

Effects of addition of nitrogen on soil fungal and bacterial biomass and carbon utilisation efficiency in a city lawn soil
Xinyu Jiang A C , Lixiang Cao B and Renduo Zhang A CA Guangdong Provincial Key Laboratory of Environmental Pollution Control and Remediation Technology, School of Environmental Science and Engineering, Sun Yat-sen University, Guangzhou 510275, China.
B School of Life Science, Sun Yat-sen University, Guangzhou 510275, China.
C Corresponding authors. Email: zhangrd@mail.sysu.edu.cn; XIN.YU.JIANG1@gmail.com
Soil Research 52(1) 97-105 https://doi.org/10.1071/SR13210
Submitted: 2 May 2013 Accepted: 23 September 2013 Published: 5 February 2014
Abstract
The aim of this study was to investigate the effects of nitrogen (N) addition on soil microbial (fungal and bacterial) biomass and carbon utilisation efficiency (CUE) in a city lawn soil. A field experiment was conducted with three N treatments (kg N ha–1 year–1): the control (0), low-N (100), and high-N (200). Soil biogeochemical properties including pH, C : N, CUE, microbial biomass C (MBC), fungal and bacterial biomass, microbial C uptake rates, and soil respiration (SR) rates were determined during a 500-day experiment. The low- and high-N treatments significantly decreased soil pH, MBC, and CUE. Available N and soil acidification caused a decline in soil MBC. Soil acidification was not beneficial for microbial biomass growth, especially for bacteria. The treatments with N changed soil biomass from bacterial-dominant to fungal-dominant.
The results also showed that the CUE of bacterial-dominant soil was higher than that of fungal-dominant soil, which is contrary to previous studies. However, SR did not increase with decreased CUE under N treatments, because the addition of N limited soil microbial C uptake rates and significantly decreased soil microbial biomass. The CUE showed a negative correlation with soil temperature for the control treatment but not for the N treatments, which suggested that added N played a more important role in CUE than did soil temperature. Our results showed that addition of further N significantly alters soil biogeochemical properties, alters the ratio of bacteria to fungi, and decreases microbial carbon utilisation, which should provide important information for model-based prediction of soil C-cycling.
Additional keywords: carbon utilisation efficiency, microbial biomass, N treatment, soil acidification, soil carbon, soil respiration.
Introduction
Carbon utilisation efficiency (CUE) represents the fraction of assimilated C allocated to microbial growth (Micks et al. 2004; López-Urrutia and Morán 2007; Allison et al. 2010), and is an important determinant of C cycling in aquatic and terrestrial ecosystems. A higher CUE often indicates an increase in C storage in soils or slower soil organic C turnover. The CUE has been used as an important parameter in several carbon–climate models (Steinweg et al. 2008) for C and nitrogen (N) cycling predictions. For example, in the CENTURY model, a CUE value of 55% is assumed, suggesting that 45% of C taken up by microorganisms is lost as respiration (Parton et al. 1987). However, it has been reported that the CUE and pattern of soil microbial substrate use change with soil temperature (Allison et al. 2010), soil water content (Schimel et al. 1999; Devêvre and Horwáth 2000) and soil C : N (Keiblinger et al. 2010), which suggests that CUE is not a fixed value. Therefore, it is necessary to study variation in CUE under different environmental conditions.
The CUE is sensitive to environmental changes such as increases in N deposition. Human activities have significantly altered global N-cycling in the last several decades, resulting in several-fold increases in N deposition and soil N availability (Hobbie et al. 2002). In an N-limited soil, N addition usually increases the microbial CUE because a larger amount of available N should reduce the microbial energetic cost of N assimilation, so that more C uptake can be used for growth. Some short-term N-treatment experiments also showed that addition of N could stimulate soil CUE. For example, Thiet et al. (2006) reported that amendment with N increased the CUE by 20% in a 15-day laboratory experiment. Teklay et al. (2007) inferred that the absence of a pronounced respiratory response to the added N in a 60-day field experiment might be attributed to increased microbial CUE. However, excessive N input can result in soil acidification and changes in soil C : N, which may significantly affect soil microbial biomass, and this implies that CUE may not increase with N addition.
The issue of the relative contributions of fungal and bacterial biomass to soil microbial CUE has been controversial for many years. Some studies have reported that fungal CUE was higher than bacterial CUE because fungi have more biomass C per unit biomass N, and soils dominated by fungi may sequester more C than soils with lower fungal abundance (Parton et al. 1987; Zak et al. 1996). However, Thiet et al. (2006) did not find any difference between fungal and bacterial CUEs in a direct comparison based on a 14-day incubation experiment. However, 14 days may be insufficient time for soil microbes to respond to soil property changes under different treatments. For example, treatments with excessive N commonly cause soil acidification (Krusche et al. 2003). According to the results of Rousk et al. (2009), lower soil pH led to a decrease in bacterial growth and an increase in fungal growth, which suggested that the CUEs of fungi and bacteria both changed and there may be a difference between fungal and bacterial CUEs under soil acidification. Boyle (1998) and (Boberg et al. 2008) also showed that N or C addition could increase or decrease individual fungal CUE, which suggests that fungal or bacterial CUEs are not consistent values and can be changed under N treatments. In this way, treatments with N over longer periods might change fungal and bacterial CUEs and cause shifts in soil CUE between different treatments.
Relative amounts of soil fungal and bacterial biomass commonly vary with soil pH and C : N ratio (Hobbie 2000; Rousk et al. 2009; Nilsson et al. 2012). Ammonium hydrolysis after application of ammonium-based fertilisers is considered a critical factor influencing soil acidification (Krusche et al. 2003; Zhao and Xing 2009). Additional N incorporated into soil organic matter increased soil N content and reduced soil C : N ratio (Hobbie and Gough 2004). In this way, both pH and C : N ratio may change under N treatments. According to previous studies, lower soil pH values generally decrease bacterial competition and thus favour fungal growth (Rousk et al. 2008, 2009). Soil fungi were found to be more sensitive to additional N than bacteria because soil organic matter with lower C : N ratios resulting from the N addition is less favourable to fungi (Bossuyt et al. 2001).
