

Uptake and translocation of inorganic and methylated arsenic species by plants
Andrea Raab A , Paul N. Williams B , Andrew Meharg B and Jörg Feldmann A CA University of Aberdeen, School of Engineering and Physical Science, Meston Walk, Aberdeen, Scotland, AB24 3UE, UK.
B University of Aberdeen, School of Biosciences, Tillydrone Ave, Aberdeen, Scotland, AB24 2TZ, UK.
C Corresponding author. Email: j.feldmann@abdn.ac.uk
Environmental Chemistry 4(3) 197-203 https://doi.org/10.1071/EN06079
Submitted: 13 December 2006 Accepted: 23 April 2007 Published: 22 June 2007
Environmental context. The molecular occurrence of arsenic in soils can vary as a result of soil conditions and/or application of arsenic-containing herbicides or fertiliser. Although large amounts of As-containing herbicides are used for different crops, there is still a lack of understanding as to how the molecular form of As determines the uptake of arsenic into plants and, in particular, the translocation into shoot and grain.
Abstract. The uptake and translocation into shoots of arsenate, methylarsonate (MA), and dimethylarsinate (DMA) by 46 different plant species were studied. The plants (n = 3 per As species) were exposed for 24 h to 1 mg of As per litre under identical conditions. Total arsenic was measured in the roots and the shoots by acid digestion and inductively coupled plasma mass spectrometry from which, besides total As values, root absorption factors and shoot-to-root transfer factors were calculated. As uptake into the root for the different plant species ranged from 1.2 to 95 (μg of As per g of dry weight) for AsV, from 0.9 to 44 for MAV and from 0.8 to 13 for DMAV, whereas in shoots the As concentration ranged from 0.10 to 17 for AsV, 0.1 to 13 for MAV, and 0.2 to 17 for DMAV. The mean root absorption factor for AsV (1.2 to 95%) was five times higher than for DMAV (0.8 to 13%) and 2.5 times higher than for MAV (0.9 to 44%). Although the uptake of arsenic in the form of AsV was significantly higher than that of MAV and DMAV, the translocation of the methylated species was more efficient in most plant species studied. Thus, an exposure of plants to DMAV or MAV can result in higher arsenic concentrations in the shoots than when exposed to AsV. Shoot-to-root transfer factors (TFs) for all plants varied with plant and arsenic species. While AsV had a median TF of 0.09, the TF of DMAV was nearly a factor of 10 higher (0.81). The median TF for MAV was in between (0.30). Although the TF for MAV correlates well with the TF for DMAV, the plants can be separated into two groups according to their TF of DMAV in relation to their TF of AsV. One group can immobilise DMAV in the roots, while the other group translocates DMAV very efficiently into the shoot. The reason for this is as yet unknown.
Introduction
Arsenic, either naturally occurring or as a result of anthropogenic influences, is one of the biggest environmental nemeses worldwide. The health of millions of people on all continents is endangered because of arsenic present in water and soil.[1,2] While a large amount of research goes into clean-up strategies for drinking water, not that much thought is given to the problems arising from contamination of soil and the growth of plants on this land. Soil contamination may occur either directly or indirectly, as a result of irrigation with arsenic-containing water, and use of As-containing fertilisers or As-containing pesticides. Arsenic contamination of soil can have direct consequences on crop yield, e.g. straight-head disease of rice, and on the other hand reduces the crop value if the arsenic concentration is over the permissible limits.[3,4] In Europe, permissible limits for As in fodder is generally between 2 and 4 mg of As per kg (moisture content 12%), for food plants a statutory limit of 1 mg As per kg of dry weight (d.w.) exists, e.g., in Spain.[5–9] The EU has not yet set maximum limits for arsenic in vegetables.
The varied molecular species of arsenic, in which it can be present in the environment, adds to the complexity. Inorganic arsenic (tri- and pentavalent) is the predominant form in most waters, while soils can contain arsenic oxides and sulfides. As-based herbicides and pesticides (such as monosodiummethyl arsonate (MSMA) and dimethylarsinic acid (DMA)) or arsenic-containing fertiliser such as chicken manure from Roxarsone fed chicken, have been applied to soil.[10,11] Besides these man-made organoarsenicals, bacteria, fungi and others are able to generate methylated arsenicals from inorganic arsenic species, which can be re-excreted by the organism and taken up by others such as plants.
