

Low-level CAM photosynthesis in a succulent-leaved member of the Urticaceae, Pilea peperomioides
Klaus Winter A C , Milton Garcia A , Aurelio Virgo A and J. Andrew C. Smith B
A C , Milton Garcia A , Aurelio Virgo A and J. Andrew C. Smith B
A Smithsonian Tropical Research Institute, PO Box 0843-03092, Balboa, Ancón, Republic of Panama.
B Department of Plant Sciences, University of Oxford, South Parks Road, Oxford, OX1 3RB, UK.
C Corresponding author. Email: winterk@si.edu
Functional Plant Biology 48(7) 683-690 https://doi.org/10.1071/FP20151
Submitted: 2 June 2020 Accepted: 28 October 2020 Published: 8 December 2020
Journal Compilation © CSIRO 2021 Open Access CC BY-NC-ND
Abstract
Pilea peperomioides Diels (Urticaceae) is a semi-succulent herbaceous species native to south-western China that has become popular in cultivation as an ornamental plant. To investigate whether this species possesses the capacity for CAM photosynthesis, measurements were made of CO2 gas exchange and titratable acidity in plants under both well-watered and water-deficit conditions. Plants were found to assimilate CO2 almost exclusively in the light via C3 photosynthesis. However, distinct transient reductions in the rate of net nocturnal CO2 release were consistently observed during the course of the dark period, and under water-deficit conditions one plant exhibited a brief period of net nocturnal CO2 uptake, providing unequivocal evidence of CAM activity. Furthermore, nocturnal increases in titratable acidity in both leaf laminas and petioles were observed in all plants exposed to wet–dry–wet cycles. This is the first report of CAM in the family Urticaceae. The results are discussed in relation to the phylogenetic position of Pilea and the partially shaded montane habitats in which this species is typically found. An updated list of all plant families currently known to contain species with CAM is presented.
Keywords: CAM photosynthesis, Chinese money plant, CO2 fixation, drought stress, nettle, photosynthesis, Pilea peperomioides, succulence, tissue acidity, Urticaceae.
Introduction
Pilea is the largest genus in the nettle family Urticaceae, with 604 accepted species native to tropical and temperate Asia, North, Central, and South America, and Africa. A phylogenetic analysis of Pilea was presented by Monro (2006). The majority of species are shade-tolerant annual or perennial herbs, often with fleshy and turgescent stems. Some have horticultural value, such as Pilea cadierei Gagnep. & Guillaumin (the aluminium plant), Pilea pubescens Liebm. (= P. involucrata Urb.; the friendship plant), Pilea nummulariifolia (Sw.) Wedd. (creeping Charlie), and Pilea microphylla (L.) Liebm. (the artillery plant, so named because of the puffs of pollen ejected by the exploding anthers).
Another increasingly popular ornamental species worldwide is Pilea peperomioides Diels, variously known as the Chinese money plant, the missionary plant, and mirror leaf. The plant is an evergreen perennial with distinctive semi-succulent, peltate leaf laminas, shiny and glabrous on the upper surface, borne on long petioles (Fig. 1). The species is native to Yunnan and Sichuan provinces in south-western China, where it grows lithophytically in montane forests on damp rocks such as humus-covered boulders and cliff-ledges. The plant first became known to the western world in the early 20th century through specimens collected in Yunnan province in 1906 and 1910 by the Scottish botanist and explorer George Forrest, which were formally named by Ludwig Diels in 1912 (Diels 1912). Following a more detailed description of the species and the first published image in 1984 (Radcliffe-Smith 1984), interest grew in explaining how this plant came to be so widely cultivated as an ornamental. It appears that living material collected in Kunming, Yunnan Province, was first introduced into Europe by the Norwegian missionary Agnar Espegren in 1946 (the material having survived transportation in a box for over a year), after which the plant was widely propagated and became popular as a house plant (Kers 1985). Despite its widespread cultivation (Dransfield and Dransfield 1984; Lancaster 1985; Walker and Levers 1993; Zou et al. 2017), this species remains little investigated.

|
Because of the succulent foliage of these plants, we decided to investigate whether the species exhibits features of CAM, since the CAM pathway has never previously been demonstrated in the family Urticaceae.
CAM photosynthesis is one of three photosynthetic pathways and is typically found in succulent plants that occupy semiarid terrestrial and epiphytic habitats in the subtropics and tropics. The CAM cycle is defined by nocturnal uptake of atmospheric CO2 via phosphoenolpyruvate carboxylase (PEPC) leading to the accumulation of malic acid in the vacuoles of chloroplast-containing cells. During the subsequent light period, malic acid is released from the vacuoles, decarboxylated, and the liberated CO2 re-fixed by Rubisco into the Calvin cycle (Osmond 1978; Winter and Smith 1996a; Holtum et al. 2005). Like C4 photosynthesis, CAM is a modification of the ancestral C3 pathway, serving to concentrate CO2 in the vicinity of Rubisco, thereby enhancing its CO2 fixation capacity. Whereas C4 plants are more water-use efficient than C3 plants, CAM allows for a further increase in water-use efficiency, because stomata are typically closed for most part of the day, and at night, when stomata open, the driving forces for water loss are strongly reduced owing to a low tissue–air vapour-pressure difference (Neales et al. 1968; Osmond et al. 1979; Winter et al. 2005). Although well-known archetypal CAM plants like cacti and agaves gain carbon mostly at night (Neales 1973a; Hanscom and Ting 1978; Nobel 1988), many CAM-exhibiting species still maintain a strong C3 component of CO2 assimilation during the day (Borland et al. 2011; Winter et al. 2015). In fact, in many species with low levels of CAM activity, the contribution of dark CO2 fixation to total carbon gain may be less than 5%, with CAM merely offsetting the loss of respiratory CO2 and, at the very low end of the CAM spectrum, not necessarily being detectable as net nocturnal CO2 uptake (Holtum and Winter 1999; Winter et al. 2019a). The ratio of dark to light CO2 fixation is species-dependent, and increases as photosynthetic organs mature. Furthermore, this ratio is strongly influenced by environmental conditions. For example, dark CO2 fixation is less sensitive to water-deficit stress than CO2 fixation in the light (Kluge and Fischer 1967; Hanscom and Ting 1978; Smith and Lüttge 1985; Winter 2019). Moreover, in plants with facultative CAM, dark CO2 fixation is induced or enhanced in response to reduced soil water availability or high salinity stress (Winter and Holtum 2011, 2014), with the halophyte Mesembryanthemum crystallinum L. being one of the most studied species (Winter and von Willert 1972; Winter and Lüttge 1976; Winter and Holtum 2007). Hence, unlike a fully developed C4 pathway, the presence and magnitude of CAM in a species is rarely an all-or-nothing phenomenon and the flexibility of many CAM-exhibiting species in terms of CO2 fixation in the dark and light provides some of the most spectacular examples of metabolic plasticity in the plant kingdom (Winter 2019).