We examined the response of soil microbial CUE to N treatments for 500 days in a subtropical city lawn. In south China, city lawns or grasslands represent about one-third of the land use in metropolitan areas (Jim and Chen 2006). In other regions around the world, such as the United States, ~8% of the land is urban, and 41% of this area is classified as residential, most of which is made up of lawns (Groffman and Pouyat 2009). Recently, atmospheric N deposition in metropolitan areas has significantly increased, which strongly affects CO2 emission from soils in these areas (Groffman et al. 2009). Given that lawns occupy a large proportion of metropolitan areas and can be greatly affected by increasing atmospheric N deposition, there is concern that N deposition could affect regional CO2 budgets. Furthermore, previous N treatments at the same experimental site showed that soil pH significantly decreased under N treatments. Therefore, we aimed to investigate: (i) whether N treatments could significantly change soil fungal and bacterial biomass; (ii) how soil fungal and bacterial CUE respond to N treatments.
Materials and methods
Site description
A city lawn at Sun Yat-sen University, Guangzhou, China (23°06′13″N, 113°17′41″E), was chosen as the experimental site. In 2008, an experimental station was established on the site, and several experiments related to soil respiration (SR) have been carried out since then (Lin et al. 2011). The botanical composition of the site includes Axonopus affinis, Eleusine indica, and Oxalis corniculata. With a subtropical monsoon climate in this region, the average annual temperature and rainfall are 21.9°C and 1735 mm, respectively. Most rainfall occurs between April and September. Detailed information about the experimental site is available in Lin et al. (2011). The soil texture of the study site is loamy with 43% sand, 35% silt, and 22% clay. The soil organic C content in the top 10 cm soil layer was 1.04%. The soil pH (soil to water 1 : 2.5) was 6.81.
Nitrogen addition experiment
Prior to N addition, nine treatment plots (3 m by 3 m) were established within the experimental site, and the distance between any two plots was >1 m. Three N addition levels were established: control (no addition), low-N (100 kg N ha–1 year–1 or 8.33 kg N ha–1 month–1), and high-N (200 kg N ha–1 year–1 or 16.67 kg N ha–1 month–1). Currently, atmospheric N deposition in southern China is in the range 30–73 kg N ha–1 year–1 and expected to increase by 50% in 2030 (Mo et al. 2007). Three treatment plots were randomly assigned to each N treatment. For each plot, four soil chamber collars (diameter 23 cm) were installed 10 cm deep into soil before the N treatments; one was used for measurement of SR rate and three for soil sampling. Before the N treatments and each measurement of SR rate, all grass on the soil surface within the chamber collars was removed by cutting. The N treatments were started on 1 July 2010 (day 0) and lasted for 18 months. Based on rates of 8.33 or 16.67 kg N ha–1 month–1, appropriate amounts of NH4NO3 was added to 100-L deionised water and the solution was applied to each plot with a hand-held sprayer at the beginning of each month. The control plots received the same amount of deionised water as the N treatments. Measurements of SR rates, CUE, and chemical and biological properties were conducted on days 0, 30, 45, 90, 180, 360, 450, and 500 after the N treatments.
Measurements of soil respiration rates
Rates of SR were measured using the Automatic Soil CO2 Exchange System (ACE; ADC BioScientific Ltd, Hoddesdon, UK). Prior to measurements of SR rate, the ACE station was placed on, and sealed to, the chamber collar. The chamber of each ACE station was 40 mm above the soil surface. Rates of SR were measured every 20 min during each measuring period of 24 h. Soil temperature (T) at 5 cm depth was measured synchronously using soil temperature sensors (ADC BioScientific Ltd). The measurement system was controlled and data were recorded by a computer. Averaged T and SR rates in each measuring time were used for the following analysis and discussion. At each sampling time, soil moisture contents of three treatments were determined using the gravimetric method (Dalal and Mayer 1986).
Soil analyses
On the dates of SR rate measurement, soil samples from the top 10 cm (diameter 10 cm) were taken from one chamber collar of each treatment plot. Each soil sample was divided into two subsamples. One was used to determine the CUE and the other to analyse the following properties. Soil total organic C (TOC) was determined using the potassium dichromate method, in which soil organic C was oxidised by potassium dichromate and then residual potassium dichromate was titrated using sodium thiosulfate (Yeomans and Bremner 1988). Total N (TN) was measured using an azotometer (Kjeltec 2300; Foss, Hillerød, Sweden). The C : N ratio was estimated as the quotient of TOC to TN. Soil pH was measured in 1 : 2.5 soil (g) : water (mL) using a pH electrode (FE20; Mettler Toledo, Switzerland).
Soil microbial biomass C (MBC) was determined with the fumigation–extraction method. Concentrations of TOC of the soil K2SO4 extract were determined by a TOC analyser (TOC-V CSH; Shimadzu Corporation, Kyoto, Japan) (Vance et al. 1987).
Fungal biomass was estimated by fluorescence microscopy. Fluorescent brightener 28 (FB28) was used to determine the total hypha bio-volume (Söderström 1977; Ingham and Klein 1984). Hyphal extraction was conducted following the method of Busse et al. (2009). Soil (10 g) was suspended in 95 mL of dilute Ringers solution (2.25 g NaCl, 0.11 g KCl, 0.12 g CaCl2, 0.05 g NaHCO3, and 1.0 L filter-sterilised-H2O). This suspension was shaken for 10 min on a wrist-action shaker. Then the suspension was diluted in 0.3 m phosphate buffer to a final dilution of 1 mg soil mL−1. All diluted samples (1 mL each) were stained with 0.5 mL FB28 (0.002 mg mL−1 filter-sterilised H2O clarified with 1 m NaOH) for 30 min at 24°C in the dark. After staining, the samples were filtered onto 0.2-μm blackened polycarbonate filters and viewed using an fluorescent microscope at 400× magnification under UV excitation. Hyphal length and width were measured based on 20 fields per sample and converted to biomass using the method of Bottomley (1994).
