

Interactions of fire and forest structure in a relict mesic forest of north Africa
Peter Z. Fulé A * , Lahcen Taiqui B and Derak Mchich C
A * , Lahcen Taiqui B and Derak Mchich C
A
B
C
Abstract
Site-specific data about fire regimes and ecosystem attributes is valuable for developing conservation strategies.
We determined fire history linked to forest attributes in Talassemtane National Park, Morocco, which conserves rare species at the northern tip of Africa.
We sampled fire-scarred conifers at three high-elevation forest sites, along with forest measurements.
Surface fires recurred frequently at all sites (mean fire intervals 15–23 years) over the period 1879–2018, burning primarily in the late summer. Fires were not highly synchronous among sites and were not climate-driven, suggesting a pattern of human ignitions at times when burning was safe. Stands were dominated by large and old Pinus and Abies trees that had survived numerous surface fires. Sites where fires continued to the present had relatively open structure with few ladder fuels. The site where 36 years had passed since the last fire had a dense mid-storey of sprouting Quercus trees.
Fire regimes and forest attributes were linked with fire-quiescent periods associated with high tree density, creating ladder fuels that could support crown fire.
Maintaining frequent-fire regimes in these forests could provide benefits for reducing fuels, avoiding undesired fires under extreme conditions, and supporting adaptation to warmer climate.
Keywords: Abies marocana, climate, dendrochronology, Morocco, Pinus nigra subsp. mauretanica, Pinus pinaster, Rif Mountains.
Introduction
Interactions of fire and forest (fuel) structure are changing rapidly in many parts of the world, often with negative consequences for sustainability of socio-ecological systems because warming climate and heavy contiguous fuels support increasingly large and severe wildfires (Peñuelas and Sardans 2021; Coop et al. 2022). Fire regimes are closely connected with evolutionary history over hundreds of millions of years (He et al. 2016) and have been affected by humans throughout our evolution (Pausas and Keeley 2009). However, in the past ~150 years, land use changes associated with colonization, industrialization, and changes in human populations and their traditional practices have often led to shifts in fire regime in many parts of the world (e.g. Niklasson and Granström 2000; Coughlan 2013). In dry climates where ecosystems are adapted to frequent, low-severity fires, and the exclusion of fires can result in the accumulation of surface and canopy fuels, resulting in high-severity wildfires that often have socially undesired effects such as native plant mortality, erosion, flooding, invasion of non-native species, and loss of land-based cultural values (Stephens et al. 2014; Parks et al. 2025). Numerous other factors such as logging, grazing, planting, non-native species, climate oscillations, and human-ignited fires affect fire-forest-climate dynamics. Due to the complex interplay of these drivers, a basic initial step in developing plans for resource conservation is the development of site-specific paleoecological and historical data about fire regimes (Leys et al. 2014; Pausas 2015) and ecosystem composition, structure, and function (Quintero-Gradilla et al. 2019) to assess changes and evaluate appropriate management actions.
The nations comprising the Mediterranean Basin that link Africa, Asia, and Europe are populated with about 500 million people (GRID Arendal 2025, https://www.grida.no/resources/5900, accessed 7 January 2025). Key ecosystem services including water, biodiversity, and traditional agricultural practices are supported by seasonally dry, forested mountain ecosystems. Historical fire regimes varied by elevation and vegetation type. Widely distributed Pinus species in the region include Pinus halapensis at low, warm sites, with adaptations to stand-replacing fires such as serotinous cones; Pinus nigra and Pinus sylvestris at mid-elevation sites, with adaptations to frequent surface fires such as thick bark; and Pinus uncinata with no specific fire-related adaptations at high, cool sites where the role of fire was diminished (Keeley et al. 2011; Pausas 2015).
Frequent fire regimes have been described empirically from fire-scarred trees in P. nigra forests in Spain (Fulé et al. 2008), Greece (Touchan et al. 2012; Christopoulou et al. 2013), Corsica (Badeau et al. 2024), and Turkey (Şahan et al. 2022). All these studies showed relatively frequent fire recurrence (mean fire interval, MFI <25 years); fire regimes in a few sites persisted until the present (Şahan et al. 2022) but in most places there were interruptions to frequent burning, especially after the mid-20th century. Several studies documented an extended recent fire-free gap followed by severe landscape-scale wildfires that killed trees that had survived centuries of surface fire (Christopoulou et al. 2013; Badeau et al. 2024). Mechanisms explaining increased fire severity include higher forest density due to tree planting and abandonment of traditional agricultural practices (Ordóñez et al. 2005; Peñuelas and Sardans 2021) and warming climate (Sarris et al. 2014). In contrast, a forest preserve with widely spaced, large, old trees had high survival after a 2001 wildfire that killed surrounding young stands, illustrating the influence of fuel quantity and connectedness on fire severity (Fulé et al. 2008).
The north African border of the Mediterranean Basin has less forested area, and is located primarily in Morocco, Algeria, and Tunisia (Food and Agriculture Organization of the United Nations 2022). This region has not been the focus of studies related to wildfire compared to the European and Asian (Turkish) side. Tree-ring studies using fire-scarred trees have been published from only one study area in north Africa, with Cedrus atlantica in Algeria (Slimani et al. 2014; Kherchouche et al. 2019; Farahat and Gärtner 2024). However, the role of fire in north African ecosystems may differ in important ways from the northern side of the Mediterranean Basin. Curt, Aini, and Dupire (2020) described the current fire regime from fire records in Algeria as being linked to areas of relatively high productivity with sufficient fuel and having high ignitions in areas close to human populations. Chergui et al. (2018) also used fire records to compare the Rif Mountains in Morroco with the province of Valencia in Spain, regions with similar climate, topography, and vegetation, but different patterns of recent land use. Valencian forests became dense after the mid 20th century due to rural land abandonment and tree planting; by the end of the century, large and severe wildfires were increasingly common (Pausas and Fernández-Muñoz 2012). However, traditional pastoralism has continued in Morocco where contemporary patterns of fire size and severity are similar to the pre-1975 patterns in Valencia but significantly less than current Valencian wildfires (Chergui et al. 2018).
The Talassemtane National Park in the Rif Mountains of Morocco in north-western Africa is a high-elevation ‘sky island’ mountain landscape conserving many endangered or rare species, including the endemic Moroccan fir (Abies marocana Trab.), one of only two forests with Abies in Africa, as well as the endemic black pine of the Atlas Mountains (Pinus nigra subsp. mauretanica) and the Barbary macaque (Macaca sylvanus). The park was designated a botanical reserve in 1972, a national park in 2004, and then incorporated in the Mediterranean Intercontinental Biosphere Reserve by UNESCO in 2006 (Aoulad-Sidi-Mhend et al. 2019). Despite being a protected area, the park’s ecosystems are vulnerable to wildfire. Castro et al. (2022) used remote sensing data to assess the area burned and severity of wildfires in the park during the first 16 years of the 21 century. They found that about half the forest loss in that period was associated with fires, with the rarest endemic forest type in the region, A. maroccana/P. nigra, being disproportionately affected. A total of 302 ha (6.3% of the area) was deforested at least temporarily due to high-severity fire during the study period (Castro et al. 2022).
