

Distribution, composition and environmental correlates of high-altitude gravelly pavement herbfields in north-eastern Victoria, Australia
Alexandra Blackburn-Smith A B * and John W. Morgan A B
A B * and John W. Morgan A B
A Research Centre for Applied Alpine Ecology, La Trobe University, Bundoora, Vic. 3086, Australia.
B Department of Environment and Genetics, La Trobe University, Bundoora, Vic. 3086, Australia.
Australian Journal of Botany 70(4) 263-274 https://doi.org/10.1071/BT21113
Submitted: 15 September 2021 Accepted: 20 May 2022 Published: 28 June 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Gravelly pavement herbfields are a rare, geographically restricted community found in high mountain areas of south-eastern Australia. Gravelly pavement plant communities occur where there is continuous waterflow, with vegetation comprising sparse, semi-aquatic species on a rock or gravel substratum.
Aims: This study aimed to further the understanding of the community on the Bogong High Plains, Victoria, by asking questions relating to their location, structure and floristic composition. In particular, we ask what determines the distribution of pavement communities at the landscape-scale; and what abiotic factors govern between- and within-site vegetation patterns?
Results: Gravelly pavement communities were associated with groundwater outflows and are likely to be formed by an interaction between surface waterflow and slope. This work adds to the current understanding of wetland types in high-mountain Australia because, to our knowledge, this relationship has not been previously described. Floristic patterns vary with slope and elevation, with high-elevation pavements on steep slopes being floristically dissimilar to those at lower elevation on flatter slopes. Species richness was consistent across sites and only a few species were found at high abundance. Within-site patterning is apparent in most pavements, attributable to abiotic factors such as rockiness and water availability, leading to characteristic vegetation patterns. Species such as the sedge Oreobolus pumilio occur on the margins of pavements where water levels are variable, whereas forbs such as Psychrophila introloba are typically found in the middle of pavements where there is constant water supply.
Conclusions: Gravelly pavement communities are groundwater-dependant ecosystems, restricted at the landscape-scale and contain many rare species and, as such, deserve higher levels of protection than is currently offered.
Keywords: alpine, groundwater-dependant ecosystem, herbfield, species distribution, sub-alpine, threatened species, vegetation patterns, wetlands.
Introduction
Australian alpine and subalpine environments are mostly restricted to south-eastern Australia (Venn et al. 2017). Australia’s mainland high mountains are relatively low in elevation, lack a nival zone and are mostly covered in soils (Costin et al. 2000; Venn et al. 2017). Alpine ecosystems encompass several plant communities characterised by species and growth forms that reflect differences in soil depth, topography and drainage (Venn et al. 2017). Wetland complexes, for example, develop when water is close to the ground surface for at least part of the year, typically next to streams, along valley floors and in areas of groundwater seepage on hillsides (Wahren et al. 1999). Australia’s mainland alpine wetlands consist of a number of well known, and extremely rare, vegetation complexes, including Sphagnum-dominated peatlands (also known as bogs) and snowpatches (Wahren 1997; Wahren et al. 1999, 2001a, 2001b; McDougall and Walsh 2007; Shannon and Morgan 2007). Sphagnum peatlands occupy valley bottoms, steam banks and seepage areas, where peaty soil is waterlogged for at least part of the year, contributing significantly to the local hydrology (Wahren et al. 2001b). Snowpatch vegetation occurs on the southern/south-eastern aspect of steep slopes at high altitudes, where snow accumulates and persists well into the summer season and snowmelt provides water to the community during the drier periods (Wahren et al. 2001a). Vegetation is typically small-statured, herb-dominated and sparse (McDougall and Walsh 2007).
Some wetland communities can be found in areas where soils are poorly developed, or hardly exist at all, existing on rocky pavements. Rock-dominated ecosystems are rare on mainland alpine Australia, yet are common in Tasmania and elsewhere in the world, e.g. fellfields, screes and snowbeds (Mark and Bliss 1970; Valachovič et al. 1997; Björk and Molau 2007; Rundel and Millar 2016). Given most alpine vegetation on mainland Australia occurs on soil, plant communities that occur in areas that lack soil are therefore particularly unusual.
One example of rock-dominated wetland communities in Australia is ‘gravelly pavement herbfields’ (sensu McDougall and Walsh 2007). The Plantago muelleri–Montia australasica alliance (Costin 1972), Oreobolus pumilio association (McVean 1969), Caltha introloba herbland, Unit 10 (McDougall 1982) and short alpine herbfields (Costin et al. 2000) are all plant communities that can occur on gravelly pavements. The vegetation of gravelly pavements on the Bogong High Plains, Victoria, is a very short, sparse herbfield-dominated by either Psychrophila introloba (formally Caltha introloba) or Oreobolus pumilio subsp. pumilio (Fig. 1). Sparseness of vegetation, and a lack of soil, is maintained by constant water flow from permanent groundwater springs, hillside seepage and late-lying snowmelt (Wahren 1997; McDougall and Walsh 2007; Tolsma and Wahren 2016).