Bacterial biomass was determined with a similar procedure. Bacteria were extracted from soil (as before) and stained using 4,6-diamidino-2-phenylindole for 30 min at 24°C in the dark. Fluorescence microscopy was used to determine cell volumes (Bottomley 1994).
Determination of CUE
The CUE was determined according to Thiet et al. (2006) and Steinweg et al. (2008). Each soil sample collected from the study site was partially dried for 6 h and then placed into 12 bottles (40 g of soil in each bottle). Sealing rings with gas tubes were placed on the bottles. Six bottles were used for amended samples and the rest for non-amended samples. For each of the amended samples, 1.8 mL cellobiose solution (20 mg mL–1) was added (equivalent to 420 μg C g–1 soil), and then deionised water was added to maintain the soil water content at the same level as at sampling time. For the non-amended samples, only deionised water was added to maintain the soil moisture content at the same level as in the amended samples. At the beginning, and then every 3 h after the addition of the cellobiose solution or deionised water, 1 g soil was collected from each of three amended samples and three non-amended samples to measure cellobiose concentrations using the sulfuric acid–anthrone method for water-soluble carbohydrates (Brink et al. 1960).
The other three amended samples and three non-amended samples were used to measure CO2 concentrations in the bottles, using an infrared gas analyser (LI-840; LI-COR Inc., Lincoln, NE, USA) with gas tubes on the bottles. The incubation period lasted for 48 h. Remaining cellobiose and cellobiose-derived CO2 were determined by subtracting the results of the non-amended samples from those of the amended samples. CurveExpert v1.38 (D. G. Hyams, Starkville, MS, USA; www.curveexpert.net/) was used to fit the cellobiose concentration data to obtain the estimated time (te) at which the cellobiose concentration was close to zero. CurveExpert v1.38 was also used to fit CO2 concentrations to obtain the cumulative CO2 emission at te. The CUE was calculated as follows (Steinweg et al. 2008):

where CR is the cumulative CO2-C emission at te (μg g–1 soil), and C0 is the concentration of cellobiose-C measured at the beginning of cellobiose addition (μg g–1 soil). Concentration of CO2 was multiplied by 0.273 to convert CO2 to C. Cellobiose concentration was multiplied by 0.421 to convert cellobiose to C. Quotients of the added cellobiose-C to te were used to represent the soil microbial C uptake rates.
Data analysis
Statistics analysis was carried out using the SPSS software package (2003; SPSS Inc., Chicago, IL, USA). Two-way analysis of variance (ANOVA) was used on different soil biogeochemical properties (soil temperature, moisture content, pH, C : N, SR rates, microbial C uptake rate, CUE, MBC, fungal biomass, bacterial biomass, and fungal : bacterial biomass) with N addition levels and sampling time as fixed variables. Pearson correlation analysis was used to test the correlation among the different properties at P = 0.01 and P = 0.05. One-way ANOVA with Tukey’s HSD test was used to test differences of the properties among the treatments for each sampling time at P = 0.05.
Results
Soil pH, C : N, SR rates, and microbial C uptake rates
Soil temperature and moisture content showed no significant difference between three treatments during the whole experiment (Fig. 1). The two-way ANOVA analysis showed that N addition significantly decreased soil pH during the whole experiment (P < 0.05). After 500 days of the N treatments, pH values of the control, low-N, and high-N treatments changed from 6.81 to 6.78, 5.96, and 5.16, respectively (Fig. 2a). As shown in Fig. 2b, the C : N values were significantly lower in the N addition treatments than in the control (P < 0.05), and the lowest C : N occurred in the high-N treatment. After 500 days of N treatments, C : N values of the control, low-N, and high-N treatments were 11.26, 10.38, and 9.86, respectively. In general, the N addition treatments significantly decreased SR rates compared with the control (Fig. 2c, P < 0.01). Except for day 180, SR rates were significantly higher in the control than in the N treatments. On day 0, differences in microbial C uptake rates among the three treatments were not significant (Fig. 2d, P > 0.05). For the control, the microbial C uptake rates declined within the first 180 days, then increased after reaching the lowest values of 17.54 ± 0.74 μg h–1. For the low-N and high-N treatments, C uptake rates decreased from 30.49 ± 0.46 to 21.05 ± 0.98 μg h–1 and from 33.90 ± 0.46 to 17.83 ± 0.58 μg h–1, respectively, during 500 days of incubation. After 360 days, N addition significantly decreased microbial C uptake rates compared with the control.

|

|
Carbon utilisation efficiency, and fungal and bacterial biomass
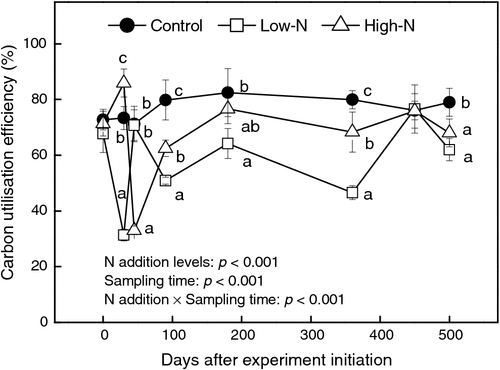
As shown in Fig. 3, soil microbial CUEs on day 0 were not significantly different among the three treatments (P > 0.05). From days 90 to 360, CUE was significantly lower in the low-N treatment than in the other two treatments (P < 0.05), while CUE of the control treatment was the highest among the treatments. Differences in CUE among the treatments on day 450 were negligible, and on day 500, CUE was significantly higher in the control than in the other two treatments (P < 0.05).
