

Response of turtle hatchlings to light emitting diodes at sea
Phillipa Wilson A B * , Kellie Pendoley C , Scott Whiting D , Charitha Pattiaratchi A , Mark Meekan B and Michele Thums B
A B * , Kellie Pendoley C , Scott Whiting D , Charitha Pattiaratchi A , Mark Meekan B and Michele Thums B
A Oceans Graduate School and the UWA Oceans Institute, The University of Western Australia, 35 Stirling Highway, Perth, WA 6009, Australia.
B Australian Institute of Marine Science, Indian Ocean Marine Research Centre, The University of Western Australia (M096), 35 Stirling Highway, Perth, WA 6009, Australia.
C Pendoley Environmental, 12A Pitt Way, Booragoon, WA 6154, Australia.
D Marine Science Program, Department of Biodiversity, Conservation and Attractions, 17 Dick Perry Avenue, Kensington, WA 6151, Australia.
Marine and Freshwater Research 73(5) 689-702 https://doi.org/10.1071/MF21217
Submitted: 26 July 2021 Accepted: 27 December 2021 Published: 25 February 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
It is well known that light pollution disrupts the early dispersal of marine turtles. But now, light emitting diodes (LEDs) are replacing traditional lights, however, we know little about how they influence hatchling dispersal. Here, we used acoustic telemetry to assess the early in-water dispersal and predation rates of hatchlings in response to different intensities of LEDs ranging from 10 to 120 W. We found no effect of LEDs on hatchling bearing when lights were in the direction they dispersed under ambient conditions. When LEDs were not in their usual direction of travel, variability in mean bearing increased, and a change in bearing occurred with the highest light intensity. We found weak evidence that predation was also higher at this light intensity compared to ambient, and also in two of the lower light intensities (10 and 30 W), but only on one experimental night. We were unable to find a relationship between hatchling speed and time spent in the tracking area with light intensity. However, reduced sample sizes (due to predation) might have affected our ability to detect effects. Although more effort is required to increase the confidence in our findings, LEDs disrupted hatchling dispersal and are therefore likely to negatively affect their survival.
Keywords: acoustic telemetry, ALAN, artificial light, coastal development, flatback turtle, in-water movement, light emitting diodes (LEDs), light intensity, Natator depressus, turtle hatchlings.
Introduction
A naturally dark night is important for a wide range of functions of diurnal, nocturnal and crepuscular animals, including navigation, foraging and regulation of circadian cycles (Gaston et al. 2012). These natural levels of ambient light are now being affected by light pollution from artificial sources (Cinzano et al. 2001), which disrupts many of these behaviours, often with consequences for fitness and survival (Longcore and Rich 2004; Minnaar et al. 2015; Raap et al. 2015; Rodríguez et al. 2017; Russart and Nelson 2018; Zapata et al. 2019). This problem has become of particular concern in coastal ecosystems due to the widespread urbanisation and industrialisation of the coastal zone (Small and Nicholls 2003). Marine turtles are one such species where the impacts of light pollution have been well documented, especially the impact of light on hatchling sea-finding, i.e. their ability to locate the sea after they emerge from nests on beaches (Hooker 1911; McFarlane 1963; Verheijen 1985; Salmon 2003; Witherington and Martin 2003; Kamrowski et al. 2014). Hatchlings can be misoriented (i.e. move towards lights on land instead of towards the sea) and disoriented (i.e. move in confused circles) by sources of artificial light (Salmon 2005), which can have lethal consequences (e.g. increased risk of predation, exhaustion and dehydration) (Philibosian 1976; Erb and Wyneken 2019). Once they reach the sea, light pollution on and near the shore (e.g. jetties, boats on anchor, coastal lighting) can also confuse and misdirect them, increasing the time taken to pass through nearshore waters (Harewood and Horrocks 2008; Thums et al. 2016), and potentially cause a ‘light trapping effect’ where they concentrate around the source and are unable to respond to other stimuli to move on (Verheijen 1958; Wilson et al. 2018). This could have survival consequences as rates of predation in nearshore waters are already high (up to 88%; Reising et al. 2015) and lights can also attract predatory fish (Becker et al. 2013).
Field and laboratory studies on sea turtles have found the degree of impact that artificial light has on their dispersal behaviour is dependent on the spectral composition of the light emissions with hatchlings more attracted to short (blue–green) over long (orange–red) light wavelengths (Osovsky and Shettleworth 1968; Witherington and Bjorndal 1991a, 1991b; Witherington 1992; Pendoley 2005; Sella et al. 2006; Fritsches 2012). These studies led to the recommendation that lights containing the least amount of short wavelength light (e.g. orange low and high pressure sodium) should be used near turtle nesting beaches if lighting cannot be avoided (Witherington and Martin 2003). Filters can also be used to eliminate any shorter light wavelengths emitted from these sources (Sella et al. 2006). Brighter lights are also more attractive and for this reason it is also recommended that light intensity is kept as low as possible near turtle nesting beaches (Witherington and Bjorndal 1991a, 1991b; Witherington and Martin 2003; Pendoley and Kamrowski 2015).
As lighting technology evolves, low pressure sodium (LPS) and high pressure sodium (HPS) lights are fast being replaced with energy-efficient light emitting diodes (LEDs) (Dudley et al. 2015; Hecht 2016; Franz and Wenzl 2017). This can be a problem because of the relatively high proportion of short wavelength (blue) light that some LEDs produce (Pawson and Bader 2014; Longcore et al. 2015). The few beach-based studies available on the impacts of LEDs have confirmed that hatchlings are attracted to these lights during sea-finding (Karnad et al. 2009; Robertson et al. 2016) and one laboratory study showed a similar effect on in-water orientation in a pool (Cruz et al. 2018). To our knowledge, this has not been tested in situ during the in-water phase of hatchling dispersal from a nesting beach.
Here, we use passive acoustic telemetry to determine if wild flatback turtle hatchlings (N. depressus) are attracted to LEDs on entering the nearshore waters from nesting beaches in Western Australia. In this region many of their nesting sites are located close to large-scale illuminated industrial infrastructure that extend beyond the shoreline, so light pollution has been identified as a major pressure to this species both on land and at sea (Department of Biodiversity, Conservation and Attractions 2017). We examine the effect of LED light intensity on attractiveness and on predation rates, which may result from the greater time spent in nearshore waters as a result of misorientation, disorientation or light trapping.
Material and methods
The study site was on the north-western side of Thevenard Island, ~30 km offshore from the mainland of Western Australia (Fig. 1). This site was selected as it was least affected by existing sources of artificial light on and around the island. Nesting of female flatback turtles occurs around the entire island from November to February but is concentrated on the beaches on the south-eastern part of the island. After an incubation duration of ~47 days (Pendoley et al. 2014), hatchlings emerge from underground nests mostly at night when sand temperatures drop (Limpus 1971), crawl to the water’s edge and swim away from shore. We collected 68 hatchlings from seven nests on the south-eastern part of the island, either from within (in the top 15 cm of the nest) or near (on the sand after emergence) their nests on the nights of 18, 19 and 20 February 2017. Straight carapace length (SCL), straight carapace width (SCW) (±0.1 cm) and weight (±0.1 g) of each hatchling was measured using digital callipers and a digital scale. Hatchlings were kept in an insulated box in a dark, quiet room until the experimental trials (24- or 48-h holding time), and then were released as part of the experiment after moon set during the last quarter moon, on the nights of 20 and 21 February 2017.

|
Prior to release, a uniquely coded Vemco V5 acoustic tag (180 kHz, 0.65 g in air and 12.7 mm long) was glued to the underside of each hatchling using a small drop of fast-acting epoxy (Selleys Araldite 5 min Adhesive). It was aligned in a vertical position with the transducer pointing down (Thums et al. 2016). The tags weighed 1.82% (weight in air) of the mean weight (35.8 ± 2.9 g) of hatchlings. Owing to the vertical alignment of the tag and the fact that hatchlings can still orient at sea even without a beach crawl (Lohmann et al. 1990), instrumented hatchlings were carried to the water’s edge and released.
