

Is site preference of N2O a tool to identify benthic denitrifier N2O?
Aurélie Mothet A C , Mathieu Sebilo A , Anniet M. Laverman B , Véronique Vaury A and André Mariotti AA UPMC Univ Paris 06, UMR Bioemco, 4 place Jussieu, Cedex 05, F-75252 Paris, France.
B UPMC CNRS Univ Paris 06, UMR Sisyphe, 4 place Jussieu, Cedex 05, F-75252 Paris, France.
C Corresponding author. Email: aurelie.mothet@upmc.fr
Environmental Chemistry 10(4) 281-284 https://doi.org/10.1071/EN13021
Submitted: 30 January 2013 Accepted: 25 June 2013 Published: 16 August 2013
Environmental context. The greenhouse gas nitrous oxide is produced by bacteria and emitted from terrestrial and aquatic environments; the origin of this compound can be determined by its 15N intramolecular distribution (site preference). The site preference of nitrous oxide was characterised experimentally in bacterial denitrifying communities under controlled conditions. This study shows the importance of the last step of denitrification on the site preference values, and that complementary methods are necessary to identify the sources of nitrous oxide.
Abstract. Site preference values of nitrous oxide emitted during different steps of benthic denitrification were determined. Compared to that of nitrous oxide as end product, the site preference during complete denitrification presents a large variation, due to the final step, and is highly correlated with nitrate reduction rate. The nitrous oxide reduction step appears decisive on the site preference values.
Nitrous oxide is a potent greenhouse gas that participates in ozone layer destruction.[1] Atmospheric N2O concentrations have increased ~20 % with the rise of anthropogenic activities, such as the intensive use of fertilisers in agriculture areas.[2] In soils and sediments, the two main processes responsible for N2O emissions are nitrification[3] and denitrification.[4] Mediated by denitrifying bacteria under anoxic conditions, denitrification emits localised and instantaneous N2O as an intermediate product by reduction of nitrate (NO3–) to dinitrogen (N2).[5] The linear asymmetric N2O molecule has a site preference (SP) that is defined by the intramolecular 15N difference between Nα (central N) and Nβ (terminal N).[6] Determined by natural stable isotope ratios (δ15N and δ18O), the N2O site preference (N2O-SP) signature has been proposed and used as an emerging tool to define the origin of N2O.[7,8] Pure culture studies, using denitrifying bacteria that reduce NO3– to N2O, have shown that the N2O-SP signature for denitrification ranged between –5 and 0 ‰.[7,8] In addition to this, several denitrification studies have investigated the N2O-SP signature originating from soil and water incubations either with N2O as an end product, substrate or intermediate and found a large variation between –8.5 to 81 ‰.[9,10] Part of this variation has been shown to be due to the fact that the N2O consumption during denitrification generates an increase of the SP values.[10,11]
Even though these investigations have increased the knowledge regarding the N2O-SP signature associated with denitrification, several questions remain unanswered. The N2O-SP signature of environmental denitrifying communities is, for example, lacking. In order to bridge the gap between the investigations on denitrification in pure cultures, the focus of this study was to determine N2O-SP signatures of environmental benthic denitrifying communities under controlled conditions. More specifically, we determined SP values of N2O produced by an environmental benthic denitrifying community during the different steps of benthic denitrification (either the production of N2O or during complete denitrification) under denitrifying conditions (anoxia, nitrate supply).
The study was conducted in an open system using flow-through reactor (FTR) experiments.[12] The use of these reactors allows the determination of denitrification-derived N2O by nitrate supply and creating anoxic conditions, and thereby exclude nitrification as an additional N2O source. Nitrate reduction rates and the accompanying N2O-SP signatures were determined simultaneously, allowing direct comparison.
