

Grizzly bear response to translocation into a novel environment
Gordon B. Stenhouse A * , Terrence A. Larsen A , Cameron J. R. McClelland A , Abbey E. Wilson B , Karen Graham A , Dan Wismer A , Paul Frame C and Isobel Phoebus A
A * , Terrence A. Larsen A , Cameron J. R. McClelland A , Abbey E. Wilson B , Karen Graham A , Dan Wismer A , Paul Frame C and Isobel Phoebus A
A Grizzly Bear Program, fRI Research, 1176 Switzer Drive, Hinton, AB T7V 1V3, Canada.
B Department of Veterinary Biomedical Sciences, University of Saskatchewan, 52 Campus Drive, Saskatoon, SK S7N 5B4, Canada.
C Alberta Environment and Parks, 9920 108 Street NW, Edmonton, AB T5K 2M4, Canada.
Wildlife Research 49(6) 540-556 https://doi.org/10.1071/WR21060
Submitted: 31 March 2021 Accepted: 14 January 2022 Published: 30 March 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution 4.0 International License (CC BY)
Abstract
Context: To mitigate conservation conflicts, some jurisdictions translocate large carnivores into novel environments where they have no previous experience. Behavioural responses of these individuals are not typically monitored to evaluate the impacts of this management approach on the animals or populations.
Aims: We examined how grizzly bears (Ursus arctos) involved in conservation conflict responded to novel environments after translocation.
Methods: We used GPS location data to compare grizzly bears translocated to a novel environment (n = 12) with bears resident within the new area (n = 12). Our analyses investigated differences between these groups in relation to exploration behaviour, habitat use and response to human-caused mortality risk.
Key results: Translocated bears had higher movement rates, greater daily displacement and revisited areas less frequently than did resident bears. They spent more time in poor-quality habitat and the habitat used was of even lower quality in the second year after translocation. Translocated bears selected for agricultural lands and active oil and gas wellsites. They also spent more time in areas with higher potential mortality risk than resident bears. However, translocated bears avoided residential areas, which resident bears selected, and crossed roads at the same rate as did resident bears. Both groups avoided campgrounds and recreation sites. Only 25% of bears engaged in further conflict behaviour after translocation and 67% of translocated bears survived over the 2-year monitoring period.
Conclusions: This work found differences in exploration behaviour and habitat use between translocated and resident bears, and showed that translocated bears can survive without reoffending during the critical few years following translocation.
Implications: Managers and the public should recognise potential impacts for translocating grizzly bears. Translocated bears require time to explore and learn within their new environment, a process that can occur without repeating conflict. When conflict behaviour does re-occur, it should be viewed as a part of animal learning and not immediately as failure. On the basis of the amount of time translocated bears spent near humans and anthropogenic features, our findings suggest that translocated bears do not pose a greater threat to humans than do resident bears.
Keywords: habitat use, human dimensions, outcome assessment, population management, radio telemetry, survival, threatened species, wildlife management.
Introduction
The growth of human populations and their activities in wildlife habitats are expected to increase over time, which is likely to result in a rise in conservation conflicts (Ripple et al. 2014) and possible impacts on animal behaviour (Champagnon et al. 2012; Gaynor et al. 2018). There is a long history in wildlife management that utilises aversive conditioning, lethal control, or translocation when wildlife comes into conflict with human activities, particularly agriculture, ranching and industrial development (Minteer and Collins 2010). Translocating wildlife is a global phenomenon that has been applied to many species (Seddon et al. 2012). However, animals are not only translocated because of conflict with humans, but are also moved to unfamiliar (hereafter novel) environments as a part of species reintroduction or population augmentation projects (Nichols and Williams 2006; Seddon et al. 2012; Berger-Tal and Saltz 2014). Translocation is a commonly used management practice (Treves and Karanth 2003); however, it comes with another set of impacts to the animal involved. To establish a home range and integrate with the resident populations of both people and conspecifics, translocated animals need to adapt to their new environment through exploration (Burns 2005). Animals in a novel environment will need to locate food and other resources (Frair et al. 2007), and may encounter higher mortality risk as a part of the learning process during exploration (Berger-Tal and Saltz 2014). Although the use of these wildlife management approaches is common practice, the consequences of these actions on animal behaviour are rarely monitored or studied (Fischer and Lindenmayer 2000). Whether for reintroduction or conservation conflict management, the common theme is that individual animals are moved into novel environments where they do not have knowledge of the resources available or anthropogenic features that could threaten their survival (e.g. roads and human settlements).
When conservation conflicts arise, mitigation strategies often seek to prevent further interactions by determining the cause of wildlife presence or attraction and then reducing, eliminating or securing the source of the attraction (e.g. garbage or livestock). Wildlife managers involved in the conflict situation often mistakenly expect an immediate cessation of conflict behaviours following the removal of attractants. These conflict behaviours may have developed after a single event, over a longer period, or as a result of social learning (i.e. female bears passing on conflict behaviours to their offspring during the cub rearing period; Morehouse et al. 2016), prior to management intervention. The lack of an immediate response by conflict animals following attractant removal, along with possible continued risk to humans and/or their property, commonly results in the decision to translocate or take lethal control action. In conservation conflict situations, destruction of the animal is the least preferred management option because of direct and immediate population consequences, and this approach often results in negative feedback from the public (Dubois et al. 2017; Lewis et al. 2019), which is of greater concern when the species involved is listed as threatened or endangered. In conflict instances involving species-at-risk, there is a strong preference to either attempt aversive conditioning practices or to capture and translocate the animal (Spencer et al. 2007). Scientists and managers are often faced with questions related to the translocation of wildlife covering topics such as return and survival rates, continuation of conflict behaviour in new areas, and risk to humans from a translocated bear. Therefore, data are needed to assist wildlife management professionals in gaining a greater understanding of how translocated animals respond to a novel environment, and that can be useful for sharing with the public when and where translocation events occur.
Grizzly bears (Ursus arctos) have been a candidate for a large number of translocation events because of public safety or property damage, coupled with how they are viewed by the public relative to conservation and their limited distribution on the landscape (Linnell et al. 1999; Massei et al. 2010). Several authors have investigated translocation success of grizzly bears, but how success is defined varies among studies (Riley et al. 1994; Blanchard and Knight 1995; Miller et al. 1999; Milligan et al. 2018). Studies have shown that the survival rate of translocated grizzly bears is much lower than that of bears not translocated within a population (Brannon 1987; Blanchard and Knight 1995). Translocated bears may return to the capture or conflict site (i.e. homing) in a short time period; however, in general, the further the translocation distance, the lower the return rate and probability of reoffending in their new environment (Brannon 1987; Blanchard and Knight 1995; Clark et al. 2002; Milligan et al. 2018). Moving a grizzly bear, or any animal, into a novel environment presents many challenges to the individual; food resources may be different, unfamiliar mortality risks may exist and social dynamics among conspecifics need to be established. How translocated animals behave in a new environment is likely to play an important role in their eventual fate. When bears are translocated and further conflict (i.e. a reoffence) is not reported, the event is typically recorded as successful. The focus of most translocation management actions, which are not commonly monitored, is on outcomes of success or failure, with little attention on the consequences to the individual animal. In particular, there remains a lack of knowledge on the individual behavioural response of translocated animals in a novel environment (Pinter-Wollman et al. 2009). Understanding individual animal responses may play an important role in wildlife management decision-making, improve translocation outcomes and support science-based conservation.
This study was undertaken to determine the influence of translocation on individual grizzly bears involved in conservation conflicts in Alberta, Canada, between 2016 and 2019. Grizzly bears have been listed as a threatened species in Alberta since 2010 (Festa-Bianchet 2010) and bears involved in conflict situations are translocated as part of regular management actions. We predicted that translocated bears move, use habitat and respond differently in relation to modelled mortality risk from bears already present on the landscape and hypothesised that translocations influence the way bears respond to a novel landscape. To test our hypothesis, we investigated how translocated bears (in relation to resident bears) respond to a novel landscape by examining: (1) their exploration (movement rates and behaviour); (2) habitat use (time spent in and revisits to high-/low-quality habitat); and (3) response to human-caused mortality risk (hereafter mortality risk) factors (time spent near and selection of anthropogenic features and habitat mortality risk, risk at site revisits, road crossings and road area use). Previous research has shown increased exploration when animals are moved to a novel environment, represented by movement behaviours (rate and displacement; Seddon et al. 2012; Berger-Tal and Saltz 2014; Hertel et al. 2019). These changes in movement behaviour can lead to increased energy expenditure (Carnahan et al. 2021) and, in combination with the use of low-quality and/or risky habitat, can result in poor body condition or low survival (Boulanger and Stenhouse 2014; Bourbonnais et al. 2014). This research will improve our understanding of translocation effects and, more broadly, will provide needed information to managers that translocate bears to address conflict situations or support conservation and population recovery efforts.