This study is a fine-scale companion to the landscape-scale fire assessment carried out by Castro et al. (2022). Our objectives were to: (1) focus on the A. maroccana/P. nigra forests of Talassemtane National Park at the stand level to quantify and characterize the fire regime and its changes using tree-ring dating of fire-scarred tree samples; (2) compare fire occurrence with data on climate (drought), fire seasonality, and stand- and landscape-level fire synchrony to assess the role of top-down (climate) and bottom-up (human ignitions) drivers of the fire regime; and (3) measure forest composition, tree structure, and tree age to compare these forest attributes with fire patterns, especially recent changes or interruption of the fire regime. Based on our analyses and interpretations of these data, we recommend consideration of an appropriate role for fire in the conservation of these important socio-ecological systems.
Materials and methods
Study area
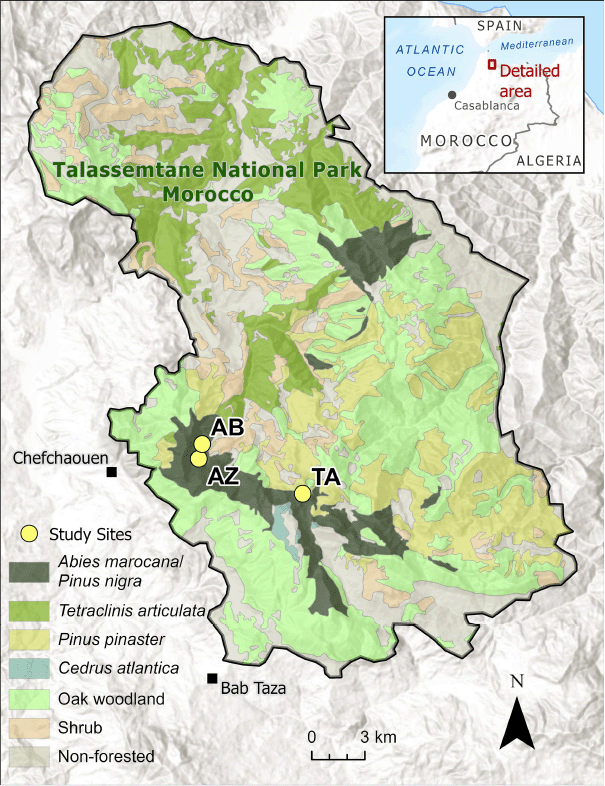
Talassemtane National Park is located in the highest peaks of the western end of the Rif Mountains in northern Morocco (Fig. 1). The land was set aside as a botanical reserve in 1972 and designated a national park in 2004 (Haut Commissariat aux Eaux et Forêts et à la Lutte Contre la Désertification (HCEFLCD) 2018). The park comprises approximately 58,000 ha, with about 75% occupied by forest lands (Haut Commissariat aux Eaux et Forêts et à la Lutte Contre la Désertification (HCEFLCD) 2018). Mesic forests of Pinus nigra, Abies marocana, and Cedrus atlantica are found only in the highest elevations, above 1400 m. The climate is Mediterranean with cool, rainy winters and warm, dry summers. There is no weather station in the high-elevation zone dominated by Abies (Sapinière) between 1400 and 2000 m, but average rainfall was estimated by extrapolation at 1705 mm per year and average temperature was estimated at 11.0°C (Haut Commissariat aux Eaux et Forêts et à la Lutte Contre la Désertification (HCEFLCD) 2018). Weather data from stations at lower elevation show an average rainfall of 1248 mm per year and average temperature of 14.9°C at the Bab Taza weather station at 880 m (Haut Commissariat aux Eaux et Forêts et à la Lutte Contre la Désertification (HCEFLCD) 2018). Average rainfall was 942 mm per year and the average temperature was 16.9°C at the weather station in Chefchaouen, at an elevation of 630 m (Haut Commissariat aux Eaux et Forêts et à la Lutte Contre la Désertification (HCEFLCD) 2018). Vegetation is distributed by elevational/climatic bands. The dominant forest types at high elevation are Moroccan fir and black pine (A. marocana and P. nigra) and maritime pine (Pinus pinaster Aiton) forest intermixed with many different oak species (e.g. Quercus rotundifolia, Quercus faginea, Quercus suber), while the lower elevation of the park is composed mostly of Tetraclinis articulata and matorral shrubs (Benabid 2000). Pinus halapensis Mill. is also found at middle and lower elevations. The park has an estimated 1380 plant species, of which 314 are endemic to Morocco and 86 are endemic to the park (Benabid 2000).
Field methods
P. nigra and A. marocana are restricted to high, mesic, and topographically complex areas of the park. We selected three study sites for sampling in March and April, 2021. Study sites were selected based on forest maps and field reconnaissance to select sites with relatively similar topography and fuel conditions, not recently killed by severe fire, and containing old trees. The three study sites were distributed in the A. maroccana/P. nigra forests of the park (Fig. 1) and ranged from 1352 m to 1600 m in elevation (Table 1). Other locations in this forest type had either been severely burned or were located on inaccessibly steep slopes or cliffs. Five fixed-area plots were installed at each site on a 120 × 120-m grid, representing a sampled area of approximately 7.2 ha per site.
| Site name (code) | Latitude (N) | Longitude (E) | Elevation (m) | Slope (%) | Aspect (°) | |
|---|---|---|---|---|---|---|
| Talaalaafid (TA) | 35.15419 | 5.14271 | 1600.4 | 49.4 | 286.6 | |
| Azilane (AZ) | 35.17214 | 5.20705 | 1352.8 | 29.2 | 13.4 | |
| Ain Boulbib (AB) | 35.17963 | 5.20502 | 1354.0 | 40 | 50 |
Geographic coordinates are the average of the five plot locations at each site.
We searched for fire-scarred trees within and between the plots, extending to the contiguous surrounding landscape. We targeted trees carrying the oldest and most complete fire records (Farris et al. 2013), predominantly P. nigra, but several Pinus pinaster trees were also sampled. Small partial cross-sections were removed from standing trees, a method that causes minimal impact (Heyerdahl and McKay 2017), as well as a few sections from downed dead trees. We collected 10 fire-scarred samples at Sites TA and AZ, and nine at Site AB, for a total of 29 samples.
