

Microbially mediated reduction of FeIII and AsV in Cambodian sediments amended with 13C-labelled hexadecane and kerogen
Athanasios Rizoulis A , Wafa M. Al Lawati A B , Richard D. Pancost C , David A. Polya A , Bart E. van Dongen A and Jonathan R. Lloyd A DA School of Earth, Atmospheric and Environmental Sciences and Williamson Research Centre for Molecular Environmental Science, The University of Manchester, Oxford Road, Manchester, M13 9PL, UK.
B Higher College of Technology, Ministry of Manpower, Al Janubyyah Street, 133, Muscat, Sultanate of Oman.
C Organic Geochemistry Unit, The Cabot Institute, School of Chemistry, Cantock’s Close, Bristol University, Bristol, BS8 1TS, UK.
D Corresponding author. Email: jon.lloyd@manchester.ac.uk
Environmental Chemistry 11(5) 538-546 https://doi.org/10.1071/EN13238
Submitted: 22 December 2013 Accepted: 5 June 2014 Published: 25 September 2014
Journal Compilation © CSIRO Publishing 2014 Open Access CC BY-NC-ND
Environmental context. The use of groundwater with elevated concentrations of arsenic for drinking, cooking or irrigation has resulted in the worst mass poisoning in human history. This study shows that organic compounds that can be found in arsenic rich subsurface sediments may be used by indigenous microorganisms, contributing to the release of arsenic from the sediments into the groundwater. This study increases our understanding of the range of organic substrates (and their sources) that can potentially stimulate arsenic mobilisation into groundwaters.
Abstract. Microbial activity is generally accepted to play a critical role, with the aid of suitable organic carbon substrates, in the mobilisation of arsenic from sediments into shallow reducing groundwaters. The nature of the organic matter in natural aquifers driving the reduction of AsV to AsIII is of particular importance but is poorly understood. In this study, sediments from an arsenic rich aquifer in Cambodia were amended with two 13C-labelled organic substrates. 13C-hexadecane was used as a model for potentially bioavailable long chain n-alkanes and a 13C-kerogen analogue as a proxy for non-extractable organic matter. During anaerobic incubation for 8 weeks, significant FeIII reduction and AsIII mobilisation were observed in the biotic microcosms only, suggesting that these processes were microbially driven. Microcosms amended with 13C-hexadecane exhibited a similar extent of FeIII reduction to the non-amended microcosms, but marginally higher AsIII release. Moreover, gas chromatography–mass spectrometry analysis showed that 65 % of the added 13C-hexadecane was degraded during the 8-week incubation. The degradation of 13C-hexadecane was microbially driven, as confirmed by DNA stable isotope probing (DNA-SIP). Amendment with 13C-kerogen did not enhance FeIII reduction or AsIII mobilisation, and microbial degradation of kerogen could not be confirmed conclusively by DNA-SIP fractionation or 13C incorporation in the phospholipid fatty acids. These data are, therefore, consistent with the utilisation of long chain n-alkanes (but not kerogen) as electron donors for anaerobic processes, potentially including FeIII and AsV reduction in the subsurface.
Introduction
Arsenic release from sediments into groundwaters used for irrigation and drinking in West Bengal and Bangladesh has led to ‘the worst mass poisoning in human history’.[1] However, this problem is more extensive, as arsenic affected aquifers have been reported globally, from Cambodia and Vietnam,[2–6] to China,[7] Chile,[8] the USA[9] and Europe.[10] Owing to the catastrophic effect of groundwater arsenic pollution on the health of millions of people around the world, the factors controlling arsenic release into groundwater have been the subject of numerous studies, which have shown that various processes can contribute to increased AsIII mobilisation.[11–18]
One of these processes, microbial reduction of AsV, is widely accepted as being an important mechanism of AsIII release in shallow reducing aquifers.[19–23] Bioavailable organic carbon serves both as a carbon source and electron donor for the microbial respiratory reduction of sedimentary AsV, which serves as an electron acceptor, resulting in the release of AsIII, which is more mobile, to the aqueous phase.[20,24] AsV reduction was shown repeatedly in a series of microcosm studies set up with sediments from arsenic affected aquifers, where simple organic proxies such as acetate or lactate were used to stimulate microbial mobilisation of arsenic.[15,19,20,22,25,26] Molecular studies in some of these systems revealed the presence of known FeIII and AsV respiring bacteria, as well as arsenate respiratory reductase gene sequences.[19,25–27]
Nevertheless, the identification of the active members of the microbial communities in these complex sedimentary environments is challenging, especially when investigating specific microbe–substrate interactions. Thus, in some studies, the use of 13C-labelled organic carbon substrates (mainly acetate) in combination with molecular microbiology techniques (DNA stable isotope probing (DNA-SIP), followed by DNA fractionation and 16S rRNA gene cloning and sequencing) have allowed the identification of the active members of the microbial community during AsIII mobilisation.[25,26,28] Alternatively to DNA-SIP, the incorporation of 13C in phospholipid fatty acids (PLFAs), which are the major components of the cell membranes and remain intact in viable cells,[29] can be used to investigate environmental microbial communities.[30] This is because the concentration and distribution of PLFAs has the capacity to reflect rapid changes in the microbial populations and link specific biomarkers to a phylogenetic or functional group of organisms.[30–32]
Despite the usefulness of previous microcosm studies that have focussed on simple organic substrates (acetate or lactate), it is known that sediments in arsenic affected aquifers are often organically lean, i.e. with less than 1 % organic matter (OM)[33–35] and they contain different types of OM. Recent studies in sediments from arsenic hot spots in West Bengal,[27,36] Cambodia,[22,37] and Taiwan[38] have shown that extractable OM, including long chain n-alkanes and other hydrocarbons of immature (e.g. petroleum) origin are present. Microbial degradation of n-alkanes has been shown previously for a range of microorganisms (pure isolates or mixed microbial communities) under aerobic and anaerobic environments.[39–42] Moreover, a recent microcosm study showed almost complete degradation of (naturally occurring) petroleum derived n-alkanes during incubation of arsenic-rich West Bengal sediments under anoxic, arsenic-reducing conditions,[27] suggesting that these hydrocarbons could play a role in mediating the release of AsIII. However, extractable OM makes up ~5 % of the sedimentary OM, meaning that the majority, ~95 %, is part of the non-extractable material (e.g. kerogen[43]). To date, microbial degradation of kerogen appears largely restricted to well-oxygenated settings,[44] and is refractory under anaerobic conditions.[45] It remains unclear if kerogen can serve as a carbon source and electron donor for the microbial respiratory reduction of sedimentary FeIII or AsV.
Therefore, the aims of this study were to investigate (i) whether environmentally relevant organic carbon substrates can be microbially degraded and (ii) to assess their affect on AsIII mobilisation. We investigated these aims by establishing a series of anoxic microcosms, using sediments collected from an arsenic-affected aquifer in Cambodia, and supplemented with 13C-labelled substrates. 13C-Hexadecane was used as a representative of long chain n-alkanes, which are part of the extractable OM (i.e. the part of the OM that can be extracted using common organic solvents). We also artificially matured 13C-labelled algal biomass to generate a labelled kerogen analogue to represent the non-extractable part of the OM. Following anaerobic incubation for 8 weeks, degradation of the 13C-labelled substrates was determined by gas chromatography–mass spectrometry (GC-MS), and their incorporation into the microbial biomass was detected by DNA-SIP fractionation. This study should provide a better understanding of the mechanisms underpinning microbial release of AsIII in natural, arsenic affected aquifers.
