

Increasing fire severity negatively affects greater glider density
Jarrah C. May-Stubbles A , Ana Gracanin A and Katarina M. Mikac A *
A and Katarina M. Mikac A *
A Centre for Sustainable Ecosystem Solutions, School of Earth, Atmospheric and Life Sciences, University of Wollongong, Northfields Road, NSW 2522, Australia.
Wildlife Research 49(8) 709-718 https://doi.org/10.1071/WR21091
Submitted: 17 June 2021 Accepted: 14 March 2022 Published: 18 May 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Fire severity and frequency is predicted to increase over the remainder of the 21st century in Australia’s temperate forests; therefore, the effects of fire severity on forest-dependant species is of major conservation concern.
Aims: Determining the short-term (<1 year) effect of fire severity on a forest-dependant species, the greater glider, Petauroides volans, in Monga National Park, south-eastern New South Wales.
Methods: Three fire-severity classes were investigated, namely, low (canopy unburnt), moderate (partial canopy burn) and high (complete canopy consumption). Fifteen randomly allocated sites were visited in total, with five sites in each fire-severity class being visited on three separate occasions. Spotlighting of two off-track transects per site was completed over a 4-week period by using multi-covariate distance sampling (MCDS) to estimate greater glider densities at each fire-severity class.
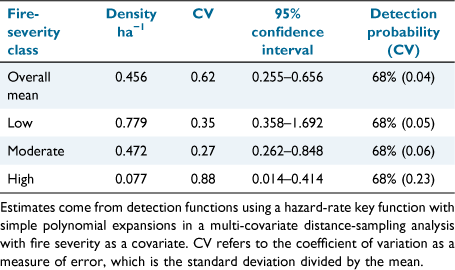
Key results: An overall mean greater glider density estimate of 0.456 ha−1 (95% confidence interval (CI) 0.256–0.654 ha−1) was calculated across all sites in Monga National Park. Density estimates varied among fire-severity classes; low 0.779 ha−1 (95% CI 0.358–1.692 ha−1), moderate 0.472 ha−1 (95% CI 0.262–0.848 ha−1), and high 0.077 ha−1 (95% CI 0.0142–0.414 ha−1).
Conclusions: Distance sampling estimates suggested that areas affected by high-severity fire have lower densities of greater gliders.
Implications: The importance of areas experiencing low fire severity as a source of greater glider refugia during wildfire events is evident. Under a changing climate, frequent major wildfire events may significantly affect population viability of greater gliders if insufficient time between fires allow for population recovery.
Keywords: arboreal, disturbance, distance sampling, gliding possum, hollow obligate, Petauroides volans, spotlighting, tree‐dwelling.
Introduction
Wildfire events have adverse effects on Australia’s forest-dependant arboreal mammals either through direct mortality (Bradstock et al. 2002), or indirect mortality through the alteration of suitable habitat (Eyre 2005; Lunney et al. 2007). Heterogeneity in burn severity across a landscape can occur during large-scale wildfires owing to habitat patches of different moisture content, vegetation types, and varying topography burning at different temperatures (Bradstock et al. 2005). Spatial heterogeneity results in different levels of fire severity across a landscape and thus allows for potential fire-habitat refuges to develop (Bradstock et al. 2005). It is from fire refuges, areas that are unburnt or have experienced low-severity burns, that species can survive and eventually repopulate areas that experienced moderate- to high-severity fire (Berry et al. 2015, 2016).
Changes in fire regimes is a recognised threat to Australian arboreal mammals (Woinarski et al. 2014). Differences in fire severity play a major role in determining post-fire survivorship and habitat use for arboreal mammals (Banks et al. 2011). This was investigated by Chia et al. (2015) who assessed changes in arboreal mammal populations, finding a negative correlation between arboreal mammal abundance and increasing fire severity. These trends are likely to be attributed to increased mortality and decreased resource availability at severely burnt sites (Lindenmayer et al. 2013; Chia et al. 2015). On the contrary, Banks et al. (2011) found that the 2009 Black Saturday fire in Victoria led to no mortality in 13 radio-tracked mountain brushtail possums. Instead, the authors attributed a reduction in possum abundance in high-severity sites, to an 80% reduction in hollow-bearing trees, a critical habitat feature for many arboreal mammals (Lindenmayer et al. 1990). Determining specific forest-dependant species responses to different levels of fire severity is fundamental for successful future conservation efforts and data-driven fire-management practices (Wintle et al. 2020).
The greater glider (Petauroides volans) is listed as vulnerable under the Australian Environment Protection and Biodiversity Conservation Act 1999 (EPBC Act). This arboreal mammal is particularly at risk from changes in fire regimes (Chia et al. 2015; Lindenmayer et al. 2020). Anthropogenic-driven climate change has increased global temperatures and has led to longer droughts in Australia (Hennessy et al. 2005; Abram et al. 2021; Nolan et al. 2021). Thus, fires are predicted to become more intense and frequent, reducing the ability of ecosystems to recover, which then threatens the ability of greater gliders to persist (Woinarski et al. 2015; Ward et al. 2020). Greater gliders are Australia’s largest gliding folivore, with three species being found in eucalypt forests along Australia’s eastern coast from central Victoria to central Queensland (McGregor et al. 2020). Classified as obligate hollow users, greater gliders utilise cavities in hollow-bearing trees for shelter, predator avoidance, and as den sites for raising young (Lindenmayer et al. 2005; Goldingay 2012). Because of their large size (from 900 to 1700 g), larger hollow entrances are required (Goldingay 2012). Hollows of this size typically require trees with a diameter at breast height >70 cm and a significant time (>100 years) for hollows to form through natural decay (Gibbons et al. 2000; Goldingay 2012). Lindenmayer et al. (1990) found that forest age and the abundance of hollow-bearing trees positively correlate to greater glider abundance. However, wildfires pose a significant risk to old-growth forests and the persistence of hollow-bearing trees (Banks et al. 2011). Furthermore, the species is particularly susceptible to wildfire because greater gliders exhibit high site fidelity, have slow reproduction and are slow dispersers (McLean et al. 2018). These changes in resource availability, combined with other habitat-altering processes, such as selective logging and land clearing, place major strain on remaining greater glider populations (McLean et al. 2018).