|
Differences of the MBC values among the three treatments were not significant on day 0 (Fig. 4a, P > 0.05). During the whole experiment, the N addition treatments significantly decreased MBC compared with the control (P < 0.01). As shown in Fig. 4b, effects of the N treatments on fungal biomass were generally not significant during the 500 days of the experiment. Differences in bacterial biomass values among the three treatments were not significant on day 0 (P > 0.05). However, N addition significantly decreased bacterial biomass over the course of the experiment (P < 0.05). Bacterial biomass values of the control varied from 70.5 to 75.6 μg g–1 soil, which was significantly higher than those of other two treatments (Fig. 4c). Ratios of fungal to bacterial biomass were significantly higher for the high-N treatment than for the other two treatments after 90 days of the N treatment (P < 0.01). For the control, ratios of fungal to bacterial biomass varied from 0.67 to 0.77 during the experiment. Ratios of fungal to bacterial biomass were >1 after 90 days for the high-N treatment and after 450 days for the low-N treatment (Fig. 4d).

|
Correlations among soil biogeochemical properties
Based on the results of correlation analysis, a significant correlation between pH and MBC was found (r = 0.658, n = 72, P < 0.01). The correlation between bacterial biomass and pH was also significant (r = 0.739, n = 72, P < 0.01), while the correlation between fungal biomass and pH was not significant (r = –0.315, n = 72, P > 0.05). The correlation between C : N and MBC was not significant (r = 0.412, n = 72, P > 0.05). The SR rates were positively correlated with bacterial biomass (r = 0.865, n = 72, P < 0.01), while the correlation between SR rates and fungal biomass was not significant (r = –0.206, n = 72, P > 0.05).
Values of CUE were positively correlated with the fungal biomass for the low-N (r = 0.797, n = 24, P < 0.05) and high-N (r = 0.835, n = 24, P < 0.05) treatments. However, the correlation between fungal biomass and CUE for the control treatment was not significant (r = 0.428, n = 24, P > 0.05). A significantly negative correlation between CUE and soil temperature was observed for the control treatment (r = –0.897, n = 24, P < 0.01).
Discussion
Previous studies showed that addition of a new C source resulted in enhanced degradation of old soil organic C due to the priming effect (Magid et al. 1999; Bell et al. 2003). In this study, part of the CO2 emitted after the cellobiose addition might come from the soil organic C, which likely made the CUE results lower than those determined using the 13C isotope method. However, because of the lag time between microbial uptake of 13C-glucose and actual mineralisation of the glucose, it is difficult to determine whether CO2 evolved at a specific time is solely from 13C-glucose and not from microbial biomass turnover (Thiet et al. 2006). Non-labelled substrates have been used to study CUEs with various treatments (Frey et al. 2001; Steinweg et al. 2008). As shown by Thiet et al. (2006), non-labelled and labelled substrates provided the same results for CUEs in fungi- and bacteria-dominant soil. In our study, our focus was the response of the estimate of CUE to N treatments. Non-labelled substrate should be suitable to investigate differences in CUE between different N treatments, although the results for CUEs in our study would be likely lower than those determined using 13C isotope-labelled substrate.
Soil microbial biomass commonly varies with soil available N and pH (Johnson et al. 1998; Aciego Pietri and Brookes 2009). Previous studies showed that N addition stimulated microbial biomass in grassland or pasture (Rousk and Bååth 2007; Hamer et al. 2009). However, decreases in microbial biomass with increasing N application in steppe have also been reported (Zhang et al. 2008; Bi et al. 2012), which is consistent with our results. The soil acidification caused by N addition in this study was more severe than that experienced in other studies (Magill et al. 1997; Lu et al. 2009). Excessive N addition commonly causes accumulation of soil NH4+. Ammonium hydrolysis is the main process for soil acidification (Li et al. 2010; Fang et al. 2012). Soil acidification can lead to mobilisation of Al or cause Al toxicity to soil microbes, which can greatly affect soil microbial biomass (Xu and Ji 2001).
Both N addition and pH can affect soil fungal or bacterial biomass. Previous studies showed that soil fungi were more sensitive to N addition and soil acidification than bacteria (Bardgett et al. 1999; Bardgett and McAlister 1999; Bossuyt et al. 2001). However, in our study, fungal biomass did not change significantly, whereas bacterial biomass decreased significantly under N addition, suggesting that soil bacteria were more sensitive to N addition. Soil fungal biomass and bacterial biomass was also correlated with soil pH (Aciego Pietri and Brookes 2009). The significant correlation between bacterial biomass and pH and the insignificant correlation between fungal biomass and pH in our study indicated that bacteria were more sensitive than fungi to soil acidification. Our results suggested that a decline in soil pH was not beneficial for soil microbial growth.
Our results did not support the theory that ecosystems with soils dominated by fungi may sequester more C than systems with lower fungal abundance. Addition of N changed the soil microbial composition from bacterial dominance to fungal dominance but decreased soil CUE in our field experiment. For the control treatment, microbial biomass was bacteria-dominant. For the N addition treatments, bacterial biomass was significantly lower than fungal biomass (Fig. 4), indicating that microbial biomass became fungi- dominant. The CUE of the control treatment was higher than those of the N addition treatments (Fig. 3), suggesting that CUE in the bacteria-dominant soil was higher than in the fungi-dominant soil. Therefore, the theory that soil dominated by fungi sequesters more C than bacteria-dominated soil could not be proved in our study. In general, fungi have a high biomass C : N ratio and a high CUE, with a positive relationship between C : N and CUE (Katharina et al. 2010). In our study, N treatments significantly decreased soil C : N and, similarly, lower soil C : N led to decline in soil fungal CUE.