At the experimental site, an array of 36 (9 × 4) acoustic receivers (Vemco VR2W) was deployed in the nearshore zone to calculate geographic positions of hatchlings as they dispersed through the nearshore zone following the approach of Thums et al. (2016) (Fig. 1, see Supplementary material). A 4 m-long aluminium boat was anchored along the most seaward (northern-most) line of receivers, either on the eastern (Night 1: bearing of 35.3° from the hatchling release point) or the western (Night 2: bearing of −44.4° from the hatchling release point relative to 0° such that westerly bearings were negative) side of the array (Fig. 1). Each night we examined hatchling dispersal through the acoustic receiver array under ambient conditions (no artificial light), and with five different intensities (10, 30, 50, 70 and 120 W) of white (4000 K) LED floodlights located on the boat (Supplementary Tables S1, S2). The highest intensity (120 W) was selected to meet the equivalent lumen output of a 400-W metal halide (MH) or HPS light, which attracted hatchlings at this site previously (Wilson et al. 2018), and intensity stepped down thereafter. All of these floodlights were highly visible from the beach. The order of the treatments was randomised each night by a person located on the boat that switched the lights on and off (Supplementary Table S2). The lights were angled slightly downwards and facing the release point of hatchlings on the beach, creating a light spill on the water. Two anchors held the boat (and the light) in a fixed position throughout each experiment.
A Nortek Aquadopp Acoustic Doppler Current Profiler (ADCP) was deployed near the middle of the receiver array between the first and second row of receivers (Fig. 1) to measure current speed and direction, water temperature and depth (measured every 2 min with 1-min mean sampling at 2 Hz, then 1 min off) and wave period, height and direction, with the latter measured every 30 min based on 20-min burst sampling at 2 Hz. Data on wind speed and direction (measured at 1-min intervals) were obtained from the Bureau of Meteorology at Onslow airport, ~27 km from the site.
Each night we allocated 30 min for each treatment with three pairs of tagged hatchlings (or two pairs and one individual when there were only five hatchlings in a treatment) released every 5 min into the receiver array within each treatment. Depending on how quickly the hatchlings dispersed through the receiver array, each hatchling pair release resulted in a maximum time exposed to the treatment of 30, 25 and 20 min respectively. This time was selected based on previous in-water studies with flatback turtle hatchlings that were found to spend a mean time of 24 min in a similar sized array when exposed to a 400-W MH light (Wilson et al. 2018). Hatchlings collected from multiple nests or at different times were distributed evenly among experimental treatments. Data from acoustic receivers were downloaded the day after the last experiment and data were sent to Vemco for calculation of turtle geographic positions as they moved through the array.
As ocean currents are known to influence hatchling trajectories (Wilson et al. 2018), we conducted the experiments on the neap tide when current speeds were low. Experiments began before low water (Supplementary Table S2), which was predicted by the Bureau of Meteorology to occur at 2328 (1.21 m) and 0133 (1.2 m) hours on the first and second night respectively.
All research protocols included in this paper have been approved by the Western Australia Department of Biodiversity, Conservation and Attractions (licence number SF011007), the University of Western Australia Animal Ethics Committee (RA/3/100/1423) and comply with the Western Australia Animal Welfare Act 2002 and the requirements of the Australian code for the care and use of animals for scientific purposes.
Light measurements
Light in the environment was quantified using a digital camera (Sky42) that consisted of a Canon PowerShot G12 camera with an unfiltered Raynox DCR-CF187PRO HD fish eye lens that enabled the entire night sky and horizon to be captured in a single circular image. The camera was deployed on the beach close to the hatchling release point and photos of the night sky were taken in each treatment. Sky Quality Camera (ver. 1.3.0.0, Euromix Ltd, Slovenia) software was used to process the images and quantify the brightness (magnitude per arcsecond2) in each treatment. Light signals from stars and the Milky Way were removed using a Milky Way brightness model that extracted known stars. Data smoothing was applied for visual display and the brightness was calculated at target bearings relative to the camera location.
The spectral characteristics of the LED lights were measured in the laboratory using an Ocean Optics USB2000 Spectrometer that recorded wavelengths between 200 and 1100 nm. The lights were aimed at a photographic reflector located 2 m away and the spectrometer was aimed at the reflector from a fixed spot (~2.1 m from the reflector and in line with the light). The reflector was used to reduce the brightness of the light so that the spectrometer could be used to measure wavelengths at close range. The proportion of light emitted in each 0.38-nm wavelength band between 365 and 550 nm (wavelengths most attractive to flatback hatchlings; Pendoley 2005; Fritsches 2012) was calculated.
Hatchling movement measurements (bearing, speed and time spent)
Detections of tagged hatchlings by the acoustic receivers were used to determine if lights influenced how long hatchlings spent in the nearshore. We calculated the time spent by each turtle in the receiver array, defined as the time from when each turtle was first detected by the receivers in the array until the time it was last detected in the array. To test for an effect of light intensity on hatchling rate of travel and direction, geographic positions from each hatchling were used to calculate mean speed and mean bearing (relative to 0° such that westerly bearings were negative) from the release point to the point where it either left the array, or where it was last detected within the tracking area. The speed between successive positions (the distance between each position divided by the time difference between the positions) was calculated using the R (ver. 3.6.3, R Foundation for Statistical Computing, Vienna, Austria, see https://www.R-project.org/) package ‘sp’ (ver. 1.2-5, E. J. Pebesma and R. S. Bivand, see https://cran.r-project.org/web/packages/sp/index.html; Bivand et al. 2013) and the mean of all these speeds was determined for each hatchling. The R package ‘geosphere’ (ver. 1.5-5, R. J. Hijmans, see https://cran.r-project.org/packages/geosphere/index.html) was used to calculate the bearing between each position and R package ‘circular’ (ver. 0.4-7, C. Agostinelli and U. Lund, see https://r-forge.r-project.org/projects/circular) was used to calculate the mean bearing of each hatchling.
Data collected by the ADCP and the Bureau of Meteorology was used to calculate several physical parameters (to use as predictor variables in models) for the period of each turtle’s transit through the receiver array: mean wave steepness (wave height/wavelength), wave height, wave period, current speed and direction, water depth, water temperature, and wind speed and direction. Peak wave direction was not calculated as wave height was too low (<10 cm) to accurately determine a direction.
Impact of light intensity on hatchling bearing
Using the mean bearing of hatchlings, we calculated the circular mean bearing and r-value for each treatment each night. The r-value ranges from 0 to 1 and is a measure of directional variance; the higher the value the more consistency in the direction of that group (Batschelet 1981). To determine if hatchling bearings within each treatment were significantly oriented as a group, we conducted a Rayleigh test of uniformity (Batschelet 1981) and for those treatments that were found to have significant orientation (P < 0.05) we determined whether the mean direction of the hatchlings in that treatment aligned with the direction of the light source by plotting the mean bearing and the 95% confidence intervals (CI) of the mean. If the CI overlapped the light source, then we determined that the hatchlings bearings aligned with the light. For treatments where this occurred, we overlayed the CI for the ambient treatment to determine if it differed from ambient. All of these analyses were conducted using functions from the CircStat2012 toolbox (Berens 2009) in Matlab (ver. R2015b, The MathWorks Inc., Natick, MA, USA).
Impact of light intensity on hatchling mean speed and time spent
To determine whether it was appropriate to pool the data across the two nights for the modelling of the response variables; mean speed and time spent, we tested each response variable with Night (Categorical: 1 or 2) using a linear model and compared the Akaike’s information criterion, corrected for small sample size (AICc) and model weights of the slope model (response ~Night) to the intercept-only model (response ~1). We also used the same modelling framework to test if there was a difference in predictor variables (see above) between the two nights. We found majority support (≥91%) for an effect of night (wAICc ≥ 0.91) on both the response and the majority of the predictor variables, thus data from each of the two nights were analysed separately (see Supplementary Fig. S1).
The response variables mean speed and time spent were used as response variables in a suite of generalised additive models to examine the relationship with the continuous variables: light treatment (intensity: 0, 10, 30, 50, 70, 120 W), current speed, wave steepness, wave period, wind direction and wind speed for each night. Wave height, current direction, water depth, water temperature, hatchling weight, SCW (mm) and SCL (mm) were excluded from the analysis as they had little variation to model. For the same reason, wave period was removed from the modelling on Night 1 when the light was located to the east and wind speed and wind direction were removed on Night 2 (light west). The gam or gamm function in the mgcv library (ver. 1.8-17, see https://cran.r-project.org/web/packages/mgcv/index.html; Wood 2017) in R was used to fit models using all combinations of the predictor variables. We used a function developed by Fisher et al. (2018) to compile all the possible combinations and extract and compile the parameters used for model selection; the AICc, and the relative model weight; the AICc weight (ωAICc). All continuous predictors were modelled as smooths, using a cubic regression spline, with k restricted to 5 following the default settings of the full subsets GAM function used in the FSSgam library (ver. 1.11, R. Fisher, see https://github.com/beckyfisher/FSSgam; Fisher et al. 2018).