Providing accurate baseline estimates of population size is fundamental in all forms of wildlife management, especially for managing at-risk populations (Sutherland et al. 2004; deTores and Elscot 2010; Finlayson et al. 2010). Line-transect distance sampling is a method that provides robust and repeatable estimate of arboreal mammal populations (deTores and Elscot 2010; Emerson et al. 2019; Crowther et al. 2020). Line-transect distance sampling uses the perpendicular distance of animals observed from the transect line to account for animal detectability, whereby a detection function is fitted which assumes that detectability will decrease as distance from the transect line increases (Buckland et al. 2001). It also assumes that animals do not move in response to the observer and that animals on the transect line are detected with certainty (Thomas et al. 2010). Emerson et al. (2019) compared the use of distance sampling with strip-width sampling for estimating the density of greater gliders in Australian eucalypt forests. The direct comparison of the two methods showed that estimates from strip-widths were less robust than were distance-sampling estimates. Despite its potential usefulness in estimating greater glider populations, distance sampling has not been used to assess populations at differing levels of fire severity.
Understanding the effect that large-scale wildfires have on greater glider population size is of utmost importance to their conservation management. Our study aimed to investigate the greater glider population at Monga National Park, in south-eastern New South Wales (NSW), within a year since the large-scale 2019–2020 Currowan and Clyde Mountain wildfires. More specifically, this study (1) investigated the density of greater gliders at our study site, (2) compared the densities of greater gliders from different fire-severity classes and (3) reported on the availability of habitat hollows at different fire-severity classes in Monga National Park, NSW.
Materials and methods
Study area
The study was conducted in Monga National Park (35′39′47′S, 149′58′16E) and a contiguous patch of forest adjacent to the Budawang National Park in the Southern Tablelands of NSW, situated 230 km south-west of Sydney (Fig. 1). It has an area of 25 144 ha extending across the Great Dividing Range south of Clyde Mountain, with an elevation range from 160 to 1070 m. The maximum temperatures vary between 12.4°C (July) and 27.1°C (February), and minimum temperatures range between 0.1°C (July) and 12.3°C (February; Australian Bureau of Meteorology 2020; Station 069132). Monthly mean rainfall ranges from 34.6 mm (July) to 69.3 mm (February). Historically, the park was managed as a state forest before being gazetted as a national park in 2001 (Stellar 2005). Thus, some areas of Monga National Park experienced historical logging as part of harvesting operations.

|
Vegetation is dominated by wet and dry sclerophyll forests of eucalyptus stands including brown barrel (Eucalyptus fastigata), ribbon gum (Eucalyptus viminalis), monkey gum (Eucalyptus cypellocarpa) and white ash (Eucalyptus fraxinoides), interspersed with rainforest patches of plumwood (Eucryphia moorei), rough tree fern (Cyathea australis) and soft tree fern (Antartica dicksonia). The area represents suitable greater glider habitat, with detections since 1995 across the extent of Monga National Park (Atlas of Living Australia 2021).
Fire-severity classification
Since 1938, there have been 66 fires in Monga National Park (MNP); of these, 47 were prescribed burns and 19 were wildfires (Mikac et al. 2021). During the Australian mega-fire event of 2019/2020 (Collins et al. 2021), the Currowan fire burnt Monga National Park from 26 November 2019 to 9 February 2020. Two large-scale wildfires burnt from west to east in the park, leading to a mosaic of differing fire severities. In total, ~98% of Monga National Park was burnt to some degree, with 87% of burns causing partial or complete canopy consumption (Mikac et al. 2021). Fire-severity classification was grouped into three discrete classes based on Fire Extent Severity Mapping (FESMv2.1) data layers and the level of canopy burn (Department of Planning, Environment, and Industry 2020). The three classes were low (canopy unburnt/minor understory burning), moderate (canopy partially burnt/understorey burnt) and high (canopy completely consumed). Ground-truthing was completed at sites to ensure FESM layers accurately assigned the fire-severity classes witnessed in the field at the time of surveys.
Survey design
Fifteen sites were visited in total, with five sites in each of the three fire-severity classes. All sites were randomly selected from a large pool of candidate sites across MNP. Random allocation of sites was completed by dividing the study area into 2 km ×2 km grid squares; from this grid, 40 candidate sites were randomly selected (Mikac et al. 2021). The 40 sites were then assigned to ‘low’, ‘moderate’, and ‘high’ fire-severity classes on the basis of FESM data. From candidate sites, five sites were randomly selected for each fire-severity class.
Within each site, two 250 m transects were established. Placement of the two transects occurred within the 2 km × 2 km grid squares around sites. Transects within sites were parallel and 200–250 m apart (for one site, transects were 150 m apart due to steep topography). Transect placement was constrained by the realised fire-severity level in the area, on the basis of FESM and ground-truthing. Transect ground-truthing involved walking transect lines during day and confirming that the level of canopy burn was kept constant as far as visibly possible. This ground-truthing ensured that all sightings were made within the chosen fire-severity class. An additional constraint of transect placement was the need for off-track transects and safe accessibility; this ensured that transect lines were placed without the inherent biases associated with surveying arboreal mammals along tracks and roads (Emerson et al. 2019). Transects were therefore inclusive of a variety of topographic regions, including ridgetops, mid-slopes, and gullies (Supplementary Appendix 1). After transects were assigned, they were marked out using reflective flagging tape to aid in subsequent night-time navigation for arboreal mammal surveys.
Arboreal-mammal surveys
Spotlight surveys were completed from 28 September to 4 November 2020 to estimate greater glider densities. Three repeat visits were made to each transect, occurring, on average, 9 days apart (maximum = 17, minimum = 5), leading to a total of 90 transect surveys over the survey period. Surveys were conducted on nights with suitable conditions (i.e. fine to partly cloudy nights); nights with heavy wind, rain or fog were avoided because greater gliders are less likely to be detected during these conditions (Davey 1990; Laurance 1990; Wintle et al. 2005).
Spotlight surveys involved two observers walking transects at a speed of 12 m min–1, and commenced no earlier than ~10 min after last light, to coincide with the time when nocturnal arboreal mammals become active (Davey 1990). Each observer (separated approximately by 20 m along the transect line) made thorough searches of the canopy, mid-storey and understorey from the transect line to an unrestricted distance (Crowther et al. 2020). Furthermore, to ensure the distance-sampling assumption of 100% detectability at 0 m from the transect line, each observer thoroughly searched the transect line (Buckland et al. 2001; Thomas et al. 2010). Observers were equipped with 1000 lumen Led Lenser H7R Work headlamps (Solingen, North Rhine-Westphalia, Germany), with a 250 m visual line of sight to detect arboreal mammals. An 8 × 40 pair of binoculars (Nikon Action 8.2) were used to identify species, with observations then being confirmed by both observers to ensure correct species identification.