|
The origin of this community is uncertain. On the Kosciuszko Main Range, gravelly pavements on flat slopes have been interpreted as the product of wetland vegetation erosion caused by free-ranging cattle (Wimbush and Costin 1979; Wahren et al. 1999; Tolsma and Wahren 2016). These pavements are typically on slopes of less than two degrees, have small gravel-like stones and are found along valley floors (Wimbush and Costin 1979; Wahren et al. 1999). By contrast, gravelly pavements on steeper slopes, with larger rocks, occurring at the head of a water source are hypothesised to be of periglacial origin (Wahren et al. 1999). In many instances, the vegetation of gravelly pavements has often been subsumed into other plant communities such as snowpatch or Sphagnum peatland vegetation (Costin et al. 2000; Tolsma and Wahren 2016), although the validity of this has been questioned (Tolsma and Wahren 2016).
Tolsma and Wahren (2016) identified patches of the Caltha introloba herbland community occurring on gravelly pavements in Victoria, ranging in size from <100 to >1500 m2 (totalling 1.8 ha), but did not quantify the within- and between-patch floristic variation. In this study, we build on the work of Tolsma and Wahren (2016) to better understand the ecology of gravelly pavement communities on the Bogong High Plains, Victoria. To do this, we ask the following: where does this community occur; how does floristic composition of pavements vary across the landscape; what abiotic correlates contribute to floristic variation within-and across-sites; and what are the patterns of species rarity in gravelly pavements?
Materials and methods
Study region
The study was conducted on the Bogong High Plains (BHP), between Mount Nelse North and Mount Cope, ∼320 km north-east of Melbourne, Victoria, Australia. Pavements have been mapped across this landscape (Tolsma and Wahren 2016), but little is known about their community composition. The region is characterised by low mean monthly daily maximum temperatures (2.5–18.9°C), frequent frosts (>100 per annum, that can occur at any time of the year) and high precipitation (>2000 mm per annum), much of which falls as snow (Williams and Costin 1994; Wahren 1997; Bureau of Meteorology 2020). Soils across the landscape are humified or raw peat, or organic loams (of variable depth) and are highly acidic (Northcote 1979; Williams and Costin 1994; McDougall and Walsh 2007). In sheltered positions, snow persists well into the summer season (December/January; Williams and Costin 1994; Wahren 1997).
The BHP has a long history of free-ranging livestock grazing, with the first cattle being introduced to the area in the 1850s (Lawrence 1995). Livestock grazing levels have been variable across time and study sites (Wahren et al. 1999), with grazing gradually being banned from the BHP between 1972 and 2005 (Lawrence 1995; Williams et al. 2014). Some pavements on the BHP have been offered intermittent protection from feral ungulates through the use of exclusion fencing, put in place to investigate the effects of cattle. The extensive pavements in New Species Gully were fenced in 1980 (Wahren et al. 1999), and a small pavement in nearby Cope Creek was fenced in 1981 (Van Rees 1984). The fences were removed after cattle grazing ceased in 2005. Large ungulates, specifically Sambar deer and feral horses, have increased in numbers in the past decade, becoming more prevalent in high-altitude areas of south-eastern Australia, including on the BHP (Brown et al. 2016; Tolsma and Shannon 2018).
Study sites
Nineteen study sites were selected across the BHP to examine the spatial patterns and floristic composition of pavements, and to quantify their abiotic setting (Fig. 2). The current study focused on some of the larger sites identified by Tolsma and Wahren (2016). Sites were distributed across five large catchments. Eleven sites have been a part of a long-term monitoring program for over 30 years, whereas the remaining eight sites were selected on the basis of prior knowledge of their location. Study sites are distributed over a 200 m elevation gradient, encompassing both alpine and subalpine habitat zones.

|
Within a pavement, variation in slope and aspect is frequent, resulting in substantial variability of micro-habitats. To characterise this within-pavement heterogeneity, and to identify the drivers of within-pavement floristic variation, ‘upper’, ‘middle’, ‘lower’ and ‘edge’ zones were designated on the basis of visual variation in abiotic conditions. Floristic and abiotic sampling was conducted within these zones. Wahren et al. (2001b) used a similar sampling approach to assess zonal variation in snowpatch vegetation.
Location and abiotic setting of pavements
The location of all study sites was recorded using a GPS (datum: WGS84). Size, position of the site within the landscape (i.e. on the valley bottom or hillside), aspect (using a sighting compass), slope (using a sextant) and surrounding vegetation type (i.e. snowpatch, closed heath, Sphagnum peatland) of each pavement were recorded. These measurements were recorded for each pavement as a whole and within each zone of each pavement.
Floristic composition of pavements
In February 2020, the within-pavement floristic variation was characterised by sampling eight 0.25 m2 quadrats in each zone of each pavement. Quadrats were sampled haphazardly within each zone, the location being determined by a blind throw. All species present, including bryophytes, and their percentage cover were recorded. Vascular species were identified in the field and all bryophytes were sent to Royal Botanic Gardens, Sydney, for confirmation of identification.
Within-site patterning of vegetation
To identify potential abiotic drivers of floristic variability among the different zones of a pavement, five 10 × 10 cm subquadrats within each floristic quadrat were sampled and substratum type and cover were recorded. Percentage cover of silt, sand, gravel, rock, boulder, bedrock and vegetation were recorded.
