

Decadal scale fire dynamics in savannas and forests of the Nilgiri Biosphere Reserve, India
Pradyumna Rajashekar A # , Aparna Krishnan A # , Varun Varma B , Jayashree Ratnam A * , Mahesh Sankaran A and Caroline E. R. Lehmann C D
B , Jayashree Ratnam A * , Mahesh Sankaran A and Caroline E. R. Lehmann C D
A
B
C
D
Abstract
The Nilgiri Biosphere Reserve (NBR) in the Western Ghats, India, contains a mosaic of savannas and forests. This landscape experiences wildfires regularly, but we lack an understanding of factors driving fire regimes in the region.
To examine the effects of climate, vegetation and human activity on wildfires in the NBR.
Using remotely sensed datasets, we examine how mean annual rainfall (MAR), topographic complexity and human activity influence fires in savannas and forests of the NBR across a wet (2001–2010) and dry decade (2011–2020).
Across both decades, savannas burned more frequently and over larger areas than forests. Burnt area and fire frequency in both habitats were higher in the wet decade. Human modification was the major driver of fire occurrence in the wet decade, but MAR took on greater importance in the dry decade.
Savannas and forests in the NBR have differing fire regimes. During dry periods, these systems are fuel limited and MAR best describes the occurrence of fires. During wet periods when fuel is not limiting, proxies for anthropogenic ignitions best explain fire occurrence.
Climate, vegetation and humans collectively determine fire regimes in the NBR. Fire management must integrate across all these factors at the landscape scale to be effective.
Keywords: fire management, forests, savannas, Southern India, traditional burning practices, Western Ghats.
Introduction
Fires are integral to regulating the distribution of terrestrial woody ecosystems such as savannas and forests, especially in the tropics where ~85% of the annual global burnt area lies (Bond et al. 2005; van der Werf et al. 2008; Hoffmann et al. 2012; McLauchlan et al. 2020). In particular, periodic fires in open-canopy savannas are crucial to maintaining their structure and function (Sankaran et al. 2004; Bond et al. 2005; Archibald et al. 2009; Hoffmann et al. 2012; Lehmann et al. 2014; Pausas and Bond 2020). On the other hand, fires are typically infrequent in closed-canopy forests except in dry years when they can result in loss of tree cover and degrade forest structure and function (Ratnam et al. 2011; Brando et al. 2014). A complex interplay of factors including inter- and intra-annual climatic variability, land topography, human presence and activity can influence fire occurrence and behaviour across landscapes (Archibald et al. 2009, 2013; Archibald 2016). Assessing the relationship between such factors and fire occurrence can provide fundamental information that can serve as a foundation for ecosystem management in savannas and forests, especially in the face of increasing anthropogenic ecosystem modification and a changing climate.
The occurrence of fires is driven by environmental, physical and anthropogenic factors that influence the frequency of ignitions, fuel quality and availability, fuel contiguity and ease of fire spread (Archibald et al. 2009). Rainfall and temperature govern the availability of moisture and energy for plant growth, set limits on plant production and hence, exert control over fuel load and moisture, which in turn influence the frequency and intensity of fire (Bradstock 2010). Land topography can influence fire spread such that topographically complex areas predominantly pose fire breaks and reduce fire continuity (Archibald et al. 2009). The opposite impact is also possible, where steeper slopes can increase rate of fire spread (Viegas 2004). Long-term rainfall and temperature patterns, as well as the rainfall and temperature preceding the dry season (during which most fires occur) in any given year can shape spatial and temporal variation in fire regimes. As global temperatures increase and dry conditions become more widespread in the tropics, understanding how climate, topography and human activity influence fire behaviour across vegetation types, and over time, will be essential in predicting the impacts of climate change on fire regimes.
While climate and topography pre-condition landscapes for the occurrence, intensity and spread of fire, human activities can alter fire regimes relative to their historic range of variability (Guyette et al. 2002; Duncan and Schmalzer 2004; Syphard et al. 2007; Archibald et al. 2009; Archibald 2016). Humans can alter fire frequencies by either suppressing fire, or alternately, increasing the frequency of ignitions (Keeley et al. 1999; Archibald 2016). Additionally, humans can change patterns of fire spread via landscape modification and fragmentation (Duncan and Schmalzer 2004; Archibald et al. 2009; Archibald 2016). Reflecting this, recent fire literature attempts to tease apart the relative importance of environmental factors (climate, topography and vegetation type) versus anthropogenic factors (human-initiated ignitions and land use), in driving fire frequency and behaviour across landscapes, with humans emerging as a primary source of ignitions globally (Marlon et al. 2008; Archibald et al. 2009; Bradstock 2010; Romero‐Ruiz et al. 2010; Archibald 2016; Wu et al. 2021).
Understanding the interplay between environmental and anthropogenic drivers of fires is of importance to fire management, particularly in areas of high human population densities and landscape change such as India. In the context of the Indian subcontinent, human presence across landscapes is both ancient and pervasive, with transformation from natural to semi-natural and modified landscapes amongst the most dramatic on the planet (Ellis et al. 2010). This, combined with the fact that fire in India is predominantly anthropogenic in origin, has shaped the understanding of fire in this region in two ways. First, it has contributed to a historical and widespread perception of fire as an anthropogenic threat to natural ecosystems (Champion and Seth 1968; Gadgil 1993; Saha and Howe 2003; Hiremath and Sundaram 2005; Kodandapani et al. 2008; Prasad et al. 2008; Takahata et al. 2010), precluding the notion that fires may be an integral component of at least some ecosystems (but see Puyravaud et al. 1994; Sankaran 2005; Kodandapani et al. 2009; Ratnam et al. 2019). This perception contrasts with other regions of the wet-dry tropics with similar climates and vegetation types, where fire, whether natural or anthropogenic in origin, is well-recognised as a fundamental driver of the occurrence and ecology of mixed tree-grass ecosystems (Bond 2008). Second, as a consequence of the first, most fire research in this region has emphasised the relationship between fire and human activities (Kodandapani et al. 2008; Takahata et al. 2010) and/or the vulnerability of vegetation regeneration to fire (Saha 2002; Saha and Howe 2003; Kodandapani et al. 2008, 2009; Mondal and Sukumar 2015; Verma et al. 2017). This has precluded a more comprehensive understanding of fire regimes as they relate to climatic and environmental drivers in the Indian subcontinent.