Commonly, N addition may decrease microbial energetic costs of N assimilation (Bowden et al. 2004), so that more C uptake can be used for microbial growth, leading to higher CUE, especially in N-limited soils. We also observed a transitory increase of CUE under the high-N treatment from day 0 to day 45. From day 0 to day 45, soil acidification was not significant; therefore, the increase in CUE should be attributed to N addition in this short period. However, soil microbial CUE significantly decreased after 90 days for both the low-N and high-N treatments (Fig. 3), which suggested that there should be other, more important, factors controlling soil CUE beside available N. Interestingly, soil acidification and a decrease in CUE occurred simultaneously. As mentioned above, soil acidification limited soil microbial biomass growth, especially for the soil bacteria, which likely caused more microbial C uptake to be used for respiration rather than growth. Therefore, N addition played a key role in the increase in CUE in the short term (from day 0 to day 45), whereas soil pH was the major factor causing a decline in soil microbial CUE in the longer term.
Presumably, lower CUE means more C to be used for respiration, resulting in a higher SR rate. However, SR rates under the N treatments did not increase in our study. Previous studies showed that excessive N inputs could decrease soil microbial C uptake capability and limit soil microbial biomass growth (Compton et al. 2004; Follett et al. 2007). In our study, addition of N significantly decreased soil MBC and soil microbial C uptake rates compared with the control treatment (Fig. 2d). These two aspects likely prevented stimulation of SR rates and C loss from the soil.
The CUE is negatively correlated with incubation temperature (Steinweg et al. 2008). Therefore, an increase in the global temperature may lead to lower CUE, resulting in decline of C storage in the soil. In our study, CUE was negatively correlated with soil temperature for the control treatment. However, since N treatments change the soil microbial community structure and metabolism pattern (Hobbie and Gough 2004), the temperature-dependence of CUE disappeared for the N addition treatments. We also observed that SR rates for the three treatments were significantly lower on day 180 than at other sampling times. The lowest soil temperature on day 180 during the whole year led to decline of soil microbial activities, resulting in the lowest SR rates.
Conclusions
Our result confirmed the hypotheses: N treatments resulted in significant soil acidification, which changed the soil from bacterial dominance to fungal dominance, and then N treatments greatly reduced soil microbial CUE. Our results showed that the CUE of fungi-dominant soil was lower than that of bacteria-dominant soil and N treatments might decrease soil fungal CUE. It was difficult to compare fungal and bacterial CUE directly in our study because the CUEs of both fungi and bacteria might change under N treatments.
However, our results did not support the theory that fungal and bacterial CUEs were similar. If the CUEs of fungi and bacteria were similar, fungal and bacterial biomass should show similar varied patterns under N treatments; however, N treatments did not affect fungal biomass significantly but greatly decreased bacterial biomass. Contrary to previous theory that N addition may decrease microbial energetic costs of N assimilation and stimulate microbial growth, we found that, with available N and soil acidification, soil bacteria were more sensitive than fungi, and soil microbial biomass decreased under N treatments. Decline in soil CUE under N treatments did not lead to higher soil respiration rates and more C loss from soil, because N treatments also decreased soil microbial biomass and microbial C uptake rates. These issues warrant further research.
Acknowledgements
This work was partly supported by grants from the Chinese National Natural Science Foundation (Nos 51039007 and 51179212) and the Fundamental Research Funds for the Central Universities.
References
Aciego Pietri JC, Brookes PC (2009) Substrate inputs and pH as factors controlling microbial biomass, activity and community structure in an arable soil. Soil Biology & Biochemistry 41, 1396–1405.| Substrate inputs and pH as factors controlling microbial biomass, activity and community structure in an arable soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXnt1Oltb0%3D&md5=2e3cdb3c820e065a5ba2ab040eb74c39CAS |
Allison SD, Wallenstein MD, Bradford MA (2010) Soil-carbon response to warming dependent on microbial physiology. Nature Geoscience 3, 336–340.
| Soil-carbon response to warming dependent on microbial physiology.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXlsFSksLY%3D&md5=20ec5aece56b89588c41f879e9d82bddCAS |
Bardgett RD, McAlister E (1999) The measurement of soil fungal: bacterial biomass ratios as an indicator of ecosystem self-regulation in temperate meadow grasslands. Biology and Fertility of Soils 29, 282–290.
| The measurement of soil fungal: bacterial biomass ratios as an indicator of ecosystem self-regulation in temperate meadow grasslands.Crossref | GoogleScholarGoogle Scholar |
Bardgett RD, Lovell RD, Hobbs PJ, Jarvis SC (1999) Seasonal changes in soil microbial communities along a fertility gradient of temperate grasslands. Soil Biology & Biochemistry 31, 1021–1030.
| Seasonal changes in soil microbial communities along a fertility gradient of temperate grasslands.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXjsFCnu7w%3D&md5=d3a957d2a013892cc72ca314a2cbea2eCAS |
Bell JM, Smith JL, Bailey VL, Bolton H (2003) Priming effect and C storage in semi-arid no-till spring crop rotations. Biology and Fertility of Soils 37, 237–244.
Bi J, Zhang NL, Liang Y, Yang HJ, Ma KP (2012) Interactive effects of water and nitrogen addition on soil microbial communities in a semiarid steppe. Journal of Plant Ecology 5, 320–329.
| Interactive effects of water and nitrogen addition on soil microbial communities in a semiarid steppe.Crossref | GoogleScholarGoogle Scholar |
Boberg J, Finlay RD, Stenlid J, Nasholm T, Lindahl BD (2008) Glucose and ammonium additions affect needle decomposition and carbon allocation by the litter degrading fungus Mycena epipterygia. Soil Biology & Biochemistry 40, 995–999.
| Glucose and ammonium additions affect needle decomposition and carbon allocation by the litter degrading fungus Mycena epipterygia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhslGqtbw%3D&md5=d2c876c55d1ccefa25538788f63c34ccCAS |
Bossuyt H, Denef K, Six J, Frey SD, Merckx R, Paustian K (2001) Influence of microbial populations and residue quality on aggregate stability. Applied Soil Ecology 16, 195–208.