To quantify the nature of the zone moisture status, in the same subquadrats as above, water ‘type’ was recorded as running, submerged, damp, or dry. As water measurements were recorded only once in the study (March 2020), this measurement can be used only as an indication of water flow through quadrats at that specific point in time. Further study is needed to explore the spatial relationship between species distribution and temporal water availability in pavements.
Data analysis
Floristic composition of pavements
Both presence/absence and percentage cover data were used to analyse variation in floristic composition among sites. To analyse cover data, sampling points (individual quadrats) were pooled by site and mean species cover for each site was calculated. Non-metric multidimensional scaling (NMDS) ordination was then performed, using the Bray–Curtis dissimilarity matrix, to analyse the floristic variation among all 19 sites. For presence/absence data, another NMDS ordination was performed, using the Jaccard similarity coefficient, to analyse changes in floristic composition among the 19 sites.
For both analyses, the relationship between floristic variation and abiotic factors was examined by fitting vectors of maximum correlation. Environmental vectors included pavement size, elevation, slope, aspect and rock cover; only environmental vectors with P < 0.05 were plotted onto the ordination. For this analysis, rock cover value was calculated as a percentage of quadrats per site that had greater than 60% rock cover. Aspect was converted to degree of northness, which allows aspect to be linearly plotted as a gradient between 1 (due north) and −1 (due south). Indicator species were plotted onto the ordination when P < 0.01. Vector fitting allows a visual representation of the degree of correlation between the configuration of the ordination (how similar samples are) and the environmental variables (size, elevation, slope, aspect and rock cover). Vectors are interpreted as increasing at sites in the direction of the arrow and decreasing at sites in the opposite direction. All analyses were conducted in R, using the packages vegan (Oksanen et al. 2019), BiodiversityR (Kindt and Coe 2005) and ggplot2 (Wickham 2016).
A rank-abundance curve was used to examine patterns of relative species abundance for all sites in this study. Individual site data were pooled, and mean cover of each species was calculated for the study. Rank abundance was then calculated and plotted in BiodiversityR as per Kindt and Coe (2005).
Within-site patterning of vegetation
The primary aim of analysing the differences in zones within pavements was to investigate how differing abiotic conditions affect local species assemblage. Three sites were omitted from this analysis because no zonation was identified in the field because of their small size.
Mean species cover was calculated for the upper, middle, lower and edge zones for each site, and analysed with NMDS ordination using the Bray–Curtis dissimilarity matrix as described above. Analysis of similarities (ANOSIM) was used to test whether vegetation was significantly different in each a priori-defined zone. Relationships between within-zone species distribution and environmental gradients were analysed by fitting vectors of maximum correlation. For this analysis, environmental vectors included slope, aspect, water cover and rock cover. Again, environmental vectors were plotted only where P < 0.05 and indicator species where P < 0.01.
Results
Location and abiotic setting of pavements
Sites were found between 1633 and 1847 m above sea level; 10 pavements were located above the treeline (alpine) and nine pavements were found within the subalpine zone. Pavement size ranged between 40 and 1295 m2. Slope ranged between 2 and 21°. Pavements occurred on all aspects; northern, eastern and south-eastern aspects were the most common, with four pavements on each. Eight sites were associated with areas of late-lying snow and all 19 sites were found in close proximity to a groundwater spring. A summary table of the abiotic setting of each site can be found in Supplementary Table S1.
Floristic composition of pavements
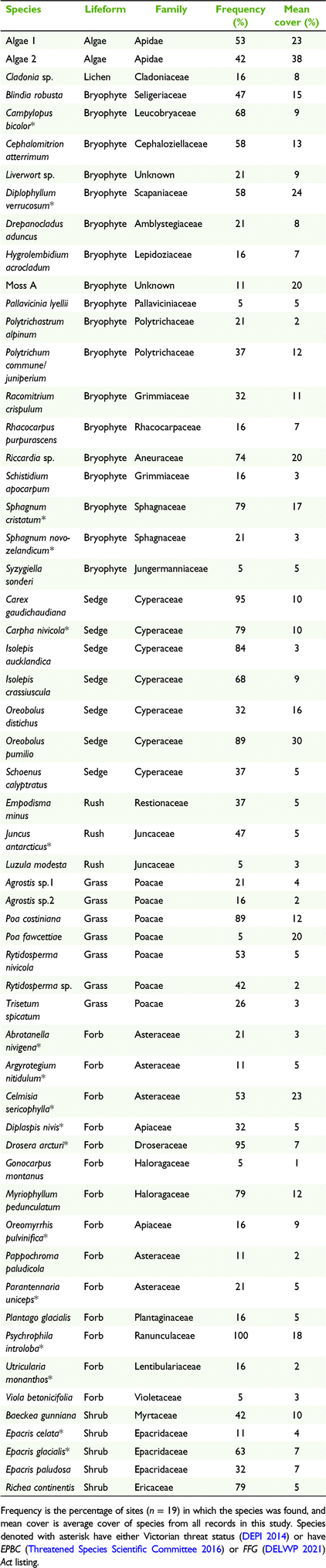
Fifty-seven species were recorded from the 19 sites on the BHP (Table 1). This included 18 bryophytes, 17 monocots, 14 forbs, five shrubs, two algae and one lichen species. Psychrophila introloba was found in every site and Oreobolus pumilio had the highest mean cover of the vascular species (30%; Table 1). Fifteen species are listed as rare, vulnerable or endangered in Victoria, with five being listed as protected under the state endangered species legislation (Flora and Fauna Guarantee (FFG) Act) and one under the national legislation (Environment Protection and Biodiversity Conservation (EPBC) Act). Three bryophytes are listed as ‘poorly known’. No exotic species were recorded. A rank-abundance curve highlighted that across all pavements, only three species had relative abundance higher than 50% (O. pumilio, P. introloba, Carex gaudichaudiana) and more than half the species had a relative abundance less than 10% (Fig. 3).
