

At-vessel mortality of tawny nurse sharks (Nebrius ferrugineus) caught in the Queensland Shark Control Program
Matthew J. Campbell A * , Tracey B. Scott-Holland B , Samuel J. Fary C and Matthew N. McMillan A
A * , Tracey B. Scott-Holland B , Samuel J. Fary C and Matthew N. McMillan A
A
B
C
Abstract
The Queensland Shark Control Program (QSCP) targets potentially dangerous sharks, using a combination of large-mesh nets and baited drumlines at beaches in Queensland, Australia. The tawny nurse shark (Nebrius ferrugineus) is caught incidentally by the QSCP, and shark control programs have been identified as a moderate threat to the species in Australia.
Assessing at-vessel mortality (AVM) is an important first step in determining the actual impact of the QSCP on the N. ferrugineus population.
Data collected by QSCP contractors were used to determine factors affecting AVM of N. ferrugineus and data were analysed using generalised linear mixed modelling.
Mean AVM was 5.95 and 30.1% for those animals caught on drumlines and in nets respectively. At-vessel mortality was found to increase with total length for net-caught N. ferrugineus. Sea-surface temperature and sex had no effect on AVM.
Animals caught on drumlines are more likely to survive capture than are those caught in nets.
The reduction in the number of nets deployed, and the introduction of alternative methods, such as catch alert (or SMART) drumlines and drone surveillance of beaches, is likely to have reduced the impact of the QSCP on the N. ferrugineus population.
Keywords: at-vessel mortality, AVM, baited drumlines, chondrichthyans, Great Barrier Reef Marine Park, Nebrius ferrugineus, QSCP, Queensland Shark Control Program, respiratory mode, shark nets.
Introduction
Chondrichthyans (sharks, rays and chimaeras) have been the subject of increasing research efforts since the 1980s (Shiffman et al. 2020), owing to a lack of data required to assess populations and inform management (Simpfendorfer et al. 2011). Of the 1043 chondrichthyans for which sufficient data are available to assess the risk of extinction, 391 (~37%) are categorised globally as critically endangered, endangered or vulnerable to extinction, primarily as a result of incidental capture in fisheries targeting other species (Dulvy et al. 2021). Further, a recent study indicated that overfishing has halved the populations of 1119 sharks and rays since 1970 (Dulvy et al. 2024). Life-history strategies including slow growth, delayed maturity, low fecundity and long life spans make many chondrichthyan species vulnerable to over-exploitation (Ellis et al. 2008) and inhibit recovery of depleted populations (Simpfendorfer et al. 2011).
One species that faces a high risk of extinction is the tawny nurse shark (Orectolobiformes; Ginglymostomidae, Nebrius ferrugineus Lesson, 1830), which is listed as ‘vulnerable’ according to the International Union for Conservation of Nature (IUCN) Red List of Threatened Species (Simpfendorfer et al. 2021). Nebrius ferrugineus occurs in tropical Indo-Pacific waters, from eastern Africa to Tahiti, where it is exploited for its meat, liver, oil, fins and hide (Last and Stevens 2009). This, combined with habitat loss throughout its range, has resulted in a 30–49% reduction in the global N. ferrugineus population (Simpfendorfer et al. 2021). However, Australia provides a refuge for N. ferrugineus, and other threatened, globally distributed coastal and shelf chondrichthyans (Kyne et al. 2021), primarily as a result of well-managed fisheries (Dulvy et al. 2024).
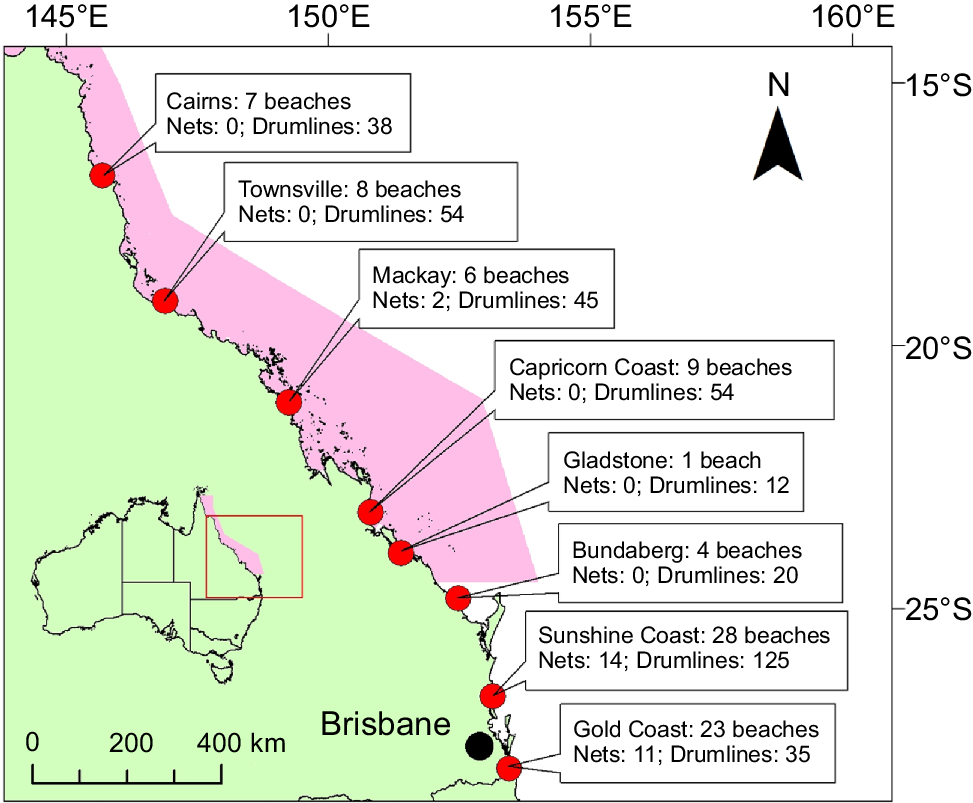
In Australia, N. ferrugineus is classified as ‘least concern’ across its distribution (Kyne et al. 2021). Although incidentally caught in two large penaeid-trawl fisheries, legislation requiring the use of turtle excluder devices (TEDs), introduced in the early 2000s, has reduced the capture of N. ferrugineus by 100% (Kyne et al. 2021). Apart from these fisheries, no N. ferrugineus have been landed by the Queensland commercial net fishery (source: https://qfish.fisheries.qld.gov.au/), whereas the recreational catch of N. ferrugineus is unknown (source: https://www.daf.qld.gov.au/business-priorities/fisheries/monitor/rec/statewide-survey#view-the-data), but is not likely to affect the Queensland population significantly (Kyne et al. 2021). As such, the only source of possibly significant fishing mortality affecting the N. ferrugineus population in Queensland waters is the Queensland Shark Control Program (QSCP). The QSCP is a bather protection program implemented in the early 1960s after several fatal shark bites (Sumpton et al. 2011). Potentially dangerous sharks, primarily tiger sharks (Galeocerdo cuvier Péron & Lesueur, 1822), bull sharks (Carcharhinus leucas Valenciennes), and white sharks (Carcharodon carcharias Linnaeus, 1758), are targeted using a combination of large-mesh nets and baited drumlines at beaches along the Queensland coastline (Sumpton et al. 2011). At the program’s inception, 76 baited drumlines and 30 nets were deployed across 29 beaches in the following 5 broad regions: Cairns, Townsville, Mackay, the Sunshine Coast and the Gold Coast. In 2024, 383 drumlines and 27 nets were deployed across the state (Fig. 1).