Greater gliders were easily identified from other arboreal mammals on the basis of their unique morphology; including bright eye-shine, long tail and tendency to stare in the direction of the observer when detected (Lindenmayer et al. 2001; Wintle et al. 2005). The exact perpendicular distance of each sighting from the transect line was recorded using a Nikon Forestry Pro 2 Laser Rangefinder (Nikon, Tokyo, Japan), and GPS location was recorded using the IOS app AVENZA MAPS (v. 3.7).
Habitat hollow counts
Habitat hollow counts were made by thoroughly searching all trees, with the aid of binoculars, within 50 m from transects during the day to estimate a relative abundance of suitable hollows. Effort was made to search each tree from all sides within the 50 m strips. Hollow entrance sizes were estimated from the ground, with hollow diameter classified into one of the following four size classes: 0–5, 5–10, 10–15, >15 cm. This was performed by the same observer to ensure consistency among sites (Harper et al. 2004). Habitat hollows were characterised by any hollow >15 cm in diameter based on greater glider hollow-entrance size preferences from Goldingay (2012). Hollow counts are an estimate of relative hollow availability, because counts from the ground cannot confirm whether hollows perceived are true hollows (Koch et al. 2008). In many cases, several habitat hollows were counted from an individual old-growth hollow-bearing tree.
Density estimates
The density of greater gliders at each fire-severity class was estimated using a multi-covariate distance-sampling (MCDS) approach outlined by Marques et al. (2007). The perpendicular distances collected during spotlight surveys were used for analysis in DISTANCE v.7.3 (Thomas et al. 2010). A histogram of the recorded distance frequencies was produced over the entire dataset and analysed to determine whether the shape criterion was satisfied (Thomas et al. 2010). This shape criterion specifies that the histogram must have perfect detection on and near the transect line, with a shoulder that extends to the point where detectability starts to drop off sharply (Buckland et al. 2001). Although the data showed irregularities and spikes between 10 and 40 m, the detection function produced a shoulder of 100% detectability up to 45 m. The dataset was truncated at 90 m because meaningful observations were being made up to this point and to remove two large outlier distances from the dataset. Furthermore, truncation was recommended to assist in model fitting because long-tailed detection functions required more model parameters, which increased variance (Thomas et al. 2010; Crowther et al. 2020).
Further DISTANCE analysis involved stratifying the truncated dataset by fire-severity classes. Stratifying the dataset produced density estimates for each class, while producing an estimated overall mean density (Thomas et al. 2010). Differences in density estimates for the three fire-severity classes were estimated using the following two approaches: (1) a conventional distance-sampling (CDS) technique where separate detection functions were applied to each severity class; and (2) an MCDS technique where a shared detection function was used to estimate between fire-severity classes. Akaike’s information criterion (AIC) values were used to select the analysis approach that had the best fit (Table 1). This was considered suitable because fire severity had only a slight effect on detectability, and the small number of observations in high-severity sites made fitting separate detection functions difficult.

|
Model selection was based on the stratified data, and the best model was chosen from the suite of recommended models (Table 1) for CDS and MCDS (Thomas et al. 2010; Emerson et al. 2019). Model performance was assessed using AIC values, distance frequency histograms to determine model fit, and the Cramer–von Mises goodness-of-fit (GOF) tests. On the basis of the lowest AIC value and an acceptable GOF, the MCDS hazard-rate model with simple polynomial expansions was chosen for estimating densities in each fire-severity class (Buckland et al. 2001; Thomas et al. 2010).
Three repeat visits to each transect were conducted, on separate nights, to increase survey effort in sites without violating the independence of observation assumption in DISTANCE, as recommended by Thomas et al. (2010). Effectively, three repeat visits to a 250 m transect led to a 750 m survey effort per transect.
Effects of fire severity on habitat hollows
The effect of fire severity on habitat hollow counts was tested using a one-way ANOVA in R studio. Further analysis involved using Tukey–Kramer post hoc pairwise comparison tests to determine significant differences among fire-severity classes (Tukey adjusted P-values, α < 0.05). Packages used in all R studio analysis were ‘tidyverse’ (Wickham et al. 2019) and ‘ggplot2’ (Wickham 2016).
Results
The total distance of transects surveyed was 22 500 m, over 33 h and 23 nights of spotlighting effort. Greater gliders were recorded at 9 of 15 (60%) sites. In total, 122 greater glider observations were made, with 58% being in low-severity sites, 36% in moderate, and 6% in high (Fig. 2). Greater gliders were spotted at a mean distance of 32 m (±s.e. 1.9), with the furthest detection at 130 m. Mean distances did not differ much for low (32 m ±s.e. 2.3) or moderate (31.8 m ±s.e. 3.6) sites; however, they were slightly larger for high-severity sites (34.1 m ±s.e. 8.5).

|
Density estimates
Distance sampling estimated the mean density of gliders across sites in Monga National Park as 0.456 ha−1 (CI 0.256–0.654; Table 2). Within the fire-severity classes, low produced the largest density estimate at 0.779 ha−1 (CI 0.344–1.692), followed by moderate 0.472 ha−1 (CI 0.262–0.848; Table 2), with high-severity sites producing the lowest density estimate at 0.077 ha−1 (CI 0.014–0.414). The averaged detection probability across our sites (68%) indicated a high detectability of greater gliders in MNP. Detection data also indicated that detectability along the transect line was an issue, in general, across the study site (Fig. 3a), particularly within low fire-severity sites (Fig. 3b), suggesting that the assumption of perfect detectability along the transect line was not satisfied and that density estimates are potentially slight underestimations.
|

|
Relative abundance of habitat hollows
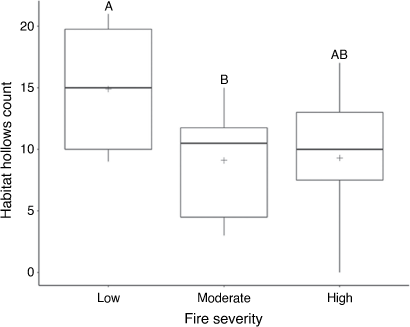
The number of habitat hollows per transect differed significantly among fire-severity classes (F2,27 = 4.0997, P < 0.05). Overall, more habitat hollows were found in low fire-severity sites than in moderate and high sites (Fig. 4). A post hoc Tukey HSD comparison confirmed this, showing a significant difference between low and moderate sites, but not between low and high sites (Fig. 4).
|
Discussion
Distance sampling was used to produce preliminary estimates of a greater glider density across surveyed sites in MNP, with differences in density being identified among fire-severity classes. Fire severity class was also linked to the relative abundance of habitat hollows available.