Mean speed was modelled with a Gaussian distribution and time spent in the receiver array was rounded to the nearest minute and then modelled as a binomial distribution with 20 trials, representing a maximum of 20 possible minutes (consistent across all turtles released) in the array. Each individual turtle (turtle ID) was included as a random effect in models with time spent. All values presented are mean ± standard deviation unless otherwise stated.
Association between light intensity and predation rates
Since previous studies have demonstrated that predation of acoustically tagged hatchlings can be identified by assessing the movement patterns of the tags in the receiver array (Thums et al. 2016; Wilson et al. 2019), we also determined the number of hatchlings that had interactions with predators. Tagged hatchlings that moved in a directed path offshore or towards the lights with no further detections in the array after ~20 min were categorised as ‘dispersed’ (Thums et al. 2016; Wilson et al. 2018). Tagged hatchlings that continued to be detected by the receivers in the array for much longer than the expected residence time (i.e. hours) were considered to be the result of an encounter with a predator and categorised as ‘predated’ (as per Wilson et al. 2019). These tags had been either ingested by fish predators or detached from the hatchlings, likely during predator strike or handling. The former were identified by complex movement patterns rather than simple directed trajectories through the array, whereas the latter were identified by hundreds to thousands of detections on a small (1–4) number of receivers over an extended period of time (hours–days) (see Supplementary Fig. S2).
Movement paths of predated tags were examined to determine the likely location of the predation event at the time the tag detached or was ingested. Positions representative of turtle behaviour before the predator encounter (i.e. where it occurred) were extracted and used in the calculation of mean speed and bearing of each hatchling. Predation rates were calculated by dividing the number of tags categorised as predated by the total number of turtles released in each treatment. We used a Chi-Square test of independence to test the hypothesis that the number of hatchlings predated in light treatments (all intensities) was different (at significance level; a = 0.05) from the number predated in ambient treatments.
Results
Environmental conditions
Most environmental parameters at the study site varied between experimental nights (Supplementary Fig. S1). Although wave height was consistent over the two nights (0.06 ± 0.01 m) (Supplementary Fig. S1a), wave period on Night 1 when lights were positioned to the east was dominated by relatively steep (0.0007 ± 0.0001) wind waves (period 7.38 ± 0.46 s). On Night 2 when lights were towards the west, waves were less steep (0.0004 ± 0.00004) and were swell dominated (period 9.51 ± 1.25 s) (Supplementary Fig. S1b, c). Mean current velocity was slightly higher on Night 1 than Night 2 (0.08 ± 0.02 and 0.06 ± 0.02 m s−1 respectively) (Supplementary Fig. S1d). Currents were driven by tidal movement and flowed towards the east (67.9–119°) on both nights (Supplementary Fig. S1e). Median water depth on Night 1 was 1.65 m, ranging from 1.57 to 1.85 m (Supplementary Fig. S1f). On Night 2, median depth was 1.51 m, ranging from 1.38 to 1.71 m (Supplementary Fig. S1f). Mean water temperature on Night 1 and 2 was 32.4 ± 0.1 and 31.6 ± 0.3°C respectively (Supplementary Fig. S1g). On Night 1, winds approached from the WSW (249.3 ± 27.5°) at speeds of 11.7 ± 3.3 km h−1 (Supplementary Fig. S1h, i). On the following night, winds were stronger (24.2 ± 2.6 km h−1) and came from the SSW (195.3 ± 2.3°) (Supplementary Fig. S1h, i).
Hatchling measurements
The SCL and SCW of hatchlings that were tagged as part of the experiment were 60.7 ± 2.1 mm and 48.5 ± 1.7 mm respectively, and weighed 35.8 ± 2.9 g (Supplementary Table S2). On Night 1, when lights were positioned to the east, hatchlings were all obtained from one nest, and on Night 2, when lights were on the western side, they were sourced from six different nests (Supplementary Table S2). On Night 1, hatchling weight (33.3 ± 1.0 g) and SCL (59.7 ± 2.0 mm) was lower and SCW (49.2 ± 1.3 mm) was higher than on Night 2 (weight 38.5 ± 1.3 g, SCL 61.9 ± 1.5 mm and SCW 47.7 ± 1.7 mm) (Supplementary Fig. S1, Supplementary Table S2).
Light camera
Zenith sky quality under ambient conditions measured 23.0 magnitude per arcsecond (mag arcsec–2: the larger the value, the darker the night sky), however, regions of horizon sky glow measuring 20.63 ± 0.21 mag arcsec–2 were visible over the dunes in a southerly and north-easterly direction originating from industrial and urban sources (Supplementary Fig. S3). When the lights of our experimental treatments were switched on, the camera measured high, uni-directional light from the light source, with the brightest point of light in all treatments measuring 16.3 ± 0.35 mag arcsec–2 (averaged across all light treatments, except ambient). The extent of the sky affected by glow increased with light intensity but there was only a small difference between some of the treatments (e.g. between the 30- and 50-W and the 50- and 70-W treatments) (Supplementary Fig. S3). In all treatments, the light spill remained confined to the region of sky around the experimental light on the boat (Supplementary Fig. S3). There was an increase in brightness on the horizon between angles 90–210° in the 30–120-W treatments due to light from our vessel reflecting off shoreline dunes (Supplementary Fig. S3).
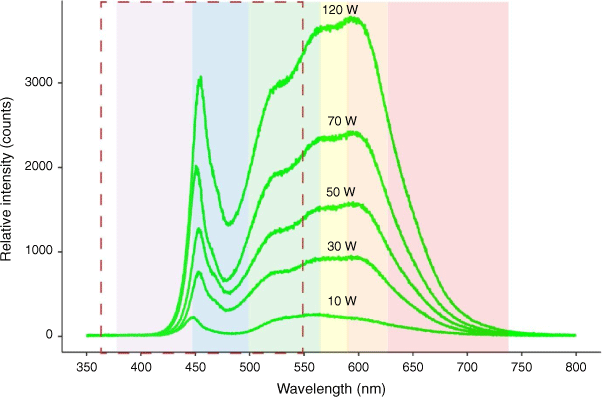
The spectral power distribution for the LEDs ranged from 340 to 1024 nm with emissions peaking at wavelengths of ~455 and 590 nm (Fig. 2). Approximately 40% of the light emitted from these lights (ranging from 39.1 to 42.2% for all five wattages) was the type of light shown to be most disruptive to the sea-finding ability of flatback turtles (365–550 nm; Pendoley 2005; Fritsches 2012).
|
Acoustic tracking
A total of 67 out of 68 tags were detected by receivers, ranging from 61 to 3915 detections per individual tag (one tag was never detected, likely due to tag malfunction or the tag not being turned on correctly, or the hatchling may have left the receiver array quickly before being detected). Of these detections, 2910 geographic positions were calculated for 65 out of the 67 tags (median 28 positions per individual, ranging from 0 to 463: median Horizontal Positioning Error of location calculation was 1.3 m (range = 0.8–1778 m). The two hatchlings that did not have geographic positions were detected hundreds of times, primarily on a single receiver located near the release point, suggesting that the tags of both had detached near the release point. Patterns in detections and geographic positions were used to identify an additional 10 tags that had detached from hatchlings either close to their release point (n = 5) or during transit (n = 5). In all of these cases, tag detachment was unlikely to be caused by glue failure and was most likely to be the result of prey handling during a predation event given their vertical placement, which makes them more likely to be dislodged during a predation attempt (Wilson et al. 2019). As such, hatchlings with tags that detached were categorised as ‘predated’.
The tags that detached near the release location (n = 7 which includes the two hatchlings that did not have geographic positions) were excluded from the modelling of speed and bearing (Supplementary Table S3). For those that were dislodged during transit (n = 5), positions prior to tags detaching were included in the modelling of mean speed and the analysis of mean bearing. Another three hatchlings were attacked by predators that ingested the tags attached to the turtle. For one of these tagged hatchlings, positions before the predation event were used in the analysis of speed and bearing as this was easily identified by assessing the movement patterns of the tags (see Wilson et al. 2019). The other two ingested tags were omitted from modelling entirely as it was difficult to identify from the movement pattern exactly when the predation event occurred. In addition, one hatchling moved at very slow speeds (<0.1 m s−1 for more than 50% of the time), which was much slower than previously documented hatchling speeds (0.5 m s−1; Wilson et al. 2018), however, the movement pattern was not entirely different as it took a directed path away from shore. As it was unclear whether this track was a turtle or a slow-moving predator, it was excluded from all analyses. In total, movement patterns from 66 hatchlings were used to calculate predation rates, positions from 57 hatchlings were used to calculate and model speed and to determine the treatments where hatchling bearing aligned with the light, and detections and positions of 51 turtles that successfully transited the receiver array (excluding predated tags; 12 detached and 3 ingested), were used in the calculations and modelling of time spent in the array.