Field plots were relatively large (400 m2, 11.28-m radius) to capture the compositional and structural diversity of the uneven-aged forests. For all trees over breast height (1.3 m), we recorded species and condition class (live, declining, recent snag, loose bark snag, clean snag, snag broken above breast height, snag broken below breast height, dead and down tree, or cut stump). Tree condition classes were assigned based on a tree, snag, and log classification system widely used in coniferous forests (Thomas 1979). We measured diameter at breast height (DBH) and diameter at stump height (DSH, stump height = 30 cm) for trees selected for coring, total height (m), and crown base height (m). We used increment borers to collect cores for determination of age structure from the first seven living conifers encountered on the circular plot moving clockwise from north. In addition, we cored all conifers ≥7 cm DBH to better assess the old-tree component of the forest. Trees were cored perpendicular to the slope at breast height (1.3 m).
Seedlings or sprouts of tree species shorter than breast height (1.3 m) were tallied on nested 100 m2 subplots by species and condition in three height classes: (1) ≤38 cm; (2) 38–80 cm; and (3) 80–134 cm. Downed woody biomass and forest floor depth were measured in a randomly selected direction from the center of the circular plots along a 15-m planar transect (Brown 1974) in each plot. We measured canopy cover every meter along the same 15-m transect with a vertical projection densiometer. We took photos from the north and east plot edge to the center.
Laboratory analysis
In the laboratory, we allowed tree cores to air dry in paper straws, then glued them to wooden mounts. We sanded core and fire-scar samples with progressively finer grits of sandpaper until the cells were clearly visible under magnification. We visually crossdated the tree ring series of the samples with each other and with an unpublished P. nigra chronology from Talassemtane National Park (J. C. Linares, pers. comm.). We measured ring widths of scanned samples with image analysis software (Coo Recorder, CybisElektronik & Data AB, Sweden). We used Cofecha software (Holmes 1983) for quality control of crossdating. We identified fire scar dates and intra-annual position or season of the wounds based on the relative position of fire injuries within each annual ring (Baisan and Swetnam 1990), which were noted as EE (early earlywood), ME (middle earlywood), LE (late earlywood), LW (latewood), D (dormant or ring boundary), or U (undetermined). Dormant-season scars could be formed before the start of growth in the spring or after cessation of growth in the late summer or fall. Consistent with other fire history studies on P. nigra in the Mediterranean region, we assigned D scars to the late summer/fall season corresponding to the date (year) of the adjacent latewood cells (Christopoulou et al. 2013; Badeau et al. 2024). Injuries that could not be unambiguously assigned to fire were not included in the data set.
The fire-scar data for each study site were assembled into FHX2-format files. We analyzed fire interval distributions, seasonality, and fire-climate links with Fire History Analysis and Exploration System (FHAES) software ver. 2.0.2 (Brewer et al. 2015) and the R package burnR (Malevich et al. 2018). All analyses of fire regime characteristics started from the first fire date with at least two scarred samples and at least 10% of recording trees. We measured the similarity of fire occurrence between sites and from site-region (all sites combined) using the Jaccard Index which ranges from 0 (complete dissimilarity) to 1 (complete similarity). We analyzed fire interval distributions with parametric statistics and also fit a Weibull function to express fire interval statistics in probabilistic terms. We assessed fire-climate relationships using Superposed Epoch Analysis (SEA) in the R package burnR (Malevich et al. 2018). Fire event data were compared with summer (June–August) reconstructions of the self-calibrating Palmer Drought Severity Index (PDSI) of Cook et al. (2015). We used data from gridpoint with the coordinates 35.12°N, 5.15°E.
We summarized forest structure data from plot measurements. Forest floor fuels were calculated from planar transect measurements using formulas from Brown (1974). Coefficients of specific gravity and fuel diameters were not available for the Moroccan species, so we used the values for P. ponderosa, which is morphologically similar to P. nigra and P. pinaster. We crossdated increment cores and added missing rings based on the curvature of the inner rings for samples that missed the pith (Applequist 1958).
Results
We crossdated all 29 fire-scarred samples; 25 were P. nigra, and four were P. pinaster. The fire history chart (Fig. 2) summarizes the fire-scar data. All the samples were small, partial cross-sections from living trees, except for two snags and one log. The oldest scarred tree had an inner ring date of 1744 and the oldest fire date was 1826. The time since the last fire differed at the three sites: Site TA had a last fire date of 2001 (except for a single-scarred tree in 2012), resulting in time-since-fire (TSF) of 19 years; Site AZ had a last fire date of 2018, TSF of 3 years; and Site AB had a last fire date of 1985 (except for a few single-scarred trees), TSF of 35 years.
Fire chart for 29 fire-scarred samples from the three study sites. Site codes are in Table 1.
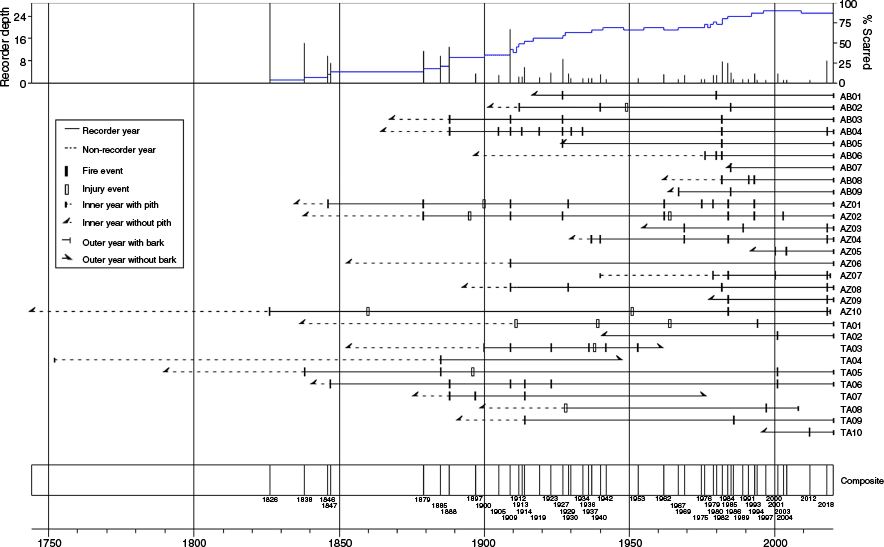
Mean fire intervals at the individual sites ranged from 15.4 years to 23.2 years (13.0–14.0 Weibull medians) (Table 2). Minimum intervals ranged from 2 years to 5 years, maximum intervals from 33 years to 78 years. Combining all sites, the mean and Weibull median were within 2 years at 7.3 and 5.7 years, respectively, with the range of intervals narrowing from 1 year to 22 years (Table 2). The fire dates at the individual sites were relatively dissimilar as measured by the Jaccard similarity index (Table 3). The highest similarity (0.222) was between Sites TA and AB. Despite the close geographical distance between Sites AZ and AB, their similarity was only 0.071.