Location of each Queensland Shark Control Program (QSCP) region, outlining the number of beaches within each region and the number of nets and drumlines deployed in 2024. The pink-shaded area represents the extent of the World Heritage-listed Great Barrier Reef Marine Park.
Because of the concerns relating to interactions with non-target animals, the use of nets by the QSCP within the boundaries of the World Heritage-listed Great Barrier Reef Marine Park (GBRMP) was progressively reduced until 2017, when all nets had been replaced by baited drumlines. In 2020, the Great Barrier Reef Marine Park Authority, the regulatory body responsible for managing activities within the GBRMP, limited the use of baited drumlines, such that (1) the equipment is serviced every 24 h where possible, (2) all animals caught by drumlines are released alive when safe to do so, and (3) G. cuvier, C. leucas and C. carcharias are tagged and relocated offshore. These conditions have been implemented by the QSCP since 17 February 2020. Prior to this time, contractors were required to service QSCP equipment at least every 3 days, weather permitting, and any target species caught were euthanased. Servicing was also undertaken on weekends, during school holidays and on public holidays, when beach use was highest.
The impact of the QSCP on the N. ferrugineus population in Queensland is currently unknown. Kyne et al. (2021) identified shark control programs as a moderate threat to N. ferrugineus in Australia; however, all N. ferrugineus must be released alive if possible after capture by QSCP gear. As such, an important first step in assessing the threat posed to the Queensland population of N. ferrugineus by the QSCP is to quantify mortality resulting from interactions with QSCP gear. Post-release mortality (PRM) is the proportion of individuals that die during, or as a result of, catch and release and accounts for any immediate or delayed effects of physical or physiological trauma experienced by an individual (Ellis et al. 2017). Although quantifying PRM is ideal, the cost and logistical constraints of experimentally deriving PRM estimates, using cage or tank experiments or satellite tagging, can be prohibitive (Musyl and Gilman 2019).
Owing to these limitations, researchers assess at-vessel mortality (AVM), which is defined as the proportion of individuals that are dead when brought on-board (or alongside) a fishing vessel (Ellis et al. 2017). Quantifying AVM improves the accuracy of fishing mortality estimates used to assess chondrichthyan stocks (Dapp et al. 2016; Matias Braccini and Waltrick 2019), and can be used to inform the management and conservation of chondrichthyans (Sulikowski et al. 2020; Gilman et al. 2022). Previous studies have shown that AVM is species-specific (Rodríguez-Cabello and Sánchez 2017; Matias Braccini and Waltrick 2019; Mucientes et al. 2022) and is influenced by factors such as respiratory mode and gear type (Dapp et al. 2016), fish size (Carruthers et al. 2009), soak time (Lyons et al. 2013; Matias Braccini and Waltrick 2019; Gulak and Carlson 2021), hooking time (Morgan and Carlson 2010; Butcher et al. 2015) and water temperature (Massey et al. 2022).
The objective of the current study was to quantify the AVM of N. ferrugineus caught in nets and on drumlines deployed by the QSCP to help inform the ecological risk posed to the species by the program.
Materials and methods
Fishing equipment and servicing
Sumpton et al. (2011) provided a description of the gear used throughout the QSCP (see https://www.daf.qld.gov.au/news-media/campaigns/sharksmart/equipment for illustrations of the gear used). In summary, the nets used are constructed of 1.6 mm diameter polypropylene twine with a mesh size of 50 cm. The nets have a drop of 12 meshes and are between 124 and 186 m long, depending on location. Generally, drumlines consist of a single 14/0 J-shaped shark hook baited with a single fresh sea mullet (Mugil cephalus Linnaeus, 1758) or shark flesh (Charcharhinus spp.) suspended 2 m below a buoy and at least 2 m from the seafloor at low tide. Suspending baits above the seafloor reduces incidental captures of benthic non-target elasmobranch species, while not affecting catch rates of potentially dangerous sharks (Lipscombe et al. 2023). Drumlines are anchored to the substrate using a length of 12-mm polypropylene rope three to four times the depth of the water. Drumlines and nets are anchored to the seafloor ~600 m from, and parallel to, the shore, in water depths between 6 and 12 m.
From the beginning of the program, in 1962, until 17 February 2020, QSCP equipment was deployed year-round, and serviced every 1–3 days, depending on weather. Servicing was undertaken during the day, mostly during the morning, and the time taken to service the gear was dependent on the number of nets or drumlines deployed. At the start of each service day, contractors recorded abiotic data such as sea-surface temperature (SST, °C). During the service, any live target sharks were euthanised and discarded ~1 km seaward of the capture location, whereas any live, non-target species, including N. ferrugineus, were released at the capture location as soon as practicable. Contractors recorded relevant data such as species, size (m, total length, TL, for sharks and Rhinopristiformes, disc width for remaining batoids), sex and alive or dead (AVM). Animals were considered dead if no body, eye or gill movement was detected or if rigor mortis was obvious. During each service, drumlines were rebaited. When required, drumlines and nets were recovered for cleaning and maintenance, and immediately replaced. During periods of forecast severe weather (e.g. cyclone), some equipment was temporarily removed from the water.