Materials and methods
Study area
Our study was conducted within two provincial bear-management areas (BMAs), the Yellowhead (28 774 km2; BMA 3) and Clearwater (17 709 km2; BMA 4), in west-central Alberta, Canada (Fig. 1). The western extent of the study area is mountainous and contains a large network of protected areas managed under national and provincial jurisdiction. From west to east, the area transitions along an elevational gradient into rolling foothills characteristic of the boreal forest. At the eastern extent is a mixture of public and private agricultural lands. Portions of the study area outside of the protected areas have varying degrees of natural resource extraction activities (e.g. forestry, oil and gas, mining), which are dependent on an extensive network of gravel roads (Boulanger and Stenhouse 2014). Recreation in the area includes hiking, camping, fishing, hunting and off-highway vehicle use.

|
The study area includes four natural subregions, including alpine, subalpine, and upper and lower foothills, which are distinguished by variations in vegetation, climate, elevation and latitudinal or physiographic differences (Achuff 1994). The most widespread tree species is lodgepole pine (Pinus contorta), with other species such as black spruce (Picea mariana), white spruce (P. glauca), trembling aspen (Populus tremuloides), balsam poplar (P. balsamifera) and balsam fir (Abies balsamea) occurring at lower densities and elevations. Several ungulate species are present in the study area and known to be consumed by grizzly bears, including moose (Alces alces), elk (Cervus canadensis), white-tailed deer (Odocoileus virginianus), mule deer (O. hemionus) and bighorn sheep (Ovis canadensis). A variety of plants also important for grizzly bears are seasonally available (Munro et al. 2006), such as alpine sweet-vetch (Hedysarum alpinium), graminoids and forbs such as clover (Trifolium spp.), cowparsnip (Heracleum lanatum), dandelion (Taraxacum officiale) and horsetail (Equisetum spp.). Berry-producing shrub species include blueberry (Vaccinium myrtiloides), huckleberry (V. membranaceum) and buffaloberry (Shepherdia canadensis), which are also commonly found and tend to be available during late summer and early fall (Munro et al. 2006; Larsen et al. 2019).
Study animals
We performed all grizzly bear capture and handling following established procedures (Cattet et al. 2008), which minimised the time bears were held prior to release. We captured resident bears by using either remote drug delivery from a helicopter or ground-based capture with culvert traps. Translocated bears were captured solely with culvert traps (Cattet et al. 2008). Grizzly bears were fitted with Followit (Lindesberg, Sweden) satellite GPS collars that provided locations every 30 or 60 min. We performed external physical examinations, recorded morphological measures (length, girth and weight), collected hair, blood and tissue samples, as well as a premolar tooth to determine the age and overall physical condition of each individual (Cattet et al. 2003). Capture and handling protocols were undertaken in accordance with the Canada Council of Animal Care and approved by the University of Saskatchewan Animal Care Committee (Approval 20010016).
Between 2016 and 2019, we collared and translocated 12 grizzly bears (six females and six males; Supplementary Table S1) involved in conservation conflicts. Translocations were a result of three types of conflictual interactions, namely, livestock depredation (eight bears), accessing human food sources (e.g. garbage; two bears) and proximity to people (two bears; Table 1). Conflict events were identified when officially reported to the government management agency. Following investigation, provincial enforcement staff captured and, in collaboration with the research team, handled, sampled and transported these animals in a culvert trap by truck to remote sites. Release sites were based on suitable habitat characteristics represented by high resource selection function (RSF) scores (Boyce et al. 2002), low levels of human use as identified by Milligan et al. (2018), and logistics in the interest of animal health and welfare. RSF scores were calculated by identifying a disproportionate use of resources (e.g. habitat, landscape), by using a 'used' and 'available' design in logistic models (wherein 1 means used and 0 means available). Translocated bears were moved either to a different location within BMA 3 or from another BMA into BMA 3 (Fig. 1). We classified bears into the following two categories depicting the distance between the capture and translocation site: short moves (<100 km; i.e. relocation; see Graham and Stenhouse 2014) and long moves (>100 km; i.e. translocation; Milligan et al. 2018). Bears were translocated, on average, 284 km from the capture site (from 32 to 449 km). Nine (75%) and three (25%) bears were translocated long and short distances respectively (Table 1).

|
We selected 12 resident bears (six females and six males) from a long-term dataset of collared grizzly bears in west-central Alberta. To establish a comparable sample group, we limited our selection to resident bears within the same management unit and with data during the same years as translocated bear GPS location data from 2016 to 2018 (no resident bear collar data were available in BMA 3 during 2019; Table S1). We selected resident bears with GPS location data across seasons (spring to fall) for bears with multiple years of data when possible. Resident bears were constrained to those that lived in subregions similar to those where translocated bears were released (i.e. the foothills rather than alpine and subalpine regions within national parks). Our final criterium was to select resident bears that matched translocated bears according to age (subadult/adult) and sex classes (female/male; Table S1). All translocated and resident bears appeared to be in good health on the basis of morphological measures when captured, with no visible injuries or diseases being detected through blood sample analysis.
GPS location data analysis
We partitioned GPS location data into three seasons according to grizzly bear feeding behaviour in Alberta (Nielsen et al. 2004c; Munro et al. 2006). Seasons were defined as spring (1 May to 15 June), summer (16 June to 31 July), and fall (1 August to 15 October). Grizzly bear GPS locations for both translocated and resident bears were resampled at hourly intervals. We categorised hourly GPS locations by time of day (day, night, twilight morning, or twilight night) on the basis of sunrise and sunset tables for the study area (National Research Council of Canada 2015). Collar life, and hence GPS location data, varied among bears, with the number of days of data obtained following translocation averaging 413 days (from 256 to 797 days; Table S1). We further classified translocated and resident bears (hereafter, bear class), age class (subadults, i.e. bears less than 5 years of age, or adults), sex (males and females) and female reproductive status (females moved with cub(s) of the year or yearlings). We analysed GPS location data by using R statistical software (R version 3.6.0; R Core Team 2019, R: a language and environment for statistical computing, available at http://www.rproject.org/) to determine whether translocated and resident grizzly bears differed in use (e.g. time spent and site revisits) and selection of features within their novel environment. For simple comparisons, means for each exploration behaviour and response to mortality risk were compared between translocated and resident bears, by using either a Student’s t-test if the data were normally distributed (daily distance travelled and daily displacement) or a Wilcoxon signed-rank test if the data were not normally distributed (road crossings and road area use). To determine selection, we created RSF models for anthropogenic features and mortality risk. RSF models were fitted using used locations and generated available location data and GLMMs (glmer function, lme4 package version 1.1.27; Bates et al. 2016, Package ‘lme4’, 1–113, available at https://cran.r-project.org/web/packages/lme4/lme4.pdf). We removed bears with fewer than 60 locations or if locations occurred over fewer than 30 consecutive days in a given year. We then used the adehabitatHR package version 0.4.19 (Calenge 2006) to calculate areas that bears reside, by using 95% minimum convex polygons (MCP), where only 95% of locations were used to account for outliers. We generated five random available locations per square kilometre for each individual MCP (Nielsen et al. 2004c; Roever et al. 2008). GPS location data were used for the following analyses and reported as mean ± standard deviation (s.d.), with statistical significance at P < 0.05.