Methodology
Study area and sample collection
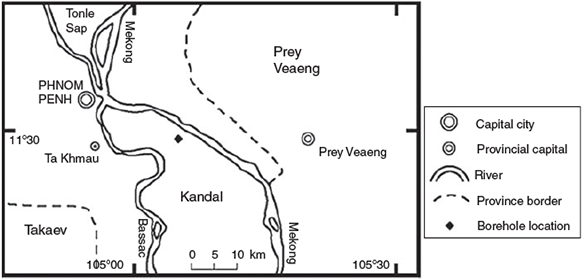
The sampling site was located in Rotaing village (Kean Svay District, Kandal Province), which lies between the Bassac and Mekong rivers, south of Phnom Penh, in Cambodia (Fig. 1). This region is known for high arsenic groundwater concentrations.[2,5,46] The sediment sample of this study was taken from a Holocene clay-rich horizon (6–7 m below ground surface) from a shallow aquifer, which is characterised by elevated (>50 μg L–1) groundwater arsenic concentrations.[4,5,47] The sediment was collected in July 2008 by drilling fluid-less within 1 m of sample collection and then up to 7 m, by augering, using groundwater from a nearby anoxic (>20-m depth) well as a drilling fluid. Upon retrieval, the full intact cores were immediately transferred into a glove bag, which was continuously flushed with N2 following the methods recommended by Rowland et al.[48] Sub-samples were sent to the UK, where they were stored under anoxic conditions, at 4 °C in the dark.
|
Sediment characterisation
Total carbon values were determined using a EuroVector EA3000 CHN elemental analyser (EuroVector, Milan, Italy). The inorganic carbon content was determined using a Strohlein Coulomat 702 analyser (Markus de Vries, Schwabach, Germany), which was modified to use phosphoric acid to release carbon dioxide from inorganic carbon in the sample. Total organic carbon (TOC) was calculated by subtracting inorganic carbon from total carbon values. Concentrations of the major and trace elements were determined on air-dried, finely ground pressed powder briquettes by X-ray fluorescence spectrometry (XRF; Axios, PANalytical, Almelo, Netherlands).
13C-Hexadecane and 13C-kerogen
13C-Hexadecane with certified label incorporation of 98.9 % was purchased from Buchem (Buchem BV, Apeldoorn, the Netherlands). 13C-Kerogen with label incorporation of 98 % was produced by artificial maturation of 13C-labelled cyanobacterial biomass purchased from Cambridge Isotope Laboratories (Cambridge Isotope Laboratories, Inc., Tewksbury, MA, USA). The detailed maturation procedure and characterisation of the 13C-labelled kerogen obtained is described elsewhere.[49] In short, 13C-labelled biomass was crushed in liquid N2, powdered and artificially matured in sealed gold cells at 330 °C and 300 bar for 24 h in the absence of water. The gold capsules were cracked and all the material was rinsed out using a combination of dichloromethane (DCM) and methanol (MeOH), both supplied by Sigma–Aldrich (Gillingham, UK). The kerogen was separated from the bitumen by sequential sonic extraction with DCM, DCM–MeOH and MeOH. Analyses by pyrolysis–gas chromatography–mass spectrometry indicates a good resemblance with pyrolysates of natural kerogens.[49]
Microcosm experiments and analyses
Four microcosm treatments were established in triplicate: (i) sediment only, (ii) sediment amended with 20 µL (2.3 mg g–1 sediment) of 13C-hexadecane, (iii) sediment amended with 0.5 mg (0.1 mg g–1 sediment) of 13C-kerogen and (iv) sediment sterilised by autoclaving (abiotic control). All microcosms were set up in 100-mL serum bottles (acid washed and sterilised by autoclaving), in a glove box under anaerobic conditions (98 : 2 %, N2/H2 atmosphere). All microcosms were set up by mixing 10 g of sediment and 20 mL of anaerobic sterile synthetic groundwater,[22] apart from the 13C-kerogen amended microcosms, which were set up by mixing 5 g of sediment and 10 mL of anaerobic sterile synthetic groundwater. The microcosms were maintained under anaerobic conditions by sealing the serum bottles with butyl rubber stoppers and aluminium crimp seals, and incubated at 20 °C in the dark. Approximately 2 mL of slurry was sampled anaerobically at the start of the incubation (t = 0), and after 2, 4, 6 and 8 weeks, with the exception of the kerogen amended microcosms, which were sampled at time points 0, 10 days and 8 weeks. Immediately after sampling, the pH and reduction potential (Eh) in the slurry sample was measured, and the concentration of 0.5 M HCl extractable FeII was determined using a ferrozine-based spectrophotometric assay.[50] In addition, arsenic speciation in the porewaters (filtered sub-samples through a 0.2-μm pore size filter) was determined by ion chromatography–inductively coupled plasma–mass spectrometry (IC-ICP-MS) using the method developed in Gault et al.[51] and described in Rowland et al.[22] Nitrate and sulfate measurements were taken in filtered slurry samples (through a 0.2-μm pore size filter) using a Metrohm 761 compact ion exchange chromatograph (Metrohm AG, Herisau, Switzerland). All microcosms were kept frozen at the end of their incubation period (8 weeks) until further analyses.
DNA isolation and 16S rRNA gene amplicon pyrosequencing
DNA was extracted from a sediment sample before incubation, a slurry sample from the sediment only treatment at the end of the incubation period (t = 8 weeks) and a slurry sample from the treatment amended with 13C-hexadecane at the end of the incubation period (t = 8 weeks). DNA isolations were carried out using the PowerSoil DNA extraction kit (MOBIO Laboratories, Carlsbad, CA, USA). Polymerase chain reaction (PCR) for amplicon pyrosequencing was performed using tagged fusion bacterial primers 27F and 338R, targeting the V1-V2 hypervariable region of the bacterial 16S rRNA gene (see Supplementary material). The pyrosequencing run was performed at The University of Manchester sequencing facility, using a Roche 454 Life Sciences GS Junior system. The 454 pyrosequencing reads were analysed using Qiime 1.6.0 release.[52] Details about the primer design and the analysis of the obtained pyrosequencing reads can be found in the Supplementary material. The pyrosequencing reads of this study have been deposited in the NCBI Sequence Read Archive (SRA) under accession number SRP046303 (BioProject ID: PRJNA260474).
DNA-SIP
DNA-SIP was performed on the extracted DNA from the treatment amended with 13C-hexadecane at the end of the incubation period (t = 8 weeks), according to the protocol by Neufeld et al.,[53] but adapted for 3.9-mL Quick-Seal, Polyallomer tubes (Beckman Coulter, Brea, CA, USA). Heavy and light DNA fractions were separated on a CsCl gradient, following centrifugation with an Optima TL-100 ultracentrifuge (Beckman Coulter) at 178 693g for 47 h at 20 °C. Eleven fractions of ~330-μL each were collected from each sample, and their density was determined using an analytical balance. The DNA in each of the collected fractions was precipitated as described previously,[53] resuspended in 30 μL of 10 mM Tris, 1 mM ethylenediaminetetraacetic acid buffer, quantified using a NanoDrop spectrophotometer (NanoDrop Products, Wilmington, DE, USA), and used as template for PCR amplification using primers 338F-GC and 519R,[54] targeting the hypervariable V3 region of the 16S rRNA gene. The PCR products were then subjected to denaturing gradient gel electrophoresis (DGGE) analysis, as previously described,[54] to investigate successful separation of the heavy (13C-labelled) and the light (unlabelled) DNA fractions.
Lipid (n-alkane and PLFA) extraction, fractionation and analysis
Lipids were extracted from 10 g of the original sediment amended with 20 µL of 13C-hexadecane (representing the t = 0 of the ‘sediment + 13C-hexadecane’ treatment), and from the remaining sediment slurries of the ‘sediment + 13C-hexadecane’ and ‘sediment + 13C-kerogen’ treatments at the end of the 8-week incubation period. The sediments (or slurries) were freeze-dried and ground, and the total lipid extracts (TLEs) were obtained by Bligh–Dyer extraction using a single-phase mixture of chloroform–methanol–KH2PO4 buffer (1 : 2 : 0.8, v/v/v, 18 mL).[55] After centrifugation at 2500 rpm for 15 min, the TLEs were concentrated using rotary evaporation. Subsequently, aliquots of the TLEs were separated into three fractions using a column packed with activated silica gel (0.5 g, 60-Å particle size). These fractions were eluted with chloroform–acetic acid (100 : 1, v/v, 5 mL; ‘simple lipid fraction’), acetone (20 mL; ‘glycolipid fraction’) and methanol (5 mL; ‘phospholipid (PL) fraction’). The simple lipid fraction was further separated into two fractions using a column packed with (activated) Al2O3 by elution with hexane–dichloromethane (9 : 1, v/v, 3 mL; ‘hydrocarbon fraction’) and dichloromethane–methanol (1 : 1, v/v, 3 mL; ‘polar fraction’).[22] PLFAs were generated and analysed as described in the Supplementary material.
Results
Sediment characteristics
The total organic carbon (TOC) content in the Cambodian sediment was 0.97 % (9.7 mg TOC g–1 of sediment), whereas the concentrations of total Fe, Mn and As were 56 000, 880 and 14 μg g–1 of sediment, as determined by XRF analysis (Table 1).