In the research presented here, we examined 24-h changes of CO2 gas-exchange and acid content in Pilea peperomioides exposed to wet–dry–wet cycles. We provide evidence for the presence of the CAM cycle in this species, and thus report for the first time the occurrence of CAM in the family Urticaceae.
Materials and methods
Potted plants of Pilea peperomioides Diels were purchased in a local supermarket in Panama City, Republic of Panama. Plastic pots had an upper diameter of 15 cm and a height of 10 cm. Shoots were ~20 cm tall. For measurements of titratable acidity, plants were kept outdoors underneath a transparent polycarbonate-roofed rain shelter for 4 weeks at the Tupper Centre of the Smithsonian Tropical Research Institute (8.963284°, –79.543474°). Daily PFD was ~70% of full natural radiation. Titratable acidity was determined in plants exposed to a wet–dry–wet cycle. Leaf laminas and petioles were excised at dusk and dawn and their fresh mass determined. The area of leaf laminas was measured with a LI-3100 leaf area meter (LI-COR Biosciences). Samples were stored in liquid nitrogen and then freeze-dried for 3 days (FreeZone 4.5, Labconco). After determination of dry mass, samples were boiled in 50 mL of 50% ethanol (v/v) for 10 min. Evaporated solution was replaced with water and samples were boiled again for 10 min. After extracts attained room temperature, their volume was increased with water to 50 mL, and H+ content determined by titration with 5 mM NaOH to pH 6.5.
Several attempts were made to measure gas exchange of single attached leaves, either by enclosing leaf laminas into a PMK 10 clamp-on cuvette (Walz GmbH), or by inserting them via their petioles into a 11 cm × 11 cm × 10 cm (internal dimensions) Perspex cuvette. In five of six cases, no CO2 signal was measurable for several days, i.e. net CO2 exchange was ± 0. In only one case did a leaf show a photosynthetic-respiratory response. We therefore switched to CO2 gas-exchange measurements of whole shoots.
Plants for CO2 gas exchange were established from plantlets that grew out of the base of purchased mother plants. Plants were grown in 1.0 L terracotta pots filled with potting mix (Miracle-Gro Lawn Products). Shoots of entire plants were enclosed inside a Perspex cuvette (internal dimensions 11 cm × 11 cm × 15 cm) that rested on the terracotta pot in which the plants grew. The roots and the pot remained outside the cuvette. After initial daily irrigation with water, water deficit treatments were imposed by withholding irrigation until net CO2 uptake during the light period had decreased markedly, after which plants were re-watered daily. The gas-exchange cuvette, pot and plant were located inside a controlled environment chamber (GC8-T, EGC) operating under 12 h light (25°C):12 h dark (20°C) cycles. Illumination was supplied by an LED grow light (SS-GU300-w, Sunshine Systems). Photon flux density was 200 µmol m–2 s–1. The cuvette was supplied with air containing 400 µL L–1 CO2 by adding pure CO2 to CO2-free air using a GMA-3/10 gas mixing system (Walz). The air flow rate was 1.1 L min–1. Net CO2 exchange was measured in a flow-through gas-exchange system consisting of Walz components such as mass-flow controlled air pumps (one LD-10R, two LD-1R) and a gas-switch unit (GU-2), and an LI-820 or LI-7000 CO2 analyser (LI-COR Biosciences) operating in the absolute mode. A CR1000 data-logger (Campbell Scientific) controlled the gas-switch unit and recorded the CO2 concentration of air entering and leaving the plant cuvette at 2-min intervals.
Results
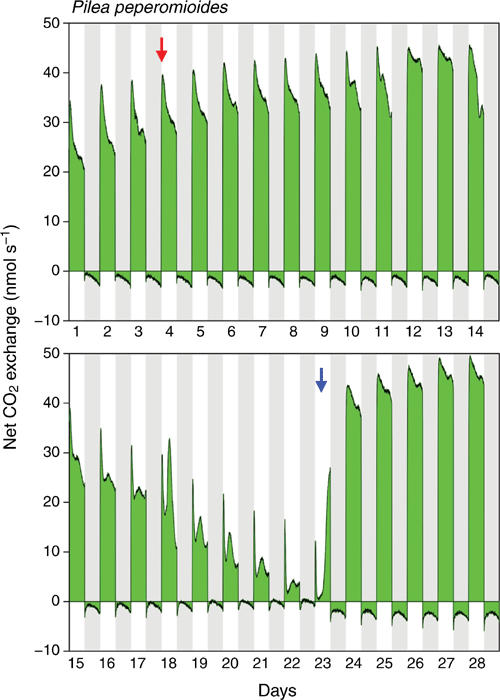
A total of 125 days of whole-shoot CO2 exchange were monitored for four separate plants. All experiments provided evidence of low-level CAM. Fig. 2 depicts the data for one of the plants. Net CO2 uptake was almost exclusively restricted to the light period, but unlike typical C3 species, especially when maintained under relatively low light-conditions as employed in the current study, neither the rates of daytime nor night-time CO2 exchange in Pilea peperomioides were constant during their respective parts of the light/dark cycle. This was particularly true for nocturnal CO2 exchange, which showed a strongly curved pattern throughout the experiment. Following an initial overshoot at the beginning of the night, net CO2 loss decreased and then increased. While in the first part of the experiment the lowest rates of net CO2 loss were observed during the very early part of the dark period (days 1–14), this point of minimal CO2 loss shifted towards the middle of the night and the curvature became more symmetrical as the experiment proceeded (days 15–28). Following cessation of watering on day 4, overall rates of daytime and night-time CO2 exchange continued to increase as the plant grew further. Daytime CO2 fixation began to decrease sharply on day 14, indicating the onset of drought stress. Simultaneously, overall rates of net CO2 loss decreased, and from day 19 onwards the nocturnal CO2 exchange trace shifted transiently to net uptake of CO2 during the middle of the dark period. Upon re-watering on day 23, daytime CO2 uptake and night-time CO2 loss rapidly recovered, while the curvature of nocturnal net CO2 loss was maintained.