Exploration of novel landscapes
Movement rates
We performed analyses to determine the relationship between grizzly bear movement rates and the explanatory variables, distinguishing resident and translocated bears, age, sex and reproductive class, year, day of the year, hour and location coordinates. We calculated movement rates by using the distance travelled between GPS location points, standardising our data to 30 and 60 min intervals (as.ltraj function, adehabitatLT package, version 0.3.25). Given the time series nature of the movement-rate data, we used gamma-distributed generalised additive models (GAM; gam function, mgcv package, version 1.8.33; Wood 2017) with smoothing functions for day of year and hour because of potential non-linear temporal effects (seasonal and daily) on movement rates (Graham and Stenhouse 2014). We additionally included a smoothing function for the location coordinates to account for spatial correlation within the data. We included interactions between bear class and the day of year, hour and location effects to test whether movement rates for resident and translocated bears changed differently over time and space. To account for subject non-independence within the data, we used the individual bear ID as a random intercept. We evaluated competing models by using Akaike information criterion (AIC) values (Burnham and Anderson 2002), and we checked model assumptions of independence, homoscedasticity, normality of residuals and random effects (Zuur et al. 2010). We found no significant effect of day post-capture or translocation event on movement rates within 30 days, when taking bear class, age/sex/reproductive class, year, day of year and location into account (GAM, t-value = 0.297, P = 0.767; data not shown). Therefore, we excluded translocation and capture days from the movement dataset used for movement rates, distance travelled and displacement analyses.
Daily distance travelled
The daily distance travelled indicates how far an individual bear moves within a day, a measure that can relate to individual variation in activity, use of resources, energy, or the likelihood of finding quality food (Hertel et al. 2019). We generated the daily distance travelled using the sum of Euclidian distances calculated between consecutive GPS location points. To avoid underestimation of the cumulative daily travel distance, we limited the data to days containing 100% (24 of 24) of the hourly locations.
Daily displacement
We examined daily displacement, a measure that shows how far a bear moves during the day from its starting location, to determine whether resident or translocated bears moved farther around the landscape. We calculated daily displacement as the Euclidian distance between GPS location points at approximately midnight each day. We subsampled our GPS location data to points only at midnight and recalculated output data to get the time and distance between those points. We retained data with time differences of 24 h to maintain data for consecutive days.
Diurnality
We assessed whether translocated bears shifted the timing of their active behaviour compared with resident bears. To do this, we calculated a diurnality index that identifies whether a bear is more active during the day or night over time (e.g. across months or seasons). We separated our GPS location points into day and night only, incorporating twilight times within their respective categories. For each bear, we generated a diurnality index by age, sex and bear class with an Eqn 1 adapted from previous studies (Hoogenboom et al. 1984; Hertel et al. 2019), as follows:
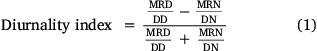
where MRD and MRN are the sums of hourly movement-rate values during the day and night, and DD and DN are the durations of the day and night respectively, determined by sunrise and sunset times (National Research Council of Canada 2015). The diurnality index varies between −1 (night active) and 1 (day active). We examined differences in the diurnality index between bear classes across months outside of the denning period.
Use of novel landscapes
Quality of habitat use
To determine the quality of habitat used from GPS location data of translocated and resident bears, we used previously established and maintained (by fRI Research) models on the relative probability of space used as a surrogate of habitat quality (RSF layers created in house by using methods from Nielsen et al. 2010 and updated annually). We calculated the percentage of time spent within poor- to high-quality habitat (RSF bin values from 1 to 10 respectively) as the number of hourly GPS locations per habitat value divided by the total number of locations. For each GPS location point, we used concurrent annual and seasonal RSF data with the most recent available data (2018). We excluded GPS location data that fell outside of the season dates or the RSF layer extent.
Habitat quality of GPS site revisits
By using the same previously created habitat-quality RSF layer as above, we calculated the proportion of time translocated and resident bears spent revisiting areas of varying habitat quality. We calculated two categories of revisits, namely, between-year and within-year revisits. Within-year revisits were calculated using the Recurse package version 1.1.2 (Bracis et al. 2018), and were defined as a location where a bear returned to within 100 m of a point, after a minimum of 14 days. Between-year revisits were defined as a bear returning to within 500 m of a point occupied in the previous year. The spatial scales of 100 m for within-year revisits and 500 m for between-year revisits differ to account for temporal differences between the two revisit categories. With the longer time period with between-year revisits, we chose a coarser spatial scale to account for landscape change and changes in food availability. For each revisit, we extracted the RSF bin value from 1 to 10. We then calculated the proportion of GPS site revisits in each bin as a percentage of the total number of GPS locations.
Mortality risk in novel landscapes
Mortality risk and time spent near humans
To model time spent in areas of high mortality risk, we used previously created (Nielsen et al. 2004b) and annually updated (by fRI research) mortality-risk layers. First, we calculated the percentage of time spent within low to high mortality-risk habitat (0–10 risk bin values respectively) by using the RSF layers described above for translocated and resident bears. We limited annual mortality risk data to the same seasonal period so as to compare with RSF values, and excluded points outside the mortality risk extent. We assessed time spent near human activity for each GPS location by determining whether it was within 100 m (McLellan and Shackleton 1989; Proctor et al. 2020) of agricultural lands (e.g. abandoned agricultural lands, fruit and vegetable crops, pastures), industry facilities, roads (e.g. gravel, paved and unimproved roads), residential lands (e.g. urban, rural and country residential building footprints), campgrounds and recreational day-use sites, and active oil and gas well sites (ABMI 2018). We then calculated a percentage of total GPS locations for translocated and resident bears spent near each human landscape class.
Mortality risk of GPS site revisits
We further investigated the difference in mortality risk of GPS within-year and between-year site revisits, using the same methods as for the habitat-quality revisits (see above), of translocated and resident bears by using the mortality risk layers. For each revisit, we extracted the mortality risk bin values from 0 to 10. The proportion of GPS site revisits in each bin were then calculated as a percentage of the total number of GPS site revisits.
Road crossings and road area use
We compared the number of road crossings and the amount of time spent within 50 m of a road between translocated and resident bears, because proximity to roads is a known factor in grizzly bear mortality (Schwartz et al. 2003; Nielsen et al. 2004b; Boulanger and Stenhouse 2014). We generated numbers of road crossings for different road types (i.e. paved, gravel, truck trail, unimproved, and other) for each individual bear year by calculating straight-line trajectories from grizzly bear GPS locations and determining the number of intersections with roads summarised by road type, time of day and time spent within 50 m of a road. We calculated road density within the area of grizzly bear utilisation (95% MCP referenced above) and computed the proportion of road crossings by individual bear and divided it by the total density of roads within each individual’s area; higher ratio values indicated more road crossings relative to density. We compared translocated and resident bear road crossing ratios in response to all roads, road types, time of day and sex of the animal.
Selection near anthropogenic features and mortality risk
To compare how translocated and resident bears perceive and select for different attributes of their environment, we created new RSF models using a RSF Type III approach (Boyce et al. 2002). This approach compares used and generated random available locations to determine whether bears select for human-activity and high-mortality risk areas disproportionately more than what is available within each bear's home range. We created separate RSFs for each bear class by using GLMMs with aggregated foraging seasons and added each human-activity (described above) variable separately to the core model (Table 2). To select the best model, we compared candidate models to the core model by using environmental variables known to influence grizzly bear habitat selection from previous research in the region (Roever et al. 2008; Nielsen et al. 2010; Berman et al. 2019; Kearney et al. 2019; McClelland et al. 2020). Our core model consisted of land cover, elevation, compound topographic index (i.e. soil wetness), distance to roads and distance to forest edge, which were tested for and added as quadratic terms, given previous evidence of non-linear relationships (Roever et al. 2008; Nielsen et al. 2010; Berman et al. 2019; Kearney et al. 2019; McClelland et al. 2020). For all models, we included a nested random intercept to account for differing response magnitudes among individual bears of different age and sex classes (Nielsen et al. 2004a; Roever et al. 2008; Kearney et al. 2019). We then created a model that encompassed all human-activity areas and mortality risk for both translocated and resident bears, and we selected a final model by using only human-activity areas deemed significant (P < 0.1). We selected models with the lowest AIC score for each bear class (Burnham and Anderson 2002). Odds ratios calculated from the best performing models determined the relative strength of selection for each human-activity area in the final models, where a ratio above one indicates selection and a ratio below one indicates avoidance.