|
FeIII reduction and mobilisation of AsIII
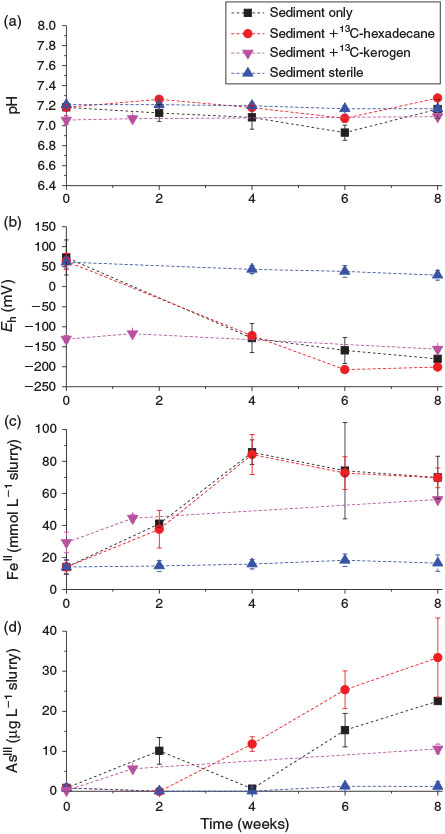
Four microcosm treatments were set up to investigate the role of OM in microbially mediated arsenic mobilisation processes. During the 8-week incubation, the pH, the Eh and the concentrations of FeII and AsIII were monitored (Fig. 2). The pH in all microcosms remained near neutral during the incubation period, with average values ranging between 6.9 and 7.3 (Fig. 2a). The starting Eh in the kerogen amended microcosms was significantly lower than in the other microcosms (–130 mV compared to between +43 and +73 mV), possibly because these microcosms were set up at a later time point than the other microcosms and the sediment may have become more reducing. After anaerobic incubation for 8 weeks, the Eh in the biotic microcosms decreased gradually to values between –156 and –200 mV but in the sterile microcosms it remained at positive values (Fig. 2b).
|
The concentration of FeII in the ‘sediment only’ and ‘sediment + 13C-hexadecane’ treatments exhibited similar patterns, increasing from 14 mmol FeII L–1 slurry at the start of the experiment to 70 mmol L–1 by the end of the incubation period (8 weeks), with the highest increase observed during the first 4 weeks (Fig. 2c). The reduction of up to 56 mM FeIII during the incubations corresponds to 112 μmoles of FeIII g–1 of sediment or 6.272 mg g–1 of sediment (i.e. 11.2 % of the total sedimentary Fe was reduced, Table 1). In the microcosms amended with 13C-kerogen, there was a lower increase in FeII concentration, from 29 to 56 mmol FeII L–1. The reduction of 27 mmol L–1 corresponds to 54 μmoles of FeIII g–1 of sediment or 3.024 μg g–1 of sediment. In the abiotic (autoclaved) microcosms, the concentration of FeII remained stable during the incubation period. For comparison, concentrations of other competing electron acceptors such as nitrate and sulfate were very low during the incubations, respectively up to 1.5 and 7 mg L–1. These concentrations were close to our analytical detection limits. Thus it was not possible to establish any conclusive nitrate- or sulfate-reducing trends during the course of the experiment (data not shown). These maximum concentrations correspond to 0.048 μmoles of NO3– and 0.146 μmoles of SO42– g–1 of sediment (Table 1).
Compared to background AsIII concentrations of 0.19–0.83 μg L–1 at the start of the incubations (t = 0), there was significant release of AsIII into the porewaters of all biotic microcosms after anaerobic incubation for 8 weeks, as the concentrations of AsIII increased to 23, 33 and 11 μg L–1 in the respective ‘sediment only’, ‘sediment + 13C-hexadecane’ and ‘sediment + 13C-kerogen’ treatments (Fig. 2d). The reduction of up to 33 μg As L–1 corresponds to 0.066 μg As g–1 of sediment (i.e. 0.47 % of the total As was reduced) or 0.00088 μmoles of As g–1 of sediment (Table 1). In contrast, the concentration of AsIII in the abiotic microcosms (‘sediment sterile’) remained stable throughout the 8-week incubation period, and up to 1.8 μg L–1.
Bacterial communities assessed by 16S rRNA gene amplicon pyrosequencing
Following 16S rRNA gene amplicon pyrosequencing, more than 2000 reads were retrieved from each sample and 125 to 188 operational taxonomic units were identified at 97 % sequence ID similarity level (Table S1 in the Supplementary material). Sequence analyses indicated a complex background community dominated by organisms affiliated with Sideroxydans species of the Gallionellaceae family. This could be a result of the long storage of the sediment before the set up of the microcosms of this study (20 months). However, compared to the microbial community before incubations (t = 0, sediment only), anaerobic incubation of the sediments (with or without supplemented 13C-hexadecane) led to a clear enrichment of sequences related to Geobacter and Sulfurospirillum genera (Table S2). Abundance of Geobacter-related sequences almost doubled from 3.6 % of the initial bacterial community (before incubations) to 6.9 and 7.6 % of the bacterial communities after incubation respectively without or with added 13C-hexadecane (Table S2). Similarly, the abundance of Sulfurospirillum-related sequences almost tripled from 1.3 % of the community before incubations to 3.7 and 4.2 % of the communities after incubation respectively without or with added 13C-hexadecane (Table S2). The majority of the Geobacter sequences of this study were related to FeIII-reducing Geobacter psychrophilus strain P35[56] and to Geobacter luticola,[57] whereas the Sulfurospirillum sequences of this study were related to AsV reducing Sulfurospirillum deleyianum strain DSM 6946.[58] In addition, Geothrix-related sequences were enriched during incubations (Table S2), with the majority of them related to FeIII reducing Geothrix fermentans strain H5.[59]
DNA-SIP fractionation of 13C-hexadecane amended microcosms
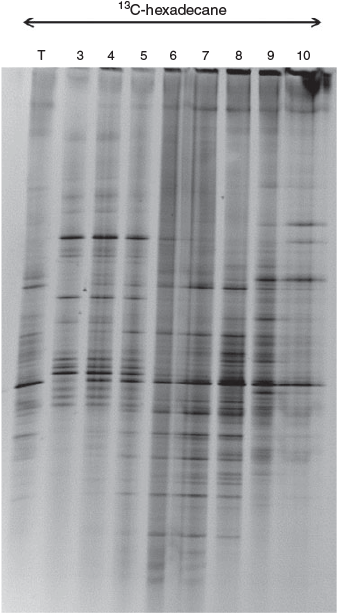
DNA-SIP and fractionation was carried out for the 13C-hexadecane amended microcosms, after the 8-week incubation. If 13C had not been incorporated into the DNA backbone of the active members of the microbial community, all DNA fractions would contain 12C-DNA, producing the same DGGE pattern. However, DNA-SIP fractionation followed by DGGE analysis of the PCR amplified DNA fractions, showed clearly that the first few fractions, which correspond to the ‘heavy’ 13C-labelled DNA, had a distinctly different banding pattern to the DGGE patterns of fractions 8 to 10, which correspond to the light DNA fraction (Fig. 3). The banding pattern of fractions 6 and 7 was a mixture of the heavy and light fractions (Fig. 3). Therefore, the different banding patterns between the collected DNA fractions not only indicate the successful 12C- and 13C-DNA separation along the density gradient, but also the successful separation of the DNA from the active members of the bacterial community (heavy DNA fractions 3–5). Moreover, the fact the DGGE profile of the total community (before DNA-SIP fractionation) resembles the pattern of the light DNA fractions (8–10), could be an indication that only a few members of the microbial community became enriched in 13C.
|
Quantification of extractable 13C-labelled n-alkanes and 13C-incorporation in the PLFAs
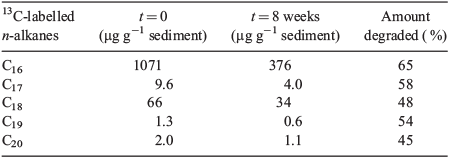
As shown in Table 2, the sediment spiked with 13C-labelled hexadecane before incubations was dominated by 13C-hexadecane (1100 μg g–1 of sediment) but also contained low amounts of other 13C-labelled n-alkanes (i.e. C17, C18, C19 and C20) with concentrations between 1.3 and 66 μg g–1 of sediment. The background sedimentary content of n-alkanes was 3.7 μg g–1 of sediment (sum of C20 to C34; data not shown). Following anaerobic incubation for 8 weeks, the amount of 13C-hexadecane had decreased to 380 μg g–1 of sediment (65 % degraded), whereas the remaining 13C-labelled n-alkanes, i.e. C17, C18, C19 and C20, had respectively degraded by 58, 48, 54 and 45 % (Table 2). The loss of 0.72 mg of hexadecane g–1 of sediment corresponds to 2.6 μmoles of hexadecane g–1 of sediment (Table 1). It was not possible to quantify 13C-incorporation in the extracted PLFAs (details in the Supplementary material).