In this study we attempt to assess the factors that contribute to fire occurrence and behaviour in the Nilgiri Biosphere Reserve (NBR) of India (Fig. 1). The NBR is a model system representative of the peninsular Indian region in many ways. Within an area a little over 5000 km2, and with strong topographic and climatic variation, it contains the major vegetation types of the larger peninsular region, from open thorn scrub to deciduous woodland savanna to evergreen forests, as well as a large complement of native fauna including large herbivores and carnivores. It also contains areas of varying human densities, and associated human modified landscapes such as plantations, agricultural fields and villages. The NBR is thus characterised by the major conservation challenges of the wider Indian geography: threatened populations of iconic species such as elephants and tigers, transformation of native vegetation via severe weed invasions, intensification of land use modification by humans, and increasing human-wildlife conflict in shared landscapes. In terms of fire management, it is characteristic of many regions of the subcontinent where, over the past half-century, official policies of fire suppression by government departments have replaced traditional burning regimes practiced by indigenous communities.
Map of India showing the location of the Nilgiri Biosphere Reserve, and the distribution of savannas and forests (see Methods) across the region. The yellow areas indicate savannas while the green areas are forests.

We combine spatially explicit information on vegetation, degree of human land modification, topography, rainfall and fire in the NBR to develop an integrated picture of the fire ecology of the region over a 20-year period. We specifically aimed to examine how environmental and physical factors regulating fuel load, fire ignition and fire spread, for which rainfall, human modification and topographic complexity respectively, served as proxies, influenced incidences of fire between 2001 and 2020. During this 20-year period, the region saw a distinct ‘wet’ decade (2001–2010) followed by a ‘dry’ decade (2011–2020), as shown in Fig. 2. We compared the influence of the above factors on patterns of fire occurrence and behaviour between savanna and forest vegetation types, as well as across the wet and dry decade.
Annual rainfall in the NBR (Nilgiri biosphere reserve) across the 20-year study period. Data were extracted from the CHIRPS global daily precipitation rasters (Funk et al. 2015), and aggregated to obtain annual rainfall values for the entire NBR. The dashed lines represent the decadal means for the wet (blue dashed line) and dry decades (red dashed line). The black dashed line is the 20 year mean.

Methods
Study region
The Nilgiri Biosphere reserve (10.82°N to 12.15°N, 76.09°E to 77.27°E; Fig. 1), situated in the Western Ghats of India, covers an area of ~5500 km2, including multiple national parks and tiger reserves, settlements, villages and small towns. The mean annual rainfall ranges from 611 to 4179 mm. Most of the rainfall across the landscape occurs between June and September, during the Indian South-west Monsoon, which is preceded by the dry season during February to May, when most fires occur (Joshi et al. 2019). Mean annual temperatures ranged between 21.8 and 23.8°C during the study period. Average maximum temperatures ranged from 28.9 to 30.8°C (Daily aggregates, Copernicus Climate Change Service (C3S) and ECMWF; Hersbach et al. 2020). NBR has an elevational range of 80 m to 2634 m above sea level (a.s.l.). The region is home to over 4000 plant species and 700 vertebrates of which ~150 angiosperms and ~150 vertebrates are endemic (Puyravaud and Davidar 2013), and is also a refuge for endangered fauna such as the tiger, Nilgiri tahr and Nilgiri marten.
Vegetation
The area is characterised by an east-west rainfall gradient, and a variety of vegetation types including savannas, forests, agricultural fields and plantations, and contains multiple settlements, villages and towns, many connected by roads and major highways. For the purposes of this study, we used a reclassified vegetation map of the Southern, Eastern and Western portions of the NBR produced by Prabhakar and Pascal (Prabhakar and Pascal 1996). Our reclassification scheme (Fig. 1, see inset) combined the various vegetation types of the NBR into structurally and functionally distinct units known to vary in their susceptibility and responses to fire (Ratnam et al. 2011), specifically savannas and forests. Woodland scrub, tree to shrub savanna and savanna woodlands were combined and labelled ‘savanna’, which covered a total area of 2497.2 km2, while dense forest, semi-evergreen forest and dense evergreen forest constituted the areas we classified as ‘forest’, and covered a total area of 1299 km2. In addition, there were a number of ‘other’ vegetation and land-use classes in the NBR (a total area of 2523.1 km2), which we did not consider in our analyses. These include a large area of human-modified production landscapes such as agriculture, silviculture, tea estates, coffee plantations, rubber plantations and arboriculture, as well as swamps, water bodies and highly urbanised areas. All of these are unlikely to burn and were excluded from our analyses. Finally, we also excluded two ‘other’ vegetation classes, montane forest-grassland mosaics (locally called shola-grasslands) and thickets (dense shrub formations, in some cases of invasive species). Temperature and disturbance regimes that are very different from the rest of the landscape placed these vegetation classes outside the scope of this analysis.
Fire frequency and annual burned area
Fire data were obtained from the ESA’s (European Space Agency https://www.esa.int/) fire CCI 5.1 burnt area product (Chuvieco et al. 2019) based on MODerate Resolution Imaging Spectroradiometer (MODIS) annual burnt area data, accessed through Google Earth Engine (Gorelick et al. 2017). This dataset has a spatial resolution of 250 m and a temporal resolution of 8 days, and is available as a monthly summary providing the date of detection of burnt pixels and the confidence of detection as two separate bands.
Using all pixels that had a detection confidence >0, we quantified burnt area data for the peak fire season in the NBR (February–May) for each year. Pixels were assigned a 1 or 0 value depending on whether they had burnt in a given year. The total area of savannas and forests burnt in any given year was determined by summing across all burnt pixels in each habitat. Data across years were combined to calculate the total number of times each pixel burned over the 20 year period.
Long-term mean annual rainfall, vegetation type and fire occurrence
Long-term mean annual rainfall (MAR) was derived from the WorldClim 1 km × 1 km high resolution dataset (Fick and Hijmans 2017). All pixels were first categorised into 50 mm rainfall bins, ranging from 600 to 4200 mm, based on their long-term MAR. For each rainfall bin, the probability of occurrence of savanna or forest was calculated by dividing the area occupied by each vegetation type by the total area occupied by both vegetation types in that rainfall bin. With these data, we assessed the distribution of the two major vegetation types – forest and savanna – as a function of long-term MAR across the NBR.