In the period between 18 February 2020 and 30 April 2024, service frequency of drumlines located within the GBRMP increased to daily, with a minimum of 21 services undertaken per month. Baits are removed and hooks are attached to the buoy to render them ineffective when not in use such as during significant weather events that would prevent scheduled servicing. No nets were in use within the bounds of the GBRMP during this period. All animals were released alive where possible, and any G. cuvier, C. leucas and C. carcharias inividuals were tagged with external acoustic tags or dart tags before being released approximately 1 km seaward of the capture location. Outside of the GBRMP, servicing continued as per the period prior to 17 February 2020; however, the contractors operating in the Cairns and Mackay regions serviced the gear inside the GBRMP 260 days year−1 and 182 days year−1 outside the GBRMP (Fig. 1), whereas contractors operating in the Capricorn Coast and Townsville regions serviced all equipment 260 days year−1.
The trial was conducted under Marine Park Permit number G17/33288.1 and Queensland Department of Agriculture and Fisheries Animal Ethics approval CA 2021/03/1482.
Statistical analysis
Data were obtained from the QSCP catch database. All catch records between 1 January 1995 and 30 April 2024 were examined and the dataset was restricted to those cases where a N. ferrugineus was caught, and data on size, sex, SST, and AVM were recorded. A trial of catch alert drumlines (CADs, aka SMART drumlines) commenced in the Capricorn Coast region in 2021, before the trial was extended to the Cairns and Mackay regions in February 2024. Catch alert drumlines are similar to the drumlines used throughout the QSCP, but include a satellite communication unit that sends an alert to the contractor when an animal is hooked. This allows contractors to immediately attend to the hooked animal, facilitating its tagging, relocation and release. Any N. ferrugineus caught using CADs were excluded from the analyses to avoid bias when estimating AVM.
At-vessel mortality was assessed using generalised linear mixed models (GLMM). At-vessel mortality was the response variable, modelled as a binomial distribution where 0 = alive and 1 = dead. Beach was added as a random term, whereas gear type (net, drumline), month (January to December), and sex (male, female) were added as categorical factors to assess their effect on the probability of survival. A dummy binary variable (0 = before, 1 = after), representing the change to daily servicing within the GBRMP, was also added to determine the effect of this change. Further, size and SST were added as continuous covariates. Categorical terms and covariates were tested for significance and retained in the model only if a significant effect was detected and their addition reduced the Akaike information criteria (AIC). Relevant two-way interactions were also tested and excluded if their addition had no significant effect on AVM. The AVM was estimated using R statistical software (ver. 4.0.2, R Foundation for Statistical Computing, Vienna, Austria, see https://www.R-project.org/, accessed 6 November 2024) using the ‘glmer’ function within the lme4 package (ver. 1.1-23, see https://CRAN.R-project.org/package=lme4; Bates et al. 2015). The ‘bootMER’ function within the lme4 package was used to generate 95% confidence intervals around the estimated probabilities.
Results
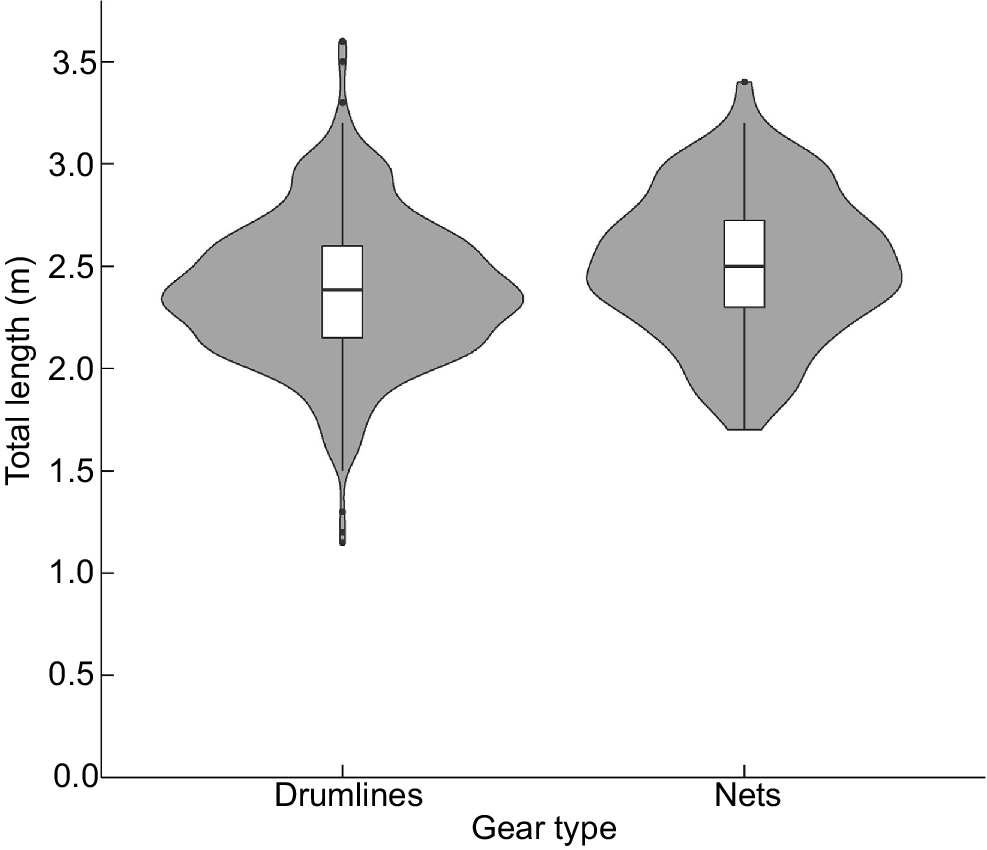
In total, 772 N. ferrugineus individuals were caught by QSCP contractors in the period between 1 January 1995 and 30 April 2024. Sea-surface temperature was absent in 47 cases, one total length was not recorded, and sex was indeterminable for 17 animals. Consequently, AVM was estimated from the capture of 707 individuals (Supplementary Tables S1 and S2), 428 (60%) of which were female. The total length of females (mean = 2.38 m, s.d. = 0.37 m) and males (mean = 2.41 m, s.d. = 0.37 m) did not differ significantly (t = −1.03, d.f. = 631, P = 0.30). However, the 92 N. ferrugineus individuals caught in nets (mean = 2.51 m, s.d. = 0.35 m) were significantly larger (t = 3.23, d.f. = 121, P < 0.01) than the 615 individuals caught on drumlines (mean = 2.38 m, s.d. = 0.36 m) (Fig. 2).