Hatchling movement in relation to artificial light position
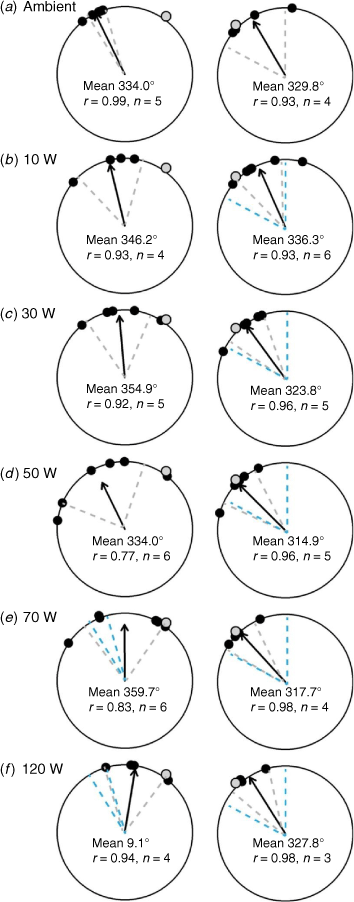
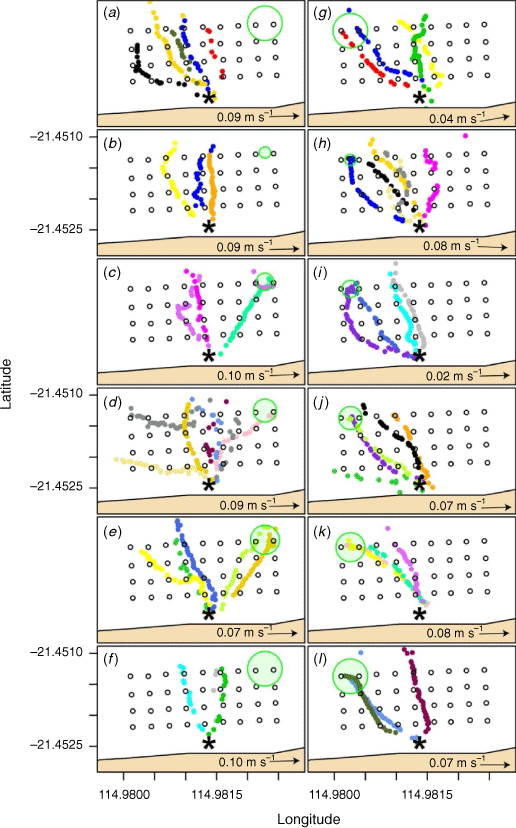
In total, 51 of the 66 (77%) hatchlings that were tagged and released in this experiment successfully transited the array. Of these, 25 were released on Night 1 when lights were positioned on the eastern side of the array and 26 were released on Night 2 when the lights were located on the western side of the array. In general, hatchlings released when lights were positioned to the east moved in a more northerly direction (−10.9° from north, 0°) compared to when lights were positioned to the west (−34.7°) (black arrows showing mean bearing in Supplementary Fig. S4). All hatchlings released under ambient conditions moved towards the north or north-west and did not disperse towards the east (Fig. 3a, 4a, g). Similarly, the mean bearing of hatchlings in each treatment was towards the north or to the north-west under all five light treatments on both nights (Fig. 3, 4b–f, h–l).
|
|
Hatchlings oriented significantly as a group (P < 0.05) in all treatments across both nights. The r-value by treatment was highest (r = 0.99) under ambient conditions on Night 1 when the boat was moored to the east (mean bearing 334°; 95% CI 326–342°) (Fig. 3). Plots of the raw data show that the mean bearing of hatchlings was relatively consistent across all treatments on Night 2 (light source located to the west), but it became more variable on Night 1 when lights were present (Fig. 3). The r-value was lowest in the 50-W treatment on Night 1 (r = 0.77) and the mean bearing aligned with the light direction only in the two brightest treatments (70 and 120 W) (Fig. 3). Of these two treatments, only the brightest treatment differed from ambient. By contrast, when lights were located to the west of the hatchling release point, the mean bearing aligned with the light direction in all of the light treatments, with a similar range in CI around the mean in all treatments (Fig. 3). The CI around the mean in all treatments was similar to the CI around the mean for the ambient treatment making it difficult to tease out any effect of light treatment on mean bearing (Fig. 3).
Impact of light intensity on predation rates
Out of the 66 turtles, a total of 12 tags detached (18%) and three were ingested (5%), which resulted in an overall predation rate of 23% of hatchlings (see Supplementary Fig. S2 for an example of the movement and detection patterns of tags assigned to each category). In general, predation was higher (29%) when lights were positioned to the east compared to the west (16%). During the experiments when lights were positioned to the east, predation only occurred during light treatments (i.e. no predation events occurred in the ambient treatment) and predation rates were highest (50%) during the 10, 30 and 120-W treatments but were not significantly (at a = 0.05) different from ambient, however, for three out of five treatments the P-value of the Chi-Square test statistic was near significant (P = 0.06) (Table 1). When the light was positioned to the west, predation rates ranged from 0 to 25% in the different treatments, with the highest (25%) coinciding with the brightest light intensity (120 W). However, 20% of hatchlings were also predated under ambient conditions and predation rates in light treatments were also not significantly different from predation rates in the ambient treatment (P > 0.05 for all comparisons) (Table 1). Thus, there was weak evidence to support the hypothesis that light increases hatchling predation when lights were positioned to the east but not enough evidence to support the hypothesis when lights were positioned to the west.

|
Mean speed and time spent in the array
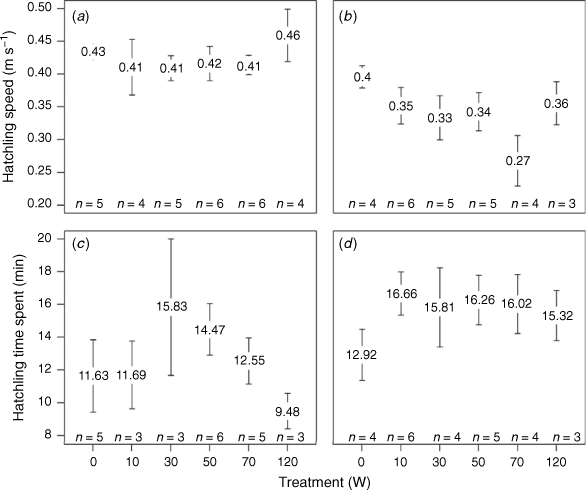
In total, hatchlings spent between 7.38 and 40.72 min in the receiver array across all treatments and nights, with a median time of 14.23 min. Our preliminary assessment of the effect of night (as outlined in the methods) on hatching speed and time spent showing majority support (100% (wAICc = 1) and 91% (wAICc = 0.91) respectively) for the model with night compared to the null model, showed that hatchlings swam faster on Night 1 (light to the east) than they did on Night 2 (light to the west) (0.42 ± 0.05 and 0.34 ± 0.07 m s−1 respectively) and they took longer to transit the array on Night 2 (15.62 ± 3.4 min) than they did on Night 1 (12.75 ± 4.3 min).
When testing the impact of light intensity on mean speed and time spent in the array light intensity was the top ranked model for hatchling speed on Night 2 only; as light intensity increased, hatchling speed generally decreased (Supplementary Fig. S5), however, there was only weak support (25%) for this model (ωAICc = 0.25) (Supplementary Table S4) and the null model was the most parsimonious (within two AICc points of the top ranked model). For mean speed on Night 1, and for time spent on both nights, the null model gained highest support (Supplementary Table S4), with plots of the raw data showing that mean speed was relatively consistent across intensities on Night 1 (Fig. 5a) and that time spent in the receiver array was highly variable on that night (Fig. 5c). Even though the models found no effect of light intensity on time spent in the array, it appeared that the time that hatchlings spent in the array was greater in the light treatments than the ambient treatment on Night 2 (Fig. 5d). The pattern mirrored that of mean speed suggesting that lower speeds in light treatments resulted in more time spent in the array than in ambient conditions (Fig. 5b, d).
|
Discussion
Using passive acoustic telemetry and a range of different intensities of LEDs, we found a significant change in hatchling bearing from ambient in the brightest treatment (120 W) only, but only when lights were positioned to the east of the release location, the opposite direction that hatchlings preferred to disperse towards under ambient conditions. We also found weak evidence (P = 0.06) for an impact of low as well as high intensity light sources (10, 30 and 120 W) on predation rates on one of the study nights. This suggests that the presence of LEDs, regardless of their intensity, may have a negative effect on the survivorship of hatchlings passing through the nearshore environment. In addition, the location of light sources in relation to hatchling general direction of travel may be important in determining its impact.