| Site name (code) | Dates | Intervals | Mean (year) | Median (year) | s.d. (year) | WMPI (year) | Weibull s.d. (year) | Min interval | Max interval | |
|---|---|---|---|---|---|---|---|---|---|---|
| Talaalaafid (TA) | 1885–2001 | 5 | 23.2 | 9 | 31.3 | 14.0 | 26.6 | 3 | 78 | |
| Azilane (AZ) | 1879–2018 | 9 | 15.4 | 10 | 10.3 | 14.0 | 9.6 | 5 | 33 | |
| Ain Boulbib (AB) | 1888–1985 | 5 | 19.4 | 18 | 20.7 | 13.0 | 20.3 | 2 | 53 | |
| All sites | 1879–2018 | 19 | 7.3 | 6 | 6.5 | 5.7 | 6.3 | 1 | 22 |
Fire interval statistics were calculated from the first to the last fire year represented by at least two fire-scarred samples. WMPI, Weibull Median Probability Interval.
| Site name (code) | TA | AZ | AB | |
|---|---|---|---|---|
| Talaalaafid (TA) | 1.000 | 0.067 | 0.222 | |
| Azilane (AZ) | 1.000 | 0.071 | ||
| Ain Boulbib (AB) | 1.000 |
Empty cells at the bottom left of the table are duplicates of the cells symmetrically located across the central diagonal.
Fire seasonality (scar position within the growing ring) could be determined on 67% of the scars. It was strongly skewed toward latter part of the growing season, with an overall percentage of 78.1% of late-earlywood, latewood, and dormant scars. The skew was even stronger in Site TA, the highest-elevation site, where 91.7% of scars were in the latter growing season.
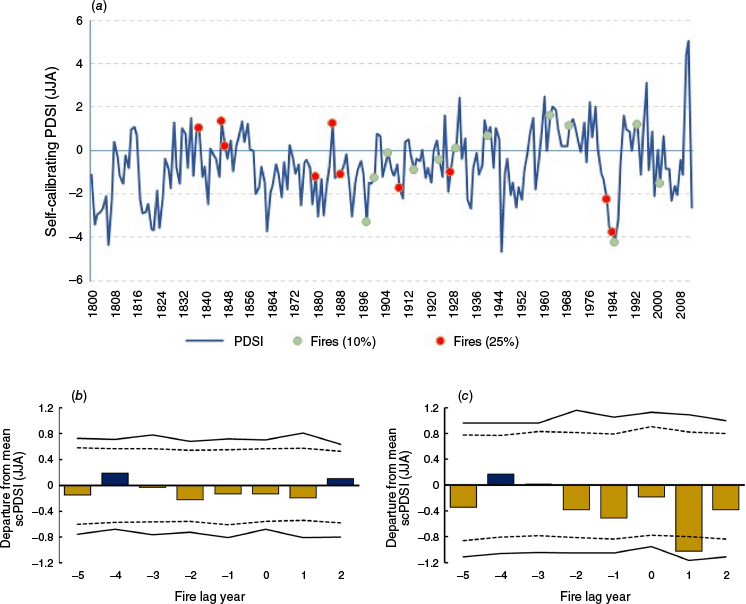
Fire-climate relationships show the occurrence of fires scarring ≥10% of trees and those scarring ≥25% of trees compared with summer PDSI (Fig. 3a). Superposed epoch analysis (SEA) with the 10%-scarred fires showed no significant relationships, as inferred from departures exceeding the 95 or 99 percentile of 1000 bootstrapped PDSI values. However, the lag year +1 after fire events were significantly dry (P < 0.05) for the 25%-scarred fires.
Fire-climate relationships. (a) Comparison of self-calibrating Palmer Drought Severity Index (scPDSI) reconstruction for June–July–August (JJA) from the Old World Drought Atlas (Cook et al. 2015), 1800–2011 C.E., with fire dates scarring 10% or 25% of the samples. (b) Superposed Epoch Analysis (SEA) illustrating departures from the mean value of scPDSI for fires scarring 10% or more of the sample trees (N = 22 fires). Horizontal dashed line represents 95 percentile, and horizontal solid line represents 99 percentile of 1000 bootstrapped scPDSI values. (c) SEA illustrating departures from the mean value of scPDSI for fires scarring 25% or more of the sample trees (N = 10 fires, 1800–2011 C.E.).
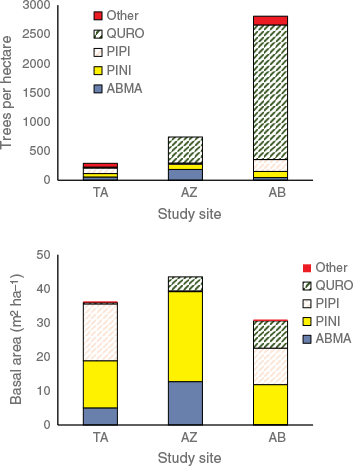
On the forest plots, we encountered six tree species plus an additional woody and shrub species in the understory; species names are in Table 4. Forest density was highly variable between sites, varying by an order of magnitude from an average of 290 trees ha−1 at Site TA to 2810 trees ha−1 at site AB (Fig. 4). The conifer species PINI, PIPI, and ABMA (see Table 4 for species codes) were fairly consistent in density and made up the majority of tree basal area (Fig. 4). The oak species QURO had widely different densities at the sites, from an average of 15 trees ha−1 in TA to 2305 trees ha−1 in AB. However, QURO trees tended to be small in size compared to the dominant conifers. Despite making up 82% of tree density at Site AB, QURO comprised only 26% of basal area.
| Species | Species code | Form | |
|---|---|---|---|
| Abies marocana | ABMA | Tree | |
| Cedrus atlantica | CEAT | Tree | |
| Juniperus oxycedrus | JUOX | Tree | |
| Pinus nigra | PINI | Tree | |
| Pinus pinaster | PIPI | Tree | |
| Quercus rotundifolia | QURO | Tree | |
| Taxus baccata | TABA | Tree | |
| Viburnum tinus | VITI | Shrub |
Forest structure at three study sites in Talassemtane National Park: tree density (upper panel); and basal area (lower panel). Site codes are in Table 1; species codes are in Table 4.