|
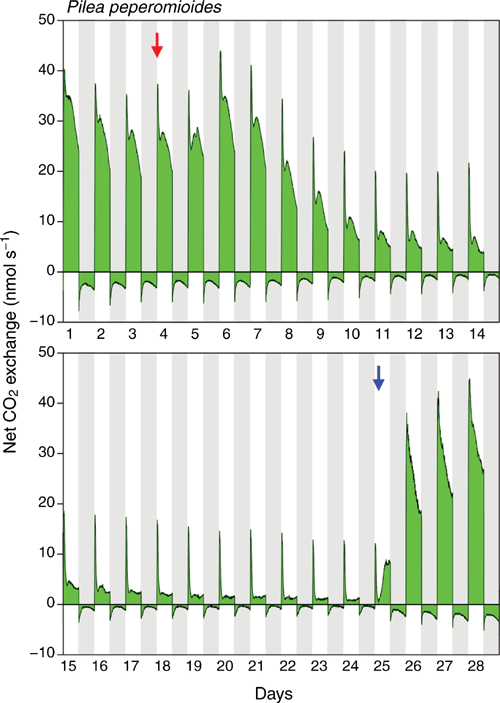
Gas-exchange responses of three other plants exposed to wet–dry–wet cycles were similar to the responses depicted in Fig. 2 in that nocturnal CO2 loss consistently showed a strongly curved pattern. This is illustrated in Fig. 3 for one of these three plants. However, during severe water deficit stress, rates of net nocturnal CO2 exchange merely approached zero at the point of minimal CO2 loss (Fig. 3, days 20–24), and net CO2 exchange never crossed the compensation line and became positive.
|
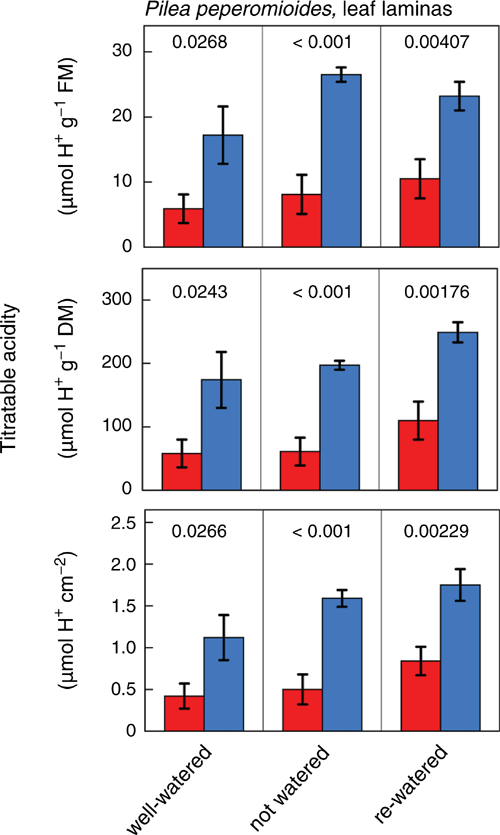
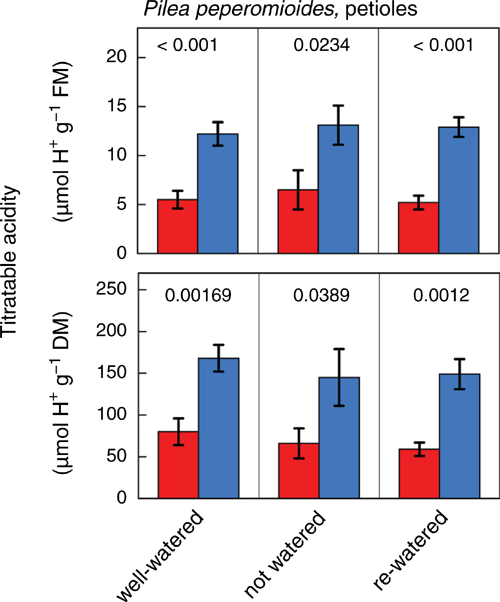
Leaf laminas (Fig. 4) and petioles (Fig. 5) of plants exposed to a wet–dry–wet cycle consistently showed significant nocturnal increases of titratable acidity content, irrespective of treatment or whether acidity was expressed on a fresh mass, dry mass or, in the case of leaf laminas, area basis. Compared with the well-watered initial state, water-deficit treatment led to a significant increase in leaf titratable acidity per unit fresh mass at dawn (P = 0.046), whereas dusk values did not differ significantly between these two states (P = 0.287). However, expressed on a dry mass or area basis, neither the dawn nor dusk values of titratable acidity differed significantly between water-deficit and well-watered conditions.
|
|
Water-deficit treatments as applied in Figs 2–4 did not lead to visible wilting of leaf laminas. Detached leaf laminas maintained for seven days on a laboratory bench under dim light (PFD of 20 µmol photons m–2 s–1 during daytime) at 23°C and 50% RH lost only 22 ± 3% of their original fresh mass (mean ± s.d., n = 5), and leaf water content per unit area declined by only 21% from 89 ± 8 mg H2O cm–2 to 70 ± 6 mg H2O cm–2.
Discussion
Although light-driven C3 photosynthesis is the principal pathway of carbon acquisition in P. peperomioides, the species is not 100% C3 as both CO2 gas exchange and titratable acidity measurements indicate the capacity for low-level CAM. The temporal pattern of nocturnal CO2 exchange shows a strong curvature, with the rate of net CO2 loss decreasing towards the middle of the dark period followed by a gradual increase thereafter (Fig. 2, predominantly under water-deficit and re-watered conditions; Fig. 3, throughout entire experiment). This pattern would be consistent with a transient increase in dark CO2 fixation that reduces net CO2 loss against a more or less constant background of dark respiration. In plants without CAM, under the constant night-temperature conditions used in this study, rates of respiratory net CO2 loss would typically remain steady and unchanged throughout the course of the dark period. Moreover, the finding that in one of the plants the nocturnal CO2 exchange trace transiently crossed the compensation line and shifted to net CO2 uptake under conditions of severe water-deficit stress (days 19–22 in Fig. 2) provides unequivocal evidence of CAM activity, and suggests that low-level CAM in P. peperomioides has a facultative component.
Titratable acidity measurements provide further evidence for the presence of CAM, as significant nocturnal increases in H+ were consistently observed in both leaf laminas (Fig. 4) and petioles (Fig. 5) irrespective of plant water status. The magnitude of acidification in leaf laminas and petioles was similar on a fresh mass and dry mass basis, respectively. Consistent with a facultative CAM component, average ΔH+, i.e. the difference between dawn and dusk values, appeared to be slightly enhanced on a fresh mass basis in leaf laminas of water-stressed compared with well-watered and re-watered plants, although this could not be confirmed statistically with the limited sample size available in this investigation.