Violin plot of length, as a function of gear type, for 707 tawny nurse sharks (Nebrius ferrugineus) caught by the QSCP between 1 January 1995 and 30 April 2024. Also shown is a box and whiskers plot for length as a function of gear type.
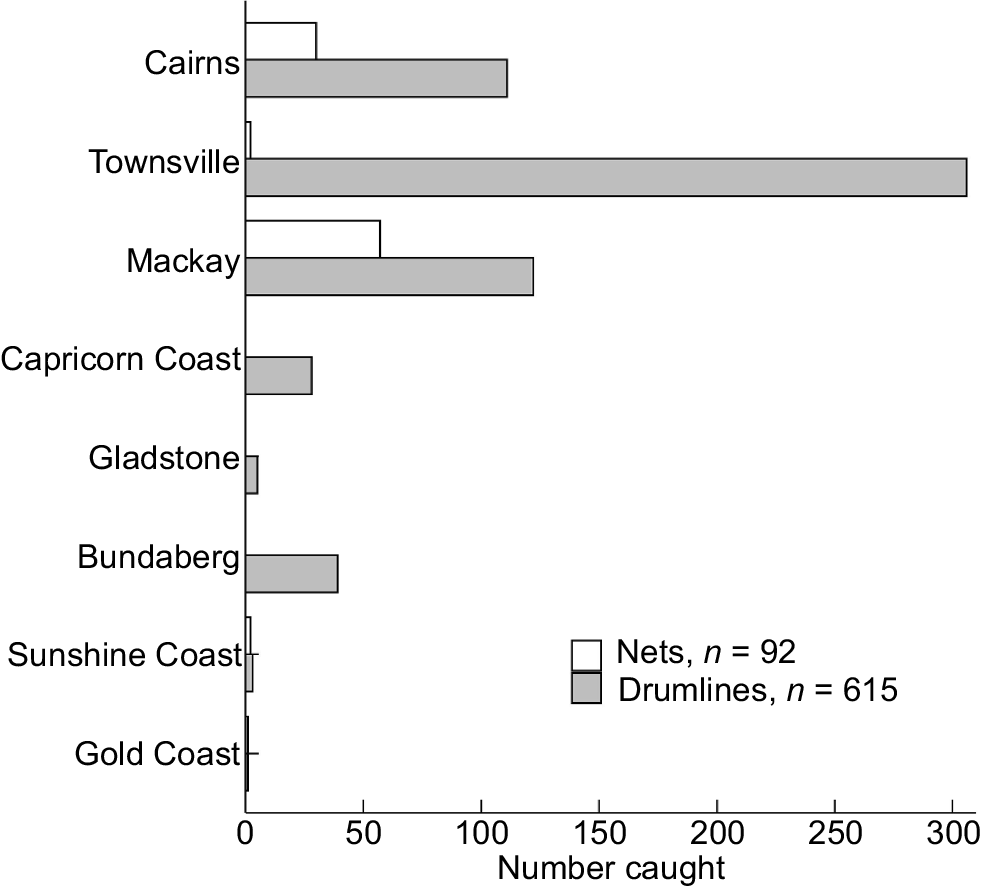
Nebrius ferrugineus was more common in northern regions (Fig. 3, Tables S1, S2). In total, 306 N. ferrugineus sharks were caught in the Townsville region, only two of which were caught in nets (Table S1). Of the 141 N. ferrugineus individuals caught in the Cairns region, 30 (~21%) were caught in nets, as were 57 of the 179 idndividuals (32%) caught in the Mackay region. Only drumlines were deployed in the Capricorn Coast, Gladstone and Bundaberg regions during the study period.
Number of tawny nurse sharks (Nebrius ferrugineus) caught in each QSCP region between 1 January 1995 and 30 April 2024, sexes combined, as a function of gear type. Note: nets were not deployed at any beach within the Capricorn Coast, Gladstone and Bundaberg regions during the study period.
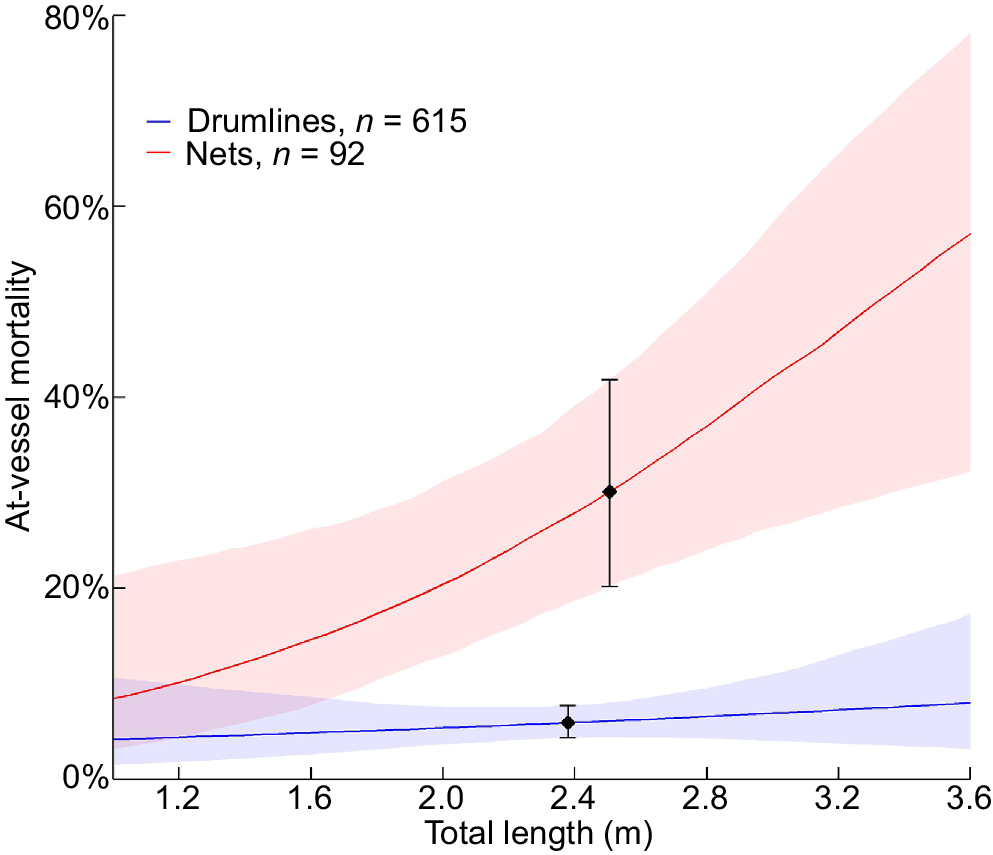
Of the 92 N. ferrugineus individuals caught in nets, 32 (~35%) were dead when caught, compared with 42 of the 615 (~7%) caught on drumlines. The GLMM indicated that the increasing size of N. ferrugineus caught in nets resulted in higher AVM (net × TL: β = 0.957, s.e. = 0.367, P = 0.009). Sex (P = 0.386), SST (P = 0.586), month (P = 0.199) and the dummy variable representing the change to daily servicing within the GBRMP (P = 0.644) had no significant effect on survival. Similarly, the drumline × TL interaction term had no effect on AVM (P = 0.556). The AVM of N. ferrugineus caught on drumlines was 5.95% (CIα = 0.05 = 4.38–7.76%) and 30.1% (CIα = 0.05 = 20.18–41.84%) for those caught in nets at the mean TL of animals caught by the respective gear types (2.38 and 2.51 m respectively) (Fig. 4).