|

|
Among-site variation of floristic composition
There were no clear groupings of pavements from across the BHP landscape seen in the ordination of sites on the basis of percentage cover or presence/absence data (Fig. 4). However, sites were spread in the ordination space, suggesting that composition and relative cover vary across them. Slope was a significant determinant of floristic variation among sites in the ordination, using both presence/absence and percentage-cover data (Fig. 4). Most floristic variation occurred parallel with the slope vector. Elevation was also a significant determinant of floristic variation among sites when analysing presence/absence data. A summary of maximum correlation of environmental variables can be found in Table 2. A summary of maximum correlation of indicator species can be found in Tables S2 and S3.

|

|
Within-site patterning of vegetation
A summary of the ordination results from each site can be found in Tables S4 and S5. ANOSIM was used to test whether the within-pavement zones were significantly different from each other. In four cases, there was no significant difference between the floristic composition of each of the pavement zones (P > 0.05; Table S4). Rock cover was a significant environmental correlate of within-site floristic variation (12 of the 16 sites had significant correlations; P < 0.05). O. pumilio was a key indicator species (P < 0.01) in the distinction of zones in 10 of the 16 sites analysed for within-site patterning (Table S5).
Ordination results from the site ‘WRC B’ are shown (Fig. 5) and used as an example for trends consistently seen in pavements in this study. ANOSIM of species composition among the zones showed that there was a significant difference in their composition (R2 = 0.473, P = 0.001). Lower levels of rock and water cover distinguished between the edge zone and the remaining zones (upper, middle and lower). P. introloba was an important species in the separation of middle and lower stratification and O. pumilio, Drosera arcturi and Sphagnum cristatum were significant in the separation of edge groups. Edge pavement vegetation was responding to lower rock and water levels (Fig. 6). This pattern was repeated in many sites, as seen in Fig. 6. (ANOSIM analysis of groupings; (a) WRC Z1 R2 = 0.499, P = 0.001, (b) Ruined Castle R2 = 0.617, P = 0.001, (c) New Species R2 = 0.353, P = 0.001, (d) Nelse B R2 = 0.329, P = 0.001).

|

|
Discussion
We build on prior research on gravelly pavement communities that identified the location of the community across the Bogong High Plains (Tolsma and Wahren 2016) but provided little insight into their floristic composition and variation. We found that pavements occur in a variety of landscape positions. Many were not closely associated with areas of late-lying snow; rather, all were associated with a source of water such as a groundwater spring, providing support for the identification of a plant community that has previously been unrecognised as a groundwater-dependant ecosystem formed by the interaction of groundwater-derived surface-water flow and slope. Slope and elevation were significant contributors to floristic variation among pavements. There was within-site floristic patterning in most sites correlated to varying levels of rock and water cover. There were very few species found in high abundance, with a large number of infrequent species being observed. Species composition varied subtly across sites on the BHP.
Location and abiotic setting of pavements
Pavements occur in a variety of landscape settings. Unlike some wetland systems, such as snowpatch wetlands that always occur on steep, south-east-facing slopes (Wahren et al. 2001b), pavements occupy a variety of slopes and aspects, confirming that pavements are not a variant of snowpatch vegetation (Tolsma and Wahren 2016). The presence of a groundwater source, particularly a groundwater spring, is a common factor across pavements on the BHP. Water outcrops occur when groundwater is pushed to the surface (McCartney et al. 2013), providing pavements with constant irrigation through the snow-free period. Importantly, many pavements occur directly below outcrops/sources where consistent overland water flow occurs. This area, with flowing surface water, has been shown to be a hostile environment to the establishment of the wetland moss Sphagnum (Wahren 1997) that dominates further downslope, and may explain why a collection of small-statured, uncommon species, encompassing bryophytes, monocots and forbs, occupies these localised places. Where water outflows are low and water collects in pools, the alpine moss Blindia robusta dominates (McCartney et al. 2013; Clements et al. 2016). Further downslope, where the topography typically flattens and water flows are very slow, peat-forming wetlands dominated by Sphagnum cristatum, Carex gaudichaudiana and Empodisma minus occur (McDougall and Walsh 2007). Hence, pavement communities occupy a zone between source pools and peat-forming wetlands (Fig. 7), a relationship not previously quantified.

