbivory: re-wilding landscapes through the recoupling of fire and grazing. Conservation Biology 23, 588–598.
| Pyric herbivory: re-wilding landscapes through the recoupling of fire and grazing.Crossref | GoogleScholarGoogle Scholar | 19183203PubMed |
Gosper CR, Yates CJ, Prober SM (2013) Floristic diversity in fire-sensitive eucalypt woodlands shows a ‘U’-shaped relationship with time since fire. Journal of Applied Ecology 50, 1187–1196.
| Floristic diversity in fire-sensitive eucalypt woodlands shows a ‘U’-shaped relationship with time since fire.Crossref | GoogleScholarGoogle Scholar |
Hall DB, Anderson VJ, Monsen SB (1999) Competitive effects of bluebunch wheatgrass, crested wheatgrass, and cheatgrass on antelope bitterbrush seedling emergency and survival. USDA Forest Service, Rocky Mountain Research Station, Research Paper RMRS-RP-16. (Ogden, UT, USA).
Harniss RO, Murray RB (1973) 30 years of vegetal change following burning of sagebrush–grass range. Journal of Range Management 26, 322–325.
| 30 years of vegetal change following burning of sagebrush–grass range.Crossref | GoogleScholarGoogle Scholar |
Horn KD (2018) Responses of the clonal Florida endemic shrub Polygonella myriophylla to fire and mechanical disturbance. American Midland Naturalist 180, 175–188.
| Responses of the clonal Florida endemic shrub Polygonella myriophylla to fire and mechanical disturbance.Crossref | GoogleScholarGoogle Scholar |
Hovick TJ, Elmore RD, Fuhlendorf SD, Engle DM, Hamilton RG (2015) Spatial heterogeneity increases diversity and stability in grassland bird communities. Ecological Applications 25, 662–672.
| Spatial heterogeneity increases diversity and stability in grassland bird communities.Crossref | GoogleScholarGoogle Scholar | 26214912PubMed |
James JJ, Davies KW, Sheley RL, Aanderud ZT (2008) Linking nitrogen partitioning and species abundance to invasion resistance in the Great Basin. Oecologia 156, 637–648.
| Linking nitrogen partitioning and species abundance to invasion resistance in the Great Basin.Crossref | GoogleScholarGoogle Scholar | 18343954PubMed |
Kunte K (2008) Competition and species diversity: removal of dominant species increases diversity in Costa Rican butterfly communities. Oikos 117, 69–76.
| Competition and species diversity: removal of dominant species increases diversity in Costa Rican butterfly communities.Crossref | GoogleScholarGoogle Scholar |
Littell RC, Milliken GA, Stroup WW, Wolfinger RD (1996) ‘SAS System for mixed models.’ (SAS Institute Inc.: Cary, NC, USA).
McCain KNS, Baer SG, Blair JM, Wilson GWT (2010) Dominant grasses suppress local diversity in restored tallgrass prairie. Restoration Ecology 18, 40–49.
| Dominant grasses suppress local diversity in restored tallgrass prairie.Crossref | GoogleScholarGoogle Scholar |
Miller RF, Rose JR (1995) Historic expansion of Juniperus occidentalis in south-eastern Oregon. The Great Basin Naturalist 55, 37–45.
Miller RF, Rose JR (1999) Fire history and western juniper encroachment in sagebrush steppe. Journal of Range Management 52, 550–559.
| Fire history and western juniper encroachment in sagebrush steppe.Crossref | GoogleScholarGoogle Scholar |
Miller RF, Wigand PE (1994) Holocene changes in semiarid pinyon–juniper woodlands; responses to climate, fire, and human activities in the US Great Basin. Bioscience 44, 465–474.
| Holocene changes in semiarid pinyon–juniper woodlands; responses to climate, fire, and human activities in the US Great Basin.Crossref | GoogleScholarGoogle Scholar |
Miller RF, Svejcar TJ, Rose JA (2000) Impacts of western juniper on plant community composition and structure. Journal of Range Management 53, 574–585.