During benthic denitrification in two types of sediments, nitrate reduction rates and the N2O-SP values were determined in an open system using FTRs (Fig. 1). The FTRs are described in detail in Laverman et al.[12] Sediments were collected from the Charmoise River (48°36′37.80″N, 2°9′19.54″E, Essonne, France), which is 7.5 km long, in October and November 2011 and in the Manche-à-Eau lagoon (16°16′38.04″N, 61°33′27.93″W, Guadeloupe, France), which has an area of 260 000 m2, in March 2012. In FTR experiments, denitrifying conditions were obtained by supplying anoxic (N2 bubbled) nitrate solutions (infinite reservoir) to the sediments.
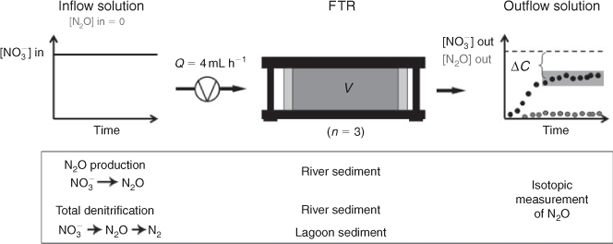
|
In order to determine the SP value of N2O as end product during NO3– reduction to N2O, by the environmental benthic denitrifying community, acetylene (C2H2) was used to block N2O reductase, the enzyme responsible for the N2O reduction to N2.[13] In addition to the determination of the SP of N2O as an end product, the SP of N2O appearing as an intermediate was measured during different microbial denitrification rates in two types of sediments (river and lagoon). In order to obtain different nitrate reduction rates, diverse nitrate concentrations were supplied to the sediment by the inflow solutions (Fig. 1). During NO3– reduction to N2O (acetylene block) 5 mM nitrate was supplied, while a range of concentrations was applied during benthic denitrification in river sediment (1, 1.5, 3 and 5 mM) and in lagoon sediment (0.83, 1, 3 and 5 mM), in order to achieve different nitrate reduction rates. The denitrification rate depends not only on NO3– concentrations, as substrate, but also on organic carbon (Corg) content. Determined by the difference between the total carbon and inorganic carbon contents, the Corg content was on average 6.3 ± 0.07 % (n = 5) in the river sediment and 12.1 ± 0.04 % (n = 6) in the lagoon sediment. These amounts of carbon are sufficient to allow denitrification over the time the sediments were incubated in our experiments.[14]
Nitrate concentrations were determined by high-performance liquid chromatography (HPLC, Dionex, AS12 column; Thermo Scientific, Sunnyvale, CA, USA) for samples of benthic reduction of NO3– to N2O and by colourimetric measurements using an autoanalyser for samples of complete denitrification within river sediment (Quaatro, Bran & Luebbe, Plaisir, France) and within lagoon sediment (Gallery; Thermo Fisher Scientific, Cergy-Pontoise, France).
Prior to the determination of benthic denitrification derived N2O, the N2O-SP signature of the nitrate reduction to nitrous oxide by azide was tested with a laboratory standard. This method, which allowed the isotopic composition of nitrate and nitrite to be obtained, consists of nitrate reduction to nitrite through a column containing activated cadmium; consequently, nitrite is reduced to nitrous oxide by sodium azide solution (NaN3, δ15N = –2.9 ± –0.4, n = 10) and the last step (N2O to N2) is blocked by the addition of sodium hydroxide.[15]
All liquid samples were analysed using an isotope ratio mass spectrometer (IRMS, DeltaVplus; Thermo Scientific, Bremen, Germany) in continuous-flow with a purge-and-trap system and coupled with a Finnigan GasBench II system (Thermo Scientific), in order to obtain the stable isotope composition of N2O (δ15N–N2O, δ18O–N2O, δ15N–NO and δ18O–NO (Nα). The GasBench II allows sampling of the gases and removal of the water contained in the liquid samples, and finally separation of the interfering gases of N2O using a chromatographic column (CP-poraPLOT U; Thermo Scientific). The N2O-SP values were calculated based on the raw values of δ15N–bulk and δ31NO (δ15Nα–N2O), which were measured by the IRMS from the same N2O reference gas. Nitrate standards were used to calibrate the isotopic composition of N2O (USGS34, δ15N = –1.8 ‰, δ18O = –27.9 ‰, USGS35, δ15N = +2.7 ‰, δ18O = +57.5 ‰ and USGS32, δ15N = +180 ‰, δ18O = +25.7 ‰). The precision is 0.5 ‰ for δ15N and 1 ‰ for δ18O. Statistical analyses were conducted using SigmaStat with Spearman’s correlation; P < 0.05 was inferred as statistically significant.