It is well known that plants take up inorganic arsenic in its tri- and pentavalent forms. The uptake mechanism for pentavalent arsenic is mostly via the high-affinity phosphate transporters and the amount can be influenced to some extent by phosphate application to the soil.[12] Trivalent arsenic is most likely taken up via glycerol transporters, which are not influenced by the amount of phosphate present.[13] Since tri- and pentavalent arsenic are easily interchangeable in the soil environment the uptake of these arsenic species can, therefore, only be limited to some extent by the application of phosphate. The uptake mechanisms for mono- and dimethylated arsenic species, either via the root system or via the shoot system, are as yet not known, but it is known that inorganic and methylated-As species are taken up via both root and shoot.[14]
There is a range of field surveys and greenhouse experiments about total arsenic concentration present in food crops and other plants available in the literature.[15] What is missing so far is a study that compares the uptake of different arsenic species via the root system under controlled, identical conditions so that comparisons can be made between plant species and whole families.
Here we present for the first time the arsenic uptake from hydroponic solution after short-term exposure of 46 plant species of the three major pentavalent arsenic species in the soil environment in order to identify whether the molecular form of arsenic governs the uptake of arsenic and its translocation into the shoot or whether every plant species shows different behaviour to the different arsenic forms.
Materials and methods
Plants
Plants were grown from commercially available seeds, with the exception of Thunbergia grandifolia and Pelargonium L’Hér. ex Ait., which were available from cuttings, and the arum species, which were grown from offshoots. Different species of the following families were grown: Apiaceae, Araceae, Asteraceae, Brassicaceae, Convolvulaceae, Cucurbitaceae, Cyperaceae, Fabaceae, Geraniaceae, Liliaceae, Poaceae, Solanaceae, Thunbergiaceae, Tropaeolaceae (for plant species and variety see Table 1). All plants were grown in a greenhouse under a 14-h light/10-h dark cycle at 18/16°C (80% relative humidity) in vermiculite (As concentration 0.4 ± 0.05 μg g–1). Growth time varied by species (6 to 20 weeks) and was dependent on available biomass (especially root) being large enough for the intended experiments. Plants were fertilised once a week (As concentration in conc. fertiliser 0.38 ± 0.027 μg g–1, dilution before use 1 : 1000). Fertilisation was stopped 1 week before exposure to arsenic. Teosinte was grown in a growth cabinet under the same conditions as all other plants, in accordance with demands of the supplier, all material was disinfected after use and biological material was burned as hazardous biological waste.[16] Before exposure the roots were washed free of vermiculite and then exposed to an arsenic-containing solution for 24 h. The plants were exposed via their root system to 100 mL of 1 mg L–1 arsenic in the form of arsenate (AsV), methylarsonic acid (MAV), or dimethylarsinic acid (DMAV). After 24 h the roots were washed using tap water (As concentration < 1 μg L–1) (10 min), immersed for 10 min in 10 × 10–3 M KH2PO4 solution (pH 6.0) to remove external inorganic arsenic from the root surface and blotted dry. The exposure time was limited to 24 h to reduce the influence of toxicity symptoms on uptake rates, since for most plants no toxicity data are available. The experiments were not designed to simulate field exposure, but to compare the behaviour of a range of plant species under identical conditions. Treatment with KH2PO4 solution removes most or all of the adhering external inorganic arsenic, but does not remove external methylated-arsenic species (A. Raab, unpubl. obs.). There is as yet no treatment known to remove externally absorbed methylated-As species from the root surface. The plants were separated into root and shoot, which were individually weighed and ground in liquid nitrogen. Subsamples of each plant (root and shoot separately) were then used for the determination of total arsenic and determination of dry weight.

|
Chemicals
All chemicals used were of analytical grade or better, except where mentioned otherwise. Stock solutions (1 g As/L) of arsenate AsV (Na2HAsO4·7H2O, 98%, BDH UK), methylarsonate MAV ((CH3)AsO3Na2, 98%, ChemService USA), and dimethylarsonate DMAV ((CH3)2AsO2Na), 98%, Strem Chemicals UK) were prepared in ultrapure water and diluted before exposure of plants. Nitric acid (65%, Fluka) and hydrogen peroxide (30%, Fluka) used for digestion of plant material were of suprapure grade. Plant material and certified reference material (IAEA 140, seaweed) was digested on a hotplate (95°C, 8 h) using 100 mg (FW) plant material, 1 mL of conc. nitric acid, and 1 mL of hydrogen peroxide.
Ultrapure water was used for dilutions and standard preparation. Rhodium was used as an internal standard. All samples were measured using external calibration. IAEA 140 (Fucus, LGC Promochem UK) was used as certified reference material, certified value 44.3 (42.2–46.4) mg kg–1 found 43.1 ± 0.8 mg kg–1 (n = 30).