To evaluate how MAR influenced fire occurrence, we estimated the probability of fire occurrence over the 20-year period as a function of MAR across the NBR. Probability of fire occurrence was calculated by averaging fire frequencies (number of times a pixel burned/20) across all pixels within that rainfall bin. We also calculated the proportion of savanna and forest burnt annually in each rainfall bin for both the wet (2001–2010) and dry decade (2011–2020).
For all analyses mentioned above, both vegetation and fire layers were first resampled to 1 km × 1 km to match the resolution of the MAR layer.
Relative importance of factors regulating fire in savannas and forests
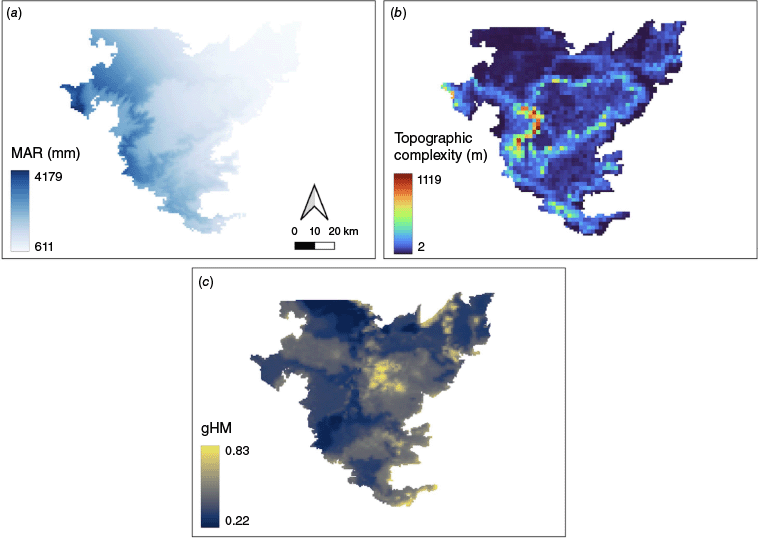
We examined the relative importance of three predictor variables – MAR, topographic complexity, and human activity and modification (Fig. 3a–c) – on fire frequency in savannas and forests. The predictor, MAR, served as a proxy for fuel accumulation, human modification for fire ignitions, and topography for fire spread and contiguity. We defined topographic complexity as the standard deviation of elevation values within a 2 km × 2 km cell. Elevation data were obtained from the Shuttle Radar Topography Mission (NASA JPL 2013; Farr et al. 2007; resolution 30 m × 30 m). Finally, human activity was indexed using the Conservation Science Partners’ Global Human Modification (CSP-gHM) (Kennedy et al. 2019) dataset – a cumulative dataset that uses 12 global metrics of human activity including agriculture, roadways and electricity distribution infrastructure accessed through Google Earth Engine (Gorelick et al. 2017). All layers were resampled to 2 km × 2 km to match the resolution of the topographic complexity layer.
Maps of three predictor variables used in our analysis: (a) MAR (mean annual rainfall) derived from WorldClim, (b) Topographic complexity showing the heterogeneity of elevation across the NBR (Nilgiri biosphere reserve), and (c) Human modification index for the NBR where values range from 0 (low human modification) to 1 (high human modification).
We used a non-parametric decision tree analysis to examine the relative importance of the above three predictors in regulating fire frequency. We used the random forest machine learning approach that constructs a multitude of uncorrelated decision trees to reach a single result. We first randomly split the dataset into training (70% of the pixels) and testing (30% of pixels) subsets. Using the training dataset we modelled fire frequency for the dry and wet decades separately, and over the entire 20 year period, as a function of the aforementioned predictors using the RandomForest package (Liaw and Wiener 2002) in R programming language (R Core Team 2024).
Our analysis was run for savanna and forest pixels separately. We plotted the variable importance values of the predictors from the random forest model for savannas and forests (over the 20 year period and for each decade separately). Importance values order predictors based on their effect on model improvement when splits are made based on the predictor, with higher importance values indicating greater relative effect of the predictor on model improvement.
We also plotted partial dependence plots for each of the three predictors to examine how they influenced fire occurrence in savannas and forests independently. Partial dependence plots graphically represent the dependence of the response on each predictor variable after accounting for the average effects of other predictors (Hastie et al. 2009). Here we show the partial dependence plots for forests and savannas across the 20 year period. Partial dependence plots for forests and savannas are also provided separately for the dry and wet decade in the supplementary information, and show similar results to the entire 20 year period.
Analyses
Analyses were conducted by importing datasets into Google Earth engine, where images were converted into comma-separated values (CSVs) files using the ‘ee.sample.Image’ function and then further analysed in R. All the analysis and plotting was done in R 4.2 (R Core Team 2024).
Results
Climate, vegetation and fire occurrence
Across the two decades of this study, incidences of fire across the NBR were highly variable, ranging from no fires in 20 years in large parts of the landscape, to as many as 9–11 fires in 20 years in a few limited hotspots (Fig. 4a). As expected, savannas were more likely to occur in the drier areas, while the opposite was true of forests (Fig. 4b), with the switch from dominance of savannas to forests occurring at ~2300 mm. (Fig. 4b). Along this same mean annual rainfall gradient, the probability of fire occurrence (calculated as the mean probability that a given pixel in that rainfall bin burns in any given year) peaked between 1800 and 2000 mm, a rainfall zone characterised by the dominance of mesic savannas. However, in absolute terms, even these peak values in mesic savannas amounted to relatively low probabilities of fire (0.06–0.075), with large parts of the landscape, notably the wetter areas dominated by forests, experiencing little to no fire (probabilities of fire ranging from 0.0 to 0.02; Fig. 4b).
(a) Map of the NBR (Nilgiri biosphere reserve) showing the number of times each pixel burned over the 20-year study period, and (b) Probability of savanna, forest and fire occurrence across the NBR as a function of MAR.

The above differences in probabilities of fire occurrence were also reflected in area of savannas and forests burnt annually, with the burnt area of savannas, in most years, being greater than that of forests, but especially so in the wet decade (Fig. 5). Critically, the burnt area of forests was uniformly low across both the wet and dry decades, whereas the burnt area of savannas was much higher in the wet than the dry decade (dashed green and yellow lines in Fig. 5).