The observation of CAM in petioles of P. peperomioides is in line with the increasing number of recent reports of CAM in small fleshy-stemmed herbs such as Basella (K. Winter, unpublished), Coleus (Winter et al. 2020b), Portulaca (Winter et al. 2019b), Sesuvium (Winter et al. 2019a), and Trianthema (Winter et al. 2020a). Although CAM is well known in perennial stem-succulents (Nobel 1988), the relatively short-lived leaf-bearing herbs with CAM-exhibiting fleshy stems and petioles may be larger contributors to CAM biodiversity than previously thought.
The adaptive significance of low-level CAM in P. peperomioides is presently unclear. Its full evaluation would require detailed studies of photosynthetic activity of plants and microclimate in situ. Although the majority of CAM plants occupy relatively open and periodically dry habitats, partially shaded montane sites where P. peperomioides has been collected in China (Radcliffe-Smith 1984) are typically not associated with CAM. However, the energetic cost of the CAM cycle is only moderately higher than that of C3 photosynthesis and does not represent an extra heavy burden on light-use efficiency (Winter and Smith 1996b; Cheung et al. 2014; Shameer et al. 2018). For example, the bromeliad Aechmea magdalenae is a well-known example of a plant with full CAM that can exist and flourish in the shaded understory of Neotropical forests (Skillman and Winter 1997; Skillman et al. 1999).
The Urticaceae belong to the large rosid clade of eudicots and is currently the only family in the order Rosales known to exhibit CAM. Other orders with CAM in the rosids are Vitales, Cucurbitales, Geraniales, Oxalidales and Malpighiales, the last of these containing three families with CAM: Clusiaceae, Euphorbiaceae, and Passifloraceae (Stevens 2001).
Table 1 presents an updated list of the families currently known to contain species with CAM. The list is a revised version of Table 1 in Winter and Smith (1996a) (see also Smith and Winter 1996) incorporating new evidence for presence and absence of CAM in taxa, and taking into account recent changes in plant relationships and taxonomic designations through mainly molecular phylogenetic studies. With addition of the Urticaceae, CAM has now been observed in a total of 37 families. Table 1 is a conservative estimate of the number of CAM families. For the time being, and in contrast to Winter and Smith (1996a), Apiaceae – a family with potentially aquatic CAM species – is no longer listed based on the assessment of Keeley (1998). As in Winter and Smith (1996a) and Smith and Winter (1996), we have not included Celastraceae, Ebenaceae, Flacourtiaceae (now Salicaceae) and Sapindaceae. Rao et al. (1979) reported substantial day-night changes in malate content in thin-leaved species of these families (one species of each). However, these plants all showed C3-type δ13C values and more work is needed to evaluate whether the CAM cycle genuinely operates in these species. Similarly, more information is required on the possible occurrence of CAM in leaves and stems of two non-succulent xerophytic tree species, Salvadora persica (Salvadoraceae) and Prosopis juliflora (Fabaceae) (Gaur 1968). In the study of Winter et al. (1976), leaves of Salvadora persica showed a C3-type δ13C value and there was no significant nocturnal accumulation of malate. We believe more work is also needed to confirm recent suggestions of fungal-induced CAM in Camellia oleifera (Theaceae) (Yuan et al. 2012) and of CAM in the coastal C4 grass Spinifex littoreus (Poaceae) (Ho et al. 2019). In both cases, continuous measurements of day/night CO2 exchange of the type shown in Fig. 2 are warranted to reveal the subtleties of weakly expressed CAM. Furthermore, in Camellia, the reported large difference between nocturnal increase of the malate anion and of H+ is stoichiometrically unusual and needs to be re-examined. Another potential ‘new’ CAM family not considered in Table 1 is the monotypic Halophytaceae, which contains the leaf-succulent Halophytum ameghinoi endemic to Patagonia. Its δ13C value of –18.6 ‰ (Holtum et al. 2018; E.J. Edwards, unpublished) is consistent with CAM, but gas-exchange and titratable acidity measurements have not yet been performed on this species. Once CAM is confirmed in these and other species, the total number of CAM families could well exceed 40.

|
Conflicts of interests
The authors declare no conflicts of interest.
Acknowledgements
This research was supported by the Smithsonian Tropical Research Institute. JACS acknowledges support from the Oxford Martin School.
References
APG IV (2016) An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: AGP IV. Botanical Journal of the Linnean Society 181, 1–20.| An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: AGP IV.Crossref | GoogleScholarGoogle Scholar |
Borland AM, Griffiths H (1997) A comparative study on the regulation of C3 and C4 carboxylation processes in the constitutive crassulacean acid metabolism (CAM) plant Kalanchoë daigremontiana and the C3-CAM intermediate Clusia minor. Planta 201, 368–378.
| A comparative study on the regulation of C3 and C4 carboxylation processes in the constitutive crassulacean acid metabolism (CAM) plant Kalanchoë daigremontiana and the C3-CAM intermediate Clusia minor.Crossref | GoogleScholarGoogle Scholar | 19343414PubMed |
Borland AM, Barrera Zambrano VA, Ceusters J, Shorrock K (2011) The photosynthetic plasticity of crassulacean acid metabolism: an evolutionary innovation for sustainable productivity in a changing world. New Phytologist 191, 619–633.
| The photosynthetic plasticity of crassulacean acid metabolism: an evolutionary innovation for sustainable productivity in a changing world.Crossref | GoogleScholarGoogle Scholar | 21679188PubMed |
Brilhaus D, Bräutigam A, Mettler-Altmann T, Winter K, Weber APM (2016) Reversible burst of transcriptional changes during induction of crassulacean acid metabolism in Talinum triangulare. Plant Physiology 170, 102–122.
| Reversible burst of transcriptional changes during induction of crassulacean acid metabolism in Talinum triangulare.Crossref | GoogleScholarGoogle Scholar | 26530316PubMed |
Cheung CYM, Poolman MG, Fell DA, Ratcliffe RG, Sweetlove LJ (2014) A diel flux balance model captures interactions between light and dark metabolism during day-night cycles in C3 and crassulacean acid metabolism leaves. Plant Physiology 165, 917–929.
| A diel flux balance model captures interactions between light and dark metabolism during day-night cycles in C3 and crassulacean acid metabolism leaves.Crossref | GoogleScholarGoogle Scholar |
Crayn DM, Winter K, Schulte K, Smith JAC (2015) Photosynthetic pathways in Bromeliaceae: phylogenetic and ecological significance of CAM and C3 based on carbon isotope ratios for 1893 species. Botanical Journal of the Linnean Society 178, 169–221.
| Photosynthetic pathways in Bromeliaceae: phylogenetic and ecological significance of CAM and C3 based on carbon isotope ratios for 1893 species.Crossref | GoogleScholarGoogle Scholar |
de Luca P, Alfani A, Virzo de Santo A (1977) CAM, transpiration, and adaptive mechanisms to xeric environments in the succulent Cucurbitaceae. Botanical Gazette 138, 474–478.
| CAM, transpiration, and adaptive mechanisms to xeric environments in the succulent Cucurbitaceae.Crossref | GoogleScholarGoogle Scholar |
Diels L (1912) Plantae Chinenses Forrestianae. New and imperfectly known species. Notes from the Royal Botanic Garden Edinburgh 5, 161–304.