|
Discussion
Degradation of hexadecane and kerogen under arsenic-reducing conditions
During the 8-week anoxic incubations, 13C-hexadecane was degraded as shown by the 65 % decrease in its concentration. Moreover, this degradation was microbially mediated, as shown by the 13C incorporation into the heavy DNA fractions of the ‘sediment + 13C-hexadecane’ treatment (Fig. 3). Previous studies have shown that n-alkanes (including n-hexadecane) can be degraded anaerobically by nitrate-reducing or sulfate-reducing bacterial isolates and consortia.[40,60–62] Moreover, Rowland et al.[27] showed marked degradation of n-alkanes (C20 to C34) in microcosms established using arsenic rich sediments. In our study, significant levels of respiration linked to reduction of the very low levels of nitrate and sulfate could not be detected during the course of the experiment, and stochiometric calculations indicated that neither nitrate or sulfate would have been dominant electron acceptors for the electrons that were potentially donated by the degradation of 2.6 μmoles of hexadecane (Table 1). From our data, it would seem clear that the microcosms of this study were predominantly FeIII-reducing systems, with other electron acceptors having minor contribution to the electron transfer balance (Fig. 2; Table 1). Interestingly, as 13C-hexadecane did not stimulate FeIII reduction we hypothesise that the majority of the electrons were donated by the unidentified indigenous OM (TOC 9.7 mg g–1 sediment, Table 1). However, as higher AsIII release was observed in the 13C-hexadecane amended microcosms (Fig. 2d), it is feasible that AsV was a minor electron sink for 13C-hexadecane oxidation (see below). Microbial degradation of 13C-hexadecane coupled to FeIII or AsV respiration, as our results suggest, could be feasible but it would constitute a novel mechanism of respiraton. Thus, it remains to be tested in enrichment microcosm experiments, using defined media supplemented with hexadecane as the sole electron donor and FeIII or AsV as the sole electron acceptor. Moreover, 16S rRNA gene and arsenate respiratory reductase (arrA) gene sequencing of the heavy DNA fractions would enable the respective identification of the 13C-hexadecane-utilising and arsenate-respiring members of the bacterial community, as shown in previous microcosm studies set up with arsenic rich sediments and amended with other 13C-labelled substrates.[25,26,28] However, sequencing efforts on the labelled fractions in our study were unsuccessful, most probably due to the very low DNA concentration in the heavy DNA fraction retrieved from the 13C-hexadecane amended sediment. Instead, we have successfully profiled the whole bacterial communities in three samples, using 16S rRNA gene amplicon pyrosequencing. The results indicated that incubation with or without 13C-hexadecane led to a clear enrichment in sequences related to FeIII-reducing Geobacter (and potentially AsV-reducing[63]) and Geothrix genera, as well as known AsV-reducing Sulfurospirillum species[58] (Table S2). Interestingly, a previous microcosm study that was established with the same sediment and supplemented with 13C-labelled acetate or 13C-labelled lactate showed that the heavy DNA fractions were also particularly enriched in sequences related to known FeIII-reducing Geobacter and AsV-respiring Sulfurospirillum species, whereas Geobacter-associated arrA gene sequences were retrieved too.[26]
Interestingly, the supplemented 13C-hexadecane also contained a mixture of other 13C-labelled n-alkanes (C17–C20), albeit in much lower amounts, and during the anaerobic incubations the shorter length n-alkanes were generally degraded to a greater degree than the longer chain n-alkanes (Table 2). However, this finding cannot be generalised, because previous studies showed that different bacterial isolates can utilise different long chain n-alkanes under aerobic or anaerobic conditions.[42] For example, under aerobic conditions Marinobacter hydrocarbonoclasticus strain 617 exhibited higher degradation of the C19–C21 than C16 n-alkanes,[64] whereas under anaerobic conditions sulfate-reducing strains Hxd3 and Pnd3 grew on n-alkanes in the range of C12–C20 and C14–C17 respectively.[65] Moreover, a slurry microcosm study showed that lower molecular weight n-alkanes (C11 to C13) were more recalcitrant than mid- to high-molecular weight n-alkanes under nitrate-reducing conditions, whereas under sulfate-reducing conditions degradation of long-chain (C32 to C39) n-alkanes was more extensive.[66] Thus, relative degradation rates of different n-alkanes are dependent on ecological and environmental conditions, including the structure of the microbial community present and electron acceptor availability.
Although 13C-hexadecane (and other n-alkanes) was clearly degraded during the incubation period by the microbial communities present, microbial degradation of the 13C-kerogen analogue could not be established in this study. DNA-SIP fractionation was not attempted for the 13C-kerogen amended microcosms because of very low DNA yields (data not shown), and the extracted PLFAs were too low in order for 13C-incorporation in the PLFA fraction to be confirmed with confidence. Thus, it appears that kerogen, the non-extractable fraction of the OM, was non-degradable under anaerobic conditions and over the time periods that we monitored. Future studies could explore again the anaerobic biodegradation of this type of OM, but over longer time scales, as this is something that has been stipulated but has not yet been confirmed conclusively.[45]
Effect of n-alkanes and kerogen on AsIII mobilisation
The effect of the addition of 13C-labelled hexadecane and 13C-labelled kerogen on AsIII mobilisation was investigated in anaerobic microcosms that were set up using arsenic-rich Cambodian sediment. The results indicated significant FeIII reduction and low AsIII mobilisation in all biotic microcosms of this study (‘sediment only’ or amended with 13C-labelled hexadecane or kerogen) but not in the sterile–abiotic microcosms (Fig. 2), confirming that these processes are biologically driven, as shown previously.[19,20,22]
The 13C-hexadecane-amended microcosms showed similar levels of FeIII reduction and a noticeable increase in AsIII release into the porewaters, in comparison to the ‘sediment only’ incubations (Fig. 2). Moreover, during incubations of the microcosms supplemented with 13C-hexadecane, there was a clear microbial degradation of the added 13C-hexadecane during the incubation (65 % of the supplied 13C-hexadecane was degraded), as discussed above. However, because there was some variability in the measured AsIII values (noted by the error bars, representing standard deviation from the mean, Fig. 2d), it is difficult to conclude unequivocally whether the observed increase in AsIII release reflects an experimental error due to sediment heterogeneity or is a result of stimulated microbial metabolic activity, due to the presence of additional bioavailable OM (the supplemented n-alkanes). Overall, the extent of FeIII reduction and AsIII release in these microcosms, measuring up to 7.84 mg FeII g–1 of sediment and up to 66 ng AsIII g–1 sediment, was comparable with the results of a previous microcosm study that was set up with sediments from the same area, but from a deeper depth (28 m, sample SYII28; Rowland et al.[22]). In the latter study, the addition of acetate did not significantly enhance FeIII reduction and AsIII release in comparison to the non-amended sediment, and up to 7 mg FeII and 46 ng AsIII g–1 sediment were detected after 4 weeks of incubation. However, Rowland et al.[22] observed much higher AsIII release (up to 400 ng g–1 of sediment) when microcosms were set up with sediment from 9-m depth and amended with acetate. This was attributed to the presence of an appropriate organic reductant in the deeper sediments, albeit at low concentrations, but its absence at shallower depths. Taking into account that AsIII may be reduced but remain sorbed and not partition into the aqueous phase, the release of AsIII in the porewaters does not depend only on the nature of bioavailable OM but also on the specific geochemistry and mineralogy. Thus, future studies on the effect of organic availability on the extent of microbially driven AsV mobilisation should also determine the speciation of arsenic in the solid phase using XANES, as it was done previously.[19]
In contrast to 13C-hexadecane amended microcosms, amendment with 13C-kerogen did not result in increased AsIII release. This could be attributed to the fact that only small amounts of 13C-kerogen was supplemented in comparison to background concentrations of OM, and to the recalcitrant nature of this OM under anoxic conditions. In fact, it has been noted that sediments of older age have higher proportions of recalcitrant OM (which has not been biodegraded), something which could explain the reverse relationship between (microbially mediated) AsIII release and sediment age.[67] However, it is not clear why in this study 13C-kerogen amended microcosms exhibited lower FeIII reduction and AsIII release than the non-amended microcosms (Fig. 2). This discrepancy is most likely due to the heterogeneous nature of the sediments used, for example with respect to background (Fe–As) mineralogy, supported by the higher initial FeII levels noted in kerogen amended microcosms.