| Influence of microbial populations and residue quality on aggregate stability.Crossref | GoogleScholarGoogle Scholar |
Bottomley PJ (1994) Light microscopic methods for studying soil microorganisms. In ‘Methods of soil analysis. Part 2. Microbiological and biochemical properties’. (Ed. RW Weaver) pp. 81–105. (Soil Science Society of America: Madison, WI)
Bowden RD, Davidson E, Savage K, Arabia C, Steudler P (2004) Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest. Forest Ecology and Management 196, 43–56.
| Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest.Crossref | GoogleScholarGoogle Scholar |
Boyle D (1998) Nutritional factors limiting the growth of Lentinula edodes and other white-rot fungi in wood. Soil Biology & Biochemistry 30, 817–823.
| Nutritional factors limiting the growth of Lentinula edodes and other white-rot fungi in wood.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXjsFylurY%3D&md5=2e60969488db172c110dda488997a011CAS |
Brink RH, Dubach P, Lynch DL (1960) Measurement of carbohydrate in soil hydrolyzates with anthrone. Soil Science 89, 157–166.
| Measurement of carbohydrate in soil hydrolyzates with anthrone.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaF3MXhsFSgtg%3D%3D&md5=a45bf4ca902c22e6f134af28dce68d53CAS |
Busse MD, Sanchez FG, Ratcliff AW, Butnor JR, Carter EA, Powers RE (2009) Soil carbon sequestration and changes in fungal and bacterial biomass following incorporation of forest residues. Soil Biology & Biochemistry 41, 220–227.
| Soil carbon sequestration and changes in fungal and bacterial biomass following incorporation of forest residues.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXotV2isw%3D%3D&md5=2dc3444e67438942e210ad98103ee55eCAS |
Compton JE, Watrud LS, Porteous LA, DeGrood S (2004) Response of soil microbial biomass and community composition to chronic nitrogen additions at Harvard forest. Forest Ecology and Management 196, 143–158.
| Response of soil microbial biomass and community composition to chronic nitrogen additions at Harvard forest.Crossref | GoogleScholarGoogle Scholar |
Dalal RC, Mayer RJ (1986) Long term trends in fertility of soils under continuous cultivation and cereal cropping in southern Queensland. II. Total organic carbon and its rate of loss from the soil profile. Soil Research 24, 281–292.
| Long term trends in fertility of soils under continuous cultivation and cereal cropping in southern Queensland. II. Total organic carbon and its rate of loss from the soil profile.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL28XkvFKmsLw%3D&md5=0e8cc56dee57a5b989b47a587da88fd4CAS |
Devêvre OC, Horwáth WR (2000) Decomposition of rice straw and microbial carbon use efficiency under different soil temperatures and moistures. Soil Biology & Biochemistry 32, 1773–1785.
| Decomposition of rice straw and microbial carbon use efficiency under different soil temperatures and moistures.Crossref | GoogleScholarGoogle Scholar |
Fang Y, Xun F, Bai WM, Zhang WH, Li LH (2012) Long-term nitrogen addition leads to loss of species richness due to litter accumulation and soil acidification in a temperate steppe. PLoS ONE.
| Long-term nitrogen addition leads to loss of species richness due to litter accumulation and soil acidification in a temperate steppe.Crossref | GoogleScholarGoogle Scholar | 23166811PubMed |
Follett RF, Paul EA, Pruessner EG (2007) Soil carbon dynamics during a long-term incubation study involving 13C and 14C measurements. Soil Science 172, 189–208.
| Soil carbon dynamics during a long-term incubation study involving 13C and 14C measurements.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXkt1KrtL0%3D&md5=e082c3979bc2bd89d3875a9350a586a4CAS |
Frey SD, Gupta VVSR, Elliott ET, Paustian K (2001) Protozoan grazing affects estimates of carbon utilization efficiency of the soil microbial community. Soil Biology & Biochemistry 33, 1759–1768.
| Protozoan grazing affects estimates of carbon utilization efficiency of the soil microbial community.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXotFOlu74%3D&md5=65bf7d769ea827a1c2e9241786862291CAS |
Groffman PM, Pouyat RV (2009) Methane uptake in urban forests and lawns. Environmental Science & Technology 43, 5229–5235.
| Methane uptake in urban forests and lawns.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXmvFWqsL4%3D&md5=4b86ec9e969dd6442776a371fdc52cd5CAS |
Groffman PM, Williams CO, Pouyat RV, Band LE, Yesilonis ID (2009) Nitrate leaching and nitrous oxide flux in urban forests and grasslands. Journal of Environmental Quality 38, 1848–1860.
| Nitrate leaching and nitrous oxide flux in urban forests and grasslands.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhtFCjsrfI&md5=fe458bf6deff16b43d8500844b445676CAS | 19643750PubMed |
Hamer U, Potthast K, Makeschin F (2009) Urea fertilisation affected soil organic matter dynamics and microbial community structure in pasture soils of Southern Ecuador. Applied Soil Ecology 43, 226–233.
| Urea fertilisation affected soil organic matter dynamics and microbial community structure in pasture soils of Southern Ecuador.Crossref | GoogleScholarGoogle Scholar |
Hobbie SE (2000) Interactions between lignin and soil nitrogen availability during leaf litter decomposition in a Hawaiian montane forest. Ecosystems 3, 484–494.
| Interactions between lignin and soil nitrogen availability during leaf litter decomposition in a Hawaiian montane forest.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXovFWgt7s%3D&md5=dabe21737f34d0eeda6f98edd99ac49bCAS |
Hobbie SE, Gough L (2004) Litter decomposition in moist acidic and non-acidic tundra with different glacial histories. Oecologia 140, 113–124.
| Litter decomposition in moist acidic and non-acidic tundra with different glacial histories.Crossref | GoogleScholarGoogle Scholar | 15164284PubMed |
Hobbie SE, Nadelhoffer KJ, Högberg P (2002) A synthesis: the role of nutrients as constraints on carbon balances in boreal and arctic regions. Plant and Soil 242, 163–170.