Annual burnt area in the forests and savannas of the NBR across the 20 year study period across the wet (2001–2010) and dry decades (2011–2020).

When we further examined these patterns as a function of MAR, we found that it was mesic savannas (with MAR in the range from 1700 to 2000 mm) that experienced the greatest increases in the proportion of area that burnt in the wet decade relative to the dry decade (Fig. 6), followed by semi-arid savannas (MAR in the range from 500 to 700 mm). During the dry decade, in addition to an overall decrease in proportion of savannas that burnt, this peak of burning shifted to the wettest savannas (MAR range from 2100 to 2300 mm). In contrast to savannas, the proportion of forest area burnt did not differ consistently across the wet and dry decade (although a slight increase was noted in the wet decade for forests in the MAR range from 1700 to 2000 mm, similar to savannas). Finally, because savannas occupy a greater land area in the NBR compared to forests, even though the burnt area of savannas is much greater than forests (Fig. 5), these differences diminish when expressed as the proportions of savannas or forests that burnt (Fig. 6).
Proportion of savanna or forest burnt as a function of mean annual rainfall (extracted for 50 mm rainfall bins from WorldClim (Fick and Hijmans 2017)). Blue lines indicate the wet decade, while red lines indicate the dry decade. The vertical lines represent one standard error around the mean proportion of forest or savanna burnt in each year.

Relative importance of factors regulating fire in savannas and forests
The three predictors in our random forest model explained a greater percentage of variation in fire frequency in savannas than in forests, and in the wet decade than in the dry decade. In savannas, the predictors explained 48% of the variation in the wet decade and 28% of the variation in the dry decade. In forests, the predictors explained 35% and 8% of the variation in the wet and dry decade, respectively. For both savannas and forests, the most important predictor of fire in the wet decade was human modification index, followed by MAR (Fig. 7). However, this was reversed in the dry decade, with MAR being the most important predictor, followed by human modification index (Fig. 7). Topographic complexity remained the third most important predictor in both the wet and dry decade.
The importance value plots from the random forest analysis for savannas in the (a) wet and (b) dry decade, and forests in the (c) wet and (d) dry decade. For variable importance only the relative values matter, while the scale is biologically irrelevant.

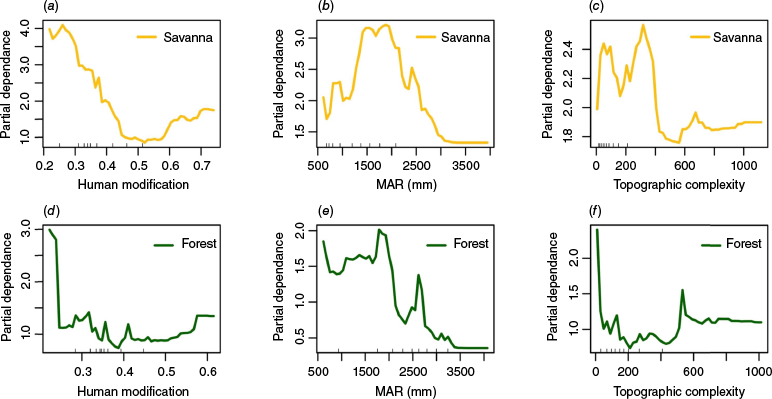
Partial dependence plots examining the effects of the three predictor variables on fire occurrence (over the 20 year period) show distinct effects of the human modification index and MAR on fires in forests and savannas (Fig. 8, Fig. S1). Low values of the human modification index were associated with higher fires in both forests and savannas. However, in forests, fires decreased sharply as human modification increased, while fires in savannas decreased more gradually as human modification of the landscape increased (Fig. 8a, d). The partial dependence plots of rainfall show that drier forests experience more fires, and fires decline in wetter forests (Fig. 8e). On the other hand, mesic savannas with intermediate rainfall experience more fires, and fires decline in very dry and very wet savannas (Fig. 8b). In both forests and savannas, fires decline with increasing topographic complexity (Fig. 8c, f).
Partial dependence plots of fire frequency on (a, d) human modification index, (b, e) mean annual rainfall (MAR) (mm), and (c, f) topographic complexity. The graphs on the top row (a–c) show results for savannas and the bottom row (d–f) for forests. The partial dependence plots show the relationship of the response to each predictor variable, averaged over the values of the other predictors. The trend, rather than the actual values, describes the relationship between predictor and response variables.
Discussion
Our results demonstrate that climate, vegetation type and human modification of the NBR landscape interact in complex ways to determine fire behaviour. Across a 20-year period, forests experienced very limited burning, regardless of whether it was a wet or dry decade. In contrast, savannas, which typically burnt more than forests, were strongly influenced by climate, burning over much greater areas (up to four times more) in the wet than the dry decade. The relative importance of climate and human modification in determining fires flipped across wet and dry decades; in the wet decade, when fuel productivity was presumably not limiting, human modification – a proxy for ignitions, was the major driver of fires, whereas rainfall became the major driver of fires in the dry decade, suggestive of fuel limitation.
Climate, vegetation and fire
As a general framework, four conditions must be met for fire to occur and spread in landscapes (Bradstock 2010). These relate to fuel loads (must be enough to carry fire), fuel moisture (fuels must be dry), ignitions (must occur when fuel is ready to burn) and fire spread (fuels must be continuous). While fuel loads are largely determined by the effects of rainfall on productivity, fuel moisture is largely a result of the length and intensity of the dry season and hence the curing rates of different types of fuel. Ignitions, on the other hand, while related to dry season lightning activity in some regions of the world, are largely anthropogenic on the subcontinent, and may increase with human presence (Saha and Howe 2003; Ratnam et al. 2016). At the same time, human activities can also fragment landscapes and disrupt connectivity, reducing the capacity for fire spread, and in turn, the overall burnt area of a region.
In this study, we interpreted long-term multi-decadal rainfall as a proxy for spatial variability in fuel loads, across savannas and forests. Variation in annual rainfall across years was interpreted as a proxy for temporal variability in fuel loads and fuel moisture across wet and dry decades. Since most ignitions are of anthropogenic origin, we used human modification index as a proxy for rates of ignition. Finally, topographic complexity and global human modification index served as indicators of ease of fire spread, with regions of greater topographic heterogeneity and higher human modification serving as barriers to the continuity of burning.