At-vessel mortality (AVM) of tawny nurse sharks (Nebrius ferrugineus) caught in nets and on drumlines between 1 January 1995 and 30 April 2024, as a function of total length. The black points are the estimated AVM of Nebrius ferrugineus at the mean total length of the individuals caught on the respective gear types (drumlins = 2.38 m; nets = 2.51 m).
Discussion
These results represent the first AVM estimates for N. ferrugineus in the primary literature. Generally, N. ferrugineus was resilient to capture on baited drumlines deployed as part of the QSCP. However, AVM was higher for animals caught in nets, particularly for larger individuals. This result is consistent with that of Sumpton et al. (2011), who demonstrated that the AVM of chondrichthyans caught in the QSCP was highest when caught in nets. Similarly, Dapp et al. (2016) found that the AVM of elasmobranchs was higher when caught in gillnets, than when caught on longlines. The higher AVM for net-caught N. ferrugineus in the current study is likely to be related to heavy entanglement around the gill region, such as that reported for the gummy shark (Mustelus antarcticus) (Frick et al. 2010), which restricts water flow through the gills, even for species that can buccal pump to irrigate the gills. Further, the correlation between AVM and the total length of net-caught N. ferrugineus observed in the current study is consistent with the results of Braccini et al. (2012), who reported that larger chondrichthyans were less likely to survive capture in bottom-set gillnets in southern Australia, than are smaller conspecifics. Restricted water flow through the gills as a result of heavy entanglement, combined with the higher oxygen consumption (Sims 1996) and proportionally less oxygen uptake (Bouyoucos et al. 2019) in larger individuals, may explain this result.
In contrast, individuals caught on hooks are able to swim while tethered by the gear (Frick et al. 2010), allowing respiration while hooked. As a result, chondrichthyans caught using baited hooks tend to exhibit lower AVM than those caught in gillnets (Dapp et al. 2016; Ellis et al. 2017). After the initial acute stress response on hooking, the ability to swim facilitates some physiological recovery (Brooks et al. 2012), which may reduce AVM and PRM. In the current study, J-hooks were used throughout the QSCP; however, trials of circle hooks are underway to determine their effect on catch and AVM. Numerous studies have shown that circle hooks reduce AVM or PRM (e.g. Godin et al. 2012; Reinhardt et al. 2018; Gilman et al. 2022), primarily by reducing the incidence of deep hooking (Gilman et al. 2016). Further, circle hooks have been shown to reduce the capture (Yan et al. 2025) and deep hooking (Santos et al. 2024) of sea turtles, a group occasionally caught incidentally in the QSCP (Gribble et al. 1998; Sumpton et al. 2011). Replacing J-hooks with circle hooks in the QSCP will likely further reduce AVM for N. ferrugineus, and outputs from the trial of circle hooks currently underway will inform future management of the QSCP.
The low AVM of hook-caught N. ferrugineus is consistent with that reported for the confamilial Atlantic nurse shark (Ginglymostoma cirratum Bonnaterre). Whitney et al. (2021) and Morgan et al. (2009) reported 0% AVM for G. cirratum caught on bottom-set longlines used in separate US fisheries. Both N. ferrugineus and G. cirratum can respire while stationary, by buccal pumping, a method of respiration that contributes to lower AVM, than for obligate ram ventilators (Dapp et al. 2016). Clearly, constraining the movement of obligate ram ventilators will reduce the volume of water moving through the gills, whereas species capable of buccal pumping can continue to respire despite being hooked. For example, Whitney et al. (2021) reported that the AVM of blacktip, blacknose, and spinner sharks was 35, 35 and 62% respectively, all of which are obligate ram ventilators. Similarly, Sumpton et al. (2011) reported 66.3–80% AVM for obligate ram ventilating carcharhinids caught in QSCP nets, compared with 10.5–19.7% AVM for batoids, which respire while stationery, through spiracles.