The variable response of hatchlings to the different intensity LED lights resulted in light intensity being only a weak predictor of hatchling speed and the time that they spent in the receiver array. This outcome was likely driven by the directional bias of hatchlings towards the north or north‐west that we detected in the ambient treatments and also the low sample size that resulted after excluding those that were taken by predators (up to 50% of hatchlings in some treatments). Although the experimental plan was to have five or six hatchlings in each treatment, predation very early after release resulted in as little as three individuals in some treatments. While recognising this limitation on sample size, our study did still find a weak effect of light intensity on hatchling speed, which suggests that a stronger effect may have been detected with a larger number of hatchlings. The small difference in the spatial extent of sky glow and intensity between the different light treatments (see Supplementary Fig. S3) may have also contributed.
The location of the light in relation to the hatchlings’ general direction of travel appears to play a role in the degree of disturbance. Most hatchlings under ambient (i.e. dark) conditions tended to move north or north-west through the receiver array, but none of the hatchlings ever moved east. This is consistent with findings from Wilson et al. (2018) from the same location where hatchlings moved mostly towards the north or towards the north-west under ambient conditions. In this study, when the light source was placed to the east of the release point, most of the hatchlings appeared to not be drawn towards it but rather, moved in a similar direction to hatchlings released under ambient conditions and the mean bearing of hatchlings only aligned with the light source in the two brightest treatments, but it was only in the 120-W treatment that the direction of hatchlings differed to the direction of hatchlings in the ambient treatment. The fact that lights lower than 120 W had little effect on hatchling behaviour on this night matches with theory which suggests that brighter lights are more attractive to hatchlings (Pendoley and Kamrowski 2015) but may also be due to the presence of short period sea waves that likely approached from the same direction as the wind (WSW) which may have acted as a stronger cue, overriding the light cue. Indeed, flatback turtle hatchlings have been shown to respond to this type of wave in a wave flume (Wilson et al. 2020). Green turtle hatchlings (Chelonia mydas) also display a similar behaviour and head into the direction of the approaching wave (Lohmann and Lohmann 1992; Okuyama et al. 2009). By contrast, when the light was placed to the west of the release point, the mean bearing of hatchlings in all of the light treatments aligned with the light source, but hatchlings released under ambient conditions also moved in this same direction. Movement towards the north-west under ambient conditions may have also been in response to wave direction as long period swell waves persisted and likely came from the north-west as this was recorded previously as the peak wave direction at this location (Wilson et al. 2018). It seems that hatchlings here favour dispersing in a westward direction, so that their tendency to move towards the different light treatments was not due to light attraction but simply a result of their directional preference. This hierarchy of cues has been reported elsewhere (Able 1991; Salmon et al. 1992; Salmon and Wyneken 1994) and it is possible that the influence of different cues may vary as hatchlings traverse nearshore waters on the way to the relative safety of deep water (Lohmann and Lohmann 1996; Okuyama et al. 2009). However, even if hatchlings were not attracted to, but simply came across the light (as that was a general direction travelled under ambient conditions) it appears that they might have been disoriented by the offshore light source as hatchlings took on average ~20% longer to transit the array when light was present (~13 min under ambient v. 16 min across most light treatments). This suggests that, if a source of light is placed in the general direction that hatchlings are favouring, it can cause them to swim slower as they linger around the light source, delaying their dispersal, thus making them more exposed to predators in the nearshore, and this can occur irrespective of intensity since all light intensities had similar effects on time spent and hatchling speed. If this behaviour can be generalised then studies involving over-water lighting (e.g. jetties, ports, oil rigs) conducted before development of infrastructure should focus on identifying the hatchlings preferred direction of offshore orientation and, at least for flatback hatchlings, avoid placing sources of artificial light in their path.
Despite artificial light intensity not having a clear effect on hatchling speed and time spent in the nearshore, we found weak evidence on one of the experimental nights to support the hypothesis that light affects predation rate. It is possible that LED lights may have attracted predators to the area (as most predation events occurred early after release as opposed to at, or near, the light source: Supplementary Table S3) or light could have aided the process of predation in some way, such as making hatchlings easier to detect through silhouetting. The highest rate of predation occurred during the brightest (120 W) treatment across both nights (50% on Night 1 and 25% on Night 2), although predation rates were also high in lower wattage treatments (50% in the 10- and 30-W treatments on Night 1) and 20% were taken under ambient conditions on one of the nights. The type of LED light used in the experiment was enriched in short wavelengths (up to 550 nm; see Fig. 2), which are transmitted further in the ocean than longer wavelength and UV light (Guthrie 1986). Coastal fish are particularly sensitive to these wavelengths (Lythgoe and Partridge 1989) and variation in intensity can also enhance attraction (Marchesan et al. 2005) so it seems possible that these lights may have contributed to the levels of predation recorded. Indeed, to exploit the attraction of fish to artificial lights they are often used on fishing nets or vessels to improve catches (Marchesan et al. 2005; Bryhn et al. 2014) and lights have also been shown to increase the predatory behaviour of fish (Bolton et al. 2017). It is possible that more predators may have been present on Night 1 compared to Night 2 and thus influencing the results we report, however, the weak or no evidence we report for the association between predation rate and light is more likely related to issues with sample size and should be explored further.
In total, 10% of hatchlings encountered predators under ambient conditions across the two nights. This is likely to represent natural levels of mortality for hatchlings transiting the nearshore (within 300 m) at this location, since equivalent predation rates have been reported here previously (tags detached from 3 out of 29 hatchlings tracked under ambient conditions; Wilson et al. 2018). Other studies also report similar low levels (below 10%) of predation at nesting sites (Witherington and Salmon 1992; Glenn 1996; Stewart and Wyneken 2004; Whelan and Wyneken 2007).
Hatchlings did not linger as long with LED lights compared to a 400-W HPS light and MH light located in the same direction (Wilson et al. 2018). Hatchlings spent 50 and 150% longer in the nearshore when a HPS and MH light was present to the west of the release point and travel speeds reduced by 12–30% respectively (Wilson et al. 2018). By contrast, hatchlings spent 24% longer (12.9 min in the ambient treatment and ~16 min in the LED treatments combined) when LEDs were present (all intensities combined) and travel speeds reduced by ~17% (0.40 and 0.33 m s−1 in the ambient and LED light treatments respectively). Different light types cannot be directly compared as they emit different wavelengths of light, and traditional bulbs such as MH and HPS emit light in all directions whereas LEDs are unidirectional so the light output (i.e. lumens) between different lighting types are also not directly comparable. Despite this, there was an obvious difference in brightness between the LED treatments and the 400-W HPS and MH lights (see Supplementary Fig. S3), which may have contributed to the difference in time spent between these experiments. Hatchlings respond more strongly to a horizontally broad, but vertically narrow area of illumination low on their horizon (Verheijen and Wildschut 1973; Lohmann et al. 1997) in comparison to a single point source of light which could represent a star or the moon (Hooker 1911). Consequently, the LED lights may have had a weaker influence on hatchling behaviour since they acted more like a point source of light. This suggests that LEDs (less than 120 W) may be less disruptive to the movement behaviour of hatchlings than a 400-W HPS light, even though the LED emits a large proportion (~40%) of light in the blue and green range (wavelengths up to 550 nm). Despite these findings, given the low sample size and high predation experienced in many of the LED light treatments, and the fact that environmental conditions were different between this experiment and the experiment by Wilson et al. (2018), these observations must be treated with caution and require further study.