|
Floristic composition of pavements
Of the 57 species recorded in this study, approximately one quarter are listed in Victoria as endangered, vulnerable, rare or poorly known (DEPI 2014). Some species, such as Plantago glacialis and Abrotanella nivigena, are entirely restricted to pavements, indicating that pavements are crucial to their ongoing persistence on the BHP. It would not take much for the local, and indeed state-wide, extinction to occur for such species.
The most speciose lifeform of pavements was bryophytes, with 18 species recorded. There are approximately 110 bryophyte species indigenous to the BHP (Meagher et al. 2002); hence, ∼16% of the total bryophyte species on the BHP are found in pavements. High bryophyte species richness is common in other high-mountain wet rocky areas such as fell fields, screes and snowbanks (Billings and Mooney 1968; Geissler 1982). Despite alpine and subalpine areas being considered a ‘hotspot’ of bryophytes and bryophyte conservation, very little is known about the ecology of bryophytes in high mountain areas of Australia (Meagher et al. 2002). Further investigation is needed to understand the distribution of these pavement bryophyte species which, in turn, may add to the pool of rare species found in pavements.
Among-site variation of floristic composition
Slope was a significant determinant of floristic variation among pavement sites in this study, drawing a strong line between two suites of species, namely, those indicative of steeper slopes and those indicative of more gentle slopes. Because of the great variation of slopes on which pavements are found (2–21°), it is likely that only some species can withstand the increased erosional pressure of water and snow experienced on steeper slopes. Elevation was also a significant determinant of floristic variation among pavement sites when analysing presence/absence data. Most floristic variation was orthogonal to the elevational vector. Species most responding to high elevations were some of those entirely restricted to pavements, and found only on the steepest, highest sites, such as, for example, P. glacialis and Oreomyrrhis pulvinifica. Sphagnum cristatum was common in the lowest-elevation pavements on flatter slopes. There was no significant difference in vegetation when pavements were grouped by water catchment (data not shown). If pavements were recent features of the landscape, it is likely that species composition would be grouped by water catchment because of their apparent limited capacity for dispersal. Most species appear to reproduce vegetatively, via rhizomes or through dispersal of dislodged fragments that are able to re-root and grow (A. Blackburn-Smith, pers. obs).
Species richness was consistent across pavements but showed some effects of pavement size; the smallest pavement had 20 species, whereas one of the largest pavements had 34 species. The rank-abundance curve highlights that, across all sites, there are very few species that are frequent across the landscape of pavements (relative abundance >50%); rather, the gravelly pavement species pool consists of a long tail of increasingly infrequent species. This implies that gamma diversity relies on the presence of many pavements, not just one or two.
Within-site patterning of vegetation
Within-site floristic patterning occurs in response to local abiotic drivers. The predominant factors influencing vegetation patterns at pavement sites were hypothesised to be moisture gradients and the amount of rock cover. Both of these were important in explaining local patch distributions. Zonation in relation to changes in abiotic environments is a feature of snowpatch vegetation in both New South Wales and Victoria (Atkin and Collier 1992; Wahren et al. 2001b; Pickering et al. 2014). In snowpatch vegetation, the dominant factor influencing floristic variation is the length of the growing season and moisture gradients, leading to upper zones typically being drier than those lower down the slope (Wahren et al. 2001b). Pavements have a similar pattern; however, owing to the typically concave shape of a pavement, moisture flows to the centre and lower zones, leaving the edges drier.
Pavement-edge zone had lower water levels and lower rock cover (i.e. higher vegetation cover). The indicator species for this zone was consistently O. pumilio. Leaf dry-matter content (LDMC) measures the ability of a leaf to retain water and is used as an indicator of plant stress-tolerance. High LDMC of O. pumilio suggests that this species is stress-tolerant (A. Blackburn-Smith, unpubl. data). Thus, the dominance of this species on the edge zone is probably due to its ability to withstand varying water levels, as well as desiccation, throughout the growing season. Upper, middle and lower zones of pavements were more similar to each other, with higher levels of water cover and rock cover (i.e. less vegetation cover). Indicator species for these wetter zones included P. introloba, C. gaudichaudiana, Myriophyllum pedunculatum and algae species. These species are not restricted to pavements and are consistently found in other areas of high moisture across the BHP, including fens and wet heathland (McDougall and Walsh 2007). LDMC observations of P. introloba suggested that, compared with O. pumilio, this species is not stress-tolerant (it has a low LDMC) and is likely to require constant water to survive (A. Blackburn-Smith, unpubl. data).
Rock cover is the main environmental correlate of floristic variation on a pavement. Species found in the edge zone of pavements are typically spreading and mat-forming, resulting in a lower cover of rock, i.e. mat-forming sedge O. pumilio, which can act as a facilitator of colonisation for S. cristatum and D. arcturi, leading to even greater vegetation cover and less rock cover. Species in wetter areas (upper, middle and lower zones) are typically sparser because they must be able to survive the erosional force of constant running water. For example, P. introloba and Myriophyllum pedunculatum appear particularly well adapted to withstand the constant erosional force of running water; P. introloba has a long taproot that anchors deep into the substratum (Wardlow et al. 1989) and M. pedunculatum hugs the substratum and roots at the nodes to anchor in.