Instrumentation
Arsenic was determined at m/z 75 using inductively coupled plasma mass spectrometry (ICP-MS) (Agilent 7500c) in normal mode. Chloride interference and selenium were measured at m/z 77 and 82, and rhodium was used as an internal standard and measured at m/z 103. Statistical calculations were performed using SigmaPlot V10.0/SigmaStat 3.5. Shoot/root transfer factors (TFs) were calculated from the total concentration of As in root and shoot dry weight (d.w.) and expressed as a ratio of As (shoot d.w.) to As (root d.w.). Root absorption factors (RAIs) were calculated from total As (root d.w.)/amount of As present in solution and expressed as a percentage.
Results and discussion
Total arsenic concentration in root and shoot
The plants of each species used for the exposure experiments were of similar size and weight within the species (P > 0.05) but not between species. Plants were used before onset of flowering.
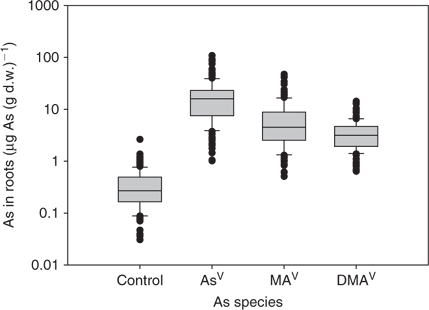
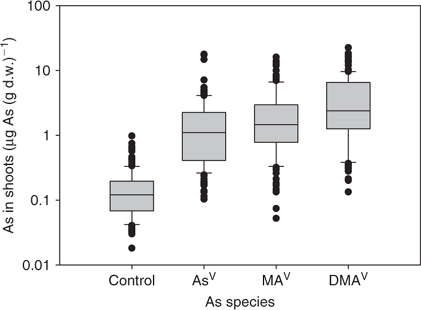
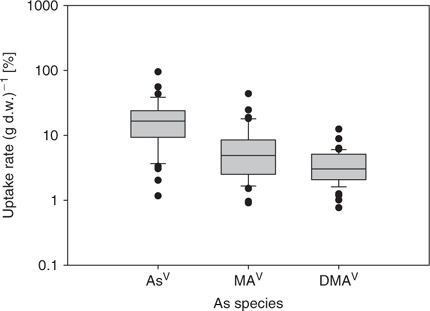
Mean total As in root was 18.5, 6.3 and 3.5 μg As (g d.w.)–1 for AsV, MAV, and DMAV, respectively. In shoot, on average 1.9, 2.4, and 4.0 μg As (g d.w.)–1 for AsV, MAV, and DMAV were found, respectively, after 24-h exposure to As in hydroponic solution. Background levels for arsenic in root were on average 0.36 μg As (g d.w.)–1 and shoot 0.16 μg As (g d.w.)–1. In Table 1 the mean ± s.d. values for each plant species are given. In Figs 1 and 2 the average concentrations are shown as box-plots for root (Fig. 1) and shoot (Fig. 2) (showing 5th and 95th percentile and the median line). RAIs were higher for inorganic arsenic than for methylated-As species (Fig. 3, showing 5th and 95th percentile and the median line). Plants absorbed on average about half the amount of MAV and a fifth of DMAV compared with AsV absorption. The highest RAI was shown by T. alata with 95% (of the available As) uptake per gram d.w. RAI factors are influenced by As adsorption onto the surface of the roots and absorption, which is the actual uptake of As into the root cells.
|
|
|
As hypothesised the uptake of arsenic is dependent on arsenic species present in the hydroponic solution and plant species. In general, methylated-arsenic species are less well taken up by plants than inorganic arsenic, but better translocated into shoots. Arsenic concentration in root was not related to As concentration in shoot after exposure to any of the tested arsenic species. Plant species with higher uptake of AsV into roots tended to have higher uptake of MAV and DMAV into roots as well. There was a strong linear relationship between the total As concentration of the different molecular forms of As in shoots of plants (DMAV/MAV r = 0.82, P < 0.005, DMAV/AsV r = 0.64, P < 0.005, MAV/AsV r = 0.62, P < 0.005). On average the concentration ratio between AsV and MAV was 3.8 and AsV and DMAV was 5.8 in root. In shoot these ratios were much lower (AsV/MAV was 0.97 and AsV/DMAV 0.61).