| Impacts of western juniper on plant community composition and structure.Crossref | GoogleScholarGoogle Scholar |
Mueggler WF, Blaisdell JP (1958) Effects on associated species of burning, rotobeating, spraying, and railing sagebrush. Journal of Range Management 11, 61–66.
| Effects on associated species of burning, rotobeating, spraying, and railing sagebrush.Crossref | GoogleScholarGoogle Scholar |
Nelson ZJ, Weisberg PJ, Kitchen SG (2014) Influence of climate and environment on post-fire recovery of mountain big sagebrush. International Journal of Wildland Fire 23, 131–142.
| Influence of climate and environment on post-fire recovery of mountain big sagebrush.Crossref | GoogleScholarGoogle Scholar |
Price O, Edwards A, Conners G, Woinarski J, Ryan G, Turner A, Russell-Smith J (2005) Fire heterogeneity in Kakadu National Park, 1980–2000. Wildlife Research 32, 425–433.
| Fire heterogeneity in Kakadu National Park, 1980–2000.Crossref | GoogleScholarGoogle Scholar |
PRISM (2020) PRISM Climate Group – Explorer. Available at http://prism.nacse.org/explorer/ [Verified 1 April 2020]
Pyle WH, Crawford JA (1996) Availability of foods of sage grouse chicks following prescribed fire in sagebrush–bitterbrush. Journal of Range Management 49, 320–324.
| Availability of foods of sage grouse chicks following prescribed fire in sagebrush–bitterbrush.Crossref | GoogleScholarGoogle Scholar |
Riginos C, Monaco TA, Veblen KE, Gunnell K, Thacker E, Dahlgren D, Messmer T (2019) Potential for post-fire recovery of greater sage-grouse habitat. Ecosphere 10, e02870
| Potential for post-fire recovery of greater sage-grouse habitat.Crossref | GoogleScholarGoogle Scholar |
Rinella MJ, Hammond DH, Bryant AM, Kozar BJ (2015) High precipitation and seeded species competition reduce seeded shrub establishment during dryland restoration. Ecological Applications 25, 1044–1053.
| High precipitation and seeded species competition reduce seeded shrub establishment during dryland restoration.Crossref | GoogleScholarGoogle Scholar | 26465041PubMed |
Rinella MJ, Espeland EK, Moffat BJ (2016) Studying long-term, large-scale grassland restoration outcome to improving seeding methods and reveal knowledge gaps. Journal of Applied Ecology 53, 1565–1574.
| Studying long-term, large-scale grassland restoration outcome to improving seeding methods and reveal knowledge gaps.Crossref | GoogleScholarGoogle Scholar |
Rittenhouse LR, Sneva FA (1976) Expressing the competitive relationship between Wyoming big sagebrush and crested wheatgrass. Journal of Range Management 29, 326–327.
| Expressing the competitive relationship between Wyoming big sagebrush and crested wheatgrass.Crossref | GoogleScholarGoogle Scholar |
Robertson JH (1947) Response of range grasses to different intensities of competition with sagebrush (Artemisia tridentata Nutt.). Ecology 28, 1–16.
| Response of range grasses to different intensities of competition with sagebrush (Artemisia tridentata Nutt.).Crossref | GoogleScholarGoogle Scholar |
Robertson JH (1971) Changes on a sagebrush-grass range in Nevada ungrazed for 30 years. Journal of Range Management 24, 397–400.
| Changes on a sagebrush-grass range in Nevada ungrazed for 30 years.Crossref | GoogleScholarGoogle Scholar |
Rossiter NA, Setterfield SA, Douglas MM, Hutley LB (2003) Testing the grass–fire cycle: alien grass invasion in the tropical savannas of northern Australia. Diversity & Distributions 9, 169–176.
| Testing the grass–fire cycle: alien grass invasion in the tropical savannas of northern Australia.Crossref | GoogleScholarGoogle Scholar |
Sage Grouse Initiative (2020) Ecosystem resilience & resistance. Available at https://map.sagegrouseinitiative.com/ecosystem [Verified 1 April 2020]
Schuman GE, Booth DT, Cockrell JR (1998) Cultural methods for establishing Wyoming big sagebrush on mined lands. Journal of Range Management 51, 223–230.