Apart from the estimates published by Whitney et al. (2021) and Morgan et al. (2009) for G. cirratum, very little AVM information is available for any of the Orectolobiformes (Ellis et al. 2017). Sumpton et al. (2011) observed that 10 of the 49 zebra sharks (Stegostoma tigrinum Forster, 1781) caught in QSCP nets died during their study (AVM = 20.4%), whereas the only individual caught on a drumline survived. Both Walker et al. (2005) and Braccini et al. (2012) reported 0% AVM for gillnet-caught spotted wobbegongs (Orectolobus maculatus) caught in a southern Australian gillnet fishery, although sample size was low (n = 4 and 5 respectively). Additionally, Braccini et al. (2012) reported an AVM of 12.5% for the rusty carpetshark (Parascyllium ferrugineum McCulloch, 1911, n = 24) and 20% for the varied carpetshark (Parascyllium variolatum Duméril, 1853, n = 4). Matias Braccini and Waltrick (2019) assessed the AVM of chondrichthyans caught by demersal longlines in Western Australia and found that the AVM of S. tigrinum (n = 8), and wobbegongs (Orectolobus ornatus de Vis, 1883 and Orectolobus spp., n = 3) was 0%. The results of this study, as well as those from data-limited prior research, indicate that species of Orectolobiformes generally exhibit low AVM. However, further research is required to determine survival in the longer term, given the potential for delayed effects of physical trauma or physiological stress (Skomal 2007).
It should be noted that the soak times for the drumlines or nets deployed during the current study was higher than those in the studies conducted by Whitney et al. (2021), Morgan et al. (2009), Walker et al. (2005) and Braccini et al. (2012). Whereas soak times in the current study were 1–3 days, the maximum soak time among the compared studies was ~20 h in the Braccini et al. (2012) study. This author found that AVM and soak time were inversely correlated and, as a result, it is difficult to compare results from the current study with those from previous studies. Further work is required to determine the effect of implementing daily servicing within the GBRMP; however, reducing the soak time of QSCP gear is likely to result in lower AVM than that derived in the current study.
The nets used throughout the QSCP caught larger N. ferrugineus than did baited drumlines. This is consistent with previous studies of chondrichthyans caught by the QSCP (e.g. Simpfendorfer 1992; Holmes et al. 2012; Haig et al. 2018). For example, Haig et al. (2018) found that C. leucas individuals caught on drumlines were, on average, 0.4 m smaller than those caught in nets. Therefore, it is reasonable to assume that the removal of nets from the GBRMP is likely to have resulted in a decrease in mean length at capture over time for species such as C. leucas, G. cuvier and N. ferrugineus, particularly when comparing current sizes to those from periods when more nets were deployed (c. pre-1990). A decline in the size at capture of sharks over time has been linked to overfishing for species interacting with the QSCP (Haig et al. 2018) and other fisheries (e.g. Braccini 2017). However, ignoring the effect of gear type, and other operational factors such as hook size, bait type and trace length, on size and species composition of individuals caught in the QSCP over a long period can lead to spurious conclusions and ill-informed recommendations.
The community has expressed concerns about the impact of the QSCP on marine fauna, particularly non-target animals such as whales, dolphins and dugongs, in QSCP nets (Gribble et al. 1998). Similarly, recent research inferring the QSCP may be responsible for declining shark populations (Roff et al. 2018), or declines in the functional diversity of sharks (Henderson et al. 2024), has been cited as evidence for the removal of nets and drumlines in Queensland. The QSCP is trialling alternatives to nets and traditional drumlines to address community concerns. Trials are currently underway to determine the efficacy of CADs (Campbell and Scott-Holland 2023) and circle hooks on the catch and survival of target and non-target animals. Further, unmanned aerial vehicles (UAVs or drones) are being trialled as a method of identifying potentially dangerous sharks (Mitchell et al. 2022), enabling lifeguards to evacuate beaches to reduce risk to swimmers.
The implementation of these measures is likely to reduce the impact of the QSCP on the Queensland population of N. ferrugineus. The current study has shown that the AVM of N. ferrugineus caught on drumlines is low, and preliminary data indicate that AVM is 0% when caught on CADs (M. Campbell, unpubl. data). As such, the QSCP is likely to pose low ecological risk to the Queensland N. ferrugineus population. However, an understanding of the actual ecological impact of the QSCP requires further research into PRM and population dynamics. Future studies should focus on (1) quantifying long-term catch and release using survivorship pop-up archival transmitting (sPAT) tags or acoustic telemetry utilising Queensland’s expanding acoustic array (Barnett et al. 2024), and (2) an ecological risk assessment to determine the effect of current levels of fishing mortality on the long-term sustainability of the N. ferrugineus population in Queensland.
Authorship contribution
Matthew Campbell provided the conceptualisation, formal analysis, investigation, methodology, validation, visualisation, writing of the original draft, and review and editing. Tracey Scott-Holland contributed data curation, investigation, writing of the original draft, and review and editing. Samuel Fary undertook investigation, writing of the original draft, and review and editing. Matthew McMillan contributed to the writing of the original draft, and review and editing.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Declaration of funding
All authors were funded by the Queensland Department of Primary Industries for this research.
Acknowledgements
The authors thank the QSCP contractors for their continued diligence in reporting catch and survival data. Glynis Fowler collated and managed the QSCP data, and we thank her for help throughout this study. Michael Mikitis and Natale Snape provided helpful comments on an earlier draft of this paper. Special thanks go to Helen McPherson for her assistance in sourcing and supplying relevant literature.
References
Barnett A, Jaine FRA, Bierwagen SL, Lubitz N, Abrantes K, Heupel MR, Harcourt R, Huveneers C, Dwyer RG, Udyawer V, Simpfendorfer CA, Miller IB, Scott-Holland T, Kilpatrick CS, Williams SM, Smith D, Dudgeon CL, Hoey AS, Fitzpatrick R, Osborne FE, Smoothey AF, Butcher PA, Sheaves M, Fisher EE, Svaikauskas M, Ellis M, Kanno S, Cresswell BJ, Flint N, Armstrong AO, Townsend KA, Mitchell JD, Campbell M, Peddemors VM, Gustafson JA, Currey-Randall LM (2024) From little things big things grow: enhancement of an acoustic telemetry network to monitor broad-scale movements of marine species along Australia’s east coast. Movement Ecology 12(1), 31.
| Crossref | Google Scholar |
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67(1), 1-48.
| Crossref | Google Scholar |
Bouyoucos IA, Simpfendorfer CA, Rummer JL (2019) Estimating oxygen uptake rates to understand stress in sharks and rays. Reviews in Fish Biology and Fisheries 29(2), 297-311.
| Crossref | Google Scholar |
Braccini M (2017) Temporal patterns in the size of the main commercial shark species of Western Australia. Marine and Freshwater Research 68(6), 1112-1117.