Density estimates
Distance sampling showed an overall mean density estimate of 0.46 ha−1 (95% CI 0.26–0.65 ha−1) greater gliders across MNP, on the basis of differences in fire-severity classes. The only other studies using distance sample for estimating greater glider density were those of Kavanagh (1984), Emerson et al. (2019) and Cripps et al. (2021). All studies were conducted in open eucalypt forests, with Kavanagh (1984) estimating densities at 0.3–1.2 ha−1 in wet eucalypt forests. Emerson et al. (2019) used an off-track conventional distance-sampling method to estimate densities of 1.36 ha−1 (95% CI 1.07–1.72) in north-eastern NSW. Recently, Cripps et al. (2021) estimated greater glider density of 0.959 ha−1 (95% CI 0.604–1.495) in open eucalypt forests in north-eastern Victoria by using a mark–recapture distance-sampling method. Overall, mean estimates from our study suggested a low-density greater glider population from MNP, with greater glider density ranging from 0.01 to 5 individuals per hectare (Henry 1984; Kavanagh 1984; Kehl and Borsboom 1984; van Der Ree et al. 2004; Vinson et al. 2021). However, it is important to note that there was deviation from the 100% detectability at 0 m distance assumption in low fire-severity sites (Fig. 3b). Thus, our density estimates are likely to be slightly underestimated because of non-perfect detection on the transect line. This issue was discussed by Emerson et al. (2019) as resulting from either surveying biases, failing to detect an animal available for detection, or from detectability biases, whereby detection is obscured by vegetation. The latter is likely to be the case, because, in this study, searches along the transect line were thorough, with detections along the transect line being obscured because of the higher levels of strata complexity and canopy cover found within low-burnt sites (Catling et al. 2002). Recent improvements to account for imperfect detection on the transect line have been made by Cripps et al. (2021), who used a mark–recapture distance-sampling method with two observers to overcome detectability issues. Their method is recommended as the preferred method for estimating population size of greater gliders because it accounts for issues in detectability (Cripps et al. 2021).
The low-density of greater gliders found at MNP is characteristic of the whole park, with all fire-severity classes producing density estimates <0.8 ha−1 (Table 2). Similar low-density populations have been recorded throughout the Eurobodalla Local Government Area, which encompasses the eastern side of MNP, classifying the greater glider population as endangered prior to the 2019–2020 wildfires. Lindenmayer et al. (2013) suggested that there are further negative impacts of fire even at unburnt sites because of landscape effects, where hollow loss in the surrounding landscape may lead to population declines over time.
Fire severity
For fire-severity classes, distance-sampling analysis showed differences in greater glider density (Table 2). Densities were higher in sites where canopy was unburnt to partially scorched (low/moderate sites) than in areas where canopy was consumed (high-severity sites), exhibiting close to zero greater gliders per hectare (Table 2). It is likely that greater gliders could have experienced higher rates of mortality in sites that experienced high-severity fire (Lindenmayer et al. 2013). Mortality is likely to have occurred, to a lesser extent, within moderate sites, with gliders potentially moving during or soon after the fire to low-severity sites that acted as refugia, such as moist gullies (Catling 1991). Inversely, it is likely that mortality was low in sites with low fire severity.
Greater gliders were observed only at one high-severity site (BTC37), with a total of seven glider observations. Although the site experienced complete canopy consumption, all other factors of the site were preferable for greater gliders. For instance, the site was within an old-growth forest patch with numerous habitat hollows available, which is typically associated with high greater glider densities (Lindenmayer et al. 1990). Additionally, the site also had several adjacent low severity-fire refuges (closest was 500 m away). Given the small home ranges (1–3 ha) and slow dispersal (Kavanagh and Wheeler 2004) of greater gliders, the two most likely explanations are that greater gliders either survived the initial fire by seeking shelter in hollows, or that individuals survived by fleeing to nearby refuges and have since returned to the area in search of suitable habitat and food resources (Lindenmayer et al. 2013). The low-severity sites are evidence of refuge usage during high-severity wildfires because the surrounding landscape was highly burnt. These sites (BTC16, BTC25 and BTC40) were habitat of low burn severity, surrounded by high-severity burns, representing the typical landscape characteristics of fire refuges used by greater gliders (Chia et al. 2015). Given this, there may have been an influx of individuals into this refugia from the surrounding landscape (Bradstock et al. 2005; Robinson et al. 2013; Chia et al. 2016).
Hollow availability
Hollows are a fundamental habitat resource for greater gliders, being positively correlated to increased glider densities (Lindenmayer et al. 1990; Comport et al. 1996; Lindenmayer et al. 2020; Vinson et al. 2021). Used for nesting sites and shelter, hollows have ecological significance in greater glider life cycles (Goldingay 2012). In the current study, on average, more hollows were found in low fire-severity sites. Although low-severity sites were in a suite of topographic areas, including a gully, mid-slope and ridge tops, the highest habitat hollow counts came from sites in gullies. Gullies, which are typically wetter than other topographic regions, are likely to have lower fire frequency and severity than are other areas (Robinson et al. 2013; Chia et al. 2015). Gullies have also been identified as areas where old-growth forests are likely to occur because they are protected from wildfires and logging practices (Penman et al. 2007; Collins et al. 2012). Lindenmayer et al. (2016), similarly, found a positive correlation between hollow-bearing tree (HBT) abundance and topographic wetness, with highest HBT densities in unburnt gullies. In our study habitat, hollow densities were lower in moderate and high sites. Our findings showed similarities to those of Lindenmayer et al. (2016) and Banks et al. (2011), who found increased rates of HBT collapse in sites experiencing moderate and high fire-severity burns. However, in the long term, high-severity fires can act to form hollows by killing old-growth trees and fire scarring (Lindenmayer et al. 2016). Given the high density of habitat hollows in low fire-severity sites in our study, it is unsurprising that high densities of greater gliders were encountered within these sites. However, what was surprising was the relative high density of gliders in moderate sites that had the lowest density of habitat hollows, because they are considered to be areas of increased rates of HBT collapse in the past (Lindenmayer et al. 2016). Hollow-limiting environments are typically associated with low greater glider population density (Smith et al. 2007). The relatively high density recorded in moderate sites indicated a level of resilience in greater gliders to moderate fire severity and habitat use, even within hollow-limiting environments. In most cases, moderate sites occurred in patches surrounded by either of the other two severity classes (Fig. 4). The spatial characteristics of the sites that experienced moderate fire severity indicated that the high number of observations could be a spill-over effect of greater gliders dispersing from neighbouring low fire-severity habitat refuges (Berry et al. 2015). A reason for this dispersion could be the larger quantity of palatable eucalyptus epicormic regrowth observed within moderate sites than within low-severity sites, leading to food resource-driven dispersion (Kavanagh and Lambert 1990). Another cause could be due to social interactions and a tendency for male greater gliders to avoid home-range overlaps and high population densities (Pope et al. 2004). These results showed interesting habitat use in areas affected by moderate fire severity a year prior, highlighting the importance of these areas even when suitable habitat hollows are limiting.