| Cultural methods for establishing Wyoming big sagebrush on mined lands.Crossref | GoogleScholarGoogle Scholar |
Seefeldt SS, Germino M, DiCristina K (2007) Prescribed fires in Artemisia tridentata ssp. vaseyana steppe have minor and transient effects on vegetation cover and composition. Applied Vegetation Science 10, 249–256.
| Prescribed fires in Artemisia tridentata ssp. vaseyana steppe have minor and transient effects on vegetation cover and composition.Crossref | GoogleScholarGoogle Scholar |
Sneva F (1982) Relation of precipitation and temperature with yield of herbaceous plants in eastern Oregon. International Journal of Biometeorology 26, 263–276.
| Relation of precipitation and temperature with yield of herbaceous plants in eastern Oregon.Crossref | GoogleScholarGoogle Scholar |
Stewart G, Hull AC (1949) Cheatgrass (Bromus tectorum L.) – an ecologic intruder in southern Idaho. Ecology 30, 58–74.
| Cheatgrass (Bromus tectorum L.) – an ecologic intruder in southern Idaho.Crossref | GoogleScholarGoogle Scholar |
Thorpe AS, Stanley AG (2011) Determining appropriate goals for restoration of imperiled communities and species. Journal of Applied Ecology 48, 275–279.
| Determining appropriate goals for restoration of imperiled communities and species.Crossref | GoogleScholarGoogle Scholar |
Turner MG (2010) Disturbance and landscape dynamics in a changing world. Ecology 91, 2833–2849.
| Disturbance and landscape dynamics in a changing world.Crossref | GoogleScholarGoogle Scholar | 21058545PubMed |
USFWS (2013) Sage-grouse (Centrocercus urophasianus) conservation objectives: final report. US Fish and Wildlife Service. (Denver, CO, USA).
Van Wilgen BW, Govender N, Biggs HC, Ntsala D, Funda XN (2004) Response of savanna fire regimes to changing fire-management policies in a large African national park. Conservation Biology 18, 1533–1540.
| Response of savanna fire regimes to changing fire-management policies in a large African national park.Crossref | GoogleScholarGoogle Scholar |
Varner JM, Gordon DR, Putz FE, Hiers JK (2005) Restoring fire to long-unburned Pinus palustris ecosystem: novel fire effects and consequences for long-unburned ecosystems. Restoration Ecology 13, 536–544.
| Restoring fire to long-unburned Pinus palustris ecosystem: novel fire effects and consequences for long-unburned ecosystems.Crossref | GoogleScholarGoogle Scholar |
West NE (Ed) (1983) Western intermountain sagebrush steppe. In ‘Ecosystems of the world: temperate deserts and semideserts.’ pp. 351–374. (Elsevier Scientific Publishing Company: Amsterdam, The Netherlands.)
Williams JW, Jackson ST (2007) Novel climates, non-analog communities, and ecological surprises. Frontiers in Ecology and the Environment 5, 475–482.
| Novel climates, non-analog communities, and ecological surprises.Crossref | GoogleScholarGoogle Scholar |
Wright HA, Bailey AW (1982) ‘Fire ecology: United States and southern Canada.’ (John Wiley & Sons, Inc.: New York, NY, USA).
Wrobleski DW, Kauffman JB (2003) Initial effects of prescribed fire on morphology, abundance, and phenology of forbs in big sagebrush communities in south-eastern Oregon. Restoration Ecology 11, 82–90.
| Initial effects of prescribed fire on morphology, abundance, and phenology of forbs in big sagebrush communities in south-eastern Oregon.Crossref | GoogleScholarGoogle Scholar |
Ziegenhagen LL, Miller RF (2009) Postfire recovery of two shrubs in the interiors of large burns in the Intermountain West, USA. Western North American Naturalist 69, 195–205.
| Postfire recovery of two shrubs in the interiors of large burns in the Intermountain West, USA.Crossref | GoogleScholarGoogle Scholar |