In summary, this study demonstrated that natural microbial communities can utilise long chain n-alkanes under anaerobic conditions, and the addition of 13C-hexadecane appeared to have a minor but potentially significant effect on AsIII release from the arsenic-rich sediments used. In contrast, 13C-kerogen appeared not to be utilised over the time scale of our study, most likely due to the recalcitrant nature of this substrate or due to the presence of other bioavailable sedimentary OM in much higher amounts than the added 13C-kerogen. Further studies are required to clarify with confidence whether n-alkanes can be utilised in situ by arsenic respiring microorganisms and, if so, whether these organic substrates may play a critical role in the mobilisation of arsenic in shallow reducing aquifers across the globe.
Acknowledgements
A. Rizoulis was supported by NERC grant NE/D013291/1 to J. R. Lloyd, R. D. Pancost and D. A. Polya. W. M. Al Lawati gratefully acknowledges receipt of a Ph.D. studentship funded by the Ministry of Manpower, Sultanate of Oman. The authors thank Paul Lythgoe and Alastair Bewsher (University of Manchester) for chemical analyses, Dr Dave Cooke for collection of sediment samples and Marina Héry for technical advice supporting the DNA-SIP work described. Field work in Cambodia was supported by the European Commission funded Asia-Link project ‘CALIBRE: Cambodia and Lao Initiative for Building human Resources for the Environment’ (contract number KH/Asia-Link/04 142966). This document can under no circumstances be regarded as reflecting the position of the European Union or any other entities that have provided support to any of the authors.
References
[1] A. H. Smith, E. O. Lingas, M. Rahman, Contamination of drinking-water by arsenic in Bangladesh: a public health emergency. Bull. World Health Organ. 2000, 78, 1093.| 1:STN:280:DC%2BD3cvmsFSqtA%3D%3D&md5=ceb53fb427645e3595643e4f259bbf06CAS | 11019458PubMed |
[2] M. Berg, C. Stengel, P. T. K. Trang, P. H. Viet, M. L. Sampson, M. Leng, S. Samreth, D. Fredericks, Magnitude of arsenic pollution in the Mekong and Red River Deltas – Cambodia and Vietnam. Sci. Total Environ. 2007, 372, 413.
| Magnitude of arsenic pollution in the Mekong and Red River Deltas – Cambodia and Vietnam.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXitl2msw%3D%3D&md5=9b6b78e337e931a57e98ec3224d079e1CAS | 17081593PubMed |
[3] L. Charlet, D. A. Polya, Arsenic in shallow, reducing groundwaters in southern Asia: an environmental health disaster. Elements. 2006, 2, 91.
| Arsenic in shallow, reducing groundwaters in southern Asia: an environmental health disaster.Crossref | GoogleScholarGoogle Scholar |
[4] D. A. Polya, A. G. Gault, N. J. Bourne, P. R. Lythgoe, D. A. Cooke, Coupled HPLC-ICP-MS analysis indicates highly hazardous concentrations of dissolved arsenic species in Cambodian groundwaters, in Plasma Source Mass Spectrometry: Applications and Emerging Technologies (Eds J. G. Holland, S. D. Tanner) 2003, pp. 127–140 (The Royal Society of Chemistry: Cambridge, UK).
[5] D. A. Polya, A. G. Gault, N. Diebe, P. Feldman, J. W. Rosenboom, E. Gilligan, D. Fredericks, A. H. Milton, M. Sampson, H. A. L. Rowland, P. R. Lythgoe, J. C. Jones, C. Middleton, D. A. Cooke, Arsenic hazard in shallow Cambodian groundwaters. Mineral. Mag. 2005, 69, 807.
| Arsenic hazard in shallow Cambodian groundwaters.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XjsFCltrY%3D&md5=22a8558b2c89375f87b82a3c8187e61eCAS |
[6] M. Berg, H. C. Tran, T. C. Nguyen, H. V. Pham, R. Schertenleib, W. Giger, Arsenic contamination of groundwater and drinking water in Vietnam: a human health threat. Environ. Sci. Technol. 2001, 35, 2621.
| Arsenic contamination of groundwater and drinking water in Vietnam: a human health threat.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXks1ems74%3D&md5=700b9d2037e997612e900c070f73b9efCAS | 11452583PubMed |
[7] H. M. Guo, Y. X. Wang, G. M. Shpeizer, S. L. Yan, Natural occurrence of arsenic in shallow groundwater, Shanyin, Datong Basin, China. J. Environ. Sci. Health Part A Tox. Hazard. Subst. Environ. Eng. 2003, 38, 2565.
| Natural occurrence of arsenic in shallow groundwater, Shanyin, Datong Basin, China.Crossref | GoogleScholarGoogle Scholar |
[8] A. M. Sancha, M. O’Ryan, Managing hazardous pollutants in Chile: arsenic, in Reviews of Environmental Contamination and Toxicology, Vol 196 (Ed. D. M. Whitacre) 2008, pp. 123–146 (Springer Science+Business Media, LLC: New York).
[9] J. D. Ayotte, D. L. Montgomery, S. M. Flanagan, K. W. Robinson, Arsenic in groundwater in eastern New England: occurrence, controls, and human health implications. Environ. Sci. Technol. 2003, 37, 2075.
| Arsenic in groundwater in eastern New England: occurrence, controls, and human health implications.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXislSitLc%3D&md5=9bad7a9db1e138fe70d4a38a51b3f13dCAS | 12785510PubMed |
[10] A. L. Lindberg, W. Goessler, E. Gurzau, K. Koppova, P. Rudnai, R. Kumar, T. Fletcher, G. Leonardi, K. Slotova, E. Gheorghiuc, M. Vahter, Arsenic exposure in Hungary, Romania and Slovakia. J. Environ. Monit. 2006, 8, 203.
| Arsenic exposure in Hungary, Romania and Slovakia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28Xht1WktQ%3D%3D&md5=dbf86c82904e41b3d4ce4e68633d9da1CAS | 16395480PubMed |
[11] C. F. Harvey, C. H. Swartz, A. B. M. Badruzzaman, N. Keon-Blute, W. Yu, M. A. Ali, J. Jay, R. Beckie, V. Niedan, D. Brabander, P. M. Oates, K. N. Ashfaque, S. Islam, H. F. Hemond, M. F. Ahmed, Arsenic mobility and groundwater extraction in Bangladesh. Science 2002, 298, 1602.
| Arsenic mobility and groundwater extraction in Bangladesh.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38Xosl2ktb0%3D&md5=15bf4a1b66c8fa3cada9b42a1c6cdd03CAS | 12446905PubMed |
[12] M. L. Polizzotto, B. D. Kocar, S. G. Benner, M. Sampson, S. Fendorf, Near-surface wetland sediments as a source of arsenic release to ground water in Asia. Nature 2008, 454, 505.
| Near-surface wetland sediments as a source of arsenic release to ground water in Asia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXovV2mtLs%3D&md5=c6b5cb4d41410848c3e8658837b86453CAS | 18650922PubMed |
[13] J. M. McArthur, P. Ravenscroft, D. M. Banerjee, J. Milsom, K. A. Hudson-Edwards, S. Sengupta, C. Bristow, A. Sarkar, S. Tonkin, R. Purohit, How paleosols influence groundwater flow and arsenic pollution: a model from the Bengal Basin and its worldwide implication. Water Resour. Res. 2008, 44, W11411.
| How paleosols influence groundwater flow and arsenic pollution: a model from the Bengal Basin and its worldwide implication.Crossref | GoogleScholarGoogle Scholar |
[14] D. Postma, F. Larsen, N. T. M. Hue, M. T. Duc, P. H. Viet, P. Q. Nhan, S. Jessen, Arsenic in groundwater of the Red River floodplain, Vietnam: controlling geochemical processes and reactive transport modeling. Geochim. Cosmochim. Acta 2007, 71, 5054.
| Arsenic in groundwater of the Red River floodplain, Vietnam: controlling geochemical processes and reactive transport modeling.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXht1Gis7jP&md5=7e79166c6904be3bbb81f824e1d7d794CAS |
[15] A. van Geen, J. Rose, S. Thoral, J. M. Garnier, Y. Zheng, J. Y. Bottero, Decoupling of As and Fe release to Bangladesh groundwater under reducing conditions. Part II. Evidence from sediment incubations. Geochim. Cosmochim. Acta 2004, 68, 3475.