|
Results
Exploration of novel landscapes
Movement rates varied significantly by bear class and age/sex/reproductive class (Table 3). Two-dimensional smoothing functions for day of year and hour, and for spatial coordinates with interaction terms for translocated or resident bears were also significant (Table 3), indicating a non-linear relationship between these variables and movement rates. Translocated bears moved faster (i.e. at a higher rate in km h−1) than did resident bears when taking age/sex/reproductive class, day of year, hour and location into account (GAM, t-value = 4.09, P < 0.001; Table 4). We found no significant difference in mean daily distance travelled between translocated and resident bears (t-test, t = −1.11, d.f. = 19, P = 0.28; translocated: 11 248 ± 2357; and resident: 9562 ± 1642 m day−1). However, we did find a significant difference in mean daily displacement (t-test, t = −2.35, d.f. = 16, P = 0.03; translocated: 6793 ± 7211; and resident: 4245 ± 3805 m day−1), where translocated bears had higher displacement than did residents (Supplementary Fig. S1). Diurnality between resident and translocated bears was similar throughout the year. However, some translocated bears showed higher individual variation and less daytime activity (Fig. S2).

|

|
Use of novel landscapes
Quality of habitat use
Translocated bears spent 24% of their time in the poorest-quality habitat (i.e. the lowest RSF bin value; 3% for resident bears), whereas resident bears spent the most time (31%) in the highest-quality habitat (RSF 10; 16% for translocated bears; Fig. 2a). Most translocated bears with multiple years of data (88%, 7 of 8 bears) spent more time in higher RSF-valued habitat during the first year of translocation than during the second. All resident bears with multiple years of data (100%, 7 of 7 bears) used similar-quality habitat each year (Fig. S3).

|
Habitat quality of GPS site revisits
When we compared within-year GPS site revisits between resident and translocated bears, we found that 32% of total locations were revisited for resident bears, whereas for translocated bears, only 8% of locations were revisited. Between-year GPS site revisit calculations showed that for resident bears, 66% of total locations were revisited from the previous year, whereas for translocated bears, only 8% of locations were within 500 m of a location from the previous year. When comparing revisits of habitat within a year, we observed that resident bears revisited higher-quality habitat more frequently than did translocated bears; 39% of all resident bear revisits occurred in the highest-quality habitat (RSF bin value 10) and 67% of all resident bear revisits occurred in the top-three RSF bin values (8–10; Fig. 2b). Conversely, 30% of translocated bear revisits occurred in the highest-quality habitat and 53% in the top-three RSF bin values. We further observed that between-years resident bears revisited more high-quality habitats than did translocated bears; 44% of all resident bear revisits occurred in the highest-quality habitat and 77% of all resident bear revisits occurred in the top-three RSF bin values. Conversely, only 27% of translocated bear revisits occurred in the highest-quality habitat, and only 46% of all translocated bear revisits occurred in the top-three RSF bin values.
Mortality risk in novel landscapes
Mortality risk and time spent near humans
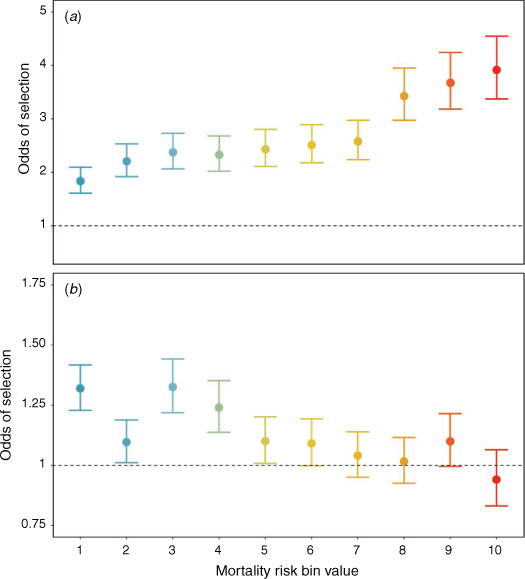
We found that translocated bears spent more time in riskier habitat than did resident bears (Fig. 3a). Translocated bears spent 43% of their time in the highest mortality-risk habitats (bins 8–10; 20% for resident bears), whereas resident bears spent 29% of their time in low-risk habitat (bins 0–2; 17% for translocated bears). Furthermore, translocated bears spent 14% of their time within 100 m of all anthropogenic features, including 9% in agricultural lands, 0.7% in residential lands, 0.8% in industrial areas, 5% in active oil and gas wellsites, and 0.03% in campgrounds and recreation sites. Comparatively, resident bears spent 2% of their time within 100 m of all anthropogenic features, including 0.1% in agricultural lands, 0.6% in residential lands, 0.5% in industrial areas, 0.6% in active oil and gas wellsites, and 0.03% in campgrounds and recreation sites.
Mortality risk of GPS site revisits
When we compared the within-year revisits of risk-habitat levels, translocated bears revisited areas of higher mortality risk more frequently than did resident bears; 8% of all translocated bear within-year revisits occurred in the highest mortality-risk areas (mortality risk bin value 10) and 42% within the top-three bin values (8–10; Fig. 3b). Conversely, 2% of resident bear within-year revisits occurred in the highest mortality-risk areas and 19% occurred in the top-three bin values. When we shift focus to between-year revisits, translocated bears also revisited areas of higher mortality risk more frequently than did resident bears; 4% of all translocated bear between-year revisits occurred in the highest mortality-risk areas and 48% in the top-three bin values. Conversely, only 2% of resident bear between-year revisits occurred in the highest risk habitat, and only 17% in the top-three bin values. We further examined the proportion of revisits as a percentage of total revisits into agricultural lands by translocated and resident bears. We found that although translocated bears had more within-year revisits than did resident bears (8% and 0.3% respectively), only a small portion of between-year revisits were present for both classes (translocated = 1%, resident = 0.2%).

|
Road crossings and road area use
Using Wilcoxon tests, no significant differences were found between translocated and resident bear road-crossing ratios in response to all roads (total crossings; Wilcoxon test W = 132, P = 0.17; translocated: 0.29 ± 0.18; and resident: 0.24 ± 0.24) or when separated by road type (gravel, paved, truck trail, unimproved, other and gravel/paved; Table S2). No significant differences were found between translocated and resident bear road-crossing ratios during the day (Wilcoxon test W = 144, P = 0.30; translocated: 0.18 ± 0.11; and resident: 0.18 ± 0.21), twilight morning (Wilcoxon test W = 152, P = 0.42; translocated: 0.01 ± 0.01; and resident: 0.01 ± 0.01), or twilight night (Wilcoxon test W = 154, P = 0.46; translocated: 0.02 ± 0.01; and resident: 0.01 ± 0.01). However, translocated bears were found to cross roads significantly more at night than were resident bears relative to road density (Wilcoxon test W = 111, P = 0.04; translocated: 0.08 ± 0.09; and resident: 0.04 ± 0.03). When separated by sex, there was no significant difference between translocated and resident bear road-crossing ratios in response to all roads (females: Wilcoxon test W = 30, P = 0.18; translocated: 0.30 ± 0.21; and resident: 0.16 ± 0.07; and males: Wilcoxon test W = 35, P = 0.70; translocated: 0.28 ± 0.17; and resident: 0.34 ± 0.35), road type, or time of day within each sex (Table S2). Furthermore, there was no significant difference between translocated and resident bears with respect to time spent within 50 m of roads (Wilcoxon test W = 167, P = 0.72; translocated: 0.31 ± 0.14; and resident: 0.47 ± 0.45).