Nitrate reduction to N2O by azide yielded an average N2O-SP value of 2.2 ± 0.9 ‰ (n = 35). Considering the small variation (±0.9 ‰) of the N2O-SP values of the laboratory standard, the NO3– reduction to N2O by azide is a safe technique to determine the isotopic composition of nitrate and nitrite.
The δ15N and δ18O of N2O as end product or as intermediate (δ15N = –44.3 to –21.5 ‰; δ18O = –10.6 to 30.2 ‰), obtained for 5 mM of nitrate supply, presented a value range consistent with that found in previous studies on soil denitrification (δ15N = –41 to 20 ‰; δ18O = –55 to 55 ‰).[16,17] However, considering these large ranges, the N2O-SP value determination may bring a stronger proof that denitrification occurs only in these sedimentary studies.
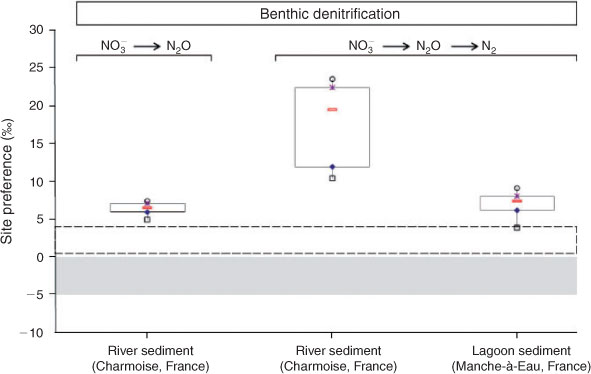
The results concerning the N2O-SP values for the different experiments at 5 mM of nitrate supply are shown in Fig. 2. Assuming that the acetylene block was complete and N2O was not further reduced to N2, the results during benthic denitrification indicated a low N2O-SP value (6.3 ± 1.0 ‰; n = 16), which was, however, higher than those from pure cultures (–5 to 0 %, Fig. 2).[7,8] The N2O-SP value derived from Pseudomonas chlororaphis and P. aureofaciens of 0 ‰[8] and of –5.1 ± 1.8 ‰ for P. denitrificans[7] suggest that these values vary among denitrifying species. Our data show that the environmental benthic denitrifying community generates N2O with a SP value different from those found in pure cultures.[7,8] Most likely the enzyme implicated in the transformation of NO to N2O (nitric oxide reductase, NOR) constrains the N2O-SP values[7] due to enzyme differences of the environmental benthic denitrifying community and may result in N2O-SP values deviating from the pure cultures tested so far. Despite the difference compared to pure cultures, the environmental benthic denitrifying community showed N2O-SP values in a narrow range, which confirms that the SP value of N2O as end product is constant during this process (NO3– to N2O).[18]
|
In the river sediments, N2O emitted during benthic denitrification showed higher average N2O-SP values, together with a large variability, (17.4 ± 6.5 ‰, n = 5) compared with those of N2O as an end product (6.3 ± 1.0 ‰, n = 16, Fig. 2). During the reduction step of N2O to N2, an enrichment of 15N in the central N-position (Nα) of the residual N2O occurs[19] and leads to an increase of the N2O-SP values.[10,11] This has been explained by the preferential consumption of 15Nβ14NαO relative to 14Nβ15NαO because the 15Nα–O bond is stronger than the 14Nα–O bond.[6] Thus, the increase of the N2O-SP values during the progress of denitrification will lead to a large variability of the N2O-SP values.