The reasons for this difference in uptake efficiency between AsV, MAV, and DMAV are most likely to be found in the different uptake mechanisms. Where high-affinity phosphate transporters (which are essential for the plant and can be regulated depending on phosphate status and, potentially, in response to As exposure as well) take up AsV, the uptake mechanisms for MAV and DMAV are not known. Even under sufficient phosphate nutrition AsV can be taken up in the form of AsIII after reduction in the soil/root environment.
Rosen et al. recently proposed a mechanism for the cellular uptake of MAIII in yeast.[17] Should MAV be taken up by plants in a similar way the lower As concentration in root after exposure to MAV would be understandable, since then MAV would first need to be reduced to MAIII at the root/soil interface. Differences in redox potential between AsV and MAV could then, among other things, explain the differences in uptake efficiency for these two As species. There is as yet no mechanism known for the cellular uptake of DMAV or DMAIII.
Studies concentrating on As uptake by plants have mostly only comprised one or two plant species, using different culture conditions from study to study. Our study is the first one to use the same culture conditions for a whole range of plant species, so that As uptake of different plant species can be compared with each other. Results published by Carbonell et al. and Marin et al. are difficult to compare with ours since the culture conditions for plants were different in each case.[18–20] However, generally it can be said that all published data show, as do our results, that soluble inorganic As is better taken up by plants than methylated-As species independent of plant species and culture conditions. Tlustos et al. found no difference in As uptake by radish plants grown in AsIII, AsV or DMAV amended soil.[21] Their explanation for this is the ability of inorganic As to bind more efficiently to soil particles than DMAV and the, therefore, higher phytoavailability of DMAV. The solubility (phytoavailability) of different As species is dependent on soil composition and redox conditions in the soil.[22] These factors are difficult to control and were, therefore, excluded from our experimental setup. Thus our data represent a ‘worst case’ scenario, under which all applied arsenic was phytoavailable.
Shoot-to-root TFs
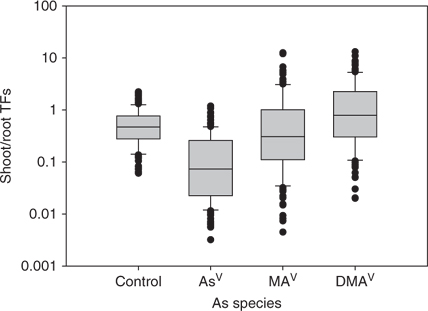
Mean shoot-to-root TFs for all exposed plants vary between 0.18 for AsV and 1.8 for DMAV. The TF for MAV was 1.0, about half that of DMAV. A box-plot diagram of the TFs for all plants is given in Fig. 4 showing the 5th and 95th percentile and the median line. Plants exposed to background levels transferred on average half the amount of arsenic into the shoot as they retained in the root (TF 0.6) (Table 2, TF average (n = 3) for each plant species and As species).
|

|
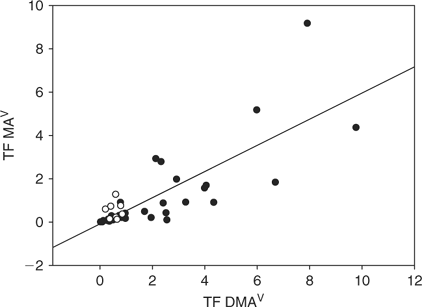
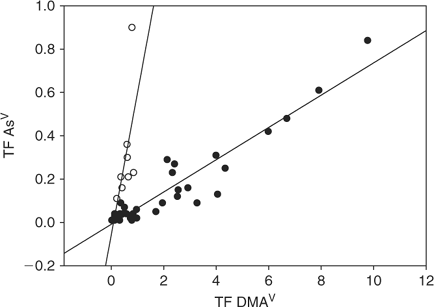
Translocation of arsenic into shoot was higher in plants exposed to DMAV than MAV or AsV. Plants with a high TF for DMAV also had generally a higher TF for MAV (Fig. 5). The same cannot be said for AsV, as certain plants (referred to in the following as group I) showed higher TF(AsV) than group II, with a similar TF(DMAV) in both groups (Fig. 6). Plants of group I showed a higher and more variable TF(AsV) (0.1 to 0.9) than others with a similar TF(DMAV). The TF(DMAV) of group I varied between 0.1 and 1, while for group II the TF(DMAV) varied between 0.1 and 10. Group I included Callistephus chinensis Cass. (L.) Nees and Lactuca sativa L. of the Asteraceae family, Cucumis sativus L. and of the Poaceae family: Avena sativa var. Melys, Panicum miliaceum L., Triticum urartu Thumanian ex Gandilyan and Melica aristata. Group II includes all other tested plant species. Since TF(MAV) and TF(DMAV) correlate (Fig. 5), it is apparent that group I plants are able to trap the methylated-As species in the roots rather than translocate them into the shoot. The effect is not dependent on plant family and the reasons are not known at the moment, but will be investigated in more detail.