Conclusions
Distance sampling successfully produced density estimates of greater gliders across MNP. Using the method, a low-density population of greater gliders in MNP was identified. This population was affected by the recent 2019–2020 wildfires, leading to concerning densities estimated within different fire-severity classes. More baseline data (and the use of more precise methods; see Cripps et al. 2021) are required in other areas affected by the 2019–2020 mega-fires along Australia’s eastern coast, to help determine the current conservation status of greater gliders. Given the predicted changes in climate leading to increased fire severity and frequency, understanding greater glider population responses to fire is critical to re-assess their vulnerability to extinction (Ward et al. 2020). Areas that experienced low fire severity act as refugia for greater gliders; however, future changes in climate may affect the recovery of populations if the frequency of wildfire events continues to increase (McLean et al. 2018). The consequences of this study are baseline estimates of greater glider density in MNP, while highlighting the effect that fire has on densities of both greater gliders, and essential habitat hollows.
Supplementary material
Supplementary material is available online.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors declare no conflict of interest.
Declaration of funding
This research was supported by a bushfire recovery grant provided by the Faculty Science, Medicine, and Health at the University of Wollongong, and the Centre of Sustainable Ecological Solutions. The supporting source was not involved in the collection of the data, preparation of the manuscript, nor the decision to submit for publication.
Acknowledgements
We acknowledge the traditional owners of the land on which this research was undertaken, the Walbanga people. The research was conducted under UOW animal ethics protocol AE19/02 and New South Wales Department of Planning, Industry and Environment Scientific Licence SL101968. The authors sincerely thank NSW National Parks and Wildlife Services ranger Chris Howard for park accessibility and ongoing assistance, and the Plumwood Conservation Trust for accommodation and access to Plumwood. Additionally, we also thank Madeline Newbery for her invaluable assistance in fieldwork activities.
References
Abram, NJ, Henley, BJ, Sen Gupta, A, et al. (2021). Connections of climate change and variability to large and extreme forest fires in southeast Australia. Communications Earth & Environment 2, 8.| Connections of climate change and variability to large and extreme forest fires in southeast Australia.Crossref | GoogleScholarGoogle Scholar |
Atlas of Living Australia (2021) Petauroides volans (greater glider). Available at https://bie.ala.org.au/species/urn:lsid:biodiversity.org.au:afd.taxon:7e891f26-c72e-4b29-98db-1cd10c4eaa6d [Accessed 23 August 2021]
Australian Bureau of Meteorology (2020) Average annual, seasonal and monthly rainfall. Available at http://www.bom.gov.au/jsp/ncc/cdio/weatherData/av?p_nccObsCode=139&p_display_type=dataFile&p_startYear=&p_c=&p_stn_num=069132 [Accessed 12 November 2020]
Banks, SC, Knight, EJ, McBurney, L, Blair, D, and Lindenmayer, DB (2011). The effects of wildfire on mortality and resources for an arboreal marsupial: resilience to fire events but susceptibility to fire regime change. PLoS One 6, e22952.
| The effects of wildfire on mortality and resources for an arboreal marsupial: resilience to fire events but susceptibility to fire regime change.Crossref | GoogleScholarGoogle Scholar | 21826221PubMed |
Berry, LE, Driscoll, DA, Banks, SC, and Lindenmayer, DB (2015). The use of topographic fire refuges by the greater glider (Petauroides volans) and the mountain brushtail possum (Trichosurus cunninghami) following a landscape-scale fire. Australian Mammalogy 37, 39–45.
| The use of topographic fire refuges by the greater glider (Petauroides volans) and the mountain brushtail possum (Trichosurus cunninghami) following a landscape-scale fire.Crossref | GoogleScholarGoogle Scholar |
Berry, LE, Lindenmayer, DB, Dennis, TE, Driscoll, DA, and Banks, SC (2016). Fire severity alters spatio-temporal movements and habitat utilisation by an arboreal marsupial, the mountain brushtail possum (Trichosurus cunninghami). International Journal of Wildland Fire 25, 1291–1302.
| Fire severity alters spatio-temporal movements and habitat utilisation by an arboreal marsupial, the mountain brushtail possum (Trichosurus cunninghami).Crossref | GoogleScholarGoogle Scholar |
Bradstock, RA, Williams, JE, Gill, MA (2002) ‘Flammable Australia: The Fire Regimes and Biodiversity of a Continent.’ (Cambridge University Press: Cambridge, UK.)