For the vegetation mosaic in the NBR we found that mesic savannas that occurred in the intermediate rainfall zone for this landscape (~1000–2100 mm) accounted for the majority of fires (Fig. 4). Previous work from the Mudumalai Tiger Reserve, a protected area within the NBR, showed that savannas and dry forests (which have grassy understoreys and we treat as savanna woodlands; see Ratnam et al. 2011), burnt more frequently and over larger areas than more moist forests (Kodandapani et al. 2004, 2008). Our results confirm that, across the NBR, the burnt area of savannas is approximately two to four times that of forests (Fig. 5). These differences in burnt area between savannas and forests can be attributed to three factors that have the potential to influence the flammability of vegetation and which vary across the different habitat types: fuel load, fuel structure and fuel moisture.
Tropical moist forests, with their shaded environments develop deep moist leaf litter layers that retard the intrusion of fire. In forests, litter moisture is directly correlated with canopy cover (Ray et al. 2005). Our analyses revealed that fires were generally rare in forests in the NBR, across both the wet and dry decades, although they increased a little in the dry decade. These data suggest that within the bounds of rainfall variation during this period, most of these forests were able to retain enough moisture to retard fires. By contrast, tropical savannas, with their open canopies, and high abundance of C4 grasses (grass species that use the C4 photosynthetic pathway to fix carbon) are known to be highly flammable (Bond 2008; Bond and Parr 2010; Ratnam et al. 2011), and this is reflected in the higher incidence of fire in savannas across the NBR. The structural attributes and productivity of the C4 grasses ensure they cure rapidly with sufficient fuel to carry fire. In the NBR, the proportion of burnt area in savannas increased with increasing MAR, up to a threshold value of ~1600 mm in the wet decade, and ~2400 mm in the dry decade (Fig. 6). Combined, these data suggest that burnt area in savannas, especially during dry periods, is determined by productivity, thus high rainfall savannas with their productive understories burn more.
Lastly, fire activity in both forests and savannas in NBR increased in the wet relative to the dry decade, with the wet decade exhibiting twice as much burnt area as the dry decade. Further, the relative importance of drivers of fire changed across these two periods. In wet years, human modification was the primary driver of fires in savannas and forests, suggesting that ignitions, rather than fuel availability, were a primary factor driving fires in these years, whereas, in the dry years, MAR, and thus fuel accumulation, was the most important factor in determining fire activity.
Human impacts on fire activity
Humans can have contrasting influences on fire frequencies and burnt area. In systems such as the NBR, where the density of lightning strikes in the dry season is low, humans are the major ignition source. People moving through the landscape, using it for purposes such as grazing or extraction of natural resources, may thus result in an increase in ignitions. On the other hand, high human presence may also result in reduced fires in modified parts of the landscape such as plantations, agricultural and pasture land. While fires may ignite in these regions, they are unlikely to spread due to barriers such as villages, roads, cleared margins around plantations and fields, and the complexity of topography. Further, livestock grazing may reduce grass and shrub fuel loads, diminishing the capacity for fire to spread.
A spatial analysis of fire in the Mudumalai Tiger Reserve in the NBR found that the probability of fire was positively related to distance from the park boundary within a 12 km buffer, suggesting that factors such as consumption of fuels by grazing livestock, and the distribution of savanna and forest at the park boundary have a strong bearing on burnt area (Kodandapani et al. 2008; also see Sankaran 2005), as has been shown in other landscapes (Lehmann et al. 2008; Holdo et al. 2009). Likewise, in a global analysis of fire, Andela et al. (2017) found that burnt area globally has declined by about 25% since the early 2000s, due to agricultural expansion and intensification, and an associated decline in the number and size of fires. Archibald et al. (2009), examining burnt area in southern Africa, found that where landscapes were already pre-conditioned to burn (due to vegetation and climate attributes) population densities greater than 10 people/km2 had a negative effect on burnt area. Indeed, this effect is also noted in the global record and over the last hundreds of years; Marlon et al. (2008) attribute a global reduction in fire activity (using charcoal as a proxy) in the past century to an increase in population density and the massive transformation of land cover and use.
Implications for fire and habitat management in NBR
Our analyses of fire across two decades in the NBR reveal some important insights, with relevance for fire and habitat management in this region. First, when placed in a global context, the proportions of annual burnt area in savannas and forests of NBR (2–10%) are very low compared to that from similar tropical savanna-forest landscapes elsewhere (up to 27% for Central India, 30% for Africa, up to 21% for Australia; Russell-Smith and Yates 2007; Zubkova et al. 2019; Pati et al. 2024). Second, it is the savanna regions of the NBR landscape, especially the mesic savannas with their productive grassy understories, that account for most of the burnt area, while burnt area in the moist forests is very low.
However, while fire is routinely understood as an integral part of the ecology of savannas elsewhere in the tropics, this understanding is notably missing from the Indian subcontinent. Despite the relatively low burnt areas recorded in these landscapes, every summer, news about ‘catastrophic’ fires that have ‘destroyed’ the ‘forests’ are all too common in local newspapers and media outlets. These are often accompanied by public outcry and dismay over ‘bad’ forest fires. Such a distorted perception of the extent of fires, which are in fact very limited in the landscape, contributes to an exaggerated sense of their negative impacts in these systems. This negative perception, combined with a failure to recognise the distinction between fire-adapted mesic savannas (which are classified as dry deciduous forests) which require burning, and moist forests from which fires should be excluded, has led to widespread official policies of fire suppression (Thekaekara et al. 2017; Rai et al. 2019). In recent years, these policies have been further strengthened by increased funding to forest departments in southern Indian states for fire-control measures. In contrast, a recent decadal-scale analysis of fires in the central Indian peninsular region (2000–2020), where many community forest reserves continue to be managed by indigenous local tribes, shows that 27–30% of this deciduous savanna-forest landscape continues to experience frequent, low-intensity fires (Kumar et al. 2022), similar to levels experienced in other savanna regions globally. These data suggest that effective fire suppression by the forest departments in the NBR may have contributed to the decreases in burnt area in this landscape in recent years.