|
|
None of the studied plant species had a shoot-to-root TF that exceeded 0.9 for AsV, whereas TFs for MAV and DMAV tended to be 6 to 10 times higher. TF values for AsV-exposed plants ranged from 0.01 (in T. grandifolia and Vicia faba) to 0.84 (in Nicotiana sanderae). Pickering et al. found for a variety of Brassica juncea grown under hydroponic conditions with phosphate (for 2 or 5 days with 18.75 mg L–1 As) a TF of < 0.1 for inorganic As, which is considerably lower than the TF factor of 0.4 found in this study for another variety of the same species.[12] The longer exposure time and different As concentration used during the experiment in addition to the different varieties are probably the reasons for the different TFs for B. juncea. Marin et al. found TFs for inorganic As of < 0.2 for rice cultivars, which are similar to the TF found for the two cultivars used in this study (TF(AsV) 0.015 and 0.008).[20]
The TFs of MAV and DMAV from shoot to root are generally higher, which might contribute to their higher phytotoxicity and, therefore, better herbicidal activity.[20] This observation is supported by Marin et al. who found that straighthead disorder of rice is increased by exposure to MAV.[20] The highest TF for MAV was found in Eruca versicaria with a value of 7.8. For DMAV the highest TF was seen for N. sanderae with 9.0, a plant species that generally showed signs of high accumulation potential and high TF.
One factor that influences the TF for the different As species is the ability of plants to complex inorganic arsenic as As–phytochelatin (PC) complexes. In experiments with Helianthus annuus it was reported that the formation of these complexes was predominantly in the root system.[23] Their formation rate is time and As-concentration dependent but nevertheless formation of these complexes starts as soon as As-exposure occurs.[23] Since As–PC complexes seem not, as such, to be transport forms of As (none were found in sap samples of H. annuus and B. juncea), their formation might reduce the translocation of inorganic arsenic.[12,23] A fact supported by the ‘virtually none’ existence of As–PC complexes in known As-hyperaccummulators Pteris vittata and P. cretica.[24] Raab et al. reported the identification of a MAIII–PC complex in H. annuus,[23] but it seems that MAV is less likely bound by PCs and/or their synthesis is not encouraged by MAV (A. Raab, unpubl. obs.). DMAIII–peptide complexes, like the one that can be synthesised from glutathione as DMAIII–GS, have not yet been observed in biological samples and are known to be very unstable.[25–27] It is, therefore, highly unlikely that DMAIII–PC complexes are formed by plants, which might offer a possible explanation for the high TF of DMAV. Only in sulfur-rich plant species of the genus Brassicaceae it has been recently found that DMAV can be complexed by glutathione as a DMASV–GS complex.[28]
The difference in complex formation between AsV and MAV on the one side, and DMAV on the other, could explain why plant species of group I and II fall within the same category when comparing TF(MAV) with TF(AsV).
Carbonnell et al. considered the TF to be dependent on the general As sensitivity of the plant species.[9] From the results of our study this cannot be confirmed, since the strain of Holcus lanatus included in the experiments was a tolerant strain for inorganic arsenic as shown by Meharg et al. but nevertheless the TFs were 0.1 for AsV, 0.9 for MAV, and 3.0 for DMAV, which are well within the range found for other non-tolerant plant species here.[29]
Conclusion
In plants hydroponically exposed to As, the uptake of inorganic arsenic into root is higher than the uptake of mono- or dimethylated As. Most of the studied plant species (n = 46) have higher transfer factors for methylated-As species than for inorganic As. Part of the reason for the better translocation of methlyated-As species might be the reduced complex formation with ligands such as glutathione or phytochelatins in roots. In general, it can be concluded that plants exposed to methylated arsenicals, either in soils with a high methylation rate of inorganic arsenic or in soils treated with MAV- or DMAV-containing herbicides, transport more As into the shoots than plants exposed to inorganic arsenic. This means that the soil conditions (soils that promote the methylation of As) or the application of methylated-arsenic herbicides may result in higher As levels in shoots of plants, which are most often the edible part of the plant. This aspect is of extreme interest, especially when herbicides are used on agricultural soils, which are used directly or indirectly for food production.
[1]
P. L. Smedley,
D. G. Kinniburgh,
Appl. Geochem. 2002, 17, 517.
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
| Crossref | GoogleScholarGoogle Scholar |