Selection near anthropogenic features and mortality risk
The final translocated bear RSF model with the lowest AIC included agricultural and residential lands, campgrounds and recreation sites, active wellsites, and mortality risk bins as significant factors (P < 0.1; Table S3). Industrial facilities were not included in the final model for translocated bears (P = 0.70; Table S3). The final resident bear RSF model included residential lands, campgrounds and recreation sites, active wellsites, and mortality risk bins (P < 0.1; Table S3). Agricultural lands and industrial facilities were not included in the final model for resident bears (agricultural lands: P = 0.20; industrial facilities: P = 0.28; Table S3). When comparing selection for areas near anthropogenic features using odds ratios, we found that translocated bears selected for agricultural lands (odds ratio = 2.4 when compared with non-agricultural lands), avoided residential lands (odds ratio = 0.4 when compared with non-residential lands), avoided campgrounds and recreation sites (odds ratio = 0.4 when compared with non-campgrounds) and selected for active well sites (odds ratio = 1.5 when compared with non-well sites; Fig. 4a). Resident bears selected for residential lands (odds ratio = 1.7 when compared with non-residential lands), avoided campgrounds and recreation sites (odds ratio = 0.1 when compared with non-campgrounds and non-recreation sites) and avoided active well sites (odds ratio = 0.9 when compared with non-well sites; Fig. 4b). When comparing selection for low versus high mortality risk across the landscape using odds ratios, translocated bears selected for areas of low mortality risk, but showed stronger selection, indicated by higher odds ratios when compared with mortality risk bin 0 (no mortality risk), for areas of higher mortality risk. Conversely, resident bears showed stronger selection for lower mortality risk when compared with mortality risk bin 0 (no mortality risk). These results corroborated our finding of time spent in the RSF mortality-risk layer.
|
Discussion
We found that grizzly bears translocated into novel environments showed different behavioural patterns and habitat use than did resident bears who were familiar with the landscape. Translocated bears showed higher movement rates and greater daily displacement. During their exploration, translocated bears used poorer-quality habitat more, suggesting that these bears responded to or utilised resources differently compared with resident bears familiar with the area. Translocated bears also spent more time in areas of greater mortality risk and agricultural areas; however, they avoided residential areas. Overall, very few translocated bears reoffended and most bears survived over the course of the monitoring period.
In this study, we found that translocated grizzly bears exhibited higher movement rates and greater daily displacement than did resident bears, behaviours that are indicative of exploration within a novel environment (Seddon et al. 2012; Berger-Tal and Saltz 2014; Hertel et al. 2019). Moving faster presumably allows translocated bears to explore more area in search of food, which may lead to encountering other bears and/or areas of higher mortality risk. However, greater movement rates can result in increased energy expenditure for these bears (Carnahan et al. 2021), which may result in poorer body condition. Overall, translocated bears travelled similar distances as did resident bears; however, they showed greater daily displacement, moving further into different areas as they travelled across more habitat (Réale et al. 2007; Hertel et al. 2019). In contrast to our results, Hertel et al. (2019) found that individual bears that travelled further also had further displacement.
We found that translocated grizzly bears spent more time in lower-quality habitat, and although we were unable to gather data on the body condition of translocated grizzly bears during the months following release, this may have had negative impacts on their body condition (Bourbonnais et al. 2014). This further suggests that bears have to range farther to find high-quality foods, because they do not know where those resources are yet, while establishing a new home range and interacting with resident bears during their first year of exploration. During their exploration of lower-quality habitat, translocated bears may use olfactory and visual cues to search for food in the novel landscape, instead of established spatial memory of food resources that resident bears have acquired while living in the area (Coogan et al. 2012; Nielsen et al. 2013). Murray et al. (2017) found that grizzly bears develop individual preferences for specific food resources. Therefore, it is possible that foraging behaviours are related to local environmental cues, which could then be difficult to generalise in novel environments.
Previous studies have shown that translocated bears have lower survival rates than do resident bears (Milligan et al. 2018), which may be due to their unfamiliarity with mortality risk in a novel environment or their willingness to take more risk. Our analysis confirmed this, with translocated bears spending more time than resident bears in high-risk habitat, which was additionally supported by weaker avoidance of features indicative of higher risk. Our findings suggest that translocated bears, despite potential previous knowledge of dangerous anthropogenic features in their home environment, recognised or responded to risk in a novel environment differently from resident bears. Translocated bears also revisited high-risk habitat more often than did resident bears, both within and among years, suggesting that these bears were not learning quickly about local risks during their exploration. These differences in response to mortality risk may indicate why lower rates of survival are typically seen with grizzly bear translocation and may even affect other aspects of translocated-bear lives such as feeding behaviour (Hertel et al. 2016). Another concern around translocated bears is their tendency to reoffend or bring potentially destructive behaviours (e.g. killing livestock, disturbing agricultural areas, or causing problems residential and recreational areas) into a new region. In this study, we found that even though translocated bears spent more time near agricultural lands than did resident bears, the overall percentage of time spent there was low, and they avoided other human-activity areas (e.g. residential and recreational camping areas). Only a small percentage of revisits along GPS travel paths made by translocated bears was associated with agricultural lands, which suggests that these bears were not returning to the same agricultural areas and repeating conflict on a continual basis, nor in successive years. When translocated bears selected agricultural lands, they used areas where no residential buildings were present, showing that these bears avoided areas in close proximity to people following their translocation. Other studies have suggested that the time bears spend near people and anthropogenic features could be a measure of ‘boldness’ (Hertel et al. 2019). Using this same metric, we conclude that both resident and translocated bears showed differences in boldness in some circumstances. Our results showed that both translocated and resident bears were not attracted to industrial areas or campgrounds where human activity tends to be high. Furthermore, past research has shown negative relationships between roads and road densities and grizzly bear survival (Boulanger et al. 2018; Proctor et al. 2020). In our study, translocated bears crossed roads at the same rate and spent the same amount of time near roads within a novel environment as did resident bears. However, translocated bears did cross roads more frequently at night, when we presume traffic volumes are most likely to be lower (Northrup et al. 2012), than did resident bears, a difference in behaviour that could indicate a higher degree of wariness by translocated bears around these common landscape features.
Despite spending more time in areas of higher mortality risk, the majority of translocated grizzly bears in our study survived. Others have noted that survival of animals, particularly large carnivores, may be compromised when they are moved to new territories (Stamps and Swaisgood 2007; Stuparyk et al. 2018). Contrary to suggestions that translocation actions have an outcome similar to lethal removal (Treves and Karanth 2003), our results showed a much higher success rate during the period of monitoring. We found that following translocation, 8 of the 12 translocated bears (67%) survived at minimum until the end of collar life, and four (33%, all males) died. Three bears were killed by management personnel in response to further conflicts (25%; two bears with livestock conflicts and one bear because of close proximity to people) and one bear was killed illegally. Note that reoffending translocated bears that were killed were similar to those that survived in terms of their use of high-risk habitat, time near anthropogenic features, and road crossings. A similar amount of repeat conflict behaviour (30%) for relocated black bears has been reported by Landriault et al. (2009); however, they found that adult females were most likely to reoffend. None of the translocated bears returned to their original capture site after either short- or long-distance moves, which differs from previous findings in Alberta that reported homing behaviour for bears moved similar short distances (Milligan et al. 2018). More recently, Bauder et al. (2020) found that translocating black bears (Ursus americanus) greater distances from the capture site did not affect repeated conflict behaviour, but reduced the probability of a translocated bear returning to an original conflict site. Studies have also suggested that prey abundance at the release site may also be a determinant of translocation success, with greater prey abundance resulting in less frequent homing behaviour and attacks on livestock (Hayward et al. 2006; Gusset et al. 2009).
Conclusions and conservation implications
Taken together, our findings have provided evidence that despite extensive movement behaviour related to exploration, translocated bears were not repeating previous conflict behaviours in the novel environments where they were released. Translocated bears spent more time in riskier habitat as defined by mortality-risk models, but avoided human settlements, buildings or facilities, suggesting that translocated bears pose a small risk to humans within our study area. Because some translocated bears in this study reoffended (3 of 12 bears), this should not be seen as failure, but rather as a part of expected exploration behaviour as they have been shown to range farther and use lower-quality habitat for at least 2 years after translocation. Therefore, we suggest that bears that reoffend should be returned to their original release site where some learning has already occurred and not moved to an additional novel environment where they will need to begin exploration activities again. Improved success rates and survival of translocated bears can occur as we integrate this new knowledge into management strategies.
In 1931, Dice suggested that the reaction of an animal to its environment is as equally important as are the characteristics of the environment in enabling the animal to survive (Dice 1931). Although there were no methods available at that time to investigate animal responses to novel environments, our findings support this important point and should be considered when translocating animals for management purposes. Because management agencies use wildlife translocation to address conservation conflict situations, or as a means to augment populations to support recovery or reintroduction actions, it is important to develop practices and protocols to monitor and evaluate the effectiveness of this technique. Regular review of outcomes would allow needed modifications in approaches to be incorporated into science-based best practices. It is important to understand the consequences of translocation for the animal (high levels of exploration over long periods) and how the animal may respond to novel environments (movements and habitat use). Furthermore, translocated bears will likely learn at different rates on the basis of individual behaviours and personality traits (Ordiz et al. 2014; Leclerc et al. 2016), past experiences (Morehouse and Boyce 2017) and individual temperaments (Réale et al. 2007; Armstrong et al. 2015), which was not possible to determine in the current study. The high levels of exploration shown by translocated grizzly bears in this study also suggest that wildlife management efforts focusing only on high-quality habitat to find the ‘ideal’ release site for translocations may need to be complemented with an understanding of exploration behaviour and movement (i.e. the bear may leave the high-quality habitat). In general, wildlife managers should translocate bears to places where there is both a high level of high-quality habitat (because bears are not initially good at finding high-quality habitat) and a land area where it is safe for them to range without reoffending (e.g. large natural areas).