| Crossref | Google Scholar |
Braccini M, Van Rijn J, Frick L (2012) High post-capture survival for sharks, rays and chimaeras discarded in the main shark fishery of Australia? PLoS ONE 7(2), e32547.
| Crossref | Google Scholar |
Brooks EJ, Mandelman JW, Sloman KA, Liss S, Danylchuk AJ, Cooke SJ, Skomal GB, Philipp DP, Sims DW, Suski CD (2012) The physiological response of the Caribbean reef shark (Carcharhinus perezi) to longline capture. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 162(2), 94-100.
| Crossref | Google Scholar |
Butcher PA, Peddemors VM, Mandelman JW, McGrath SP, Cullis BR (2015) At-vessel mortality and blood biochemical status of elasmobranchs caught in an Australian commercial longline fishery. Global Ecology and Conservation 3, 878-889.
| Crossref | Google Scholar |
Campbell M, Scott-Holland T (2023) Queensland shark control program: catch alert drumline trial 2022–2023. (Queensland Department of Agriculture and Fisheries: Brisbane, Qld, Australia) Available at https://era.daf.qld.gov.au/id/eprint/14175/
Carruthers EH, Schneider DC, Neilson JD (2009) Estimating the odds of survival and identifying mitigation opportunities for common bycatch in pelagic longline fisheries. Biological Conservation 142(11), 2620-2630.
| Crossref | Google Scholar |
Dapp DR, Walker TI, Huveneers C, Reina RD (2016) Respiratory mode and gear type are important determinants of elasmobranch immediate and post-release mortality. Fish and Fisheries 17(2), 507-524.
| Crossref | Google Scholar |
Dulvy NK, Pacoureau N, Rigby CL, Pollom RA, Jabado RW, Ebert DA, Finucci B, Pollock CM, Cheok J, Derrick DH, Herman KB, Sherman CS, VanderWright WJ, Lawson JM, Walls RHL, Carlson JK, Charvet P, Bineesh KK, Fernando D, Ralph GM, Matsushiba JH, Hilton-Taylor C, Fordham SV, Simpfendorfer CA (2021) Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Current Biology 31(21), 4773-4787.e8.
| Crossref | Google Scholar | PubMed |
Dulvy NK, Pacoureau N, Matsushiba JH, Yan HF, VanderWright WJ, Rigby CL, Finucci B, Sherman CS, Jabado RW, Carlson JK, Pollom RA, Charvet P, Pollock CM, Hilton-Taylor C, Simpfendorfer CA (2024) Ecological erosion and expanding extinction risk of sharks and rays. Science 386(6726), eadn1477.
| Crossref | Google Scholar |
Ellis JR, McCully Phillips SR, Poisson F (2017) A review of capture and post-release mortality of elasmobranchs. Journal of Fish Biology 90(3), 653-722.
| Crossref | Google Scholar | PubMed |
Frick LH, Reina RD, Walker TI (2010) Stress related physiological changes and post-release survival of Port Jackson sharks (Heterodontus portusjacksoni) and gummy sharks (Mustelus antarcticus) following gill-net and longline capture in captivity. Journal of Experimental Marine Biology and Ecology 385(1–2), 29-37.
| Crossref | Google Scholar |
Gilman E, Chaloupka M, Swimmer Y, Piovano S (2016) A cross-taxa assessment of pelagic longline by-catch mitigation measures: conflicts and mutual benefits to elasmobranchs. Fish and Fisheries 17(3), 748-784.
| Crossref | Google Scholar |
Gilman E, Chaloupka M, Benaka LR, Bowlby H, Fitchett M, Kaiser M, Musyl M (2022) Phylogeny explains capture mortality of sharks and rays in pelagic longline fisheries: a global meta-analytic synthesis. Scientific Reports 12(1), 18164.
| Crossref | Google Scholar |
Godin AC, Carlson JK, Burgener V (2012) The effect of circle hooks on shark catchability and at-vessel mortality rates in longlines fisheries. Bulletin of Marine Science 88(3), 469-483.
| Crossref | Google Scholar |
Gribble NA, McPherson G, Lane B (1998) Effect of the Queensland Shark Control Program on non-target species: whale, dugong, turtle and dolphin: a review. Marine and Freshwater Research 49(7), 645-651.
| Crossref | Google Scholar |
Gulak SJB, Carlson JK (2021) Less soak time saves those upon the line: capture times and hooking mortality of sharks caught on bottom longlines. North American Journal of Fisheries Management 41(3), 791-808.
| Crossref | Google Scholar |
Haig JA, Lambert GI, Sumpton WD, Mayer DG, Werry JM (2018) Habitat features influence catch rates of near-shore bull shark (Carcharhinus leucas) in the Queensland Shark Control Program, Australia 1996–2012. Estuarine, Coastal and Shelf Science 200, 289-300.
| Crossref | Google Scholar |
Henderson CJ, Gilby BL, Turschwell MP, Goodridge Gaines LA, Mosman JD, Schlacher TA, Borland HP, Olds AD (2024) Long term declines in the functional diversity of sharks in the coastal oceans of eastern Australia. Communications Biology 7(1), 611.
| Crossref | Google Scholar |
Holmes BJ, Sumpton WD, Mayer DG, Tibbetts IR, Neil DT, Bennett MB (2012) Declining trends in annual catch rates of the tiger shark (Galeocerdo cuvier) in Queensland, Australia. Fisheries Research 129–130, 38-45.
| Crossref | Google Scholar |
Kyne PM, Heupel MR, White WT, Simpfendorfer CA (2021) The action plan for Australian sharks and rays 2021. (National Environmental Science Program, Marine Biodiversity Hub: Hobart, Tas., Australia) Available at https://www.nespmarine.edu.au/system/files/Shark_Action_Plan_FINAL_Sept7_2021_WEB_RGB.pdf
Lipscombe RS, Scott A, Morris S, Peddemors VM, Smoothey AF, Butcher PA (2023) The influence of bait position on the catch of target and non-target sharks in a SMART drumline bather protection program. Fisheries Research 257, 106501.
| Crossref | Google Scholar |
Lyons K, Jarvis ET, Jorgensen SJ, Weng K, O’Sullivan J, Winkler C, Lowe CG (2013) The degree and result of gillnet fishery interactions with juvenile white sharks in southern California assessed by fishery-independent and -dependent methods. Fisheries Research 147, 370-380.