Dransfield J, Dransfield S (1984) Pilea peperomioides in China. The Kew Magazine 1, 114
Gaur YD (1968) Preliminary studies on titratable acidity in xerophytic plants: Salvadora persica Linn. and Prosopis juliflora D.C. Experientia 24, 239–240.
| Preliminary studies on titratable acidity in xerophytic plants: Salvadora persica Linn. and Prosopis juliflora D.C.Crossref | GoogleScholarGoogle Scholar | 5661414PubMed |
Guralnick LJ, Jackson MD (2001) The occurrence and phylogenetics of crassulacean acid metabolism in the Portulacaceae. International Journal of Plant Sciences 162, 257–262.
| The occurrence and phylogenetics of crassulacean acid metabolism in the Portulacaceae.Crossref | GoogleScholarGoogle Scholar |
Guralnick LJ, Ting IP, Lord EM (1986) Crassulacean acid metabolism in the Gesneriaceae. American Journal of Botany 73, 336–345.
| Crassulacean acid metabolism in the Gesneriaceae.Crossref | GoogleScholarGoogle Scholar |
Hanscom Z, Ting IP (1978) Irrigation magnifies CAM-photosynthesis in Opuntia basilaris (Cactaceae). Oecologia 33, 1–15.
| Irrigation magnifies CAM-photosynthesis in Opuntia basilaris (Cactaceae).Crossref | GoogleScholarGoogle Scholar | 28309263PubMed |
Herrera A, Delgado J, Paraguatey I (1991) Occurrence of inducible crassulacean acid metabolism in leaves of Talinum triangulare (Portulacaceae). Journal of Experimental Botany 42, 493–499.
| Occurrence of inducible crassulacean acid metabolism in leaves of Talinum triangulare (Portulacaceae).Crossref | GoogleScholarGoogle Scholar |
Heyduk K, McKain MR, Lalani F, Leebens-Mack J (2016) Evolution of a CAM anatomy predates the origins of crassulacean acid metabolism in the Agavoideae (Asparagaceae). Molecular Phylogenetics and Evolution 105, 102–113.
| Evolution of a CAM anatomy predates the origins of crassulacean acid metabolism in the Agavoideae (Asparagaceae).Crossref | GoogleScholarGoogle Scholar | 27591171PubMed |
Ho C-L, Chiang J-M, Lin T-C, Martin CE (2019) First report of C4/CAM-cycling photosynthetic pathway in a succulent grass, Spinifex littoreus (Brum. F.) Merr., in coastal regions of Taiwan. Flora 254, 194–202.
| First report of C4/CAM-cycling photosynthetic pathway in a succulent grass, Spinifex littoreus (Brum. F.) Merr., in coastal regions of Taiwan.Crossref | GoogleScholarGoogle Scholar |
Holthe PA, Patel A, Ting IP (1992) The occurrence of CAM in Peperomia. Selbyana 13, 77–87.
Holtum JAM, Winter K (1999) Degrees of crassulacean acid metabolism in tropical epiphytic and lithophytic ferns. Australian Journal of Plant Physiology 26, 749–757.
Holtum JAM, Smith JAC, Neuhaus HE (2005) Intracellular transport and pathways of carbon flow in plants with crassulacean acid metabolism. Functional Plant Biology 32, 429–449.
| Intracellular transport and pathways of carbon flow in plants with crassulacean acid metabolism.Crossref | GoogleScholarGoogle Scholar | 32689145PubMed |
Holtum JAM, Winter K, Weeks MA, Sexton TR (2007) Crassulacean acid metabolism in the ZZ plant, Zamioculcas zamiifolia (Araceae). American Journal of Botany 94, 1670–1676.
| Crassulacean acid metabolism in the ZZ plant, Zamioculcas zamiifolia (Araceae).Crossref | GoogleScholarGoogle Scholar |
Holtum JAM, Hancock LP, Edwards EJ, Winter K (2018) Crassulacean acid metabolism in the Basellaceae (Caryophyllales). Plant Biology 20, 409–414.
| Crassulacean acid metabolism in the Basellaceae (Caryophyllales).Crossref | GoogleScholarGoogle Scholar | 29369469PubMed |
Horn JW, Xi Z, Riina R, Peirson JA, Yang Y, Dorsey BL, Berry PE, Davis CC, Wurdack KJ (2014) Evolutionary bursts in Euphorbia (Euphorbiaceae) are linked with photosynthetic pathway. Evolution 68, 3485–3504.
| Evolutionary bursts in Euphorbia (Euphorbiaceae) are linked with photosynthetic pathway.Crossref | GoogleScholarGoogle Scholar | 25302554PubMed |
Huang W, Han S, Xing Z, Li W (2020) Responses of leaf anatomy and CO2 concentrating mechanisms of the aquatic plant Ottelia cordata to variable CO2. Frontiers in Plant Science 11, 1261
| Responses of leaf anatomy and CO2 concentrating mechanisms of the aquatic plant Ottelia cordata to variable CO2.Crossref | GoogleScholarGoogle Scholar | 32922428PubMed |
Jones CS, Cardon ZG, Czaja AD (2003) A phylogenetic view of low-level CAM in Pelargonium (Geraniaceae). American Journal of Botany 90, 135–142.
| A phylogenetic view of low-level CAM in Pelargonium (Geraniaceae).Crossref | GoogleScholarGoogle Scholar | 21659089PubMed |
Keeley JE (1998) CAM photosynthesis in submerged aquatic plants. Botanical Review 64, 121–175.
| CAM photosynthesis in submerged aquatic plants.Crossref | GoogleScholarGoogle Scholar |
Kers LE (1985) Pilea peperomioides – a mystery solved. The Kew Magazine 2, 334–336.