Data availability
The datasets generated during and/or analysed during the current study may be requested from the corresponding author.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
This work was financially supported by Alberta Environment and Parks, and fRI Research. Both sources supported the preparation of the data, the writing of the manuscript, and the decision to submit for publication.
Author contributions
GS and PF conceived the ideas; GS, PF, TL and IP contributed in project development; GS, CM and IP designed methodology; GS, TL and additional staff collared bears for data; DW extracted data; KG managed databases; CM and IP analysed the data; GS, IP, and AW led the writing of the paper, with text, figures and edits from CM, KG, TL. All authors contributed critically to the drafts.
Supplementary material
Supplementary material is available online.
Acknowledgements
We thank the staff with the Grizzly Bear Program at fRI Research for capture and handling of resident bears and report preparation, and enforcement officer staff with the Government of Alberta for assistance with capture and handling of translocated bears. The authors thank M. Cattet for input on the data analysis and preparation of the paper. We also thank the three anonymous reviewers for their insightful and thoughtful comments that improved this paper.
References
Alberta Biodiversity Monitoring Institute (ABMI) (2018) ABMI Human Footprint Inventory: Wall-to-Wall Human Footprint Inventory. (AMBI: Edmonton, AB, Canada) Available at https://abmi.ca/home.htmlAchuff PL (1994) ‘Natural Regions, Subregions and Natural History Themes of Alberta: a Classification for Protected Areas Management-Updated and Revised.’ (Alberta Environment Protection, Parks Services)
Armstrong D, Hayward M, Moro D, Seddon P (2015) ‘Advances in Reintroduction Biology of Australian and New Zealand Fauna.’ (CSIRO Publishing: Melbourne, Vic., Australia)
Bauder, JM, Roberts, NM, Ruid, D, Kohn, B, and Allen, ML (2020). Black bear translocations in response to nuisance behaviour indicate increased effectiveness by translocation distance and landscape context. Wildlife Research 47, 426.
| Black bear translocations in response to nuisance behaviour indicate increased effectiveness by translocation distance and landscape context.Crossref | GoogleScholarGoogle Scholar |
Berger-Tal, O, and Saltz, D (2014). Using the movement patterns of reintroduced animals to improve reintroduction success. Current Zoology 60, 515–526.
| Using the movement patterns of reintroduced animals to improve reintroduction success.Crossref | GoogleScholarGoogle Scholar |
Berman, EE, Coops, NC, Kearney, SP, and Stenhouse, GB (2019). Grizzly bear response to fine spatial and temporal scale spring snow cover in Western Alberta. PLoS One 14, 1–15.
| Grizzly bear response to fine spatial and temporal scale spring snow cover in Western Alberta.Crossref | GoogleScholarGoogle Scholar |
Blanchard, BM, and Knight, RR (1995). Biological consequences of relocating grizzly bears in the Yellowstone ecosystem. Journal of Wildlife Management 59, 560–565.
Boulanger, J, and Stenhouse, GB (2014). The impact of roads on the demography of grizzly bears in Alberta. PLoS One 9, e115535.
| The impact of roads on the demography of grizzly bears in Alberta.Crossref | GoogleScholarGoogle Scholar | 25532035PubMed |
Boulanger, J, Nielsen, SE, and Stenhouse, GB (2018). Using spatial mark-recapture for conservation monitoring of grizzly bear populations in Alberta. Scientific Reports 8, 1–15.
| Using spatial mark-recapture for conservation monitoring of grizzly bear populations in Alberta.Crossref | GoogleScholarGoogle Scholar |
Bourbonnais, ML, Nelson, TA, Cattet, MRL, Darimont, CT, Stenhouse, GB, and Janz, DM (2014). Environmental factors and habitat use influence body condition of individuals in a species at risk, the grizzly bear. Conservation Physiology 2, 1–14.
| Environmental factors and habitat use influence body condition of individuals in a species at risk, the grizzly bear.Crossref | GoogleScholarGoogle Scholar |
Boyce, MS, Vernier, PR, Nielsen, SE, and Schmiegelow, FKA (2002). Evaluating resource selection functions. Ecological Modelling 157, 281–300.
| Evaluating resource selection functions.Crossref | GoogleScholarGoogle Scholar |
Bracis, C, Bildstein, KL, and Mueller, T (2018). Revisitation analysis uncovers spatio-temporal patterns in animal movement data. Ecography 41, 1801–1811.
| Revisitation analysis uncovers spatio-temporal patterns in animal movement data.Crossref | GoogleScholarGoogle Scholar |
Brannon, RD (1987). Nuisance grizzly bears, Ursus arctos, translocations in the greater Yellowstone area. Tha Canadian Feild-Naturalist 101, 569–575.
Burnham KP, Anderson DR (2002) ‘Model selection and multi-model inference: a practical information-theoretic approach.’ 2nd edn. (Spring-Verlag: New York, NY, USA)
Burns, CE (2005). Behavioral ecology of disturbed landscapes: the response of territorial animals to relocation. Behavioral Ecology 16, 898–905.
| Behavioral ecology of disturbed landscapes: the response of territorial animals to relocation.Crossref | GoogleScholarGoogle Scholar |
Calenge, C (2006). The package ‘adehabitat’ for the R software: a tool for the analysis of space and habitat use by animals. Ecological Modelling 197, 516–519.
| The package ‘adehabitat’ for the R software: a tool for the analysis of space and habitat use by animals.Crossref | GoogleScholarGoogle Scholar |
Carnahan, AM, Manen, FT, van Haroldson, MA, Stenhouse, GB, and Robbins, CT (2021). Quantifying energetic costs and defining energy landscapes experienced by grizzly bears. Journal of Experimental Biology , .
| Quantifying energetic costs and defining energy landscapes experienced by grizzly bears.Crossref | GoogleScholarGoogle Scholar |
Cattet, MR, Christison, K, Caulkett, NA, and Stenhouse, GB (2003). Physiologic responses of grizzly bears to different methods of capture. Journal of Wildlife Diseases 39, 649–654.
| Physiologic responses of grizzly bears to different methods of capture.Crossref | GoogleScholarGoogle Scholar | 14567227PubMed |
Cattet, M, Boulanger, J, Stenhouse, G, Powell, RA, and Reynolds-Hogland, MJ (2008). An Evaluation of Long-Term Capture Effects in Ursids: Implications for Wildlife Welfare and Research. Journal of Mammalogy 89, 973–990.
| An Evaluation of Long-Term Capture Effects in Ursids: Implications for Wildlife Welfare and Research.Crossref | GoogleScholarGoogle Scholar |
Champagnon, J, Elmberg, J, Guillemain, M, Gauthier-Clerc, M, and Lebreton, J-D (2012). Conspecifics can be aliens too: a review of effects of restocking practices in vertebrates. Journal for Nature Conservation 20, 231–241.
| Conspecifics can be aliens too: a review of effects of restocking practices in vertebrates.Crossref | GoogleScholarGoogle Scholar |
Clark, J, Huber, D, and Servheen, C (2002). Bear Reintroductions: Lessons and Challenges. Ursus 13, 335–345.
| Bear Reintroductions: Lessons and Challenges.Crossref | GoogleScholarGoogle Scholar |
Coogan, SC, Nielsen, SE, and Stenhouse, GB (2012). Spatial and temporal heterogeneity creates a ‘brown tide’ in root phenology and nutrition. International Scholarly Research Network 2012, 1–10.
Dice, LR (1931). The Relation of Mammalian Distribution to Vegetation Types. The Scientific Monthly 33, 312–317.