Future research and management implications
Gravelly pavement herbfields on the BHP were previously overlooked as groundwater-dependant ecosystems; the consistent supply of water to pavements from a groundwater source seems the key feature defining their distribution in the landscape, their size and within-site vegetation patterning. The interaction between spring and slope is the most likely mode of origin for these communities, with water acting as an erosional force, creating a community of sparse, small-statured vegetation, similar to the role of wind in the formation of fjaeldmark communities (Costin et al. 2000). An investigation into the type and amount of water coming into these systems, and how water levels vary across pavements over a growing season, would help inform land managers on how best to protect these systems, especially in light of climate change. The Australian Alps are predicted to become drier, with less precipitation and a longer snow-free season (Whetton et al. 1996; Hughes 2003; Williams and Wahren 2005; Pickering 2007). At present, it is unknown how these systems would respond to changes in hydrological regime shifts; however, groundwater-dependent communities have been highlighted as areas of particular concern when it comes to the predicted changes in climate (Pickering et al. 2004; Pickering 2007). Clements (2009) hypothesised that Sphagnum may currently be excluded from groundwater-source pools because of the chemical composition of groundwater, specifically the high levels of carbon dioxide found in groundwater, and that pavements play a role in regulating that chemical composition to a state where Sphagnum can thrive in peatlands below pavements. More research is needed to confirm this hypothesis; such an investigation may inform how the floristic composition of pavements could change in the future with less water availability.
While some pavements have been afforded intermittent protection from past feral ungulate impacts, historic cattle-excluding fences have now been removed and new threats such as feral deer and horses have complete access to pavements. Over the past 15 years, not only has feral horse evidence become significantly more prevalent in areas previously not accessed by horses, but the intensity of the impacts has also increased, particularly in wetland areas (Tolsma and Shannon 2018). Similarly, deer pose a significant threat to the flora and soils of wetland areas, including pavements (Z. Walker, unpubl. data).
Conclusions
We identified how gravelly pavement floristic composition varies across the landscape, the abiotic correlates that contribute to floristic variation within-and across-sites, and identified the patterns of species rarity in pavements. We conclude that gravelly pavement herbfields on the Bogong High Plains are floristically rare and geographically restricted groundwater-dependent ecosystems in need of greater recognition. Most importantly, we identified the relationship between groundwater spring and slope, as a mode of origin and persistence for this community, a relationship that has not previously been described in the literature. This community is highly susceptible to threats such as feral ungulate herbivory, trampling and changing hydrological regimes. In light of these threats, it is critically important that a management plan for the protection of this ecosystem is developed and implemented.
Supplementary material
Supplementary material is available online.
Data availability
Data will be available upon request to the first author (ABS).
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
This research did not receive any specific funding.
Acknowledgements
Warwick Papst, Zac Walker, Henrik Wahren, Dick Williams, Hayley Sime and Nina Kerr provided guidance, knowledge, help and support throughout various aspects of this study. We thank Falls Creek Resort Management for access to study sites within the resort, Parks Victoria Mount Beauty ranger staff (in particular, Elaine Thomas and Mick Keenan), the Research Centre for Applied Alpine Ecology (for research support) and Matt Renner at the Royal Botanic Gardens Sydney for his bryophyte identification services. Two anonymous reviewers helped improve the manuscript.
References
Atkin OK, Collier DE (1992) Relationship between soil nitrogen and floristic variation in late snow areas of the Kosciuszko alpine region [Australia]. Australian Journal of Botany 40, 139–149.| Relationship between soil nitrogen and floristic variation in late snow areas of the Kosciuszko alpine region [Australia].Crossref | GoogleScholarGoogle Scholar |
Billings WD, Mooney HA (1968) The ecology of arctic and alpine plants. Biological Reviews 43, 481–529.
| The ecology of arctic and alpine plants.Crossref | GoogleScholarGoogle Scholar |
Björk RG, Molau U (2007) Ecology of alpine snowbeds and the impact of global change. Arctic, Antarctic, and Alpine Research 39, 34–43.
| Ecology of alpine snowbeds and the impact of global change.Crossref | GoogleScholarGoogle Scholar |
Brown D, Thomas E, Herbert K, Primrose K (2016) Evaluating the effects of feral deer management on endangered alpine peatlands: the Alpine National Park deer control trial. Plant Protection Quarterly 31, 63–66.
| Evaluating the effects of feral deer management on endangered alpine peatlands: the Alpine National Park deer control trial.Crossref | GoogleScholarGoogle Scholar |
Bureau of Meteorology (2020) Falls Creek, Victoria. Monthly climate statistics. Available at http://www.bom.gov.au/climate/averages/tables/cw_083084_All.shtml [Verified 13 September 2021]
Clements AR (2009) Macroinvertebrate communities and the physio-chemical conditions in groundwater-fed bryophyte pools at Whiterock Creek, Bogong High Plains, Victoria, Australia. Honours Thesis, La Trobe University, Albury-Wodonga, NSW, Australia.