Interestingly, no predation was reported under HPS and MH lighting in a previous study at this location (Wilson et al. 2018), although the two studies differed in timing and environmental conditions. In the study by Wilson et al. (2018), hatchlings were tracked on the spring tides around high water and on the new moon, whereas the current study occurred on the neap tide just before low water during last quarter moon, but after moon set. In another study, predation of green turtle hatchings was consistently higher at low water than at high water, particularly when compared to the spring high tide, and it was also lowest under new moon conditions compared to all other moon phases (Gyuris 1994) so the differences in depth, current speeds and moon phase may have been driving the higher predation rates reported in this study instead of the LED lights.
The raw data showed that there was large variability in most of our response and predictor variables between the two nights that potentially also contributed to our inability to find consistent effects of light intensity on hatchling speed and time spent in the nearshore. Hatchlings tracked on Night 1 when lights were located to the east had lower weight, swam faster and spent less time in the receiver array than those tracked on Night 2. Water temperatures, water depth and current speeds were higher on the night that hatchlings swam faster. Hatchling speed was also higher when winds came from the west than when winds came from the south-south-west. The westerly winds on Night 1 produced wind waves (wave periods <8 s) likely approaching from the same westerly direction, and on this night, hatchlings swam faster. South-south-west winds on Night 2 produced mostly swell waves (wave period >8 s) as winds were travelling in an offshore direction from the land and hatchlings swam slower. Hatchlings can distinguish between waves of different periods and are more responsive to shorter wave periods (Manning et al. 1997; Wilson et al. 2020) so it is possible that the presence of waves with shorter periods encouraged them to quickly disperse offshore. This was suggested by Wilson et al. 2018 as the presence of steeper waves (a combination of shorter wave periods and higher wave heights) helped reduce the period of time spent in the nearshore. As the physical (wind, waves, water temperature) and biological conditions (hatchling size) between nights were so different, it was impossible to tease out the relative importance of each of these variables.
Concluding remarks
Although more effort is required to increase the confidence in our findings, this study raises many questions in regard to the effect of LEDs on this species of turtle and their predators. Given our issues with sample size and the differences in the physical and biological parameters between nights, some aspects of the impact of light intensity on hatchling in-water movement behaviour remain unresolved and underscores the difficulty of acquiring adequate replication to account for such variability in an experiment where the subjects are prone to high rates of natural predation. The type of LEDs used in this experiment (up to 120 W) increased the time that some of the hatchlings spent in the nearshore but did not cause them to linger as long as has been shown at this site previously with MH and HPS lights (Wilson et al. 2018). This suggests that the impact of LEDs on the time spent in the nearshore may be lower than with MH and HPS, especially at these relatively lower wattages and when the light source is placed outside of their usual direction of travel. However, although lower intensity lights (<120 W) may not have resulted in hatchling attraction, there is suggestion that they may be more attractive to their predators as predation rates were near to being significantly (P = 0.06) higher when LED light was present compared to ambient conditions for some treatments. Given predation rates on hatchlings can be high in nearshore areas (Gyuris 1994; Pilcher et al. 2000; Reising et al. 2015; Wilson et al. 2019) and that the brightest light influenced the direction of dispersal, the use of white LED floodlights in coastal areas, even at low intensity, appear to pose a threat to hatchlings and perhaps light avoidance or other measures such as task lighting, shielding, and avoiding shining light directly on the water might be more appropriate mitigation measures than simply reducing intensity.
Data availability
The data that support this study are available at the Australian Institute of Marine Science Data Centre at https://apps.aims.gov.au/metadata/view/cbec5004-701b-436f-b286-a99c476c0881.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Declaration of funding
P. Wilson was supported by an Australian Government Research Training Program (RTP) Scholarship and a top-up scholarship from The University of Western Australia. Project funding was provided predominantly from the Department of Biodiversity, Conservation and Attractions through the North-west Shelf Flatback Turtle Conservation Program with contributions from all author organisations.
Supplementary material
Supplementary material is available online.
Acknowledgements
We thank Rebecca Fisher for her advice on experimental design and data analysis, Luciana Ferreira, Selina Warburton, Alex Mason and Minh Vo for their assistance with field experiments and Adam Mitchell for processing the light camera images. We also thank Rick Roberts for donating the boat that was used for the experiments. The authors appreciate the valuable comments provided by reviewers on earlier versions of this manuscript.
References
Able, KP (1991). Common themes and variations in animal orientation systems. American Zoologist 31, 157–167.| Common themes and variations in animal orientation systems.Crossref | GoogleScholarGoogle Scholar |
Batschelet E (1981) ‘Circular Statistics in Biology.’ (Academic Press: London, UK)
Becker, A, Whitfield, AK, Cowley, PD, Jarnegren, J, and Naesje, TF (2013). Potential effects of artificial light associated with anthropogenic infrastructure on the abundance and foraging behaviour of estuary-associated fishes. Journal of Applied Ecology 50, 43–50.
| Potential effects of artificial light associated with anthropogenic infrastructure on the abundance and foraging behaviour of estuary-associated fishes.Crossref | GoogleScholarGoogle Scholar |
Berens, P (2009). CircStat: a Matlab toolbox for circular statistics. Journal of Statistical Software 31, 1–21.
| CircStat: a Matlab toolbox for circular statistics.Crossref | GoogleScholarGoogle Scholar |
Bivand R, Pebesma E, Gomez-Rubio V (2013) ‘Applied spatial data analysis with R’, 2nd edn. (Springer: New York, NY, USA)
Bolton, D, Mayer-Pinto, M, Clark, GF, Dafforna, KA, Brassil, WA, Becker, A, and Johnston, EL (2017). Coastal urban lighting has ecological consequences for multiple trophic levels under the sea. The Science of the Total Environment 576, 1–9.
| Coastal urban lighting has ecological consequences for multiple trophic levels under the sea.Crossref | GoogleScholarGoogle Scholar | 27780095PubMed |
Bryhn, AC, Konigson, SJ, Lunneryd, S-G, and Bergenius, MAJ (2014). Green lamps as visual stimuli affect the catch efficiency of floating cod (Gadus morhua) pots in the Baltic sea. Fisheries Research 157, 187–192.
| Green lamps as visual stimuli affect the catch efficiency of floating cod (Gadus morhua) pots in the Baltic sea.Crossref | GoogleScholarGoogle Scholar |
Cinzano, P, Falchi, F, and Elvidge, CD (2001). The first World Atlas of the artificial night sky brightness. Monthly Notices of the Royal Astronomical Society 328, 689–707.
| The first World Atlas of the artificial night sky brightness.Crossref | GoogleScholarGoogle Scholar |
Cruz, LM, Shillinger, GL, Robinson, NJ, Tomillo, PS, and Paladino, FV (2018). Effect of light intensity and wavelength on the in-water orientation of olive ridley turtle hatchlings. Journal of Experimental Marine Biology and Ecology 505, 52–56.
| Effect of light intensity and wavelength on the in-water orientation of olive ridley turtle hatchlings.Crossref | GoogleScholarGoogle Scholar |
Department of Biodiversity, Conservation and Attractions (2017) ‘North West Shelf Flatback Turtle Conservation Program Strategic Conservation Plan 2014–21.’ (Department of Biodiversity, Conservation and Attractions: Perth, Western Australia)
Dudley, JM, Erkintalo, M, and Genty, G (2015). Environment, wildlife and LED illumination. Optics and Photonics News 26, 42–47.
| Environment, wildlife and LED illumination.Crossref | GoogleScholarGoogle Scholar |
Erb, V, and Wyneken, J (2019). Nest-to-surf mortality of loggerhead sea turtle (Caretta caretta) hatchlings on Florida’s east coast. Frontiers in Marine Science 6, 271.
| Nest-to-surf mortality of loggerhead sea turtle (Caretta caretta) hatchlings on Florida’s east coast.Crossref | GoogleScholarGoogle Scholar |
Fisher, R, Wilson, SK, Sin, TM, Lee, AC, and Langlois, TJ (2018). A simple function for full subsets multiple regression in ecology with R. Ecology and Evolution 8, 6104–6113.
| A simple function for full subsets multiple regression in ecology with R.Crossref | GoogleScholarGoogle Scholar | 29988441PubMed |
Franz, M, and Wenzl, FP (2017). Critical review on life cycle inventories and environmental assessments of LED-lamps. Critical Reviews in Environmental Science and Technology 47, 2017–2078.
| Critical review on life cycle inventories and environmental assessments of LED-lamps.Crossref | GoogleScholarGoogle Scholar |
Fritsches, KA (2012). Australian loggerhead sea turtle hatchlings do not avoid yellow. Marine and Freshwater Behaviour and Physiology 45, 79–89.