Dubois, S, Fenwick, N, Ryan, EA, Baker, L, Baker, SE, Beausoleil, NJ, Carter, S, Cartwright, B, Costa, F, Draper, C, Griffin, J, Grogan, A, Howald, G, Jones, B, Littin, KE, Lombard, AT, Mellor, DJ, Ramp, D, Schuppli, CA, and Fraser, D (2017). International consensus principles for ethical wildlife control. Conservation Biology 31, 753–760.
| International consensus principles for ethical wildlife control.Crossref | GoogleScholarGoogle Scholar | 28092422PubMed |
Festa-Bianchet M (2010) ‘Status of the Grizzly Bear (Ursus arctos) in Alberta: Update 2010.’ (Alberta Sustainable Resource Development: Edmonton, AB, Canada)
Fischer, J, and Lindenmayer, DB (2000). An assessment of the published results of animal relocations. Biological Conservation 96, 1–11.
| An assessment of the published results of animal relocations.Crossref | GoogleScholarGoogle Scholar |
Gaynor, KM, Hojnowski, CE, Carter, NH, and Brashares, JS (2018). The influence of human disturbance on wildlife nocturnality. Science 360, 1232–1235.
| The influence of human disturbance on wildlife nocturnality.Crossref | GoogleScholarGoogle Scholar | 29903973PubMed |
Frair, Jl, Merrill, EH, Allen, JR, and Boyce, MS (2007). Know Thy Enemy: Experience Affects Elk Translocation Success in Risky Landscapes. Journal of Wildlife Management 71, 541–554.
| Know Thy Enemy: Experience Affects Elk Translocation Success in Risky Landscapes.Crossref | GoogleScholarGoogle Scholar |
Graham, K, and Stenhouse, GB (2014). Home range, movements, and denning chronology of the Grizzly Bear (Ursus arctos) in west-central Alberta. Canadian Field-Naturalist 128, 223–234.
| Home range, movements, and denning chronology of the Grizzly Bear (Ursus arctos) in west-central Alberta.Crossref | GoogleScholarGoogle Scholar |
Gusset, M, Swarner, MJ, Mponwane, L, Keletile, K, and McNutt, JW (2009). Human–wildlife conflict in northern Botswana: Livestock predation by endangered African wild dog lycaon pictus and other carnivores. Oryx 43, 67–72.
| Human–wildlife conflict in northern Botswana: Livestock predation by endangered African wild dog lycaon pictus and other carnivores.Crossref | GoogleScholarGoogle Scholar |
Hayward, MW, Adendorff, J, Moolman, L, Hayward, GJ, and Kerley, GIH (2006). The successful reintroduction of leopard Panthera pardus to the Addo Elephant National Park. African Journal of Ecology 45, 103–104.
| The successful reintroduction of leopard Panthera pardus to the Addo Elephant National Park.Crossref | GoogleScholarGoogle Scholar |
Hertel, AG, Zedrosser, A, Mysterud, A, Støen, O-G, Steyaert, SMJG, and Swenson, JE (2016). Temporal effects of hunting on foraging behavior of an apex predator: Do bears forego foraging when risk is high? Oecologia 182, 1019–1029.
| Temporal effects of hunting on foraging behavior of an apex predator: Do bears forego foraging when risk is high?Crossref | GoogleScholarGoogle Scholar | 27696003PubMed |
Hertel, AG, Leclerc, M, Warren, D, Pelletier, F, Zedrosser, A, and Mueller, T (2019). Don’t poke the bear: using tracking data to quantify behavioural syndromes in elusive wildlife. Animal Behaviour 147, 91–104.
| Don’t poke the bear: using tracking data to quantify behavioural syndromes in elusive wildlife.Crossref | GoogleScholarGoogle Scholar |
Hoogenboom, I, Daan, S, Dallinga, JH, and Schoenmakers, M (1984). Seasonal change in the daily timing of behaviour of the common vole, Microtus arvalis. Oecologia 61, 18–31.
| Seasonal change in the daily timing of behaviour of the common vole, Microtus arvalis.Crossref | GoogleScholarGoogle Scholar | 28311381PubMed |
Kearney, SP, Coops, NC, Stenhouse, GB, Nielsen, SE, Hermosilla, T, White, JC, and Wulder, MA (2019). Grizzly bear selection of recently harvested forests is dependent on forest recovery rate and landscape composition. Forest Ecology and Management 449, 117459.
| Grizzly bear selection of recently harvested forests is dependent on forest recovery rate and landscape composition.Crossref | GoogleScholarGoogle Scholar |
Landriault, LJ, Brown, GS, Hamr, J, and Mallory, FF (2009). Age, sex and relocation distance as predictors of return for relocated nuisance black bears Ursus americanus in Ontario, Canada. Wildlife Biology 15, 155–164.
| Age, sex and relocation distance as predictors of return for relocated nuisance black bears Ursus americanus in Ontario, Canada.Crossref | GoogleScholarGoogle Scholar |
Larsen, TA, Nielsen, SE, Cranston, J, and Stenhouse, GB (2019). Do remnant retention patches and forest edges increase grizzly bear food supply? Forest Ecology and Management 433, 741–761.
| Do remnant retention patches and forest edges increase grizzly bear food supply?Crossref | GoogleScholarGoogle Scholar |
Leclerc, M, Vander, E, Zedrosser, A, Swenson, JE, Kindberg, J, and Pelletier, F (2016). Quantifying consistent individual differences in habitat selection. Oecologia 180, 697–705.
| Quantifying consistent individual differences in habitat selection.Crossref | GoogleScholarGoogle Scholar | 26597548PubMed |
Lewis, JH, Alldredge, MW, Dreher, BP, George, JL, Wait, S, Petch, B, and Runge, JP (2019). Summarizing Colorado’s black bear two‐strike directive 30 years after inception. Wildlife Society Bulletin 43, 599–607.
| Summarizing Colorado’s black bear two‐strike directive 30 years after inception.Crossref | GoogleScholarGoogle Scholar |
Linnell, J, Odden, J, and Smith, M (1999). Large carnivores that kill livestock: do ‘problem individuals’ really exist? Wildlife Society Bulletin 27, 698–705.
Massei, G, Quy, RJ, Gurney, J, and Cowan, DP (2010). Can translocations be used to mitigate human - wildlife conflicts? Wildlife Research 37, 428.
| Can translocations be used to mitigate human - wildlife conflicts?Crossref | GoogleScholarGoogle Scholar |
McClelland, CJR, Coops, NC, Kearney, SP, Burton, AC, Nielsen, SE, and Stenhouse, GB (2020). Variations in grizzly bear habitat selection in relation to the daily and seasonal availability of annual plant-food resources. Ecological Informatics 58, 101116.
| Variations in grizzly bear habitat selection in relation to the daily and seasonal availability of annual plant-food resources.Crossref | GoogleScholarGoogle Scholar |
McLellan, BN, and Shackleton, DM (1989). Grizzly bears and resource-extraction industries: habitat displacement in response to seismic exploration, timber harvesting and road maintenance. Journal of Applied Ecology 26, 371–380.
| Grizzly bears and resource-extraction industries: habitat displacement in response to seismic exploration, timber harvesting and road maintenance.Crossref | GoogleScholarGoogle Scholar |
Miller, B, Ralls, K, Reading, RP, Scott, JM, and Estes, J (1999). Biological and technical considerations of carnivore translocation: a review. Animal Conservation 2, 59–68.
| Biological and technical considerations of carnivore translocation: a review.Crossref | GoogleScholarGoogle Scholar |
Milligan, S, Brown, L, Hobson, D, Frame, P, and Stenhouse, G (2018). Factors affecting the success of grizzly bear translocations. The Journal of Wildlife Management 82, 519–530.
| Factors affecting the success of grizzly bear translocations.Crossref | GoogleScholarGoogle Scholar |
Minteer, BA, and Collins, JP (2010). Move it or lose it? The ecological ethics of relocating species under climate change. Ecological Applications 20, 1801–1804.
| Move it or lose it? The ecological ethics of relocating species under climate change.Crossref | GoogleScholarGoogle Scholar | 21049870PubMed |
Morehouse, AT, and Boyce, MS (2017). Troublemaking carnivores: conflicts with humans in a diverse assemblage of large carnivores. Ecology and Society 22, art4.