Kluge M, Fischer K (1967) Über Zusammenhänge zwischen dem CO2-Austausch und der Abgabe von Wasserdampf durch Bryophyllum daigremontianum Berg. Planta 77, 212–223.
| Über Zusammenhänge zwischen dem CO2-Austausch und der Abgabe von Wasserdampf durch Bryophyllum daigremontianum Berg.Crossref | GoogleScholarGoogle Scholar | 24522539PubMed |
Kluge M, Ting IP (1978) ‘Crassulacean acid metabolism.’ (Springer: Berlin, Germany)
Kluge M, Knapp I, Kramer D, Schwerdtner I, Ritter H (1979) Crassulacean acid metabolism (CAM) in leaves of Aloe arborescens Mill. Planta 145, 357–363.
| Crassulacean acid metabolism (CAM) in leaves of Aloe arborescens Mill.Crossref | GoogleScholarGoogle Scholar | 24317763PubMed |
Koch K, Kennedy RA (1980) Characteristics of crassulacean acid metabolism in the succulent C4 dicot, Portulaca oleracea L. Plant Physiology 65, 193–197.
| Characteristics of crassulacean acid metabolism in the succulent C4 dicot, Portulaca oleracea L.Crossref | GoogleScholarGoogle Scholar | 16661159PubMed |
Lancaster R (1985) Pilea peperomioides in China. The Kew Magazine 2, 334
Lüttge U (2006) Photosynthetic flexibility and ecophysiological plasticity: Questions and lessons from Clusia, the only CAM tree, in the neotropics. New Phytologist 171, 7–25.
| Photosynthetic flexibility and ecophysiological plasticity: Questions and lessons from Clusia, the only CAM tree, in the neotropics.Crossref | GoogleScholarGoogle Scholar | 16771979PubMed |
Martin CE, Gravatt DA, Loeschen VS (1994) Crassulacean acid metabolism in three species of Commelinaceae. Annals of Botany 74, 457–463.
| Crassulacean acid metabolism in three species of Commelinaceae.Crossref | GoogleScholarGoogle Scholar |
Martin SL, Davis R, Protti P, Lin T-C, Lin S-H, Martin CE (2005) The occurrence of crassulacean acid metabolism in epiphytic ferns, with an emphasis on the Vittariaceae. International Journal of Plant Sciences 166, 623–630.
| The occurrence of crassulacean acid metabolism in epiphytic ferns, with an emphasis on the Vittariaceae.Crossref | GoogleScholarGoogle Scholar |
Medina E (1974) Dark CO2 fixation, habitat preference and evolution within the Bromeliaceae. Evolution 28, 677–686.
Monro AK (2006) The revision of species-rich genera: a phylogenetic framework for the strategic revision of Pilea (Urticaceae) based on cpDNA, nrDNA, and morphology. American Journal of Botany 93, 426–441.
| The revision of species-rich genera: a phylogenetic framework for the strategic revision of Pilea (Urticaceae) based on cpDNA, nrDNA, and morphology.Crossref | GoogleScholarGoogle Scholar | 21646202PubMed |
Mooney HA, Troughton JH, Berry JS (1977) Carbon isotope ratio measurements of succulent plants in southern Africa. Oecologia 30, 295–305.
| Carbon isotope ratio measurements of succulent plants in southern Africa.Crossref | GoogleScholarGoogle Scholar | 28309179PubMed |
Neales TF (1973a) The effect of night-temperature on CO2 assimilation, transpiration, and water use efficiency in Agave americana L. Australian Journal of Biological Sciences 26, 705–714.
| The effect of night-temperature on CO2 assimilation, transpiration, and water use efficiency in Agave americana L.Crossref | GoogleScholarGoogle Scholar |
Neales TF (1973b) Effect of night temperature on the assimilation of carbon dioxide by mature pineapple plants, Ananas comosus (L.) Merr. Australian Journal of Biological Sciences 26, 539–546.
| Effect of night temperature on the assimilation of carbon dioxide by mature pineapple plants, Ananas comosus (L.) Merr.Crossref | GoogleScholarGoogle Scholar |
Neales TF, Patterson AA, Hartney VJ (1968) Physiological adaptation to drought in the carbon assimilation and water loss of xerophytes. Nature 219, 469–472.
| Physiological adaptation to drought in the carbon assimilation and water loss of xerophytes.Crossref | GoogleScholarGoogle Scholar |
Nobel PS (1988) ‘Environmental biology of agaves and cacti.’ (Cambridge University Press: Cambridge, UK)
Nuernbergk EL (1961) Endogener Rhythmus und CO2-Stoffwechsel bei Pflanzen mit diurnalem Säurerhythmus. Planta 56, 28–70.
| Endogener Rhythmus und CO2-Stoffwechsel bei Pflanzen mit diurnalem Säurerhythmus.Crossref | GoogleScholarGoogle Scholar |
Osmond CB (1978) Crassulacean acid metabolism: a curiosity in context. Annual Review of Plant Physiology 29, 379–414.
| Crassulacean acid metabolism: a curiosity in context.Crossref | GoogleScholarGoogle Scholar |
Osmond CB, Ludlow MM, Davis R, Cowan IR, Powles SP, Winter K (1979) Stomatal responses to humidity in Opuntia inermis in relation to control of CO2 and H2O exchange patterns. Oecologia 41, 65–76.
| Stomatal responses to humidity in Opuntia inermis in relation to control of CO2 and H2O exchange patterns.Crossref | GoogleScholarGoogle Scholar | 28310360PubMed |
Radcliffe-Smith A (1984) 5. Pilea peperomioides: Urticaceae. The Kew Magazine 1, 14–19.
Rao IM, Swamy PM, Das VSR (1979) Some characteristics of crassulacean acid metabolism in five nonsucculent scrub species under natural semiarid conditions. Zeitschrift für Pflanzenphysiologie 94, 201–210.
| Some characteristics of crassulacean acid metabolism in five nonsucculent scrub species under natural semiarid conditions.Crossref | GoogleScholarGoogle Scholar |
Ruess BR, Eller BM (1985) The correlation between crassulacean acid metabolism and water uptake in Senecio medley-woodii. Planta 166, 57–66.
| The correlation between crassulacean acid metabolism and water uptake in Senecio medley-woodii.Crossref | GoogleScholarGoogle Scholar | 24241312PubMed |
Schütte KH, Steyn R, van der Westhuizen M (1967) Crassulacean acid metabolism in South African succulents: a preliminary investigation into its occurrence in various families. Journal of South African Botany 33, 107–110.