| Decoupling of As and Fe release to Bangladesh groundwater under reducing conditions. Part II. Evidence from sediment incubations.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXmvVGju7g%3D&md5=5de40bed5a77cc31766be46e01774db0CAS |
[16] M. Lawson, D. A. Polya, A. J. Boyce, C. Bryant, D. Mondal, A. Shantz, C. J. Ballentine, Pond-derived organic carbon driving changes in arsenic hazard found in Asian groundwaters. Environ. Sci. Technol. 2013, 47, 7085.
| 1:CAS:528:DC%2BC3sXptFeisb0%3D&md5=e8b44f76eb273f56a777dd117cd1be0eCAS | 23755892PubMed |
[17] B. J. Mailloux, E. Trembath-Reichert, J. Cheung, M. Watson, M. Stute, G. A. Freyer, A. S. Ferguson, K. M. Ahmed, M. J. Alam, B. A. Buchholz, J. Thomas, A. C. Layton, Y. Zheng, B. C. Bostick, A. van Geen, Advection of surface-derived organic carbon fuels microbial reduction in Bangladesh groundwater. Proc. Natl. Acad. Sci. USA 2013, 110, 5331.
| Advection of surface-derived organic carbon fuels microbial reduction in Bangladesh groundwater.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXntVehtbw%3D&md5=f62d70f6a2ffd857a9becb10c041d5c2CAS | 23487743PubMed |
[18] D. A. Polya, L. Charlet, Rising arsenic risk? Nat. Geosci. 2009, 2, 383.
| Rising arsenic risk?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXms1Ortbc%3D&md5=d91435bc2e0064ee363a05645439a394CAS |
[19] M. Héry, B. E. van Dongen, F. Gill, D. Mondal, D. J. Vaughan, R. D. Pancost, D. A. Polya, J. R. Lloyd, Arsenic release and attenuation in low organic carbon aquifer sediments from West Bengal. Geobiology 2010, 8, 155.
| Arsenic release and attenuation in low organic carbon aquifer sediments from West Bengal.Crossref | GoogleScholarGoogle Scholar | 20156294PubMed |
[20] F. S. Islam, A. G. Gault, C. Boothman, D. A. Polya, J. M. Charnock, D. Chatterjee, J. R. Lloyd, Role of metal-reducing bacteria in arsenic release from Bengal delta sediments. Nature 2004, 430, 68.
| Role of metal-reducing bacteria in arsenic release from Bengal delta sediments.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXlt1Cqt7c%3D&md5=07a3ade80cca1d42f0e375dd9a62d0aaCAS | 15229598PubMed |
[21] J. Akai, K. Izumi, H. Fukuhara, H. Masuda, S. Nakano, T. Yoshimura, H. Ohfuji, H. M. Anawar, K. Akai, Mineralogical and geomicrobiological investigations on groundwater arsenic enrichment in Bangladesh. Appl. Geochem. 2004, 19, 215.
| Mineralogical and geomicrobiological investigations on groundwater arsenic enrichment in Bangladesh.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXisFKntg%3D%3D&md5=c1250b47e608cf04a319bb5774241b88CAS |
[22] H. A. L. Rowland, R. L. Pederick, D. A. Polya, R. D. Pancost, B. E. Van Dongen, A. G. Gault, D. J. Vaughan, C. Bryant, B. Anderson, J. R. Lloyd, The control of organic matter on microbially mediated iron reduction and arsenic release in shallow alluvial aquifers, Cambodia. Geobiology 2007, 5, 281.
| The control of organic matter on microbially mediated iron reduction and arsenic release in shallow alluvial aquifers, Cambodia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhtFCgurfN&md5=42d0f2ef51b32b8f089e7d50d1435518CAS |
[23] R. S. Oremland, J. F. Stolz, Arsenic, microbes and contaminated aquifers. Trends Microbiol. 2005, 13, 45.
| Arsenic, microbes and contaminated aquifers.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXoslaltg%3D%3D&md5=31936b4578ef8ddb033476629e7a763cCAS | 15680760PubMed |
[24] R. S. Oremland, J. F. Stolz, The ecology of arsenic. Science 2003, 300, 939.
| The ecology of arsenic.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXjsVyjsLs%3D&md5=840e38f53bf5d9201af72c47691ceb50CAS | 12738852PubMed |
[25] G. Lear, B. Song, A. G. Gault, D. A. Polya, J. R. Lloyd, Molecular analysis of arsenate-reducing bacteria within Cambodian sediments following amendment with acetate. Appl. Environ. Microbiol. 2007, 73, 1041.
| Molecular analysis of arsenate-reducing bacteria within Cambodian sediments following amendment with acetate.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXitlyqsrg%3D&md5=ad2324dfc0f226db2f4a3d7a899583f4CAS | 17114326PubMed |
[26] M. Héry, A. Rizoulis, H. Sanguin, D. A. Cooke, R. D. Pancost, D. A. Polya, J. R. Lloyd, Microbial ecology of arsenic-mobilizing Cambodian sediments: lithological controls uncovered by stable-isotope probing. Environ. Microbiol. 2014, [Published online early 5 March 2014]
| Microbial ecology of arsenic-mobilizing Cambodian sediments: lithological controls uncovered by stable-isotope probing.Crossref | GoogleScholarGoogle Scholar | 24467551PubMed |
[27] H. A. L. Rowland, C. Boothman, R. Pancost, A. G. Gault, D. A. Polya, J. R. Lloyd, The role of indigenous microorganisms in the biodegradation of naturally occurring petroleum, the reduction of iron, and the mobilization of arsenite from West Bengal aquifer sediments. J. Environ. Qual. 2009, 38, 1598.
| The role of indigenous microorganisms in the biodegradation of naturally occurring petroleum, the reduction of iron, and the mobilization of arsenite from West Bengal aquifer sediments.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXosFKqsbk%3D&md5=95d68e3a8c14c35ea7e70dfe2f90dffeCAS |
[28] M. Héry, A. G. Gault, H. A. L. Rowland, G. Lear, D. A. Polya, J. R. Lloyd, Molecular and cultivation-dependent analysis of metal-reducing bacteria implicated in arsenic mobilisation in south-east asian aquifers. Appl. Geochem. 2008, 23, 3215.
| Molecular and cultivation-dependent analysis of metal-reducing bacteria implicated in arsenic mobilisation in south-east asian aquifers.Crossref | GoogleScholarGoogle Scholar |
[29] D. C. White, W. M. Davis, J. S. Nickels, J. D. King, R. J. Bobbie, Determination of the sedimentary microbial biomass by extractable lipid phosphate. Oecologia 1979, 40, 51.
| Determination of the sedimentary microbial biomass by extractable lipid phosphate.Crossref | GoogleScholarGoogle Scholar |
[30] R. P. Evershed, Z. M. Crossman, I. D. Bull, H. Mottram, J. A. J. Dungait, P. J. Maxfield, E. L. Brennand, 13C-Labelling of lipids to investigate microbial communities in the environment. Curr. Opin. Biotechnol. 2006, 17, 72.
| 13C-Labelling of lipids to investigate microbial communities in the environment.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhtlKlu7c%3D&md5=0dbeba5a75f2958ee9a270fc86f4b6c4CAS | 16423522PubMed |
[31] M. P. Waldrop, M. K. Firestone, Microbial community utilization of recalcitrant and simple carbon compounds: impact of oak-woodland plant communities. Oecologia 2004, 138, 275.
| Microbial community utilization of recalcitrant and simple carbon compounds: impact of oak-woodland plant communities.Crossref | GoogleScholarGoogle Scholar | 14614618PubMed |
[32] L. Zelles, Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: a review. Biol. Fertil. Soils 1999, 29, 111.
| Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: a review.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXisleks7c%3D&md5=07e3bc408db5504979762b493a6a4997CAS |
[33] A. G. Gault, F. S. Islam, D. A. Polya, J. M. Charnock, C. Boothman, D. Chatterjee, J. R. Lloyd, Microcosm depth profiles of arsenic release in a shallow aquifer, West Bengal. Mineral. Mag. 2005, 69, 855.
| Microcosm depth profiles of arsenic release in a shallow aquifer, West Bengal.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XjsFClt78%3D&md5=d876c66e7ab2ab252cc4c3cdfb79993bCAS |
[34] J. M. McArthur, D. M. Banerjee, K. A. Hudson-Edwards, R. Mishra, R. Purohit, P. Ravenscroft, A. Cronin, R. J. Howarth, A. Chatterjee, T. Talukder, D. Lowry, S. Houghton, D. K. Chadha, Natural organic matter in sedimentary basins and its relation to arsenic in anoxic ground water: the example of West Bengal and its worldwide implications. Appl. Geochem. 2004, 19, 1255.