| Troublemaking carnivores: conflicts with humans in a diverse assemblage of large carnivores.Crossref | GoogleScholarGoogle Scholar |
Morehouse, AT, Graves, TA, Mikle, N, and Boyce, MS (2016). Nature vs. nurture: evidence for social learning of conflict behaviour in grizzly bears. PLoS ONE 11, e0165425.
| Nature vs. nurture: evidence for social learning of conflict behaviour in grizzly bears.Crossref | GoogleScholarGoogle Scholar | 27851753PubMed |
Munro, RHM, Nielsen, SE, Price, MH, Stenhouse, GB, and Boyce, MS (2006). Seasonal and diel patterns of grizzly bear diet and activity in west-central Alberta. Journal of Mammalogy 87, 1112–1121.
| Seasonal and diel patterns of grizzly bear diet and activity in west-central Alberta.Crossref | GoogleScholarGoogle Scholar |
Murray, MH, Fassina, S, Hopkins, JB, Whittington, J, and St Clair, CC (2017). Seasonal and individual variation in the use of rail-associated food attractants by grizzly bears (Ursus arctos) in a national park. PloS One 12, e0175658.
| Seasonal and individual variation in the use of rail-associated food attractants by grizzly bears (Ursus arctos) in a national park.Crossref | GoogleScholarGoogle Scholar | 28542218PubMed |
National Research Council of Canada (2015) Sunrise/sunset calculator with advanced options and sun angles. Available at http://www.nrc-cnrc.gc.ca/eng/services/sunrise/advanced.html
Nichols, JD, and Williams, BK (2006). Monitoring for conservation. Trends in Ecology and Evolution 21, 668–673.
| Monitoring for conservation.Crossref | GoogleScholarGoogle Scholar | 16919361PubMed |
Nielsen, SE, Boyce, MS, and Stenhouse, GB (2004a). Grizzly bears and forestry I: selection of clearcuts by grizzly bears in west-central Alberta, Canada. Forest Ecology and Management 199, 51–65.
Nielsen, SE, Herrero, S, Boyce, MS, Mace, RD, Benn, B, Gibeau, ML, and Jevons, S (2004b). Modelling the spatial distribution of human-caused grizzly bear mortalities in the Central Rockies ecosystem of Canada. Biological Conservation 120, 101–113.
| Modelling the spatial distribution of human-caused grizzly bear mortalities in the Central Rockies ecosystem of Canada.Crossref | GoogleScholarGoogle Scholar |
Nielsen, SE, Munro, RHM, Bainbridge, EL, Stenhouse, GB, and Boyce, MS (2004c). Grizzly bears and forestry II. Distribution of grizzly bear foods in clearcuts of west-central Alberta, Canada. Forest Ecology and Management 199, 67–82.
| Grizzly bears and forestry II. Distribution of grizzly bear foods in clearcuts of west-central Alberta, Canada.Crossref | GoogleScholarGoogle Scholar |
Nielsen, SE, McDermid, G, Stenhouse, GB, and Boyce, MS (2010). Dynamic wildlife habitat models: seasonal foods and mortality risk predict occupancy-abundance and habitat selection in grizzly bears. Biological Conservation 143, 1623–1634.
Nielsen, SE, Shafer, ABA, Boyce, MS, and Stenhouse, GB (2013). Does learning or instinct shape habitat selection? PLoS One 8, 1–5.
Northrup, JM, Pitt, J, Muhly, TB, Stenhouse, GB, Musiani, M, and Boyce, MS (2012). Vehicle traffic shapes grizzly bear behaviour on a multiple-use landscape. Journal of Applied Ecology 49, 1159–1167.
Ordiz, A, Kindberg, J, Sæbø, S, Swenson, JE, and Støen, O-G (2014). Brown bear circadian behavior reveals human environmental encroachment. Biological Conservation 173, 1–9.
| Brown bear circadian behavior reveals human environmental encroachment.Crossref | GoogleScholarGoogle Scholar |
Pinter-Wollman, N, Isbell, LA, and Hart, LA (2009). Assessing translocation outcome: comparing behavioral and physiological aspects of translocated and resident African elephants (Loxodonta africana). Biological Conservation 142, 1116–1124.
| Assessing translocation outcome: comparing behavioral and physiological aspects of translocated and resident African elephants (Loxodonta africana).Crossref | GoogleScholarGoogle Scholar |
Proctor, MF, McLellan, BN, Stenhouse, GB, Mowat, G, Lamb, CT, and Boyce, MS (2020). Effects of roads and motorized human access on grizzly bear populations in British Columbia and Alberta, Canada. Ursus 2019, 16.
| Effects of roads and motorized human access on grizzly bear populations in British Columbia and Alberta, Canada.Crossref | GoogleScholarGoogle Scholar |
Réale, D, Reader, SM, Sol, D, McDougall, PT, and Dingemanse, NJ (2007). Integrating animal temperament within ecology and evolution. Biological Reviews 82, 291–318.
| Integrating animal temperament within ecology and evolution.Crossref | GoogleScholarGoogle Scholar | 17437562PubMed |
Riley, SJ, Aune, K, Mace, RD, and Madel, MJ (1994). Translocation of Nuisance Grizzly Bears in Northwestern Montana. Bears: Their Biology and Management 9, 567.
| Translocation of Nuisance Grizzly Bears in Northwestern Montana.Crossref | GoogleScholarGoogle Scholar |
Ripple, WJ, Estes, JA, Beschta, RL, Wilmers, CC, Ritchie, EG, Hebblewhite, M, Berger, J, Elmhagen, B, Letnic, M, Nelson, MP, Schmitz, OJ, Smith, DW, Wallach, AD, and Wirsing, AJ (2014). Status and ecological effects of the world’s largest carnivores. Science 343, 1241484.
| Status and ecological effects of the world’s largest carnivores.Crossref | GoogleScholarGoogle Scholar | 24408439PubMed |
Roever, CL, Boyce, MS, and Stenhouse, GB (2008). Grizzly bears and forestry II: grizzly bear habitat selection and conflicts with road placement. Forest Ecology and Management 256, 1262–1269.
| Grizzly bears and forestry II: grizzly bear habitat selection and conflicts with road placement.Crossref | GoogleScholarGoogle Scholar |
Schwartz, CC, Harris, RB, and Haroldson, MA (2003). Temporal, spatial, and environmental influences on the demographics of grizzly bears in the Greater Yellowstone ecosystem. Wildlife Monographs 161, 1–68.
Seddon PJ, Strauss WM, Innes J (2012) Animal translocations: what are they and why do we do them? In ‘Reintroduction Biology: Integrating Science and Management’, 1st edn. (Eds JG Ewen, DP Armstrong, KA Parker, PJ Seddon) pp. 1–32. (Wiley-Blackwell: West Sussex, UK)
Spencer, RD, Beausoleil, RA, and Martorello, DA (2007). How agencies respond to human–black bear conflicts: a survey of wildlife agencies in North America. Ursus 18, 217–229.
| How agencies respond to human–black bear conflicts: a survey of wildlife agencies in North America.Crossref | GoogleScholarGoogle Scholar |
Stamps, JA, and Swaisgood, RR (2007). Someplace like home: experience, habitat selection and conservation biology. Applied Animal Behaviour Science 102, 392–409.
| Someplace like home: experience, habitat selection and conservation biology.Crossref | GoogleScholarGoogle Scholar |
Stuparyk, B, Horn, CJ, Karabatsos, S, and Torres, JA (2018). A Meta-analysis of Animal Survival Following Translocation: Comparisons Between Conflict and Conservation Efforts. Canadian Wildlife Biology and Management 7, 3–17.
Treves, A, and Karanth, KU (2003). Human-Carnivore Conflict and Perspectives on Carnivore Management Worldwide. Conservation Biology 17, 1491–1499.
| Human-Carnivore Conflict and Perspectives on Carnivore Management Worldwide.Crossref | GoogleScholarGoogle Scholar |
Wood SN (2017) ‘Generalized Additive Models: An Introduction with R.’ 2nd edn. (CRC Press: Boca Raton, FL, USA)
Zuur, AF, Ieno, EN, and Elphick, CS (2010). A protocol for data exploration to avoid common statistical problems. Methods in Ecology and Evolution 1, 3–14.
| A protocol for data exploration to avoid common statistical problems.Crossref | GoogleScholarGoogle Scholar |