These policies of fire-suppression are in sharp contrast to traditional fire management practices of indigenous communities across this landscape (Thekaekara et al. 2017; Rai et al. 2019). Indigenous communities across this region historically practised early dry season ‘cool’ burning, especially of mesic savannas, as a means to maintain the grassy understoreys, collect various types of non-timber forest produce and prevent intense late dry season fires. Indigenous elders from the Soliga tribe of this region describe a process where the suppression of traditional fire regimes over the last four decades has allowed the widespread invasion of a woody invader Lantana camara, which has replaced the grassy understories, and the proliferation of a parasitic mistletoe Taxillus tomentosus that infects and kills adult native trees (Rai et al. 2019). These accounts are borne out by ecological studies that link reduced fire frequencies with increases in Lantana and Taxillus proliferation (Ticktin et al. 2012; Sundaram et al. 2015). Other studies have established that native tree communities in these mesic savannas across southern peninsular India are fire-adapted (Mondal and Sukumar 2015; Ratnam et al. 2019). Likewise, a recent analysis of fire frequency and species diversity in a central Indian landscape where indigenous burning regimes are still ongoing shows that areas that have experienced frequent low-intensity fires over the past 15 years support higher tree diversity than areas that have experienced no fires over this period (Pati et al. 2024).
It is abundantly clear that controlled burning, especially in mesic savannas, and ideally with the engagement of local communities and the application of their lived knowledge, should be an integral part of the management policies for mesic savannas, to restore them to their historical, pre-invaded states (Thekaekara et al. 2017; Rai et al. 2019; Ratnam et al. 2019; Sangha et al. 2024). The data from this 20-year study suggest that at the current time, mesic savannas in this landscape probably do not experience fires at either the optimal time or frequency to maintain a healthy balance of trees and grasses. In the past, the frequent (annual, bi-annual) low-intensity, early dry season fires practiced by the indigenous communities maintained these systems in an open state (Thekaekara et al. 2017; Rai et al. 2019). However, given the widespread and intense woody invasion that has occurred since the cessation of these practices, it may be the case that occasional, high-intensity fires in the late dry season may now be required to restore the system to its more open state of the past (e.g. Murphy et al. 2023). Once this open state is restored, it may then be maintained by the indigenous burning regimes that were practised earlier. However, the experimental studies needed to test these ideas are lacking at this time.
On the other hand, our data suggest that fire-sensitive moist forests in this landscape appear to experience rare and limited fires, to which these systems, like all wet tropical forests, may be adapted (Cochrane 2003), at least under current regimes of temperature and rainfall. Fire prevention and suppression are therefore appropriate management strategies for these fire-sensitive moist forests.
Data availability
All data sets used in this analysis are publicly available (see Methods for details of where they were accessed from). Codes used for analyses will be available upon request.
Declaration of funding
This work was support by core funding to the National Centre for Biological Sciences-Tata Institute of Fundamental Research via the Department of Atomic Energy Grant RTI 4006, and grant funding from the R.M.Tulpule Charitable Trust. Supporting sources were not involved in the procurement and processing of data or preparation of the manuscript.
Acknowledgements
We thank the editors of this special issue for the invitation to contribute this paper. We thank the National Centre for Biological Sciences-Tata Institute of Fundamental Research for core funding (Department of Atomic Energy Grant RTI 4006) that supported this work. AK thanks Hindustan Uniliver Ltd. for fellowship support, MS thanks the R.M.Tulpule Charitable Trust for funding support and CL acknowledges the support provided by the Endeavour Research Fellowship, Australia.
References
Andela N, Morton DC, Giglio L, Chen Y, van der Werf GR, Kasibhatla PS, DeFries RS, Collatz GJ, Hantson S, Kloster S, Bachelet D, Forrest M, Lasslop G, Li F, Mangeon S, Melton JR, Yue C, Randerson JT (2017) A human-driven decline in global burned area. Science 356(6345), 1356-1362.
| Crossref | Google Scholar |
Archibald S (2016) Managing the human component of fire regimes: lessons from Africa. Philosophical Transactions of the Royal Society B: Biological Sciences 371(1696), 20150346.
| Crossref | Google Scholar |
Archibald S, Roy DP, van Wilgen BW, Scholes RJ (2009) What limits fire? An examination of drivers of burnt area in Southern Africa. Global Change Biology 15(3), 613-630.
| Crossref | Google Scholar |
Archibald S, Lehmann CE, Gómez-Dans JL, Bradstock RA (2013) Defining pyromes and global syndromes of fire regimes. Proceedings of the National Academy of Sciences 110(16), 6442-6447.
| Crossref | Google Scholar | PubMed |
Bond WJ, Woodward FI, Midgley GF (2005) The global distribution of ecosystems in a world without fire. New Phytologist 165(2), 525-538.
| Crossref | Google Scholar | PubMed |
Bond WJ (2008) What limits trees in C4 grasslands and savannas? Annual Review of Ecology, Evolution, and Systematics 39(1), 641-659.
| Crossref | Google Scholar |
Bond WJ, Parr CL (2010) Beyond the forest edge: ecology, diversity and conservation of the grassy biomes. Biological Conservation 143(10), 2395-2404.
| Crossref | Google Scholar |
Bradstock RA (2010) A biogeographic model of fire regimes in Australia: current and future implications. Global Ecology and Biogeography 19(2), 145-158.
| Crossref | Google Scholar |
Brando PM, Balch JK, Nepstad DC, Morton DC, Putz FE, Coe MT, et al. (2014) Abrupt increases in Amazonian tree mortality due to drought–fire interactions. Proceedings of the National Academy of Sciences 111(17), 6347-6352.
| Crossref | Google Scholar | PubMed |
Chuvieco E, Pettinari ML, Lizundia Loiola J, Storm T, Padilla Parellada M (2019) ESA Fire climate change initiative (Fire_cci): MODIS Fire_cci burned area grid product, version 5.1 [Data set]. 10.5285/3628CB2FDBA443588155E15DEE8E5352
Cochrane MA (2003) Fire science for rainforests. Nature 421(6926), 913-919.
| Crossref | Google Scholar | PubMed |
Duncan BW, Schmalzer PA (2004) Anthropogenic influences on potential fire spread in a pyrogenic ecosystem of Florida, USA. Landscape Ecology 19, 153-165.
| Crossref | Google Scholar |
Ellis EC, Klein Goldewijk K, Siebert S, Lightman D, Ramankutty N (2010) Anthropogenic transformation of the biomes, 1700 to 2000. Global Ecology and Biogeography 19(5), 589-606.