Shameer S, Baghalian K, Cheung CTM, Ratcliffe RG, Sweetlove LJ (2018) Computational analysis of the productivity potential of CAM. Nature Plants 4, 165–171.
| Computational analysis of the productivity potential of CAM.Crossref | GoogleScholarGoogle Scholar | 29483685PubMed |
Shao H, Gontero B, Maberly SC, Jiang HS, Cao Y, Li W, Huang WM (2017) Responses of Ottelia alismoides, an aquatic plant with three CCMs, to variable CO2 and light. Journal of Experimental Botany 68, 3985–3995.
| Responses of Ottelia alismoides, an aquatic plant with three CCMs, to variable CO2 and light.Crossref | GoogleScholarGoogle Scholar | 28369629PubMed |
Silvera K, Santiago L, Winter K (2005) Distribution of crassulacean acid metabolism in orchids of Panama: evidence of selection for weak and strong modes. Functional Plant Biology 32, 397–407.
| Distribution of crassulacean acid metabolism in orchids of Panama: evidence of selection for weak and strong modes.Crossref | GoogleScholarGoogle Scholar | 32689142PubMed |
Sipes DL, Ting IP (1985) Crassulacean acid metabolism and crassulacean acid metabolism modifications in Peperomia camptotricha. Plant Physiology 77, 59–63.
| Crassulacean acid metabolism and crassulacean acid metabolism modifications in Peperomia camptotricha.Crossref | GoogleScholarGoogle Scholar | 16664028PubMed |
Skillman JB, Winter K (1997) High photosynthetic capacity in a shade-tolerant crassulacean acid metabolism plant. Implications for sunfleck use, nonphotochemical energy dissipation, and susceptibility to photoinhibition. Plant Physiology 113, 441–450.
| High photosynthetic capacity in a shade-tolerant crassulacean acid metabolism plant. Implications for sunfleck use, nonphotochemical energy dissipation, and susceptibility to photoinhibition.Crossref | GoogleScholarGoogle Scholar | 12223618PubMed |
Skillman JB, Garcia M, Winter K (1999) Whole-plant consequences of crassulacean acid metabolism for a tropical forest understory plant. Ecology 80, 1584–1593.
| Whole-plant consequences of crassulacean acid metabolism for a tropical forest understory plant.Crossref | GoogleScholarGoogle Scholar |
Smith BN, Epstein S (1971) Two categories of 13C/12C ratios for higher plants. Plant Physiology 47, 380–384.
| Two categories of 13C/12C ratios for higher plants.Crossref | GoogleScholarGoogle Scholar | 16657626PubMed |
Smith JAC, Lüttge U (1985) Day–night changes in leaf water relations associated with the rhythm of crassulacean acid metabolism in Kalanchoë daigremontiana. Planta 163, 272–282.
| Day–night changes in leaf water relations associated with the rhythm of crassulacean acid metabolism in Kalanchoë daigremontiana.Crossref | GoogleScholarGoogle Scholar |
Smith JAC, Winter K (1996) Taxonomic distribution of crassulacean acid metabolism. In ‘Crassulacean acid metabolism. Biochemistry, ecophysiology and evolution’. (Eds K Winter, JAC Smith) pp. 427–436. (Springer: Berlin, Germany)
Stevens PF (2001) Angiosperm phylogeny website. Version 14, July 2017. http://www.mobot.org/MOBOT/research/APweb/ [Verified 4 November 2020]
Tinoco Ojanguren C, Vásquez-Yánes C (1983) Especies CAM en la selva húmeda tropical de los Tuxtlas, Veracruz. Boletín de la Sociedad Botánica de México 45, 150–153.
Tsen EWJ, Holtum JAM (2012) Crassulacean acid metabolism (CAM) in an epiphytic ant-plant, Myrmecodia beccarii Hook.f. (Rubiaceae). Photosynthesis Research 113, 311–320.
| Crassulacean acid metabolism (CAM) in an epiphytic ant-plant, Myrmecodia beccarii Hook.f. (Rubiaceae).Crossref | GoogleScholarGoogle Scholar |
Virzo de Santo A, Alfani A, Greco L, Fioretto A (1980) Environmental influences on CAM activity of Cissus quadrangularis. Journal of Experimental Botany 31, 75–82.
| Environmental influences on CAM activity of Cissus quadrangularis.Crossref | GoogleScholarGoogle Scholar |
von Willert DJ, Armbrüster N, Drees T, Zaborowski M (2005) Welwitschia mirabilis: CAM or not CAM – what is the answer? Functional Plant Biology 32, 389–395.
| Welwitschia mirabilis: CAM or not CAM – what is the answer?Crossref | GoogleScholarGoogle Scholar | 32689141PubMed |
Vovides AP, Etherington JR, Dresser PQ, Groenhof A, Iglesias C, Flores Ramirez J (2002) CAM-cycling in the cycad Dioon edule Lindl. in its natural tropical deciduous forest habitat in central Veracruz, Mexico. Botanical Journal of the Linnean Society 138, 155–162.
| CAM-cycling in the cycad Dioon edule Lindl. in its natural tropical deciduous forest habitat in central Veracruz, Mexico.Crossref | GoogleScholarGoogle Scholar |
Walker CC, Levers M (1993) Pilea peperomioides – a Chinese member of the stinging nettle family. British Cactus & Succulent Journal 11, 98–99.
Winter K (1979) δ13C values of some succulent plants from Madagascar. Oecologia 40, 103–112.
| δ13C values of some succulent plants from Madagascar.Crossref | GoogleScholarGoogle Scholar | 28309607PubMed |
Winter K (2019) Ecophysiology of constitutive and facultative CAM photosynthesis. Journal of Experimental Botany 70, 6495–6508.
| Ecophysiology of constitutive and facultative CAM photosynthesis.Crossref | GoogleScholarGoogle Scholar | 30810162PubMed |
Winter K, Holtum JAM (2007) Environment or development? Lifetime net CO2 exchange and control of the expression of crassulacean acid metabolism in Mesembryanthemum crystallinum. Plant Physiology 143, 98–107.
| Environment or development? Lifetime net CO2 exchange and control of the expression of crassulacean acid metabolism in Mesembryanthemum crystallinum.Crossref | GoogleScholarGoogle Scholar | 17056756PubMed |
Winter K, Holtum JAM (2011) Induction and reversal of crassulacean acid metabolism in Calandrinia polyandra: effects of soil moisture and nutrients. Functional Plant Biology 38, 576–582.