Bradstock, RA, Bedward, M, Gill, AM, and Cohn, JS (2005). Which mosaic? A landscape ecological approach for evaluating interactions between fire regimes, habitat and animals. Wildlife Research 32, 409–423.
| Which mosaic? A landscape ecological approach for evaluating interactions between fire regimes, habitat and animals.Crossref | GoogleScholarGoogle Scholar |
Buckland, ST, Anderson, DR, Burnham, KP, Laake, JL, Borchers, DL, Thomas, L (2001) ‘Introduction to Distance Sampling: Estimating Abundance of Biological Populations.’ (Oxford University Press: Oxford, UK)
Catling P (1991) Ecological effects of prescribed burning practices on the mammals of southeastern Australia. In ‘Conservation of Australia’s Forest Fauna’. (Ed. D Lunney) pp. 353–363. (Royal Zoological Society: Sydney, NSW, Australia)
Catling, PC, Coops, N, and Burt, RJ (2002). The distribution and abundance of ground-dwelling mammals in relation to time since wildfire and vegetation structure in south-eastern Australia. Wildlife Research 28, 555–565.
| The distribution and abundance of ground-dwelling mammals in relation to time since wildfire and vegetation structure in south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Chia, EK, Bassett, M, Nimmo, DG, Leonard, SWJ, Ritchie, EG, Clarke, MF, and Bennett, AF (2015). Fire severity and fire‐induced landscape heterogeneity affect arboreal mammals in fire‐prone forests. Ecosphere 6, 1–14.
| Fire severity and fire‐induced landscape heterogeneity affect arboreal mammals in fire‐prone forests.Crossref | GoogleScholarGoogle Scholar |
Chia, EK, Bassett, M, Leonard, SWJ, Holland, GJ, Ritchie, EG, Clarke, MF, and Bennett, AF (2016). Effects of the fire regime on mammal occurrence after wildfire: site effects vs landscape context in fire-prone forests. Forest Ecology and Management 363, 130–139.
| Effects of the fire regime on mammal occurrence after wildfire: site effects vs landscape context in fire-prone forests.Crossref | GoogleScholarGoogle Scholar |
Collins, L, Bradstock, RA, Tasker, EM, and Whelan, RJ (2012). Can gullies preserve complex forest structure in frequently burnt landscapes? Biological Conservation 153, 177–186.
| Can gullies preserve complex forest structure in frequently burnt landscapes?Crossref | GoogleScholarGoogle Scholar |
Collins, L, Bradstock, RA, Clarke, H, Clarke, MF, Nolan, RH, and Penman, TD (2021). The 2019/2020 mega-fires exposed Australian ecosystems to an unprecedented extent of high-severity fire. Environmental Research Letters 16, 044029.
| The 2019/2020 mega-fires exposed Australian ecosystems to an unprecedented extent of high-severity fire.Crossref | GoogleScholarGoogle Scholar |
Comport, SS, Ward, SJ, and Foley, WJ (1996). Home ranges, time budgets and food-tree use in a high-density tropical population of greater gliders, Petauroides volans minor (Pseudocheiridae: Marsupialia). Wildlife Research 23, 401–419.
| Home ranges, time budgets and food-tree use in a high-density tropical population of greater gliders, Petauroides volans minor (Pseudocheiridae: Marsupialia).Crossref | GoogleScholarGoogle Scholar |
Cripps, JK, Nelson, JL, Scroggie, MP, Durkin, LK, Ramsey, DSL, and Lumsden, LF (2021). Double-observer distance sampling improves the accuracy of density estimates for a threatened arboreal mammal. Wildlife Research 48, 756–768.
| Double-observer distance sampling improves the accuracy of density estimates for a threatened arboreal mammal.Crossref | GoogleScholarGoogle Scholar |
Crowther, MS, Dargan, JR, Madani, G, Rus, AI, Krockenberger, MB, McArthur, C, Moore, BD, Lunney, D, and Mella, VSA (2020). Comparison of three methods of estimating the population size of an arboreal mammal in a fragmented rural landscape. Wildlife Research 48, 105–114.
| Comparison of three methods of estimating the population size of an arboreal mammal in a fragmented rural landscape.Crossref | GoogleScholarGoogle Scholar |
Davey, S (1990). Methods for surveying the abundance and distribution of arboreal marsupials in a south coast forest of New South Wales. Wildlife Research 17, 427–445.
| Methods for surveying the abundance and distribution of arboreal marsupials in a south coast forest of New South Wales.Crossref | GoogleScholarGoogle Scholar |
Department of Planning, Industry and Environment (2020) Fire extent severity mapping. Version 2.1. Available at https://datasets.seed.nsw.gov.au/dataset/fire-extent-and-severity-mapping-fesm
de Tores, JP, and Elscot, S (2010). Estimating the population size of a threatened arboreal marsupial: use of distance sampling to dispense with ad hoc survey techniques. Wildlife Research 37, 512–523.
| Estimating the population size of a threatened arboreal marsupial: use of distance sampling to dispense with ad hoc survey techniques.Crossref | GoogleScholarGoogle Scholar |
Emerson, LD, Ballard, G-A, and Vernes, K (2019). Conventional distance sampling versus strip transects and abundance indices for estimating abundance of greater gliders (Petauroides volans) and eastern ringtail possums (Pseudocheirus peregrinus). Wildlife Research 46, 518–532.
| Conventional distance sampling versus strip transects and abundance indices for estimating abundance of greater gliders (Petauroides volans) and eastern ringtail possums (Pseudocheirus peregrinus).Crossref | GoogleScholarGoogle Scholar |
Eyre, TJ (2005). Hollow‐bearing trees inlarge glider habitat in south‐east Queensland,Australia: abundance, spatial distribution and management. Pacific Conservation Biology 11, 23–37.
| Hollow‐bearing trees inlarge glider habitat in south‐east Queensland,Australia: abundance, spatial distribution and management.Crossref | GoogleScholarGoogle Scholar |
Finlayson, GR, Diment, AN, Mitrovski, P, Thompson, GG, and Thompson, SA (2010). Estimating western ringtail possum (Pseudocheirus occidentalis) density using distance sampling. Australian Mammalogy 32, 197–200.
| Estimating western ringtail possum (Pseudocheirus occidentalis) density using distance sampling.Crossref | GoogleScholarGoogle Scholar |
Gibbons, P, Lindenmayer, DB, Barry, SC, and Tanton, MT (2000). Hollow formation in eucalypts from temperate forests in southeastern Australia. Pacific Conservation Biology 6, 218.
| Hollow formation in eucalypts from temperate forests in southeastern Australia.Crossref | GoogleScholarGoogle Scholar |
Goldingay, RL (2012). Characteristics of tree hollows used by Australian arboreal and scansorial mammals. Australian Journal of Zoology 59, 277–294.
| Characteristics of tree hollows used by Australian arboreal and scansorial mammals.Crossref | GoogleScholarGoogle Scholar |
Harper, MJ, McCarthy, MA, van der Ree, R, and Fox, JC (2004). Overcoming bias in ground-based surveys of hollow-bearing trees using double-sampling. Forest Ecology and Management 190, 291–300.