| Crossref | Google Scholar |
Farr TG, Rosen PA, Caro E, Crippen R, Duren R, Hensley S, Kobrick M, Paller M, Rodriguez E, Roth L, Seal D, Shaffer S, Shimada J, Umland J, Werner M, Oskin M, Burbank D, Alsdorf D (2007) The Shuttle Radar Topography Mission. Reviews of Geophysics 45(2),.
| Crossref | Google Scholar |
Fick SE, Hijmans RJ (2017) WorldClim 2: new 1‐km spatial resolution climate surfaces for global land areas. International Journal of Climatology 37(12), 4302-4315.
| Crossref | Google Scholar |
Funk C, Peterson P, Landsfeld M, Pedreros D, Verdin J, Shukla S, et al. (2015) The climate hazards infrared precipitation with stations—a new environmental record for monitoring extremes. Scientific Data 2(1), 1-21.
| Crossref | Google Scholar |
Gorelick N, Hancher M, Dixon M, Ilyushchenko S, Thau D, Moore R (2017) Google Earth Engine: planetary-scale geospatial analysis for everyone. Remote Sensing of Environment 202, 18-27.
| Crossref | Google Scholar |
Guyette RP, Muzika RM, Dey DC (2002) Dynamics of an anthropogenic fire regime. Ecosystems 5, 472-486.
| Google Scholar |
Hersbach H, Bell B, Berrisford P, Hirahara S, Horányi A, Muñoz‐Sabater J, et al. (2020) The ERA5 global reanalysis. Quarterly Journal of the Royal Meteorological Society 146(730), 1999-2049.
| Crossref | Google Scholar |
Hiremath AJ, Sundaram B (2005) The fire-lantana cycle hypothesis in Indian forests. Conservation and Society 3(1), 26-42.
| Google Scholar |
Hoffmann WA, Geiger EL, Gotsch SG, Rossatto DR, Silva LC, Lau OL, et al. (2012) Ecological thresholds at the savanna‐forest boundary: how plant traits, resources and fire govern the distribution of tropical biomes. Ecology Letters 15(7), 759-768.
| Crossref | Google Scholar | PubMed |
Holdo RM, Holt RD, Fryxell JM (2009) Grazers, browsers, and fire influence the extent and spatial pattern of tree cover in the Serengeti. Ecological Applications 19(1), 95-109.
| Crossref | Google Scholar |
Keeley JE, Fotheringham CJ, Morais M (1999) Reexamining fire suppression impacts on brushland fire regimes. Science 284(5421), 1829-1832.
| Crossref | Google Scholar | PubMed |
Kennedy CM, Oakleaf JR, Theobald DM, Baruch-Mordo S, Kiesecker J (2019) Managing the middle: a shift in conservation priorities based on the global human modification gradient. Global Change Biology 25(3), 811-826.
| Crossref | Google Scholar | PubMed |
Joshi AA, Ratnam J, Sankaran M (2019) Frost maintains forests and grasslands as alternate states in a montane tropical forest–grassland mosaic; but alien tree invasion and warming can disrupt this balance. Journal of Ecology 108(1), 122-132.
| Crossref | Google Scholar |
Kodandapani N, Cochrane M, Sukumar R (2004) Conservation threat of increasing fire frequencies in the Western Ghats, India. Conservation Biology 18(6), 1553-1561.
| Crossref | Google Scholar |
Kodandapani N, Cochrane MA, Sukumar R (2008) A comparative analysis of spatial, temporal, and ecological characteristics of forest fires in seasonally dry tropical ecosystems in the Western Ghats, India. Forest Ecology and Management 256(4), 607-617.
| Crossref | Google Scholar |
Kumar G, Kumar A, Saikia P, Roy PS, Khan ML (2022) Ecological impacts of forest fire on composition and structure of tropical deciduous forests of central India. Physics and Chemistry of the Earth, Parts a/b/c 128, 103240.
| Crossref | Google Scholar |
Lehmann CER, Prior LD, Williams RJ, Bowman DMJS (2008) Spatio‐temporal trends in tree cover of a tropical mesic savanna are driven by landscape disturbance. Journal of Applied Ecology 45(4), 1304-1311.
| Crossref | Google Scholar |
Lehmann CER, Anderson TM, Sankaran M, Higgins SI, Archibald S, Hoffmann WA, Hanan NP, Williams RJ, Fensham RJ, Felfili J, Hutley LB, Ratnam J, San Jose J, Montes R, Franklin D, Russell-Smith J, Ryan CM, Durigan G, Hiernaux P, Haidar R, Bowman DMJS, Bond WJ (2014) Savanna vegetation-fire-climate relationships differ among continents. Science 343(6170), 548-552.
| Crossref | Google Scholar |
Liaw A, Wiener M (2002) Classification and regression by randomForest. R News 2(3), 18-22.
| Google Scholar |
Marlon JR, Bartlein PJ, Carcaillet C, Gavin DG, Harrison SP, Higuera PE, et al. (2008) Climate and human influences on global biomass burning over the past two millennia. Nature Geoscience 1(10), 697-702.
| Crossref | Google Scholar |
McLauchlan KK, Higuera PE, Miesel J, Rogers BM, Schweitzer J, Shuman JK, et al. (2020) Fire as a fundamental ecological process: research advances and frontiers. Journal of Ecology 108(5), 2047-2069.
| Crossref | Google Scholar |
Mondal N, Sukumar R (2015) Regeneration of juvenile woody plants after fire in a seasonally dry tropical forest of Southern India. Biotropica 47(3), 330-338.
| Crossref | Google Scholar |
Murphy BP, Whitehead PJ, Evans J, Yates CP, Edwards AC, MacDermott HJ, et al. (2023) Using a demographic model to project the long‐term effects of fire management on tree biomass in Australian savannas. Ecological Monographs 93(2), e1564.
| Crossref | Google Scholar |
NASA JPL (2013) NASA Shuttle Radar Topography Mission Global 1 arc second [Data set]. Accessed 2023-05 from 10.5067/MEaSUREs/SRTM/SRTMGL1.003
Pati PK, Kaushik P, Malasiya D, Ray T, Khan ML, Khare PK (2024) Impacts of forest fire frequency on structure and composition of tropical moist deciduous forest communities of Bandhavgarh Tiger Reserve, Central India. Trees, Forests and People 15, 100489.