| Induction and reversal of crassulacean acid metabolism in Calandrinia polyandra: effects of soil moisture and nutrients.Crossref | GoogleScholarGoogle Scholar | 32480910PubMed |
Winter K, Holtum JAM (2014) Facultative crassulacean acid metabolism (CAM) plants: powerful tools for unravelling the functional elements of CAM photosynthesis. Journal of Experimental Botany 65, 3425–3441.
| Facultative crassulacean acid metabolism (CAM) plants: powerful tools for unravelling the functional elements of CAM photosynthesis.Crossref | GoogleScholarGoogle Scholar | 24642847PubMed |
Winter K, Holtum JAM (2017) Facultative crassulacean acid metabolism (CAM) in four small C3 and C4 leaf-succulents. Australian Journal of Botany 65, 103–108.
| Facultative crassulacean acid metabolism (CAM) in four small C3 and C4 leaf-succulents.Crossref | GoogleScholarGoogle Scholar |
Winter K, Lüttge U (1976) Balance between C3 and CAM pathway of photosynthesis. In ‘Water and plant life – problems and modern approaches’. (Eds OL Lange, L Kappen, ED Schulze) pp. 323–334. (Springer: Berlin, Germany)
Winter K, Smith JAC (1996a) An introduction to crassulacean acid metabolism. Biochemical principles and ecological diversity. In ‘Crassulacean acid metabolism. Biochemistry, ecophysiology and evolution’. (Eds K Winter, JAC Smith) pp. 1–13. (Springer: Berlin, Germany)
Winter K, Smith JAC (1996b) Crassulacean acid metabolism. Current status and perspectives. In ‘Crassulacean acid metabolism. Biochemistry, ecophysiology and evolution’. (Eds K Winter, JAC Smith) pp. 389–426. (Springer: Berlin, Germany)
Winter K, von Willert DJ (1972) NaCl-induzierter Crassulaceensäurestoffwechsel bei Mesembryanthemum crystallinum. Zeitschrift für Pflanzenphysiologie 67, 166–170.
| NaCl-induzierter Crassulaceensäurestoffwechsel bei Mesembryanthemum crystallinum.Crossref | GoogleScholarGoogle Scholar |
Winter K, Troughton JH, Evenari M, Läuchli A, Lüttge U (1976) Mineral ion composition and occurrence of CAM like diurnal malate fluctuations in plants of coastal and desert habitats of Israel and the Sinai. Oecologia 25, 125–143.
| Mineral ion composition and occurrence of CAM like diurnal malate fluctuations in plants of coastal and desert habitats of Israel and the Sinai.Crossref | GoogleScholarGoogle Scholar | 28308995PubMed |
Winter K, Aranda J, Holtum JAM (2005) Carbon isotope composition and water-use efficiency in plants with crassulacean acid metabolism. Functional Plant Biology 32, 381–388.
| Carbon isotope composition and water-use efficiency in plants with crassulacean acid metabolism.Crossref | GoogleScholarGoogle Scholar | 32689140PubMed |
Winter K, Holtum JAM, Smith JAC (2015) Crassulacean acid metabolism: a continuous or discrete trait? New Phytologist 208, 73–78.
| Crassulacean acid metabolism: a continuous or discrete trait?Crossref | GoogleScholarGoogle Scholar | 25975197PubMed |
Winter K, Wallace BJ, Stocker GC, Roksandic Z (1983) Crassulacean acid metabolism in Australian vascular epiphytes and some related species. Oecologia 57, 129–141.
| Crassulacean acid metabolism in Australian vascular epiphytes and some related species.Crossref | GoogleScholarGoogle Scholar | 28310165PubMed |
Winter K, Garcia M, Virgo A, Holtum JAM (2019a) Operating at the very low end of the crassulacean acid metabolism spectrum: Sesuvium portulacastrum (Aizoaceae). Journal of Experimental Botany 70, 6561–6570.
| Operating at the very low end of the crassulacean acid metabolism spectrum: Sesuvium portulacastrum (Aizoaceae).Crossref | GoogleScholarGoogle Scholar | 30535159PubMed |
Winter K, Sage RF, Edwards EJ, Virgo A, Holtum JAM (2019b) Facultative crassulacean acid metabolism in a C3 – C4 intermediate. Journal of Experimental Botany 70, 6571–6579.
| Facultative crassulacean acid metabolism in a C3 – C4 intermediate.Crossref | GoogleScholarGoogle Scholar | 30820551PubMed |
Winter K, Garcia M, Virgo A, Ceballos J, Holtum JAM (2020a) Does the C4 plant Trianthema portulacastrum L. (Aizoaceae) exhibit weakly expressed crassulacean acid metabolism (CAM)? Functional Plant Biology
Winter K, Virgo A, Garcia M, Aranda J, Holtum JAM (2020b) Constitutive and facultative crassulacean acid metabolism (CAM) in Cuban oregano, Coleus amboinicus (Lamiaceae). Functional Plant Biology
| Constitutive and facultative crassulacean acid metabolism (CAM) in Cuban oregano, Coleus amboinicus (Lamiaceae).Crossref | GoogleScholarGoogle Scholar | 33213694PubMed |
Wong SC, Hew CS (1976) Diffusive resistance, titratable acidity, and CO2 fixation in two tropical epiphytic ferns. American Fern Journal 66, 121–124.
| Diffusive resistance, titratable acidity, and CO2 fixation in two tropical epiphytic ferns.Crossref | GoogleScholarGoogle Scholar |
Yuan M, Xu F, Wang S-D, Zhang D-W, Zhang Z-W, Cao Y, Xu X-C, Luo M-H, Yuan S (2012) A single leaf of Camellia oleifera has two types of carbon assimilation pathway, C3 and crassulacean acid metabolism. Tree Physiology 32, 188–199.
| A single leaf of Camellia oleifera has two types of carbon assimilation pathway, C3 and crassulacean acid metabolism.Crossref | GoogleScholarGoogle Scholar | 22337600PubMed |
Zhang Y, Yin L, Jiang H-S, Wi L, Gontero B, Maberly SC (2014) Biochemical and biophysical CO2 concentrating mechanisms in two species of freshwater macrophyte within the genus Ottelia (Hydrocharitaceae). Photosynthesis Research 121, 285–297.
| Biochemical and biophysical CO2 concentrating mechanisms in two species of freshwater macrophyte within the genus Ottelia (Hydrocharitaceae).Crossref | GoogleScholarGoogle Scholar | 24203583PubMed |
Zou S, Zeng Y, Duan W, Li R (2017) Discussion about plant landscape configuration in the construction of urban rainwater gardens. Advances in Intelligent Systems Research 150, 77–83.