| Overcoming bias in ground-based surveys of hollow-bearing trees using double-sampling.Crossref | GoogleScholarGoogle Scholar |
Hennessy K, Lucas C, Nicholls N, Bathols J, Suppiah R, Ricketts J (2005) ‘Climate Change Impacts on Fire-weather in South-east Australia.’ (Climate Impacts Group, CSIRO Atmospheric Research and the Australian Government Bureau of Meteorology: Melbourne, Vic., Australia)
Henry S (1984) Social organisation of the greater glider (Petauroides volans) in Victoria. In ‘Possums and Gliders’. (Eds AP Smith, ID Hume) pp. 221–228. (Surrey Beatty: Sydney, NSW, Australia)
Kavanagh RP (1984) Seasonal changes in habitat use by gliders and possums in southeastern New South Wales. In ‘Possums and Gliders’. (Eds AP Smith, ID Hume) pp. 527–543. (Australian Mammal Society: Sydney, NSW, Australia)
Kavanagh, RP, and Lambert, MJ (1990). Food selection by the greater glider, Petauroides volans: is foliar nitrogen a determinant of habitat quality? Australian Wildlife Research 17, 285–299.
| Food selection by the greater glider, Petauroides volans: is foliar nitrogen a determinant of habitat quality?Crossref | GoogleScholarGoogle Scholar |
Kavanagh R, Wheeler R (2004) Home-range of the greater glider Petauroides volans in tall montane forest of southeastern New South Wales, and changes following logging. In ‘The Biology of Australian Possums and Gliders’. (Eds RL Goldingay, SM Jackson.) pp. 413–425. (Surrey Beatty and Sons: Sydney, NSW, Australia)
Kehl J, Borsboom A (1984) Home range, den tree use and activity patterns in the greater glider, Petauroides volans. In ‘Possums and Gliders’ (Eds AP Smith, ID Hume) pp. 229–236. (Surrey Beatty: Sydney, NSW, Australia)
Koch, AJ, Munks, SA, Driscoll, D, and Kirkpatrick, JB (2008). Does hollow occurrence vary with forest type? A case study in wet and dry Eucalyptus obliqua forest. Forest Ecology and Management 255, 3938–3951.
| Does hollow occurrence vary with forest type? A case study in wet and dry Eucalyptus obliqua forest.Crossref | GoogleScholarGoogle Scholar |
Laurance, WF (1990). Effects of weather on marsupial folivore activity in a north Queensland upland tropical rainforest. Australian Mammalogy 13, 41–47.
| Effects of weather on marsupial folivore activity in a north Queensland upland tropical rainforest.Crossref | GoogleScholarGoogle Scholar |
Lindenmayer, D, Cunningham, R, Tanton, M, Smith, A, and Nix, H (1990). Habitat requirements of the mountain brushtail possum and the greater glider in the montane ash-type eucalypt forests of the central highlands of Victoria. Australian Wildlife Research 17, 467–478.
| Habitat requirements of the mountain brushtail possum and the greater glider in the montane ash-type eucalypt forests of the central highlands of Victoria.Crossref | GoogleScholarGoogle Scholar |
Lindenmayer, DB, Cunningham, RB, Donnelly, CF, Incoll, RD, Pope, ML, Tribolet, CR, Viggers, KL, and Welsh, AH (2001). How effective is spotlighting for detecting the greater glider (Petauroides volans)? Wildlife Research 28, 105–109.
| How effective is spotlighting for detecting the greater glider (Petauroides volans)?Crossref | GoogleScholarGoogle Scholar |
Lindenmayer, DB, Pope, ML, and Cunningham, RB (2005). Patch use by the greater glider (Petauroides volans) in a fragmented forest ecosystem. II. Characteristics of den trees and preliminary data on den-use patterns. Wildlife Research 31, 569–577.
| Patch use by the greater glider (Petauroides volans) in a fragmented forest ecosystem. II. Characteristics of den trees and preliminary data on den-use patterns.Crossref | GoogleScholarGoogle Scholar |
Lindenmayer, DB, Blanchard, W, McBurney, L, Blair, D, Banks, SC, Driscoll, D, Smith, AL, and Gill, AM (2013). Fire severity and landscape context effects on arboreal marsupials. Biological Conservation 167, 137–148.
| Fire severity and landscape context effects on arboreal marsupials.Crossref | GoogleScholarGoogle Scholar |
Lindenmayer, DB, Blanchard, W, Blair, D, McBurney, L, and Banks, SC (2016). Environmental and human drivers influencing large old tree abundance in Australian wet forests. Forest Ecology and Management 372, 226–235.
| Environmental and human drivers influencing large old tree abundance in Australian wet forests.Crossref | GoogleScholarGoogle Scholar |
Lindenmayer, DB, Blanchard, W, Blair, D, McBurney, L, Taylor, C, Scheele, BC, Westgate, MJ, Robinson, N, and Foster, C (2020). The response of arboreal marsupials to long‐term changes in forest disturbance. Animal Conservation 24, 246–258.
| The response of arboreal marsupials to long‐term changes in forest disturbance.Crossref | GoogleScholarGoogle Scholar |
Lunney, D, Gresser, S, O’Neill, LE, Matthews, A, and Rhodes, J (2007). The impact of fire and dogs on koalas at Port Stephens, New South Wales, using population viability analysis. Pacific Conservation Biology 13, 189–201.