During benthic denitrification, the SP of N2O produced in the lagoon sediment, exhibiting an average of 6.8 ± 2.8 ‰ (n = 6), was lighter and less variable than that emitted from the river sediment (17.4 ± 6.5 ‰, n = 5, Fig. 2). This difference in N2O-SP values from the two sediments during complete denitrification is most likely due to the presence and activity of distinct denitrifying communities, similar to the differences between N2O-SP values obtained during NO3– reduction to N2O between environmental and pure cultures.
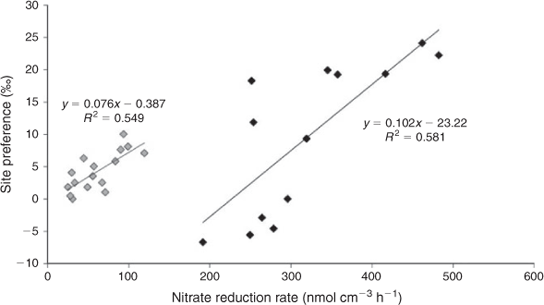
During biological NO3– reduction to N2O, the small variation in N2O-SP values (varying between 4.2 and 7.8 ‰) corresponded to a wide range of nitrate reduction rates (100 to 300 nmol cm–3 h–1). Thus, this result confirms that the step of NO3– reduction to N2O results in a specific low N2O-SP value independent of the nitrate reduction rate. On the contrary, in the two sediments investigated, the N2O-SP values exhibited a large variation during complete denitrification (Figs 2, 3).
|
Fig. 3 shows the N2O-SP values for the two sediments, obtained for the entire range of nitrate supply, indicating a significant correlation between nitrate reduction rates and the N2O-SP values (P = 0.0086; r = 0.48) during complete denitrification with N2O appearing as intermediate. In the lagoon sediment, the NO3– reduction rates are lower than those found in the river sediment. This statement consolidates the idea of the presence and the activity of distinct species of denitrifying communities in the two sediments. Moreover, the variation of N2O-SP values related to the variation of the NO3– reduction rate, during benthic denitrification in the two sediments, is in good agreement with previous studies that showed that the reduction of N2O to N2 leads to an increase of N2O-SP values.[10,11] In addition, the denitrification potential by the denitrifying bacteria is enhanced with the higher nitrate concentration and leads to a N2O reduction to N2. Thus, the last step of denitrification is crucial in the determination of the N2O-SP values in field studies where production and reduction of N2O occur simultaneously. On the contrary, without this last step of denitrification the values of N2O-SP will be invariant and close to those found by Toyoda et al.,[7] Sutka et al.[8] and this study for N2O production only.
This study showed that the SP of N2O as an end product of the benthic denitrifying communities is independent of the nitrate reduction rate and constant during the NO3– reduction to N2O step of denitrification. However, the SP value of N2O tends to increase during complete denitrification, with N2O as intermediate, and shows a strong positive correlation with the nitrate reduction rate. In fact, a high nitrate concentration enhances the denitrifier activity and increases the N2O-SP values by 15N enrichment of residual N2O during the last step in the denitrification process. Consequently, in field studies, during which production and reduction of N2O occur simultaneously, the SP of N2O can show a wide range of values, which does not depend on the composition of the denitrifying community only. This study underscores the importance of the last step of denitrification for the determination of the N2O-SP values and that care should be taken using the SP of N2O as the origin for denitrification in the case of field studies. In the identification of nitrous oxide sources, the determination of N2O-SP values should be associated with other methods such as the use of the natural abundances of isotopes and the isotopic enrichment techniques or the identification of the denitrifying species involved.
Acknowledgements
The authors acknowledge A. Boc and O. Gros for hosting us in their laboratory in Guadeloupe, O. Gros and C. Yan for assistance in the field and E. Aubry and E. Huché-Pillet for chemical analysis and isotopic measurements.