| Crossref | Google Scholar |
Massey Y, Sabarros PS, Bach P (2022) Drivers of at-vessel mortality of the blue shark (Prionace glauca) and oceanic whitetip shark (Carcharhinus longimanus) assessed from monitored pelagic longline experiments. Canadian Journal of Fisheries and Aquatic Sciences 79(9), 1407-1419.
| Crossref | Google Scholar |
Matias Braccini J, Waltrick D (2019) Species-specific at-vessel mortality of sharks and rays captured by demersal longlines. Marine Policy 99, 94-98.
| Crossref | Google Scholar |
Mitchell JD, Scott-Holland TB, Butcher PA (2022) Factors affecting shark detection from drone patrols in southeast Queensland, eastern Australia. Biology 11(11), 1552.
| Crossref | Google Scholar |
Morgan A, Carlson JK (2010) Capture time, size and hooking mortality of bottom longline-caught sharks. Fisheries Research 101(1-2), 32-37.
| Crossref | Google Scholar |
Morgan A, Cooper PW, Curtis T, Burgess GH (2009) Overview of the US east coast bottom longline shark fishery, 1994-2003. Marine Fisheries Review 71(1), 23.
| Google Scholar |
Mucientes G, Vedor M, Sims DW, Queiroz N (2022) Unreported discards of internationally protected pelagic sharks in a global fishing hotspot are potentially large. Biological Conservation 269, 109534.
| Crossref | Google Scholar |
Musyl MK, Gilman EL (2019) Meta-analysis of post-release fishing mortality in apex predatory pelagic sharks and white marlin. Fish and Fisheries 20(3), 466-500.
| Crossref | Google Scholar |
Reinhardt JF, Weaver J, Latham PJ, Dell’Apa A, Serafy JE, Browder JA, Christman M, Foster DG, Blankinship DR (2018) Catch rate and at-vessel mortality of circle hooks versus J-hooks in pelagic longline fisheries: a global meta-analysis. Fish and Fisheries 19(3), 413-430.
| Crossref | Google Scholar |
Rodríguez-Cabello C, Sánchez F (2017) Catch and post-release mortalities of deep-water sharks caught by bottom longlines in the Cantabrian Sea (NE Atlantic). Journal of Sea Research 130, 248-255.
| Crossref | Google Scholar |
Roff G, Brown CJ, Priest MA, Mumby PJ (2018) Decline of coastal apex shark populations over the past half century. Communications Biology 1(1), 223.
| Crossref | Google Scholar |
Santos CC, Rosa D, Gonçalves JMS, Coelho R (2024) A review of reported effects of pelagic longline fishing gear configurations on target, bycatch and vulnerable species. Aquatic Conservation: Marine and Freshwater Ecosystems 34(1), e4027.
| Crossref | Google Scholar |
Shiffman DS, Ajemian MJ, Carrier JC, Daly-Engel TS, Davis MM, Dulvy NK, Grubbs RD, Hinojosa NA, Imhoff J, Kolmann MA, Nash CS, Paig-Tran EWM, Peele EE, Skubel RA, Wetherbee BM, Whitenack LB, Wyffels JT (2020) Trends in chondrichthyan research: an analysis of three decades of conference abstracts. Copeia 108(1), 122-131.
| Crossref | Google Scholar |
Simpfendorfer C (1992) Biology of Tiger Sharks (Galeocerdo cuvier) caught by the Queensland Shark Meshing Program off Townsville, Australia. Marine and Freshwater Research 43(1), 33-43.
| Crossref | Google Scholar |
Simpfendorfer CA, Heupel MR, White WT, Dulvy NK (2011) The importance of research and public opinion to conservation management of sharks and rays: a synthesis. Marine and Freshwater Research 62(6), 518-527.
| Crossref | Google Scholar |
Simpfendorfer C, Derrick D, Bin Ali A, Fahmi V, V. Q, Tanay D, Seyha L, Haque AB, Fernando D, Bineesh KK, Utzurrum JAT, Yuneni RR, Maung A (2021) Tawny nurse shark Nebrius ferrugineus. In ‘The IUCN Red List of Threatened Species 2021’. e.T41835A173437098. (International Union for Conservation of Nature and Natural Resources) Available at https://www.iucnredlist.org/species/41835/173437098
Sims DW (1996) The effect of body size on the standard metabolic rate of the lesser spotted dogfish. Journal of Fish Biology 48(3), 542-544.
| Crossref | Google Scholar |
Skomal GB (2007) Evaluating the physiological and physical consequences of capture on post-release survivorship in large pelagic fishes. Fisheries Management and Ecology 14(2), 81-89.
| Crossref | Google Scholar |
Sulikowski JA, Golet W, Hoffmayer ER, Driggers WB, III, Natanson LJ, Carlson A, Sweezey BB (2020) Observing post-release mortality for dusky sharks, Carcharhinus obscurus, captured in the U.S. pelagic longline fishery. Fisheries Research 221, 105341.
| Crossref | Google Scholar |
Sumpton WD, Taylor SM, Gribble NA, McPherson G, Ham T (2011) Gear selectivity of large-mesh nets and drumlines used to catch sharks in the Queensland Shark Control Program. African Journal of Marine Science 33(1), 37-43.
| Crossref | Google Scholar |
Walker TI, Hudson RJ, Gason AS (2005) Catch evaluation of target, by-product and by-catch species taken by gillnets and longlines in the shark fishery of south-eastern Australia. Journal of Northwest Atlantic Fishery Science 35, 505-530.
| Crossref | Google Scholar |
Whitney NM, Lear KO, Morris JJ, Hueter RE, Carlson JK, Marshall HM (2021) Connecting post-release mortality to the physiological stress response of large coastal sharks in a commercial longline fishery. PLoS ONE 16(9), e0255673.
| Crossref | Google Scholar |
Yan H, Zhou C, Gilman E, Cao J, Wan R, Zhang F, Zhu J, Xu L, Song L, Dai X, Tian S (2025) A meta-analysis of bycatch mitigation methods for sea turtles vulnerable to swordfish and tuna longline fisheries. Fish and Fisheries 26(1), 45-64.
| Crossref | Google Scholar |