Clements AR, Suter PJ, Fussell M, Silvester E (2016) Macroinvertebrate communities in spring-fed alpine source pools. Hydrobiologia 777, 119–138.
| Macroinvertebrate communities in spring-fed alpine source pools.Crossref | GoogleScholarGoogle Scholar |
Costin AB (1972) Carbon-14 dates from the Snowy Mountains area, southeastern Australia, and their interpretation. Quaternary Research 2, 579–590.
| Carbon-14 dates from the Snowy Mountains area, southeastern Australia, and their interpretation.Crossref | GoogleScholarGoogle Scholar |
Costin AB, Gray M, Totterdell CJ (2000) ‘Kosciuszko alpine flora’, 2nd edn. (CSIRO Publishing: Melbourne, Vic., Australia)
DELWP (2021) Flora and Fauna Guarantee Act 1998 - Threatened List. State of Victoria Department of Environment, Land, Water and Planning, Melbourne, Victoria. Available at https://www.environment.vic.gov.au/conserving-threatened-species/threatened-list [Accessed 14 June 2022]
DEPI (2014) Advisory list of rare or threatened plants in Victoria. State of Victoria, Department of Primary Industries, Melbourne, Vic., Australia. Available at https://www.environment.vic.gov.au/__data/assets/pdf_file/0021/50448/Advisory-List-of-Rare-or-Threatened-Plants-in-Victoria-2014.pdf [Accessed 13 September 2021]
Geissler P (1982) Alpine communities. In ‘Bryophyte ecology’. (Ed. AJE Smith) pp. 167–189. (Springer: Dordrecht, Netherlands)
Hughes L (2003) Climate change and Australia: trends, projections and impacts. Austral Ecology 28, 423–443.
| Climate change and Australia: trends, projections and impacts.Crossref | GoogleScholarGoogle Scholar |
Kindt R, Coe R (2005) ‘Tree diversity analysis: a manual and software for commone statistical methods for ecology and biodiversity studies.’ (World Agroforestry Center: Nairobi, Kenya)
Lawrence RE (1995) The effects of grazing activity on the hydrology of the Bogong High Plains, Australia. The Rangeland Journal 17, 138–153.
| The effects of grazing activity on the hydrology of the Bogong High Plains, Australia.Crossref | GoogleScholarGoogle Scholar |
Mark AF, Bliss LC (1970) The high-alpine vegetation of Central Otago, New Zealand. New Zealand Journal of Botany 8, 381–451.
| The high-alpine vegetation of Central Otago, New Zealand.Crossref | GoogleScholarGoogle Scholar |
McCartney VA, Silvester E, Morgan JW, Suter PJ (2013) Physical and chemical drivers of vegetation in groundwater-source pools on the Bogong High Plains, Victoria. Australian Journal of Botany 61, 566–573.
| Physical and chemical drivers of vegetation in groundwater-source pools on the Bogong High Plains, Victoria.Crossref | GoogleScholarGoogle Scholar |
McDougall K (1982) ‘The alpine vegetation of the Bogong High Plains.’ Environmental Studies Publication no. 357. (Soil Conservation Authority, Environmental Studies Division, The State of Victoria Ministry for Conservation: Melbourne, Vic., Australia)
McDougall K, Walsh N (2007) Treeless vegetation of the Australian Alps. Cunninghamia 10, 1–57.
McVean DN (1969) Alpine vegetation of the central Snowy Mountains of New South Wales. Journal of Ecology 57, 67–86..
Meagher D, Downing A, Curnow J, Clarke J (2002) The bryophyte flora of Kosciuszko. In ‘Biodiversity in the snowy mountains’. (Ed. K Green) pp. 14–22 (Australian institute of Alpine Studies: Jindabyne, NSW, Australia)
Northcote KH (1979) ‘Factual key for the recognition of Australian soils.’ (Rellim Technical Publications: Adelaide, SA, Australia)
Oksanen AF, Blanchet G, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara B, Simpson GL, Solymos P, Henry M, Stevens H, Szoecs E, Wagner H (2019) Vegan: community ecology package. R package version 2.5-6. Available at https://CRAN.R-project.org/package=vegan [Verified 13 September 2021]
Pickering C (2007) Climate change and other threats in the Australian Alps. In ‘Protected areas: buffering nature against climate change. Proceedings of a WWF and IUCN world comission on protected areas symposium’. (Eds M Taylor, P Figgis) pp. 28–34. (WWF Australia: Sydney, NSW, Australia)
Pickering C, Good R, Green K (2004) Potential effects of global warming on the biota of the Australian Alps. Report for the Australian Greenhouse Office, Canberra, ACT, Australia.
Pickering C, Green K, Barros AA, Venn S (2014) A resurvey of late-lying snowpatches reveals changes in both species and functional composition across snowmelt zones. Alpine Botany 124, 93–103.
| A resurvey of late-lying snowpatches reveals changes in both species and functional composition across snowmelt zones.Crossref | GoogleScholarGoogle Scholar |
Rundel PW, Millar CI (2016) Alpine ecosystems. In ‘Ecosystems of California’. (Eds E Zavaleta, H Mooney) pp. 613–634. (University of California Press: Berkeley, CA, USA)
Shannon JM, Morgan JW (2007) Floristic variation in Sphagnum-dominated peatland communities of the Central Highlands, Victoria. Cunninghamia 10, 59–76.