| Natural organic matter in sedimentary basins and its relation to arsenic in anoxic ground water: the example of West Bengal and its worldwide implications.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXkt1WksL0%3D&md5=1c30e71d154a3c6b92091a7ffa71db05CAS |
[35] Y. Zheng, A. van Geen, M. Stute, R. Dhar, Z. Mo, Z. Cheng, A. Horneman, I. Gavrieli, H. J. Simpson, R. Versteeg, M. Steckler, A. Grazioli-Venier, S. Goodbred, M. Shahnewaz, M. Shamsudduha, M. A. Hoque, K. M. Ahmed, Geochemical and hydrogeological contrasts between shallow and deeper aquifers in two villages of Araihazar, Bangladesh: implications for deeper aquifers as drinking water sources. Geochim. Cosmochim. Acta 2005, 69, 5203.
| Geochemical and hydrogeological contrasts between shallow and deeper aquifers in two villages of Araihazar, Bangladesh: implications for deeper aquifers as drinking water sources.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXhtlerurrM&md5=877b7154ab06b5dc9171d54ded6e2075CAS |
[36] H. A. L. Rowland, D. A. Polya, J. R. Lloyd, R. D. Pancost, Characterisation of organic matter in a shallow, reducing, arsenic-rich aquifer, West Bengal. Org. Geochem. 2006, 37, 1101.
| Characterisation of organic matter in a shallow, reducing, arsenic-rich aquifer, West Bengal.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XptVSisr4%3D&md5=f3d8e8fb5433e776817f0b3cb2ded92fCAS |
[37] B. E. van Dongen, H. A. L. Rowland, A. G. Gault, D. A. Polya, C. Bryant, R. D. Pancost, Hopane, sterane and n-alkane distributions in shallow sediments hosting high arsenic groundwaters in Cambodia. Appl. Geochem. 2008, 23, 3047.
| Hopane, sterane and n-alkane distributions in shallow sediments hosting high arsenic groundwaters in Cambodia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhtlWkur3M&md5=d1b0c8c06d078cda317c8f10916207e1CAS |
[38] W. M. Al Lawati, J.-S. Jean, T. R. Kulp, M.-K. Lee, D. A. Polya, C.-C. Liu, B. E. van Dongen, Characterisation of organic matter associated with groundwater arsenic in reducing aquifers of southwestern Taiwan. J. Hazard. Mater. 2013, 262, 970.
| Characterisation of organic matter associated with groundwater arsenic in reducing aquifers of southwestern Taiwan.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXhsl2gt7bM&md5=834c6f23b079893ef8f91c8d4ac80ffaCAS | 22964390PubMed |
[39] C. De Pasquale, E. Palazzolo, L. Lo Piccolo, P. Quatrini, Degradation of long-chain n-alkanes in soil microcosms by two actinobacteria. J. Environ. Sci. Health Part A Tox. Hazard. Subst. Environ. Eng. 2012, 47, 374.
| Degradation of long-chain n-alkanes in soil microcosms by two actinobacteria.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XitVCgtro%3D&md5=7d8c312804ce04b7d1cc4fa9bc27c42dCAS |
[40] A. V. Callaghan, M. Tierney, C. D. Phelps, L. Y. Young, Anaerobic biodegradation of n-hexadecane by a nitrate-reducing consortium. Appl. Environ. Microbiol. 2009, 75, 1339.
| Anaerobic biodegradation of n-hexadecane by a nitrate-reducing consortium.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXjtVyqs7s%3D&md5=7be964c2b67d8bece64d4b0442b14ccbCAS | 19114507PubMed |
[41] V. Grossi, C. Cravo-Laureau, R. Guyoneaud, A. Ranchou-Peyruse, A. Hirschler-Réa, Metabolism of n-alkanes and n-alkenes by anaerobic bacteria: a summary. Org. Geochem. 2008, 39, 1197.
| Metabolism of n-alkanes and n-alkenes by anaerobic bacteria: a summary.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXovFOltr0%3D&md5=59c085860003019db7b1b27d0c40945bCAS |
[42] A. Wentzel, T. E. Ellingsen, H. K. Kotlar, S. B. Zotchev, M. Throne-Holst, Bacterial metabolism of long-chain n-alkanes. Appl. Microbiol. Biotechnol. 2007, 76, 1209.
| Bacterial metabolism of long-chain n-alkanes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhtVGmt7%2FN&md5=536601b309eb6149c1d023e2eae63862CAS | 17673997PubMed |
[43] B. Durant, Sedimentary organic matter and kerogen. Definition and quantitative importance of kerogen, in Kerogen-Insoluble Organic Matter from Sedimentary Rocks (Ed. B. Durand) 1980, pp 13–34 (Editions Technip: Paris).
[44] S. T. Petsch, R. A. Berner, T. I. Eglinton, A field study of the chemical weathering of ancient sedimentary organic matter. Org. Geochem. 2000, 31, 475.
| A field study of the chemical weathering of ancient sedimentary organic matter.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXktFartLw%3D&md5=992d5d001873461435876b21521ca383CAS |
[45] S. T. Petsch, T. I. Eglinton, K. J. Edwards, 14C-Dead living biomass: evidence for microbial assimilation of ancient organic carbon during shale weathering. Science 2001, 292, 1127.
| 14C-Dead living biomass: evidence for microbial assimilation of ancient organic carbon during shale weathering.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXjs1Churs%3D&md5=de08c72424686517dba7f0a0c6fbfa30CAS | 11283356PubMed |
[46] J. Buschmann, M. Berg, C. Stengel, M. L. Sampson, Arsenic and manganese contamination of drinking water resources in Cambodia: coincidence of risk areas with low relief topography. Environ. Sci. Technol. 2007, 41, 2146.
| Arsenic and manganese contamination of drinking water resources in Cambodia: coincidence of risk areas with low relief topography.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhs1SqtLs%3D&md5=a16a604437615f9276b03b299b805685CAS | 17438755PubMed |
[47] H. A. L. Rowland, A. G. Gault, P. Lythgoe, D. A. Polya, Geochemistry of aquifer sediments and arsenic-rich groundwaters from Kandal Province, Cambodia. Appl. Geochem. 2008, 23, 3029.
| Geochemistry of aquifer sediments and arsenic-rich groundwaters from Kandal Province, Cambodia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhtlWkurzF&md5=7bdec4825396a5ae958e5b2ad04acee9CAS |
[48] H. A. L. Rowland, A. G. Gault, J. M. Charnock, D. A. Polya, Preservation and XANES determination of the oxidation state of solid-phase arsenic in shallow sedimentary aquifers in Bengal and Cambodia. Mineral. Mag. 2005, 69, 825.
| Preservation and XANES determination of the oxidation state of solid-phase arsenic in shallow sedimentary aquifers in Bengal and Cambodia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XjsFCltrc%3D&md5=257eac8535e53d023dbbbb9d5b7bc321CAS |
[49] W. M. Al Lawati, The Role Of Organics In The Mobilization Of Arsenic In Shallow Aquifers 2012, Ph.D. Thesis, The University of Manchester.
[50] D. R. Lovley, E. J. P. Phillips, Availability of ferric iron for microbial reduction in bottom sediments of the freshwater tidal Potomac River. Appl. Environ. Microbiol. 1986, 52, 751.
| 1:CAS:528:DyaL28XmtFekur4%3D&md5=e53b1035bbf215729fa3f202e24d08aaCAS | 16347168PubMed |
[51] A. G. Gault, J. Jana, S. Chakraborty, P. Mukherjee, M. Sarkar, B. Nath, D. A. Polya, D. Chatterjee, Preservation strategies for inorganic arsenic species in high iron, low-Eh groundwater from West Bengal, India. Anal. Bioanal. Chem. 2005, 381, 347.
| Preservation strategies for inorganic arsenic species in high iron, low-Eh groundwater from West Bengal, India.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXht1KrtLc%3D&md5=48d105b47414a5daa8c0e93952c2cee7CAS | 15558247PubMed |
[52] J. G. Caporaso, J. Kuczynski, J. Stombaugh, K. Bittinger, F. D. Bushman, E. K. Costello, N. Fierer, A. G. Peña, J. K. Goodrich, J. I. Gordon, G. A. Huttley, S. T. Kelley, D. Knights, J. E. Koenig, R. E. Ley, C. A. Lozupone, D. McDonald, B. D. Muegge, M. Pirrung, J. Reeder, J. R. Sevinsky, P. J. Tumbaugh, W. A. Walters, J. Widmann, T. Yatsunenko, J. Zaneveld, R. Knight, QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335.