| Crossref | Google Scholar |
Pausas JG, Bond WJ (2020) Alternative biome states in terrestrial ecosystems. Trends in Plant Science 25(3), 250-263.
| Crossref | Google Scholar | PubMed |
Prasad VK, Badarinath KV, Eaturu A (2008) Biophysical and anthropogenic controls of forest fires in the Deccan Plateau, India. Journal of Environmental Management 86(1), 1-13.
| Crossref | Google Scholar | PubMed |
Puyravaud JP, Davidar P (2013) The Nilgiris Biosphere Reserve: an unrealized vision for conservation. Tropical Conservation Science 6(4), 468-476.
| Crossref | Google Scholar |
Puyravaud JP, Pascal JP, Dufour C (1994) Ecotone structure as an indicator of changing forest-savanna boundaries (Linganamakki region, southern India). Journal of Biogeography 21, 581-593.
| Crossref | Google Scholar |
Rai ND, Benjaminsen TA, Krishnan S, Madegowda C (2019) Political ecology of tiger conservation in India: adverse effects of banning customary practices in a protected area. Singapore Journal of Tropical Geography 40(1), 124-139.
| Crossref | Google Scholar |
Ratnam J, Bond WJ, Fensham RJ, Hoffmann WA, Archibald S, Lehmann CE, et al. (2011) When is a ‘forest’ a savanna, and why does it matter? Global Ecology and Biogeography 20(5), 653-660.
| Crossref | Google Scholar |
Ratnam J, Tomlinson KW, Rasquinha DN, Sankaran M (2016) Savannahs of Asia: antiquity, biogeography, and an uncertain future. Philosophical Transactions of the Royal Society B: Biological Sciences 371(1703), 20150305.
| Crossref | Google Scholar | PubMed |
Ratnam J, Chengappa SK, Machado SJ, Nataraj N, Osuri AM, Sankaran M (2019) Functional traits of trees from dry deciduous “forests” of southern India suggest seasonal drought and fire are important drivers. Frontiers in Ecology and Evolution 7, 8.
| Crossref | Google Scholar |
Ray D, Nepstad D, Moutinho P (2005) Micrometeorological and canopy controls of fire susceptibility in a forested Amazon landscape. Ecological Applications 15(5), 1664-1678.
| Crossref | Google Scholar |
R Core Team (2024) R: A language and environment for statistical computing. (R Foundation for Statistical Computing: Vienna, Austria) Available at https://www.R-project.org/
Romero‐Ruiz M, Etter A, Sarmiento A, Tansey K (2010) Spatial and temporal variability of fires in relation to ecosystems, land tenure and rainfall in savannas of northern South America. Global Change Biology 16(7), 2013-2023.
| Crossref | Google Scholar |
Russell-Smith J, Yates CP (2007) Australian savanna fire regimes: context, scales, patchiness. Fire Ecology 3(1), 48-63.
| Crossref | Google Scholar |
Saha S (2002) Anthropogenic fire regime in a deciduous forest of central India. Current Science 82, 1144-1147.
| Google Scholar |
Saha S, Howe HF (2003) Species composition and fire in a dry deciduous forest. Ecology 84(12), 3118-3123.
| Crossref | Google Scholar |
Sangha KK, Madegowda C, Balasubramanian M (2024) Reshaping conservation incorporating Indigenous perspectives. Global Ecology and Conservation 54, e03197.
| Crossref | Google Scholar |
Sankaran M (2005) Fire, grazing and the dynamics of tall-grass savannas in the Kalakad-Mundanthurai Tiger Reserve, South India. Conservation and Society 3(1), 4-25.
| Google Scholar |
Sankaran M, Ratnam J, Hanan NP (2004) Tree–grass coexistence in savannas revisited–insights from an examination of assumptions and mechanisms invoked in existing models. Ecology Letters 7(6), 480-490.
| Crossref | Google Scholar |
Sundaram B, Hiremath AJ, Krishnaswamy J (2015) Factors influencing the local scale colonisation and change in density of a widespread invasive plant species, Lantana camara, in South India. NeoBiota 25, 27-46.
| Crossref | Google Scholar |
Syphard AD, Radeloff VC, Keeley JE, Hawbaker TJ, Clayton MK, Stewart SI, Hammer RB (2007) Human influence on California fire regimes. Ecological Applications 17(5), 1388-1402.
| Crossref | Google Scholar | PubMed |
Takahata C, Amin R, Sarma P, Banerjee G, Oliver W, Fa JE (2010) Remotely-sensed active fire data for protected area management: eight-year patterns in the Manas National Park, India. Environmental Management 45, 414-423.
| Crossref | Google Scholar | PubMed |
Thekaekara T, Vanak AT, Ankila Hiremath J, Rai ND, Ratnam J, Raman S (2017) Notes from the other side of a forest fire. Economic & Political Weekly 52(25–26), 22-25.
| Google Scholar |
Ticktin T, Ganesan R, Paramesha M, Setty S (2012) Disentangling the effects of multiple anthropogenic drivers on the decline of two tropical dry forest trees. Journal of Applied Ecology 49(4), 774-784.
| Crossref | Google Scholar |
van der Werf GR, Randerson JT, Giglio L, Gobron N, Dolman AJ (2008) Climate controls on the variability of fires in the tropics and subtropics. Global Biogeochemical Cycles 22(3), GB3028.
| Crossref | Google Scholar |
Verma S, Singh D, Mani S, Jayakumar S (2017) Effect of forest fire on tree diversity and regeneration potential in a tropical dry deciduous forest of Mudumalai Tiger Reserve, Western Ghats, India. Ecological Processes 6, 32.
| Crossref | Google Scholar |
Viegas DX (2004) Slope and wind effects on fire propagation. International Journal of Wildland Fire 13(2), 143-156.
| Crossref | Google Scholar |
Wu C, Venevsky S, Sitch S, Mercado LM, Huntingford C, Staver AC (2021) Historical and future global burned area with changing climate and human demography. One Earth 4(4), 517-530.
| Crossref | Google Scholar |
Zubkova M, Boschetti L, Abatzoglou JT, Giglio L (2019) Changes in fire activity in Africa from 2002 to 2016 and their potential drivers. Geophysical Research Letters 46(13), 7643-7653.
| Crossref | Google Scholar | PubMed |