Threatened Species Scientific Committee (2016) Conservation advice, Argyrotegium nitidilim, Shining cudweed. Threatened Species Scientific Committee: Canberra, ACT, Australia. Available at http://www.environment.gov.au/biodiversity/threatened/species/pubs/82043-conservation-advice-16122016.pdf [Verified 13 September 2021]
Tolsma AD, Shannon J (2018) Assessing the impacts of feral horses on the Bogong High Plains, Victoria. Unpublished client report for Parks Victoria. Department of Environment, Land, Water and Planning, Melbourne, Vic., Australia.
Tolsma AD, Wahren CH (2016) Mapping and monitoring Caltha introloba Herbland Community. Unpublished client report. Department of Environment, Land, Water and Planning, Melbourne, Vic., Australia.
Valachovič M, Dierssen K, Dimopoulos P, Hadač E, Loidi J, Mucina L, Rossi G, Tendero FV, Tomaselli M (1997) The vegetation on screes: a synopsis of higher syntaxa in Europe. Folia Geobotanica 32, 173–192.
| The vegetation on screes: a synopsis of higher syntaxa in Europe.Crossref | GoogleScholarGoogle Scholar |
Van Rees H (1984) ‘Behaviour and diet of free-ranging cattle on the Bogong High Plains Victoria.’ Environmental Studies Series no. 409. (Department of Conservation, Forests and Lands: Melbourne, Vic., Australia)
Venn S, Kirkpatrick J, McDougall K, Walsh N, Whinam J, Williams RJ (2017) ‘Alpine, sub-alpine and sub-Antarctic vegetation of Australia. In ‘Australian Vegetation’. 3rd edn. (Ed. DA Keith) pp. 461–489. (Cambridge University Press: Cambridge, UK)
Wahren CH (1997) Vegetation dynamics on the Bogong High Plains. PhD Thesis, Monash University, Melbourne, Vic., Australia.
Wahren C-H, Williams RJ, Papst WA (1999) Alpine and subalpine wetland vegetation on the Bogong High Plains, south-eastern Australia. Australian Journal of Botany 47, 165–188.
| Alpine and subalpine wetland vegetation on the Bogong High Plains, south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Wahren C-HA, Williams RJ, Papst WA (2001a) Vegetation change and ecological processes in alpine and subalpine Sphagnum bogs of the Bogong High Plains, Victoria, Australia. Arctic, Antarctic, and Alpine Research 33, 357–368.
| Vegetation change and ecological processes in alpine and subalpine Sphagnum bogs of the Bogong High Plains, Victoria, Australia.Crossref | GoogleScholarGoogle Scholar |
Wahren C-H, Williams RJ, Papst WA (2001b) Alpine and subalpine snow patch vegetation on the Bogong High Plains, SE Australia. Journal of Vegetation Science 12, 779–790.
| Alpine and subalpine snow patch vegetation on the Bogong High Plains, SE Australia.Crossref | GoogleScholarGoogle Scholar |
Wardlow IF, Moncur MW, Totterdell CJ (1989) The growth and development of Caltha introloba F.Muell. II. The regulation of germination, growth and photosynthesis by temperature. Australian Journal of Botany 37, 291–303.
| The growth and development of Caltha introloba F.Muell. II. The regulation of germination, growth and photosynthesis by temperature.Crossref | GoogleScholarGoogle Scholar |
Whetton PH, Haylock MR, Galloway R (1996) Climate change and snow-cover duration in the Australian Alps. Climatic Change 32, 447–479.
| Climate change and snow-cover duration in the Australian Alps.Crossref | GoogleScholarGoogle Scholar |
Wickham H (2016) ‘ggplot2: elegant graphics for data analysis.’ (Springer-Verlag: New York, NY, USA)
Williams RJ, Costin AB (1994) Alpine and subalpine vegetation. In ‘Australian vegetation’. (Ed. HR Groves) pp. 467–500. (Press Syndicate of the University of Cambridge: Melbourne, Vic., Australia)
Williams RJ, Wahren CH (2005) Potential impacts of global climate change on vegetation in Australian alpine landscapes: climate change, landuse, vegetation dynamics and biodiversity conservation. In ‘Global change and mountain regions: an overview of current knowledge’. (Eds UM Huber, HKM Bugmann, MA Reasoner) pp. 401–408. (Springer: Dordrecht, Netherlands)
Williams RJ, Papst W, McDougall K, Mansergh I, Heinz D, Camac J, Nash M, Morgan J, Hoffman A (2014) Alpine ecosystems. In ‘Biodiversity and environmental change: monitoring, challenges and direction’. (Eds D Lindenmayer, E Burns, N Thurgate, A Lowe) pp. 167–212. (CSIRO Publishing: Melbourne, Vic., Australia)
Wimbush DJ, Costin AB (1979) Trends in vegetation at Kosciusko. III. Alpine range transects, 1959-1978. Australian Journal of Botany 27, 833–871.
| Trends in vegetation at Kosciusko. III. Alpine range transects, 1959-1978.Crossref | GoogleScholarGoogle Scholar |