| A synthesis: the role of nutrients as constraints on carbon balances in boreal and arctic regions.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XlvFWjsr4%3D&md5=1490d2b25b3359326cf6c402b73d5b56CAS |
Ingham ER, Klein DA (1984) Soil fungi: relationships between hyphal activity and staining with fluorescein diacetate. Soil Biology & Biochemistry 16, 273–278.
| Soil fungi: relationships between hyphal activity and staining with fluorescein diacetate.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL2cXls1Krt7o%3D&md5=968559391f80049b2f05db45361e3669CAS |
Jim CY, Chen WY (2006) Perception and attitude of residents toward urban green spaces in Guangzhou (China). Environmental Management 38, 338–349.
| Perception and attitude of residents toward urban green spaces in Guangzhou (China).Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD28vkvFGnsg%3D%3D&md5=41697e82f0cfa098027c088e7eb1e01eCAS | 16752045PubMed |
Johnson D, Leake JR, Lee JA, Campbell CD (1998) Changes in soil microbial biomass and microbial activities in response to 7 years simulated pollutant nitrogen deposition on a heathland and two grasslands. Environmental Pollution 103, 239–250.
| Changes in soil microbial biomass and microbial activities in response to 7 years simulated pollutant nitrogen deposition on a heathland and two grasslands.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXotVWqtL0%3D&md5=99cd1bba3800e7ad5df0756ebad587fcCAS |
Katharina MK, Edward KH, Wolfgang W, Ute S, Ieda H, Gunther E, Sandra B, Joseph S, Katja S, Andreas R, Sophie ZB (2010) The effect of resource quantity and resource stoichiometry on microbial carbon-use-efficiency. FEMS Microbiology Ecology 73, 430–440.
Keiblinger KM, Hall EK, Wanek W, Szukics U, Hämmerle I, Ellersdorfer G, Böck S, Strauss J, Sterflinger K, Richter A, Zechmeister-Boltenstern S (2010) The effect of resource quantity and resource stoichiometry on microbial carbon-use efficiency. FEMS Microbiology Ecology 73, 430–440.
Krusche AV, De Camargo PB, Cerri CE, Ballester MV, Lara LBLS, Victoria RL, Martinelli LA (2003) Acid rain and nitrogen deposition in a sub-tropical watershed (Piracicaba): ecosystem consequences. Environmental Pollution 121, 389–399.
| Acid rain and nitrogen deposition in a sub-tropical watershed (Piracicaba): ecosystem consequences.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XptlWjsLY%3D&md5=226ec8cd7ef409beed0ef8ea7c619c98CAS | 12685767PubMed |
Li LJ, Zeng DH, Yu ZY, Fan ZP, Mao R (2010) Soil microbial properties under N and P additions in a semi-arid, sandy grassland. Biology and Fertility of Soils 46, 653–658.
| Soil microbial properties under N and P additions in a semi-arid, sandy grassland.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXoslyktrY%3D&md5=81d1d115fc0fa61fb36064645c85f84fCAS |
Lin Z, Zhang R, Tang J, Zhang J (2011) Effects of high soil water content and temperature on soil respiration. Soil Science 176, 150–155.
| Effects of high soil water content and temperature on soil respiration.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXis1yjsbY%3D&md5=ba58036ccbf1ae738513817772968bf5CAS |
López-Urrutia A, Morán XAG (2007) Resource limitation of bacterial production distorts the temperature dependence of oceanic carbon cycling. Ecology 88, 817–822.
| Resource limitation of bacterial production distorts the temperature dependence of oceanic carbon cycling.Crossref | GoogleScholarGoogle Scholar | 17536698PubMed |
Lu XK, Mo JM, Gundersern P, Zhu WX, Zhou GY, Li DJ, Zhang X (2009) Effect of simulated N deposition on soil exchangeable cations in three forest types of subtropical China. Pedosphere 19, 189–198.
| Effect of simulated N deposition on soil exchangeable cations in three forest types of subtropical China.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXkvFCltb4%3D&md5=cf87cde660dd65c4ea018ac55cce298eCAS |
Magid J, Kjaergaard GA, Kuikman PJ (1999) Drying and rewetting of a loamy sand soil did not increase turnover of native organic matter, but retarded the decomposition of added 14C-labelled plant material. Soil Biology & Biochemistry 31, 595–602.
| Drying and rewetting of a loamy sand soil did not increase turnover of native organic matter, but retarded the decomposition of added 14C-labelled plant material.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXitlansbk%3D&md5=280b6c94f1309aef9c0c049fe84e463dCAS |
Magill AH, Aber JD, Hendricks JJ, Bowden RD, Melillo JM, Steudler PA (1997) Biogeochemical response of forest ecosystems to simulated chronic nitrogen deposition. Ecological Applications 7, 402–415.
| Biogeochemical response of forest ecosystems to simulated chronic nitrogen deposition.Crossref | GoogleScholarGoogle Scholar |
Micks P, Aber JD, Boone RD, Davidson EA (2004) Short-term soil respiration and nitrogen immobilization response to nitrogen applications in control and nitrogen-enriched temperate forests. Forest Ecology and Management 196, 57–70.
| Short-term soil respiration and nitrogen immobilization response to nitrogen applications in control and nitrogen-enriched temperate forests.Crossref | GoogleScholarGoogle Scholar |
Mo JM, Zhang W, Zhu WX, Fang YT, Li DJ, Zhao P (2007) Response of soil respiration to simulated N deposition in a disturbed and a rehabilitated tropical forest in southern China. Plant and Soil 296, 125–135.
| Response of soil respiration to simulated N deposition in a disturbed and a rehabilitated tropical forest in southern China.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXnsFejsrc%3D&md5=cc8d2a06431af58fceb980f29d734a26CAS |
Nilsson LO, Wallander H, Gundersen P (2012) Changes in microbial activities and biomasses over a forest floor gradient in C-to-N ratio. Plant and Soil 355, 75–86.