| QIIME allows analysis of high-throughput community sequencing data.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXksFalurg%3D&md5=8e8440f7b29c9f97830e90daed0dbef0CAS | 20383131PubMed |
[53] J. D. Neufeld, J. Vohra, M. G. Dumont, T. Lueders, M. Manefield, M. W. Friedrich, J. C. Murrell, DNA stable-isotope probing. Nat. Protoc. 2007, 2, 860.
| DNA stable-isotope probing.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhtFGnur7E&md5=37219e41e5b2c1d87e6c3237d973a071CAS | 17446886PubMed |
[54] D. R. Elliott, J. D. Scholes, S. F. Thornton, A. Rizoulis, S. A. Banwart, S. A. Rolfe, Dynamic changes in microbial community structure and function in phenol-degrading microcosms inoculated with cells from a contaminated aquifer. FEMS Microbiol. Ecol. 2010, 71, 247.
| Dynamic changes in microbial community structure and function in phenol-degrading microcosms inoculated with cells from a contaminated aquifer.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXnvVOmsA%3D%3D&md5=a4fd8823abf5724fe1f5f5349e8a45f5CAS | 19930459PubMed |
[55] Å. Frostegård, A. Tunlid, E. Bååth, Microbial biomass measured as total lipid phosphate in soils of different organic content. J. Microbiol. Methods 1991, 14, 151.
| Microbial biomass measured as total lipid phosphate in soils of different organic content.Crossref | GoogleScholarGoogle Scholar |
[56] K. P. Nevin, D. E. Holmes, T. L. Woodard, E. S. Hinlein, D. W. Ostendorf, D. R. Lovley, Geobacter bemidjiensis sp. nov. and Geobacter psychrophilus sp. nov., two novel Fe(III)-reducing subsurface isolates. Int. J. Syst. Evol. Microbiol. 2005, 55, 1667.
| Geobacter bemidjiensis sp. nov. and Geobacter psychrophilus sp. nov., two novel Fe(III)-reducing subsurface isolates.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXovV2nt7Y%3D&md5=4589c39d665e22b4aab94cb3011d470eCAS | 16014499PubMed |
[57] S. Viulu, K. Nakamura, Y. Okada, S. Saitou, K. Takamizawa, Geobacter luticola sp. nov., an Fe(III)-reducing bacterium isolated from lotus field mud. Int. J. Syst. Evol. Microbiol. 2013, 63, 442.
| Geobacter luticola sp. nov., an Fe(III)-reducing bacterium isolated from lotus field mud.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXmtlWrtL8%3D&md5=e1209b578ce95cb6c16dfe988bb37819CAS | 22493170PubMed |
[58] J. Sikorski, A. Lapidus, A. Copeland, T. G. Del Rio, M. Nolan, S. Lucas, F. Chen, H. Tice, J.-F. Cheng, E. Saunders, D. Bruce, L. Goodwin, S. Pitluck, G. Ovchinnikova, A. Pati, N. Ivanova, K. Mavromatis, A. Chen, K. Palaniappan, P. Chain, M. Land, L. Hauser, Y.-J. Chang, C. D. Jeffries, T. Brettin, J. C. Detter, C. Han, M. Rohde, E. Lang, S. Spring, M. Göker, J. Bristow, J. A. Eisen, V. Markowitz, P. Hugenholtz, N. C. Kyrpides, H.-P. Klenk, Complete genome sequence of Sulfurospirillum deleyianum type strain (5175T). Stand. Genomic Sci. 2010, 2, 149.
| Complete genome sequence of Sulfurospirillum deleyianum type strain (5175T).Crossref | GoogleScholarGoogle Scholar | 21304697PubMed |
[59] D. J. Lonergan, H. L. Jenter, J. D. Coates, E. J. P. Phillips, T. M. Schmidt, D. R. Lovley, Phylogenetic analysis of dissimilatory Fe(III)-reducing bacteria. J. Bacteriol. 1996, 178, 2402.
| 1:CAS:528:DyaK28Xit1Ors7s%3D&md5=c6332f7bc5031958f6dc47d7ebbe8983CAS | 8636045PubMed |
[60] H. Wilkes, R. Rabus, T. Fischer, A. Armstroff, A. Behrends, F. Widdel, Anaerobic degradation of n-hexane in a denitrifying bacterium: further degradation of the initial intermediate (1-methylpentyl)succinate via C-skeleton rearrangement. Arch. Microbiol. 2002, 177, 235.
| Anaerobic degradation of n-hexane in a denitrifying bacterium: further degradation of the initial intermediate (1-methylpentyl)succinate via C-skeleton rearrangement.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38Xitl2itLg%3D&md5=2b1c7f285893f4c86140383be68568f6CAS | 11907679PubMed |
[61] C. Cravo-Laureau, V. Grossi, D. Raphel, R. Matheron, A. Hirschler-Rea, Anaerobic n-alkane metabolism by a sulfate-reducing bacterium, Desulfatibacillum aliphaticivorans strain CV2803. Appl. Environ. Microbiol. 2005, 71, 3458.
| Anaerobic n-alkane metabolism by a sulfate-reducing bacterium, Desulfatibacillum aliphaticivorans strain CV2803.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXmt1ylsLs%3D&md5=6d65493ed551283aa771ad16fbff8186CAS | 16000749PubMed |
[62] I. A. Davidova, K. E. Duncan, O. K. Choi, J. M. Suflita, Desulfoglaeba alkanexedens gen. nov., sp nov., an n-alkane-degrading, sulfate-reducing bacterium. Int. J. Syst. Evol. Microbiol. 2006, 56, 2737.
| Desulfoglaeba alkanexedens gen. nov., sp nov., an n-alkane-degrading, sulfate-reducing bacterium.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhtVWmsLk%3D&md5=5331f998d5b6f9fb3d60451fdb7d2370CAS | 17158970PubMed |
[63] J. R. Lloyd, A. G. Gault, M. Héry, J. D. MacRae, Microbial transformations of arsenic in the subsurface, in Microbial Metal and Metalloid Metabolism: Advances and Applications (Eds J. F. Stolz, R. S. Oremland) 2011, pp. 77–90 (ASM Press: Washington, DC).
[64] P. Doumenq, E. Aries, L. Asia, M. Acquaviva, J. Artaud, M. Gilewicz, G. Mille, J. C. Bertrand, Influence of n-alkanes and petroleum on fatty acid composition of a hydrocarbonoclastic bacterium: Marinobacter hydrocarbonoclasticus strain 617. Chemosphere 2001, 44, 519.
| Influence of n-alkanes and petroleum on fatty acid composition of a hydrocarbonoclastic bacterium: Marinobacter hydrocarbonoclasticus strain 617.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXkvVehsbo%3D&md5=70fc969f6f6ac73bb904f83e5a480023CAS | 11482638PubMed |
[65] F. Aeckersberg, F. A. Rainey, F. Widdel, Growth, natural relationships, cellular fatty acids and metabolic adaptation of sulfate-reducing bacteria that utilize long-chain alkanes under anoxic conditions. Arch. Microbiol. 1998, 170, 361.
| Growth, natural relationships, cellular fatty acids and metabolic adaptation of sulfate-reducing bacteria that utilize long-chain alkanes under anoxic conditions.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXntleltbY%3D&md5=ff765c143960487f82ef6349d81137dbCAS | 9818355PubMed |
[66] M. Hasinger, K. E. Scherr, T. Lundaa, L. Bräuer, C. Zach, A. P. Loibner, Changes in iso- and n-alkane distribution during biodegradation of crude oil under nitrate and sulphate reducing conditions. J. Biotechnol. 2012, 157, 490.
| Changes in iso- and n-alkane distribution during biodegradation of crude oil under nitrate and sulphate reducing conditions.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XjtF2gtLw%3D&md5=3fa2e8c40f96120776b8b32b1e5671c5CAS | 22001845PubMed |
[67] D. Postma, F. Larsen, T. Nguyen Thi, T. Pham Thi Kim, R. Jakobsen, N. Pham Quy, L. Tran Vu, V. Pham Hung, A. S. Murray, Groundwater arsenic concentrations in Vietnam controlled by sediment age. Nat. Geosci. 2012, 5, 656.
| Groundwater arsenic concentrations in Vietnam controlled by sediment age.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhtFCgs7jP&md5=915a0d7d1be7a1bfebf707119fc9c80bCAS |