References
[1] P. J. Crutzen, The influence of nitrogen oxides on the atmospheric ozone content. Quart. J. R. Met. Soc. 1970, 96, 320.| The influence of nitrogen oxides on the atmospheric ozone content.Crossref | GoogleScholarGoogle Scholar |
[2] S. A. Montzka, E. J. Dlugokencky, J. H. Butler, Non-CO2 greenhouse gases and climate change. Nature 2011, 476, 43.
| Non-CO2 greenhouse gases and climate change.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXpslShsr4%3D&md5=bac44ca59cacfc74ddde32fa1e3cbea9CAS | 21814274PubMed |
[3] T. Yoshida, M. Alexander, Nitrous oxide formation by Nitrosomonas europaea and heterotrophic microorganisms. Soil Sci. Soc. Am. Proc. 1970, 34, 880.
| Nitrous oxide formation by Nitrosomonas europaea and heterotrophic microorganisms.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE3MXitVKntw%3D%3D&md5=b606dce9ad21157cda6946359c41cf94CAS |
[4] J. Wijler, C. C. Delwiche, Investigations on the denitrifying process in soil. Plant Soil 1954, 5, 155.
| Investigations on the denitrifying process in soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaG2cXktlOnsA%3D%3D&md5=9262e35bde71506012435bab39434f57CAS |
[5] M. K. Firestone, E. A. Davidson, Microbiological basis of NO and N2O production and consumption in soils, in Exchanges of Trace Gases Between Terrestrial Ecosystems and the Atmosphere (Eds M. O. Andreae, D. S. Schimel) 1989, pp. 7–21 (Wiley: New York).
[6] N. Yoshida, S. Toyoda, Constraining the atmospheric N2O budget from intramolecular site preference in N2O isotopomers. Nature 2000, 405, 330.
| Constraining the atmospheric N2O budget from intramolecular site preference in N2O isotopomers.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXjs1KltLw%3D&md5=a460c4de2f1d8869d5bfe002a0d543a8CAS | 10830958PubMed |
[7] S. Toyoda, H. Mutobe, H. Yamagishi, N. Yoshida, Y. Tanji, Fractionation of N2O isotopomers during production by denitrifier. Soil Biol. Biochem. 2005, 37, 1535.
| Fractionation of N2O isotopomers during production by denitrifier.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXltFSksb0%3D&md5=d95b1cce3f3282e75ca210bef2ae46ffCAS |
[8] R. L. Sutka, N. E. Ostrom, P. H. Ostrom, J. A. Breznak, H. Gandhi, A. J. Pitt, F. Li, Distinguishing nitrous oxide production from nitrification and denitrification on the basis of isotopomer abundances. Appl. Environ. Microbiol. 2006, 72, 638.
| Distinguishing nitrous oxide production from nitrification and denitrification on the basis of isotopomer abundances.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XmtFeitA%3D%3D&md5=f41c394e657000f9f7fe50544bcc79eeCAS | 16391101PubMed |
[9] R. Bol, T. Röckmann, M. Blackwell, S. Yamulki, Influence of flooding on δ15N, δ18O, 1δ15N and 2δ15N signatures of N2O released from estuarine soils–a laboratory experiment using tidal flooding chambers. Rapid Commun. Mass Spectrom. 2004, 18, 1561.
| Influence of flooding on δ15N, δ18O, 1δ15N and 2δ15N signatures of N2O released from estuarine soils–a laboratory experiment using tidal flooding chambers.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXlvFegs7g%3D&md5=da69a5417939b0d005e12bc460e7458dCAS | 15282780PubMed |
[10] R. Well, H. Flessa, F. Jaradat, S. Toyoda, N. Yoshida, Measurement of isotopomer signatures of N2O in groundwater. J. Geophys. Res. 2005, 110, G02006.
| Measurement of isotopomer signatures of N2O in groundwater.Crossref | GoogleScholarGoogle Scholar |
[11] N. E. Ostrom, A. Pitt, R. Sutka, P. H. Ostrom, A. S. Grandy, K. M. Huizinga, G. P. Robertson, Isotopologue effects during N2O reduction in soils and in pure culture of denitrifiers. J. Geophys. Res. 2007, 112, G02005.