| Changes in microbial activities and biomasses over a forest floor gradient in C-to-N ratio.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xns1eks7Y%3D&md5=c6d7d3b51dd8c899887f4d8fbf8229d1CAS |
Parton WJ, Schimel DS, Cole CV, Ojima DS (1987) Analysis of factors controlling soil organic matter levels in Great Plains grasslands. Soil Science Society of America Journal 51, 1173–1179.
| Analysis of factors controlling soil organic matter levels in Great Plains grasslands.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL2sXmtlGnsbw%3D&md5=83adee9d531a52b4c726cce830d3407cCAS |
Rousk J, Bååth E (2007) Fungal and bacterial growth in soil with plant materials of different C/N ratios. FEMS Microbiology Ecology 62, 258–267.
| Fungal and bacterial growth in soil with plant materials of different C/N ratios.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhsValtr7I&md5=7be35778c24cc413772c807c975a0e6cCAS | 17991019PubMed |
Rousk J, Demoling LA, Bahr A, Bååth E (2008) Examining the fungal and bacterial niche overlap using selective inhibitors in soil. FEMS Microbiology Ecology 63, 350–358.
| Examining the fungal and bacterial niche overlap using selective inhibitors in soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXivFeqs7w%3D&md5=238cf208a867f049ee3518dc396322b9CAS | 18205814PubMed |
Rousk J, Brookes PC, Bååth E (2009) Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization. Applied and Environmental Microbiology 75, 1589–1596.
| Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXjsFKgsLo%3D&md5=90c606e8fe28332ea97e73b1cc889256CAS | 19151179PubMed |
Schimel JP, Gulledge JM, Clein-Curley JS, Lindstrom JE, Braddock JF (1999) Moisture effects on microbial activity and community structure in decomposing birch litter in the Alaskan taiga. Soil Biology & Biochemistry 31, 831–838.
| Moisture effects on microbial activity and community structure in decomposing birch litter in the Alaskan taiga.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXjtFSrsL4%3D&md5=c864454d0c297a2ba93b9180f4e73f9bCAS |
Söderström BE (1977) Vital staining of fungi in pure cultures and in soil with fluorescein diacetate. Soil Biology & Biochemistry 9, 59–63.
| Vital staining of fungi in pure cultures and in soil with fluorescein diacetate.Crossref | GoogleScholarGoogle Scholar |
Steinweg JM, Plante AF, Conant RT, Paula EA, Tanaka DL (2008) Patterns of substrate utilization during long-term incubations at different temperatures. Soil Biology & Biochemistry 40, 2722–2728.
| Patterns of substrate utilization during long-term incubations at different temperatures.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXht1KnsrjE&md5=92a6fe666979670c578399700fd8b1a3CAS |
Teklay T, Nordgren A, Nyberg G, Malmer A (2007) Carbon mineralization of leaves from four Ethiopian agroforestry species under laboratory and field conditions. Applied Soil Ecology 35, 193–202.
| Carbon mineralization of leaves from four Ethiopian agroforestry species under laboratory and field conditions.Crossref | GoogleScholarGoogle Scholar |
Thiet RK, Frey SD, Six J (2006) Do growth yield efficiencies differ between soil microbial communities differing in fungal: bacterial ratios? Reality check and methodological issues. Soil Biology & Biochemistry 38, 837–844.
| Do growth yield efficiencies differ between soil microbial communities differing in fungal: bacterial ratios? Reality check and methodological issues.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XivVaqt78%3D&md5=e13183bdf3d5768a50f3228dfcfc179eCAS |
Vance ED, Brookes PC, Jenkinson DS (1987) An extraction method for measuring microbial biomass C. Soil Biology & Biochemistry 19, 703–707.
| An extraction method for measuring microbial biomass C.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL1cXjs1KqsA%3D%3D&md5=619d5036dacd709b54824b460cae8c88CAS |
Xu RK, Ji GL (2001) Effects of H2SO4 and HNO3 on soil acidification and aluminum speciation in variable and constant charge soils. Water, Air, and Soil Pollution 129, 33–43.
| Effects of H2SO4 and HNO3 on soil acidification and aluminum speciation in variable and constant charge soils.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXlsVGjtL8%3D&md5=62d6efae0a5a97999a015c3646a45baaCAS |
Yeomans JC, Bremner JM (1988) A rapid and precise method for routine determination of organic carbon in soil. Communications in Soil Science and Plant Analysis 19, 1467–1476.
| A rapid and precise method for routine determination of organic carbon in soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL1cXlt1Oru7w%3D&md5=d82b7e08c8f3f16790508facae7221c1CAS |
Zak DR, Ringelberg DB, Pregitzer KS, Randlett DL, White DC, Curtis PS (1996) Soil microbial communities beneath Populus grandidentata grown under elevated atmospheric CO2. Ecological Applications 6, 257–262.
| Soil microbial communities beneath Populus grandidentata grown under elevated atmospheric CO2.Crossref | GoogleScholarGoogle Scholar |
Zhang NL, Wan SQ, Li LH, Bi J, Zhao MM, Ma KP (2008) Impacts of urea N addition on soil microbial community in a semi-arid temperate steppe in northern China. Plant and Soil 311, 19–28.
| Impacts of urea N addition on soil microbial community in a semi-arid temperate steppe in northern China.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhtVyju7rK&md5=08d813e0c84b992a26368e62453c4a2fCAS |
Zhao X, Xing G (2009) Variation in the relationship between nitrification and acidification of subtropical soils as affected by the addition of urea or ammonium sulfate. Soil Biology & Biochemistry 41, 2584–2587.
| Variation in the relationship between nitrification and acidification of subtropical soils as affected by the addition of urea or ammonium sulfate.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhtlGitr7N&md5=692f27f287ab4f9d7247979f1f6e625fCAS |