| Australian loggerhead sea turtle hatchlings do not avoid yellow.Crossref | GoogleScholarGoogle Scholar |
Gaston, KJ, Davies, TW, Bennie, J, and Hopkins, J (2012). Reducing the ecological consequences of night-time light pollution: options and developments. Journal of Applied Ecology 49, 1256–1266.
| Reducing the ecological consequences of night-time light pollution: options and developments.Crossref | GoogleScholarGoogle Scholar |
Glenn L (1996) The consequences of human manipulation of the coastal environment on hatchling loggerhead sea turtles (Caretta caretta, L). In ‘Proceedings of the Sixteenth Annual Symposium on Sea Turtle Biology and Conservation’, 28 February–1 March 1996, Hilton Head, SC, USA. (Eds R Byles, Y Fernandez) pp. 58–59. (NOAA)
Guthrie DM (1986) Role of vision in fish behaviour. In ‘The behaviour of teleost fishes’. (Ed. TJ Pilcher) pp. 75–113. (Billing & Sons Limited: Worcester, UK)
Gyuris, E (1994). The rate of predation by fishes on hatchlings of the green turtle (Chelonia mydas). Coral Reefs 13, 137–144.
| The rate of predation by fishes on hatchlings of the green turtle (Chelonia mydas).Crossref | GoogleScholarGoogle Scholar |
Harewood, A, and Horrocks, J (2008). Impacts of coastal development on hawksbill hatchling survival and swimming success during the initial offshore migration. Biological Conservation 141, 394–401.
| Impacts of coastal development on hawksbill hatchling survival and swimming success during the initial offshore migration.Crossref | GoogleScholarGoogle Scholar |
Hecht, J (2016). The early-adopter blues. IEEE Spectrum 53, 44–50.
Hooker D (1911) Certain reactions to color in the young loggerhead turtle. In ‘Papers from the Marine Biological Laboratory at Tortugas Laboratory of the Carnegie Institution of Washington’. pp. 69–76. (Carnegie Institution of Washington: Philadelphia, PA, USA)
Kamrowski, RL, Limpus, C, Pendoley, K, and Hamann, M (2014). Influence of industrial light pollution on the sea-finding behaviour of flatback turtle hatchlings. Wildlife Research 41, 421–434.
| Influence of industrial light pollution on the sea-finding behaviour of flatback turtle hatchlings.Crossref | GoogleScholarGoogle Scholar |
Karnad, D, Isvaran, K, Kar, CS, and Shanker, K (2009). Lighting the way: Towards reducing misorientation of olive ridley hatchlings due to artificial lighting at Rushikulya, India. Biological Conservation 142, 2083–2088.
| Lighting the way: Towards reducing misorientation of olive ridley hatchlings due to artificial lighting at Rushikulya, India.Crossref | GoogleScholarGoogle Scholar |
Limpus, CJ (1971). The flatback turtle, Chelonia depressa Garman in Southeast Queensland, Australia. Herpetologica 27, 431–446.
Lohmann, KJ, and Lohmann, CMF (1992). Orientation to oceanic waves by green turtle hatchlings. The Journal of Experimental Biology 171, 1–13.
| Orientation to oceanic waves by green turtle hatchlings.Crossref | GoogleScholarGoogle Scholar |
Lohmann, KJ, and Lohmann, CMF (1996). Orientation and open-sea navigation in sea turtles. The Journal of Experimental Biology 199, 73–81.
| Orientation and open-sea navigation in sea turtles.Crossref | GoogleScholarGoogle Scholar |
Lohmann, KJ, Salmon, M, and Wyneken, J (1990). Functional autonomy of land and sea orientation systems in sea turtle hatchlings. The Biological Bulletin 179, 214–218.
| Functional autonomy of land and sea orientation systems in sea turtle hatchlings.Crossref | GoogleScholarGoogle Scholar | 29314980PubMed |
Lohmann KJ, Witherington B, Lohmann CM, Salmon M (1997) Orientation, navigation, and natal beach homing in sea turtles. In ‘The Biology of Sea Turtles. Vol. 1’. (Eds PL Lutz, JA Musick) pp. 107–136. (CRC Press, Inc.: Boca Raton, FL, USA)
Longcore, T, and Rich, C (2004). Ecological light pollution. Frontiers in Ecology and the Environment 2, 191–198.
| Ecological light pollution.Crossref | GoogleScholarGoogle Scholar |
Longcore, T, Aldern, HL, Eggers, JF, Flores, S, Franco, L, Hirshfield-Yamanishi, E, Petrinec, LN, Yan, WA, and Barroso, AM (2015). Tuning the white light spectrum of light emitting diode lamps to reduce attraction of nocturnal arthropods. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 370, 20140125.
| Tuning the white light spectrum of light emitting diode lamps to reduce attraction of nocturnal arthropods.Crossref | GoogleScholarGoogle Scholar | 25780237PubMed |
Lythgoe, JN, and Partridge, JC (1989). Visual pigments and the acquisition of visual information. The Journal of Experimental Biology 146, 1–20.
| Visual pigments and the acquisition of visual information.Crossref | GoogleScholarGoogle Scholar | 2689557PubMed |
Manning, EL, Cate, HS, and Lohmann, KJ (1997). Discrimination of ocean wave features by hatchling loggerhead sea turtles, Caretta caretta. Marine Biology 127, 539–544.
| Discrimination of ocean wave features by hatchling loggerhead sea turtles, Caretta caretta.Crossref | GoogleScholarGoogle Scholar |
Marchesan, M, Spoto, M, Verginella, L, and Ferrero, EA (2005). Behavioural effects of artificial light on fish species of commercial interest. Fisheries Research 73, 171–185.
| Behavioural effects of artificial light on fish species of commercial interest.Crossref | GoogleScholarGoogle Scholar |
McFarlane, RW (1963). Disorientation of loggerhead hatchlings by artificial road lighting. Copeia 1963, 153.
| Disorientation of loggerhead hatchlings by artificial road lighting.Crossref | GoogleScholarGoogle Scholar |
Minnaar, C, Boyles, JG, Minnaar, IA, Sole, CL, and McKechnice, AE (2015). Stacking the odds: light pollution may shift the balance in an ancient predator–prey arms race. Journal of Applied Ecology 52, 522–531.
| Stacking the odds: light pollution may shift the balance in an ancient predator–prey arms race.Crossref | GoogleScholarGoogle Scholar |
Okuyama, J, Abe, O, Nishizawa, H, Kobayashhi, M, Yoseda, K, and Arai, N (2009). Ontogeny of the dispersal migration of green turtle (Chelonia mydas) hatchlings. Journal of Experimental Marine Biology and Ecology 379, 43–50.
| Ontogeny of the dispersal migration of green turtle (Chelonia mydas) hatchlings.Crossref | GoogleScholarGoogle Scholar |
Osovsky, N, and Shettleworth, SJ (1968). Wavelength preferences and brightness cues in the water finding behaviour of sea turtles. Behaviour 32, 211–257.
| Wavelength preferences and brightness cues in the water finding behaviour of sea turtles.Crossref | GoogleScholarGoogle Scholar |
Pawson, SM, and Bader, MK-F (2014). LED lighting increases the ecological impact of light pollution irrespective of colour temperature. Ecological Applications 24, 1561–1568.
| LED lighting increases the ecological impact of light pollution irrespective of colour temperature.Crossref | GoogleScholarGoogle Scholar | 29210222PubMed |
Pendoley K (2005) Sea turtles and the environmental management of industrial activities in north west Western Australia. Ph.D thesis, Murdoch University, Perth, WA, Australia.