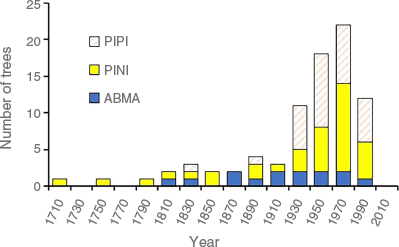
The age distribution of conifers in the Pinus and Abies genera (Pinaceae) covered 300 years (Fig. 5). The oldest tree encountered on the study sites was a PINI with a center date at coring height of 1705 C.E. The oldest ABMA dated to 1813 and the oldest PIPI to 1838.
Age distribution of conifers (Pinaceae) >7 cm DBH (all sites combined). Dates are center dates at the coring height. Species codes are in Table 4.
Forest regeneration density ranged from 3900 to 7380 stems ha−1, with high variability within as well as between each site (Table 5). The sprouting species QURO dominated regeneration at all sites, making up approximately 60% at the high-elevation (Site TA) and up to 92% at Site AZ. Conifers made up the greatest proportion of regeneration at TA. Rare species (CEAT, TABA, VITI) were each found at only one site. Regeneration densities were greatest in the 0–40 cm height class (59%), declining to 27 and 14% in the next two height classes (Table 5).
| Site name (code) | Mean density (ha−1) ± s.e. | % ABMA | % CEAT | % JUOX | % PINI | % PIPI | % QURO | % TABA | % VITI | % ABMA | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Talaalaafid (TA) | 7380 ± 1154 | 21.9 | 2.1 | 1.9 | 4.9 | 9.5 | 59.7 | 0.0 | 0.0 | 21.9 | |
| Azilane (AZ) | 4860 ± 778 | 4.7 | 0.0 | 0.3 | 0.8 | 0.5 | 92.0 | 1.8 | 0.0 | 4.7 | |
| Ain Boulbib (AB) | 3900 ± 1062 | 0.3 | 0.0 | 8.3 | 0.0 | 6.8 | 81.6 | 0.0 | 0.7 | 0.3 |
Species codes are in Table 4.
Forest floor characteristics included litter depths ranging from 0.1 cm to 2.9 cm, duff depths of 2.1–3.8 cm, and woody debris ranging from 4.7 to 15.6 Mg ha−1 (Table 6). Woody debris was distributed differently across the sites. At Site AZ, 56% of the woody debris was made up by 1–100 H timelag fuels and 49% of the coarse woody debris (1000 H timelag class) was comprised of rotten wood. At Site AZ, 34% of the woody debris was made up by 1–100 H timelag fuels and 87% of the coarse woody debris was rotten. At Site AB, 91% of the woody debris was made up by 1–100 H timelag fuels and none of the coarse woody debris was rotten. Canopy cover was relatively consistent across all three sites, 60–72% (Table 6).
| Site name (code) | Litter depth (cm) | Duff depth (cm) | 1 H (Mg ha−1) | 10 H (Mg ha−1) | 100 H (Mg ha−1) | 1000 H (Mg ha−1) | Total woody (Mg ha−1) | Canopy cover (%) | |
|---|---|---|---|---|---|---|---|---|---|
| Talaalaafid (TA) | 2.1 (0.3) | 2.6 (0.7) | 1.0 (0.3) | 4.0 (1.3) | 3.8 (1.8) | 6.9 (5.5) | 15.6 (4.5) | 60 | |
| Azilane (AZ) | 0.9 (0.2) | 2.1 (0.8) | 0.4 (0.1) | 2.7 (0.4) | 1.9 (0.8) | 9.4 (9.4) | 14.4 (9.9) | 72 | |
| Ain Boulbib (AB) | 1.9 (0.4) | 3.8 (1.5) | 0.6 (0.2) | 3.1 (1.3) | 0.6 (0.6) | 0.4 (0.4) | 4.7 (1.6) | 72 |
Columns 1–1000 H refer to timelag classes of woody fuels.
Discussion
Addressing our first research objective, which was to characterize the fire regime, we found that high-elevation conifer forests of Talassemtane National Park experienced a regime of frequent (MFI <25 years) surface fires over the past ± 140 years. Over the period 1879–2018, there was a fire every 7.3 years, on average, that scarred at least two trees in at least one of the three 7.2-ha study sites. Currently, the three sites differ in terms of time since the last fire. Site AB last burned in 1985, Site TA burned in 2001, and Site AZ burned in 2018. In broad terms, these patterns are similar in frequency to other studies in P. nigra forests of the Mediterranean Basin, but surface fire regimes in Talassemtane National Park have continued later than in many other regions. Fire regimes in P. nigra forests of Spain became less frequent after the 1960s–1980s (Fulé et al. 2008), and Greece (Christopoulou et al. 2013) and Corsica (Badeau et al. 2024) in the 1970s. In Anatolia, P. nigra fire regimes were highly variable with some sites showing fire-regime interruption as early as the late 19th century while others continued to burn up to the present (Şahan et al. 2022). In the only other dendrochronological fire history study from north Africa, surface fires in C. atlantica forests of Algeria were first interrupted in the late 19th century and no fires at all were recorded after 1930 (Kherchouche et al. 2019).
Fire regimes are highly scale-dependent (Falk et al. 2011). Our tree-ring-based fire history in this study found fire-scar evidence in 2003, 2004, 2012, and 2018. Satellite-based fire detection in Talassemtane National Park from 2003 to 2018 found fires in 2003, 2007, 2012, 2014, 2015, 2017, and 2018, but none of those fire polygons overlapped the study site locations in the current study (Castro et al. 2022). The reason for the discrepancy is that Castro et al. (2022) studied fires detected by MODIS (Moderate Resolution Imaging Spectoradiometer) satellite imagery (500-m resolution) and confirmed by pre-fire and post-fire delta normalized burn ratio (dNBR) analysis of Landsat (30-m resolution) imagery. This process depended on fires having sufficient radiative energy to be detected by MODIS and sufficient ecological effect (severity, lethality) to kill or scorch vegetation at a scale sufficient for Landsat detection. While several such fires occurred at landscape scales in the park in the 2003–2018 period, covering approximately 4670 ha in total (Castro et al. 2022), they were qualitatively different from the low-intensity surface fires detected through fire scars on living trees, showing that complex fire regimes at the landscape scale combine a range of fire sizes and effects, requiring multiple approaches for assessment (Taylor et al. 2020).