| Isotopologue effects during N2O reduction in soils and in pure culture of denitrifiers.Crossref | GoogleScholarGoogle Scholar |
[12] A. M. Laverman, P. van Cappellen, D. van Rotterdam-Los, C. Pallud, J. Abell, Potential rates and pathways of microbial nitrate reduction in coastal sediments. FEMS Microbiol. Ecol. 2006, 58, 179.
| Potential rates and pathways of microbial nitrate reduction in coastal sediments.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhtF2rtrvJ&md5=9e66a93ea212d6b03929ac2eca6615a0CAS | 17064260PubMed |
[13] T. Yoshinari, R. Hynes, R. Knowles, Acetylene inhibition of nitrous-oxide reduction and measurement of denitrification and nitrogen-fixation in soil. Soil Biol. Biochem. 1977, 9, 177.
| Acetylene inhibition of nitrous-oxide reduction and measurement of denitrification and nitrogen-fixation in soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE2sXkt1WntbY%3D&md5=467db299734a58a1a4f085d56b801ad0CAS |
[14] J. Abell, A. M. Laverman, P. van Cappellen, Bioavailability of organic matter in a freshwater estuarine sediment: long-term degradation experiments with and without nitrate supply. Biogeochemistry 2009, 94, 13.
| Bioavailability of organic matter in a freshwater estuarine sediment: long-term degradation experiments with and without nitrate supply.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhtVKktrjK&md5=6851dd66eed65bf85c10cad4671c4180CAS |
[15] P. Semaoune, M. Sebilo, J. Templier, S. Derenne, Is there any isotopic fractionation of nitrate associated with diffusion and advection? Environ. Chem. 2012, 9, 158.
| Is there any isotopic fractionation of nitrate associated with diffusion and advection?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XmsFGnt74%3D&md5=12824245a51c0b17705c8ac06b0dcab8CAS |
[16] J. Tilsner, N. Wrage, J. Lauf, G. Gebauer, Emission of gaseous nitrogen oxides from an extensively managed grassland in NE Bavaria, Germany – II. Stable isotope natural abundance of N2O. Biogeochemistry 2003, 63, 249.
| Emission of gaseous nitrogen oxides from an extensively managed grassland in NE Bavaria, Germany – II. Stable isotope natural abundance of N2O.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXjtVGrsbk%3D&md5=73dcdc67bb4bccb6d2fce2f1ad13f7feCAS |
[17] O. V. Menyailo, B. A. Hungate, Stable isotope discrimination during soil denitrification: production and consumption of nitrous oxide. Global Biogeochem. Cycles 2006, 20, GB3025.
| Stable isotope discrimination during soil denitrification: production and consumption of nitrous oxide.Crossref | GoogleScholarGoogle Scholar |
[18] B. N. Popp, M. B. Westley, S. Toyoda, T. Miwa, J. E. Dore, N. Yoshida, T. M. Rust, F. J. Sansone, M. E. Russ, N. E. Ostrom, P. H. Ostrom, Nitrogen and oxygen isotopomeric constraints on the origins and sea-to-air flux of N2O in the oligotrophic subtropical North Pacific gyre. Global Biogeochem. Cycles 2002, 16, 12-1.
| Nitrogen and oxygen isotopomeric constraints on the origins and sea-to-air flux of N2O in the oligotrophic subtropical North Pacific gyre.Crossref | GoogleScholarGoogle Scholar |
[19] C. C. Barford, J. P. Montoya, M. A. Altabet, R. Mitchell, Steady-state nitrogen isotope effects of N2 and N2O production in Paracoccus denitrificans. Appl. Environ. Microbiol. 1999, 65, 989.
| 1:CAS:528:DyaK1MXhslWkt7o%3D&md5=703ea42f1add014c90734df0607b767cCAS | 10049852PubMed |