Pendoley, K, and Kamrowski, RL (2015). Influence of horizon elevation on the sea-finding behaviour of hatchling flatback turtles exposed to artificial light glow. Marine Ecology Progress Series 529, 279–288.
| Influence of horizon elevation on the sea-finding behaviour of hatchling flatback turtles exposed to artificial light glow.Crossref | GoogleScholarGoogle Scholar |
Pendoley, KL, Bell, CD, McCracken, R, Ball, KR, Sherborne, J, Oates, JE, Becker, P, Vitenbergs, A, and Whittock, PA (2014). Reproductive biology of the flatback turtle Natator depressus in Western Australia. Endangered Species Research 23, 115–123.
| Reproductive biology of the flatback turtle Natator depressus in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Philibosian, R (1976). Disorientation of hawskbill turtle hatchlings, Eretmochelys imbricata, by stadium lights. Copeia 1976, 824.
| Disorientation of hawskbill turtle hatchlings, Eretmochelys imbricata, by stadium lights.Crossref | GoogleScholarGoogle Scholar |
Pilcher NJ, Enderby S, Stringell T, Bateman L (2000) Nearshore turtle hatchling distribution and predation. In ‘Sea turtles of the Indo-Pacific: research management and conservation. Proceedings of the Second ASEAN Symposium and Workshop on Sea Turtle Biology and Conservation’. (Eds N Pilcher, G Ismail) pp. 151–166. (ASEAN Academic Press: London, UK)
Raap, T, Pinxten, R, and Eens, M (2015). Light pollution disrupts sleep in free-living animals. In Nature. Scientific Reports 5, 13557.
| Light pollution disrupts sleep in free-living animals. In Nature.Crossref | GoogleScholarGoogle Scholar | 26337732PubMed |
Reising, M, Salmon, M, and Stapleton, S (2015). Hawksbill nest site selection affects hatchling survival at a rookery in Antigua, West Indies. Endangered Species Research 29, 179–187.
| Hawksbill nest site selection affects hatchling survival at a rookery in Antigua, West Indies.Crossref | GoogleScholarGoogle Scholar |
Robertson, K, Booth, DT, and Limpus, CJ (2016). An assessment of ‘turtle-friendly’ lights on the sea-finding behaviour of loggerhead turtle hatchlings (Caretta caretta). Wildlife Research 43, 27–37.
| An assessment of ‘turtle-friendly’ lights on the sea-finding behaviour of loggerhead turtle hatchlings (Caretta caretta).Crossref | GoogleScholarGoogle Scholar |
Rodríguez, A, Holmes, ND, Ryan, PG, Wilson, K, Faulquier, L, Murillo, Y, Raine, AF, Penniman, JF, Neves, V, Rodrıguez, B, Negro, JJ, Chiaradia, A, Dann, P, Anderson, T, Metzger, B, Shirai, M, Deppe, L, Wheeler, J, Hodum, P, Gouveia, C, Carmo, V, Carreira, GP, Delgado-Alburqueque, L, Guerra-Correa, C, Couzi, F, Travers, M, and Le Corre, M (2017). Seabird mortality induced by land‐based artificial lights. Conservation Biology 31, 986–1001.
| Seabird mortality induced by land‐based artificial lights.Crossref | GoogleScholarGoogle Scholar | 28151557PubMed |
Russart, KLG, and Nelson, RJ (2018). Artificial light at night alters behavior in laboratory and wild animals. Journal of Experimental Zoology. Part A, Ecological and Integrative Physiology 329, 401–408.
| Artificial light at night alters behavior in laboratory and wild animals.Crossref | GoogleScholarGoogle Scholar | 29806740PubMed |
Salmon, M (2003). Artificial night lighting and sea turtles. Biologist 50, 163–168.
Salmon M (2005) Protecting sea turtles from artificial night lighting at Florida’s oceanic beaches. In ‘Ecological Consequences of Artificial Night Lighting’. (Eds C Rich, T Longcore) pp. 141–168. (Island Press: Washington, DC, USA)
Salmon, M, and Wyneken, J (1994). Orientation by hatchling sea turtles: mechanisms and implications. Herpetological Natural History 2, 13–24.
Salmon, M, Wyneken, J, Fritz, E, and Lucas, M (1992). Seafinding by hatchling sea turtles: role of brightness, silhouette and beach slope as orientation cues. Behaviour 122, 56–77.
| Seafinding by hatchling sea turtles: role of brightness, silhouette and beach slope as orientation cues.Crossref | GoogleScholarGoogle Scholar |
Sella, KN, Salmon, M, and Witherington, BE (2006). Filtered streetlights attract hatchling marine turtles. Chelonian Conservation and Biology 5, 255–261.
| Filtered streetlights attract hatchling marine turtles.Crossref | GoogleScholarGoogle Scholar |
Small, C, and Nicholls, RJ (2003). A global analysis of human settlement in coastal zones. Journal of Coastal Research 19, 584–599.
Stewart, KR, and Wyneken, J (2004). Predation risk to loggerhead hatchlings at a high-density nesting beach in Southeast Florida. Bulletin of Marine Science 74, 325–335.
Thums, M, Whiting, SD, Reisser, J, Pendoley, KL, Pattiaratchi, CB, Proietti, M, Hetzel, Y, Fisher, R, and Meekan, MG (2016). Artificial light on water attracts turtle hatchlings during their near shore transit. Royal Society Open Science 3, 160142.
| Artificial light on water attracts turtle hatchlings during their near shore transit.Crossref | GoogleScholarGoogle Scholar | 27293795PubMed |
Verheijen, FJ (1958). The mechanisms of the trapping effect of artificial light sources upon animals. Netherlands Journal of Zoology 13, 1–107.
Verheijen, FJ (1985). Photopollution: artificial light optic spatial control systems fail to cope with. Indicents, caustations, remedies. Experimental Biology 44, 1–18.
| 3896840PubMed |
Verheijen, FJ, and Wildschut, JT (1973). The photic orientation of hatchling sea turtles during water finding behaviour. Netherlands Journal of Sea Research 7, 53–67.
| The photic orientation of hatchling sea turtles during water finding behaviour.Crossref | GoogleScholarGoogle Scholar |
Whelan, CL, and Wyneken, J (2007). Estimating predation levels and site-specific survival of hatchling loggerhead seaturtles (Caretta caretta) from south Florida beaches. Copeia 2007, 745–754.
| Estimating predation levels and site-specific survival of hatchling loggerhead seaturtles (Caretta caretta) from south Florida beaches.Crossref | GoogleScholarGoogle Scholar |
Wilson, P, Thums, M, Pattiaratchi, C, Meekan, MG, Pendoley, K, Fisher, R, and Whiting, S (2018). Artificial light disrupts the nearshore dispersal of neonate flatback turtles (Natator depressus). Marine Ecology Progress Series 600, 179–192.
| Artificial light disrupts the nearshore dispersal of neonate flatback turtles (Natator depressus).Crossref | GoogleScholarGoogle Scholar |
Wilson, P, Thums, M, Pattiaratchi, C, Whiting, S, Pendoley, K, Ferreira, LC, and Meekan, M (2019). High predation of marine turtle hatchlings near a coastal jetty. Biological Conservation 236, 571–579.
| High predation of marine turtle hatchlings near a coastal jetty.Crossref | GoogleScholarGoogle Scholar |
Wilson, P, Thums, M, Pattiaratchi, C, Whiting, S, Meekan, M, and Pendoley, K (2020). Nearshore wave characteristics as cues for swimming orientation in flatback turtle hatchlings. Journal of Experimental Marine Biology and Ecology 535, 1–7.
Witherington BE (1992) ‘Sea-finding behavior and the use of photic orientation cues by hatchling sea turtles.’ (University of Florida: Gainesville, FL, USA)
Witherington, BE, and Bjorndal, KA (1991a). Influences of artificial lighting on the seaward orientation of hatchling loggerhead turtles Caretta caretta. Biological Conservation 55, 139–149.
| Influences of artificial lighting on the seaward orientation of hatchling loggerhead turtles Caretta caretta.Crossref | GoogleScholarGoogle Scholar |
Witherington, BE, and Bjorndal, KA (1991b). Influences of wavelength and intensity on hatchling sea-turtle phototaxis: implications for sea-finding behavior. Copeia 1991, 1060–1069.
| Influences of wavelength and intensity on hatchling sea-turtle phototaxis: implications for sea-finding behavior.Crossref | GoogleScholarGoogle Scholar |
Witherington, BE, and Salmon, M (1992). Predation on loggerhead turtle hatchlings after entering the sea. Journal of Herpetology 26, 226–228.
| Predation on loggerhead turtle hatchlings after entering the sea.Crossref | GoogleScholarGoogle Scholar |
Witherington BE, Martin RE (2003) Understanding, assessing, and resolving light-pollution problems on sea turtle nesting beaches. Florida Fish and Wildlife Conservation Commission, Technical Report TR-2, Florida Marine Research Institute, Saint Petersburg, FL, USA.
Wood SN (2017) ‘Generalized additive models: an introduction with R’, 2nd edn. (CRC Press: Boca Raton, FL, USA)
Zapata, MJ, Sullivan, SMP, and Gray, SM (2019). Artificial lighting at night in estuaries—implications from individuals to ecosystems. Estuaries and Coasts 42, 309–330.
| Artificial lighting at night in estuaries—implications from individuals to ecosystems.Crossref | GoogleScholarGoogle Scholar |