Despite high fire frequency, there were relatively few years with synchronous fires at multiple sites (Fig. 2) and an index of similarity showed little correspondence in fire dates (Table 3). The pattern of frequent but spatially isolated surface fires could be attributed in part to the severe topographic relief, sometimes bare cliffs, interrupting fuel continuity. Another possible reason for isolated fires could be human ignitions, done intentionally under conditions favorable for low-intensity burning. Intentional burning is highly linked to the expansion of agricultural activities, particularly cannabis cultivation (Chergui et al. 2024). The SEA analysis is consistent with this inference, with a minimal connection between drought (PDSI) and fire occurrence, suggesting the possibility of intentional ignitions under less-severe fire conditions. In Algeria, Kherchouche et al. (2019) described evidence of pastoralists in north Africa burning forested areas for millennia to control predators. Traditional pastoralists burned to limit forest encroachment on mountain pastures in the Pyrenées (Coughlan 2013) and to enhance forage production for livestock throughout the Mediterranean (Duffy 2019). Traditional burning often came into conflict with colonial policies that favored tree protection over pastoralism, although anti-fire policies varied in timeframe and effectiveness depending on the colonizing nation(s) and local factors (Duffy 2019).
The only SEA result that was statistically significant was a dry PDSI departure in lag year +1 post-fire. In fire regimes where climate is a major driver, the driest conditions tend to occur in Year 0, which is the fire year. Our result at Year +1 may be related to the seasonality of fires (mid- to late-summer) and the specific climate variable used. As a drought index, the PDSI incorporates lagging elements in the water balance model (Dai 2011). A fire- and moisture-based interpretation of the significant Year +1 lag in Fig. 3c is that larger fires (25%-scarred) may have tended to occur in dry periods, as indicated by the dry PDSI values in all the lags between Year −2 and Year +2. The fire year itself may have had quite dry conditions not fully reflected in that year’s PDSI value due to the lagging effect of preceding moisture, with the full effect of dryness becoming apparent in the significantly dry Year +1.
Our second research objective was to link the fire regime with forest composition and structure. Since Talassemtane National Park plays a key role in the protection of the native endemic species A. marocana and P. nigra subsp. mauretanica, the finding that all three study sites were dominated by large and old conifers of native species that were up to 300 years old, with evidence of survival of numerous forest fires, is a positive sign of the effectiveness of the park’s policies. However, it is important to remember that severe fire remains a threat, as indicated by the fire mortality reported in this vegetation type by Castro et al. (2022) and by our observation in the field of recent severe fires close to the study sites.
The vulnerability of the study sites to severe fire is most clearly connected to the time since the interruption of the frequent fire regime. All three sites had relatively similar basal areas (31–44 m2 ha−1) and canopy cover (60–72%), with fairly shallow forest floors (litter plus duff, means 3.0–5.8 cm) and relatively low woody debris fuel loading (5–16 Mg ha−1). However, tree density of small-diameter trees was exceptionally high at Site AB, which was the site with the longest time since fire (TSF) of 36 years (Fig. 6). Q. rotundifolia is an important component at the lower-elevation sites, and made up 82% of the tree density despite comprising only 26% of basal area. This species also dominated regeneration (60–92%) at all three sites. Because the sprouting capability of the species favors voluminous regeneration, the 36-year gap since the last fire at Site AB may have allowed the oaks to grow into dense stands. Dense small trees create ladder fuels that facilitate the transition of surface fires into crown fires (Menning and Stephens 2007).
Comparison of forest structure at the three study sites in the context of fire regime interruption (All photos by P. Z. Fulé).

Little data are available on the relationship of fire regime and forest structure in P. nigra, but Fulé et al. (2008) reported characteristics of open forest structure with few ladder or surface fuels in an old Spanish P. nigra forest where a frequent surface-fire regime continued up to the present. Christopoulou et al. (2013) and Badeau et al. (2024) documented severe wildfires in P. nigra forests of Greece and France in very dry years following relatively brief periods (1–2 decades) of fire exclusion. Sarris et al. (2014) suggested that extremes of dry and hot climate could override other factors, such as fuels and fire suppression, in Mediterranean forests. In pine–oak forests of North America, the pattern of low fuels associated with the frequent fire regime, and development of high ladder fuels after fire regime interruption are well documented (Barton and Poulos 2018; Huffman et al. 2020).
Our final research objective was to recommend consideration of an appropriate role for fire in the conservation of these important socio-ecological systems. Fire-related traits are deeply embedded in the evolutionary history of P. nigra and P. pinaster (He et al. 2016). Abies species tend to occur in more mesic environments but are often associated with frequent fire regimes in dry Abies forests such as in Mexico (Sáenz-Ceja et al. 2022) or the south-western USA (O’Connor et al. 2014). More broadly, fire is a common and important disturbance factor in almost all Mediterranean ecosystems (Grove and Rackham 2001), especially those where sufficient fuel loading and continuity exist (Curt et al. 2020). Thus, intentional maintenance of the surface fire regime through a program of prescribed burning and/or extensive management of wildland fires that are producing ecologically beneficial effects can be considered inherent to conserving the ecological integrity of the park.
There are practical advantages to fire use as well for controlling dense regeneration and ladder fuel development, as seen in the study site with the longest time since fire at the park. Repeated surface fires can be a practical, cost-effective, and environmentally benign form of active management (Fulé et al. 2011).
On the one hand, traditional fire knowledge is undoubtedly maintained within local communities in Talassemtane National Park (Seijo et al. 2023) and in the region as a whole (Chergui et al. 2018). On the other hand, clear challenges exist in balancing fire use with conservation of rare vegetation types (Ben-Said et al. 2022), threats of excessive burning associated with the cultivation of cannabis (Cannabis sativa) (Chouvy and Macfarlane 2018; Castro et al. 2022), and the difficulty of adjusting contemporary fire suppression policies and building expertise in fire use. As climate continues to warm in the Mediterranean Basin, it is likely that extremes of drought and temperature will become increasingly strong drivers of severe fires in P. nigra and associated forests (Sarris et al. 2014; Badeau et al. 2024). The Rif Mountains and Talassemtane National Park, which have conserved mesic northern forest types in Africa throughout the Holocene, will be tested with fire. Further study of past fire patterns and fire-climate relationships in this biodiverse region would help provide scientists and managers with data to improve conservation strategies in the face of change.
Data availability
Fire history data will be uploaded to the International Multiproxy Paleofire Database (IMPD, https://www.ncei.noaa.gov/products/paleoclimatology/fire-history). Forest structural data are available from the authors on request.
Conflicts of interest
P. Z. Fulé is an Associate Editor of International Journal of Wildland Fire but was not involved in the peer review or any decision-making process for this paper. The authors have no other conflicts of interest.
Declaration of funding
This research was funded in part by a Fulbright Scholar grant to P.Z.F. through the Moroccan-American Commission for Educational & Cultural Exchange (MACECE) and funding from the Charles O. and Mary Minor Professorship, School of Forestry, Northern Arizona University. These funds supported the stay of P. Z. Fulé in Morocco in 2021 for field and laboratory research.