Marques, TA, Thomas, L, Fancy, SG, and Buckland, ST (2007). Improving estimates of bird density using multiple-covariate distance sampling. The Auk 124, 1229–1243.
| Improving estimates of bird density using multiple-covariate distance sampling.Crossref | GoogleScholarGoogle Scholar |
McGregor, DC, Padovan, A, Georges, A, Krockenberger, A, Yoon, H-J, and Youngentob, KN (2020). Genetic evidence supports three previously described species of greater glider, Petauroides volans, P. minor, and P. armillatus. Scientific reports 10, 19284.
| Genetic evidence supports three previously described species of greater glider, Petauroides volans, P. minor, and P. armillatus.Crossref | GoogleScholarGoogle Scholar | 33159131PubMed |
McLean, CM, Kavanagh, RP, Penman, T, and Bradstock, R (2018). The threatened status of the hollow dependent arboreal marsupial, the greater glider (Petauroides volans), can be explained by impacts from wildfire and selective logging. Forest Ecology and Management 415–416, 19–25.
| The threatened status of the hollow dependent arboreal marsupial, the greater glider (Petauroides volans), can be explained by impacts from wildfire and selective logging.Crossref | GoogleScholarGoogle Scholar |
Mikac, KM, Knipler, ML, Gracanin, A, and Newbery, MS (2021). Ground dwelling mammal response to fire: a case study from Monga National Park after the 2019/2020 Clyde Mountain fire. Austral Ecology , .
| Ground dwelling mammal response to fire: a case study from Monga National Park after the 2019/2020 Clyde Mountain fire.Crossref | GoogleScholarGoogle Scholar |
Nolan, RH, Collins, L, Leigh, A, Ooi, MKJ, Curran, TJ, Fairman, TA, Resco de Dios, V, and Bradstock, R (2021). Limits to post‐fire vegetation recovery under climate change. Plant, Cell & Environment 44, 3471–3489.
| Limits to post‐fire vegetation recovery under climate change.Crossref | GoogleScholarGoogle Scholar |
Penman, TD, Kavanagh, RP, Binns, DL, and Melick, DR (2007). Patchiness of prescribed burns in dry sclerophyll eucalypt forests in south-eastern Australia. Forest Ecology and Management 252, 24–32.
| Patchiness of prescribed burns in dry sclerophyll eucalypt forests in south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Pope, ML, Lindenmayer, DB, and Cunningham, RB (2004). Patch use by the greater glider (Petauroides volans) in a fragmented forest ecosystem. I. Home range size and movements. Wildlife Research 31, 559–568.
| Patch use by the greater glider (Petauroides volans) in a fragmented forest ecosystem. I. Home range size and movements.Crossref | GoogleScholarGoogle Scholar |
Robinson, NM, Leonard, SWJ, Ritchie, EG, Bassett, M, Chia, EK, Buckingham, S, Gibb, H, Bennett, AF, and Clarke, MF (2013). Refuges for fauna in fire‐prone landscapes: their ecological function and importance. Journal of Applied Ecology 50, 1321–1329.
| Refuges for fauna in fire‐prone landscapes: their ecological function and importance.Crossref | GoogleScholarGoogle Scholar |
Smith, GC, Mathieson, M, and Hogan, L (2007). Home range and habitat use of a low-density population of greater gliders, Petauroides volans (Pseudocheiridae: Marsupialia), in a hollow-limiting environment. Wildlife Research 34, 472–483.
| Home range and habitat use of a low-density population of greater gliders, Petauroides volans (Pseudocheiridae: Marsupialia), in a hollow-limiting environment.Crossref | GoogleScholarGoogle Scholar |
Stellar R (2005) ‘Monga intacta.’ (Robyn Stellar: Braidwood, NSW, Australia)
Sutherland, WJ, Pullin, AS, Dolman, PM, and Knight, TM (2004). The need for evidence-based conservation. Trends in Ecology & Evolution 19, 305–308.
| The need for evidence-based conservation.Crossref | GoogleScholarGoogle Scholar |
Thomas, L, Buckland, ST, Rexstad, EA, Laake, JL, Strindberg, S, Hedley, SL, Bishop, JRB, Marques, TA, and Burnham, KP (2010). Distance software: design and analysis of distance sampling surveys for estimating population size. Journal of Applied Ecology 47, 5–14.
| Distance software: design and analysis of distance sampling surveys for estimating population size.Crossref | GoogleScholarGoogle Scholar | 20383262PubMed |
van Der Ree R, Ward S, Handasyde K (2004) Distribution and conservation status of possums and gliders in Victoria. In ‘The Biology of Australian Possums and Gliders’. (Eds RL Goldingay, SM Jackson) pp. 91–110. (Surrey Beatty and Sons: Sydney, NSW, Australia)
Vinson, SG, Johnson, AP, and Mikac, KM (2021). Current estimates and vegetation preferences of an endangered population of the vulnerable greater glider at Seven Mile Beach National Park. Austral Ecology 46, 303–314.
| Current estimates and vegetation preferences of an endangered population of the vulnerable greater glider at Seven Mile Beach National Park.Crossref | GoogleScholarGoogle Scholar |
Ward, M, Tulloch, AIT, Radford, JQ, Williams, BA, Reside, AE, Macdonald, SL, Mayfield, HJ, Maron, M, Possingham, HP, Vine, SJ, and O’Connor, JL (2020). Impact of 2019–2020 mega-fires on Australian fauna habitat. Nature Ecology & Evolution 4, 1321–1326.
| Impact of 2019–2020 mega-fires on Australian fauna habitat.Crossref | GoogleScholarGoogle Scholar |
Wickham H (2016). ggplot2:Elegant Graphics for Data Analysis. Springer‐Verlag New York. ISBN: 978‐3‐319‐24277‐4. https://ggplot2.tidyverse.org
Wickham, H, Averick, M, Bryan, J, et al. (2019). Welcome to the Tidyverse. Journal of Open Source Software 4, 1686.
| Welcome to the Tidyverse.Crossref | GoogleScholarGoogle Scholar |
Wintle, BA, Kavanagh, RP, McCARTHY, MA, and Burgman, MA (2005). Estimating and dealing with detectability in occupancy surveys for forest owls and arboreal marsupials. The Journal of Wildlife Management 69, 905–917.
| Estimating and dealing with detectability in occupancy surveys for forest owls and arboreal marsupials.Crossref | GoogleScholarGoogle Scholar |
Wintle, BA, Legge, S, and Woinarski, JCZ (2020). After the megafires: What next for Australian wildlife? Trends in Ecology & Evolution 35, 753–757.
| After the megafires: What next for Australian wildlife?Crossref | GoogleScholarGoogle Scholar |
Woinarski JC, Burbidge A, Harrison P (2014) ‘The Action Plan for Australian Mammals 2012.’ (CSIRO Publishing: Melbourne, Vic., Australia)
Woinarski, JCZ, Burbidge, AA, and Harrison, PL (2015). Ongoing unraveling of a continental fauna: decline and extinction of Australian mammals since European settlement. Proceedings of the National Academy of Sciences 112, 4531–4540.
| Ongoing unraveling of a continental fauna: decline and extinction of Australian mammals since European settlement.Crossref | GoogleScholarGoogle Scholar |