Acknowledgements
We thank Elmehdi Jadar, Kaoutar Aounane, Emma Sautter, Joachim Manian, Mohamed Jadar, Gite Afeska, Abdelilah Ghallab, Amanda Stan, and Erik Schiefer for their support in this study.
References
Aoulad-Sidi-Mhend A, Maaté A, Amri I, Hlila R, Chakiri S, Maaté S, Martín-Martín M (2019) The Geological Heritage of the Talassemtane National Park and the Ghomara coast Natural Area (NW of Morocco). Geoheritage 11(3), 1005-1025.
| Crossref | Google Scholar |
Applequist MB (1958) A simple pith locator for use with off-center increment cores. Journal of Forestry 56(2), 141.
| Google Scholar |
Badeau J, Guibal F, Fulé PZ, Chauchard S, Moneglia P, Carcaillet C (2024) 202 years of changes in Mediterranean fire regime in Pinus nigra forest, Corsica. Forest Ecology and Management 554, 121658.
| Crossref | Google Scholar |
Baisan CH, Swetnam TW (1990) Fire history on a desert mountain range: Rincon Mountain wilderness. Journal of Forest Research 20, 1559-1569.
| Crossref | Google Scholar |
Barton AM, Poulos HM (2018) Pine vs. oaks revisited: Conversion of Madrean pine-oak forest to oak shrubland after high-severity wildfire in the Sky Islands of Arizona. Forest Ecology and Management 414, 28-40.
| Crossref | Google Scholar |
Ben-Said M, Linares JC, Antonio Carreira J, Taïqui L (2022) Spatial patterns and species coexistence in mixed Abies marocana-Cedrus atlantica forest in Talassemtane National Park. Forest Ecology and Management 506, 119967.
| Crossref | Google Scholar |
Brewer PW, Velásquez ME, Sutherland EK, Falk DA (2015) Fire History Analysis and Exploration System - FHAES v2.0.0. (Zenodo). 10.5281/ZENODO.34142
Castro I, Stan AB, Taiqui L, Schiefer E, Ghallab A, Derak M, Fulé PZ (2022) Detecting fire-caused forest loss in a Moroccan protected area. Fire 5(2), 51.
| Crossref | Google Scholar |
Chergui B, Fahd S, Santos X, Pausas JG (2018) Socioeconomic factors drive fire-regime variability in the Mediterranean Basin. Ecosystems 21(4), 619-628.
| Crossref | Google Scholar |
Chergui B, Fahd S, Santos X, Pausas JG (2024) Moroccan cannabis farms threaten biodiversity. Science 385(6712), 941.
| Crossref | Google Scholar | PubMed |
Chouvy P-A, Macfarlane J (2018) Agricultural innovations in Morocco’s cannabis industry. International Journal of Drug Policy 58, 85-91.
| Crossref | Google Scholar | PubMed |
Christopoulou A, Fulé PZ, Andriopoulos P, Sarris D, Arianoutsou M (2013) Dendrochronology-based fire history of Pinus nigra forests in Mount Taygetos, Southern Greece. Forest Ecology and Management 293, 132-139.
| Crossref | Google Scholar |
Cook ER, Seager R, Kushnir Y, Briffa KR, Büntgen U, Frank D, Krusic PJ, Tegel W, Van Der Schrier G, Andreu-Hayles L, Baillie M, Baittinger C, Bleicher N, Bonde N, Brown D, Carrer M, Cooper R, Čufar K, Dittmar C, Esper J, Griggs C, Gunnarson B, Günther B, Gutierrez E, Haneca K, Helama S, Herzig F, Heussner K-U, Hofmann J, Janda P, Kontic R, Köse N, Kyncl T, Levanič T, Linderholm H, Manning S, Melvin TM, Miles D, Neuwirth B, Nicolussi K, Nola P, Panayotov M, Popa I, Rothe A, Seftigen K, Seim A, Svarva H, Svoboda M, Thun T, Timonen M, Touchan R, Trotsiuk V, Trouet V, Walder F, Ważny T, Wilson R, Zang C (2015) Old World megadroughts and pluvials during the Common Era. Science Advances 1(10), e1500561.
| Crossref | Google Scholar | PubMed |
Coop JD, Parks SA, Stevens‐Rumann CS, Ritter SM, Hoffman CM (2022) Extreme fire spread events and area burned under recent and future climate in the western USA. Global Ecology and Biogeography 31(10), 1949-1959.
| Crossref | Google Scholar |
Coughlan MR (2013) Errakina : Pastoral fire use and landscape memory in the Basque Region of the French Western Pyrenees. Journal of Ethnobiology 33(1), 86-104.
| Crossref | Google Scholar |
Curt T, Aini A, Dupire S (2020) Fire activity in Mediterranean forests (The Algerian Case). Fire 3(4), 58.
| Crossref | Google Scholar |
Dai A (2011) Characteristics and trends in various forms of the Palmer Drought Severity Index during 1900–2008. Journal of Geophysical Research 116(D12), D12115.
| Crossref | Google Scholar |
Falk DA, Heyerdahl EK, Brown PM, Farris C, Fulé PZ, McKenzie D, Swetnam TW, Taylor AH, Van Horne ML (2011) Multi-scale controls of historical forest-fire regimes: New insights from fire-scar networks. Frontiers in Ecology and the Environment 9(8), 446-454.
| Crossref | Google Scholar |
Farris C, Baisan C, Falk D, Van Horne M, Fulé P, Swetnam T (2013) A comparison of targeted and systematic fire-scar sampling for estimating historical fire frequency in Southwestern ponderosa pine forests. International Journal of Wildland Fire 22(8), 1021-1033.
| Crossref | Google Scholar |
Farahat E, Gärtner H (2024) Dendrochronological studies in North Africa: reality and prospects. Frontiers in Ecology and Evolution 12, 1299472.
| Crossref | Google Scholar |
Food and Agriculture Organization of the United Nations (2022) Global Forest Resources Assessment. Global and regional estimates of carbon stocks and stock changes 1990–2020. Available at https://www.fao.org/interactive/forest-resources-assessment/2020/en/
Fulé PZ, Ribas M, Gutiérrez E, Vallejo R, Kaye MW (2008) Forest structure and fire history in an old Pinus nigra forest, eastern Spain. Forest Ecology and Management 255(3–4), 1234-1242.
| Google Scholar |
Fulé PZ, Ramos-Gómez M, Cortés-Montaño C, Miller AM (2011) Fire regime in a Mexican forest under indigenous resource management. Ecological Applications 21(3), 764-775.
| Crossref | Google Scholar | PubMed |
He T, Belcher CM, Lamont BB, Lim SL (2016) A 350‐million‐year legacy of fire adaptation among conifers. Journal of Ecology 104(2), 352-363.
| Crossref | Google Scholar |
Heyerdahl EK, McKay SJ (2017) Condition of live fire-scarred ponderosa pine twenty-one years after removing partial cross-sections. Tree-Ring Research 73(2), 149-153.
| Crossref | Google Scholar |
Holmes RL (1983) Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin 43, 69-75.
| Google Scholar |
Huffman DW, Floyd ML, Hanna DP, Crouse JE, Fulé PZ, Sánchez Meador AJ, Springer JD (2020) Fire regimes and structural changes in oak-pine forests of the Mogollon Highlands ecoregion: Implications for ecological restoration. Forest Ecology and Management 465, 118087.
| Crossref | Google Scholar |
Kherchouche D, Slimani S, Touchan R, Touati D, Malki H, Baisan CH (2019) Fire human-climate interaction in Atlas cedar forests of Aurès, Northern Algeria. Dendrochronologia 55, 125-134.
| Crossref | Google Scholar |
Leys B, Finsinger W, Carcaillet C (2014) Historical range of fire frequency is not the Achilles’ heel of the Corsican black pine ecosystem. Journal of Ecology 102(2), 381-395.
| Crossref | Google Scholar |
Malevich SB, Guiterman CH, Margolis EQ (2018) burnr: Fire history analysis and graphics in R. Dendrochronologia 49, 9-15.
| Crossref | Google Scholar |
Menning KM, Stephens SL (2007) Fire climbing in the forest: a semiqualitative, semiquantitative approach to assessing ladder fuel hazards. Western Journal of Applied Forestry 22(2), 88-93.
| Crossref | Google Scholar |
Niklasson M, Granström A (2000) Numbers and sizes of fires: long-term spatially explicit fire history in a Swedish boreal landscape. Ecology 81(6), 1484-1499.
| Crossref | Google Scholar |
O’Connor CD, Falk DA, Lynch AM, Swetnam TW (2014) Fire severity, size, and climate associations diverge from historical precedent along an ecological gradient in the Pinaleño Mountains, Arizona, USA. Forest Ecology and Management 329, 264-278.
| Crossref | Google Scholar |
Ordóñez JL, Retana J, Espelta JM (2005) Effects of tree size, crown damage, and tree location on post-fire survival and cone production of Pinus nigra trees. Forest Ecology and Management 206(1–3), 109-117.
| Crossref | Google Scholar |
Parks SA, Guiterman CH, Margolis EQ, Lonergan M, Whitman E, Abatzoglou JT, Falk DA, Johnston JD, Daniels LD, Lafon CW, Loehman RA, Kipfmueller KF, Naficy CE, Parisien M-A, Portier J, Stambaugh MC, Williams AP, Wion AP, Yocom LL (2025) A fire deficit persists across diverse North American forests despite recent increases in area burned. Nature Communications 16(1), 1493.
| Crossref | Google Scholar | PubMed |
Pausas JG (2015) Evolutionary fire ecology: lessons learned from pines. Trends in Plant Science 20(5), 318-324.
| Crossref | Google Scholar | PubMed |
Pausas JG, Fernández-Muñoz S (2012) Fire regime changes in the Western Mediterranean Basin: from fuel-limited to drought-driven fire regime. Climatic Change 110(1–2), 215-226.
| Crossref | Google Scholar |
Pausas JG, Keeley JE (2009) A burning story: the role of fire in the history of life. BioScience 59(7), 593-601.
| Crossref | Google Scholar |
Peñuelas J, Sardans J (2021) Global change and forest disturbances in the Mediterranean Basin: breakthroughs, knowledge gaps, and recommendations. Forests 12(5), 603.
| Crossref | Google Scholar |
Quintero-Gradilla SD, Jardel-Peláez EJ, Cuevas-Guzmán R, García-Oliva F, Martínez-Yrizar A (2019) Cambio postincendio en la estructura y composición del estrato arbóreo y carga de combustibles en un bosque de Pinus douglasiana de México. Madera y Bosques 25(3), 1-14.
| Crossref | Google Scholar |
Sáenz-Ceja JE, Martínez-Ramos M, Mendoza ME, Pérez-Salicrup DR (2022) Fire scar characteristics in two tropical montane conifer species from central Mexico. International Journal of Wildland Fire 31(7), 684-692.
| Crossref | Google Scholar |
Şahan EA, Köse N, Güner HT, Trouet V, Tavşanoğlu Ç, Akkemik Ü, Dalfes HN (2022) Multi-century spatiotemporal patterns of fire history in black pine forests, Turkey. Forest Ecology and Management 518, 120296.
| Crossref | Google Scholar |
Sarris D, Christopoulou A, Angelonidi E, Koutsias N, Fulé PZ, Arianoutsou M (2014) Increasing extremes of heat and drought associated with recent severe wildfires in southern Greece. Regional Environmental Change 14(3), 1257-1268.
| Crossref | Google Scholar |
Seijo F, Linares JC, Sánchez-Salguero R, Taiqui L, Zavala MA (2023) Cultural dimensions of forest conservation under global change: the case of relict Mediterranean fir forests. Landscape Ecology 38(12), 3675-3694.
| Crossref | Google Scholar |
Slimani S, Touchan R, Derridj A, Kherchouche D, Gutiérrez E (2014) Fire history of Atlas cedar (Cedrus atlantica Manetti) in Mount Chélia, northern Algeria. Journal of Arid Environments 104, 116-123.
| Crossref | Google Scholar |
Stephens SL, Burrows N, Buyantuyev A, Gray RW, Keane RE, Kubian R, Liu S, Seijo F, Shu L, Tolhurst KG, Van Wagtendonk JW (2014) Temperate and boreal forest mega‐fires: characteristics and challenges. Frontiers in Ecology and the Environment 12(2), 115-122.
| Crossref | Google Scholar |
Taylor AH, Airey-Lauvaux C, Estes B, Harris L, Skinner CN (2020) Spatial patterns of nineteenth century fire severity persist after fire exclusion and a twenty-first century wildfire in a mixed conifer forest landscape, Southern Cascades, USA. Landscape Ecology 35(12), 2777-2790.
| Crossref | Google Scholar |
Touchan R, Baisan C, Mitsopoulos ID, Dimitrakopoulos AP (2012) Fire history in European black pine (Pinus nigra Arn.) forests of the Valia Kalda, Pindus Mountains, Greece. Tree-Ring Research 68(1), 45-50.
| Crossref | Google Scholar |