

‘Moa’s Ark’ or ‘Goodbye Gondwana’: is the origin of New Zealand’s terrestrial invertebrate fauna ancient, recent, or both?
Gonzalo Giribet A C and Sarah L. Boyer BA Museum of Comparative Zoology, Department of Organismic and Evolutionary Biology, Harvard University, 26 Oxford Street, Cambridge, MA 02138, USA.
B Biology Department, Macalester College, 1600 Grand Avenue, Saint Paul, MN 55105, USA.
C Corresponding author. Email: ggiribet@oeb.harvard.edu
Invertebrate Systematics 24(1) 1-8 https://doi.org/10.1071/IS10009
Submitted: 10 March 2010 Accepted: 29 March 2010 Published: 17 May 2010
Abstract
Evaluating the hypothesis of New Zealand’s total submersion during the Oligocene requires the strictest tests, including sound phylogenetic data and dating of phylogenies. Although New Zealand has been traditionally considered to host ancient biota that originated by vicariance after it separated from Australia ~80 Mya, the ancient origins of its biota have been recently questioned, with some authors even suggesting that all current land organisms had to arrive to the islands after it re-emerged from the ocean 22 million years ago. Here we examine examples of short-range endemic soil-dwelling invertebrates and find compelling evidence that at least some of them are the result of old lineages that diversified in New Zealand before the hypothesised submersion event 22 million year ago. We conclude that New Zealand indeed has old lineages as well as recently diversified lineages and compare this situation with that of other more stable areas of the Neotropics.
Introduction
New Zealand is, without doubt, one of the most fascinating archipelagos in the world. Its geology and probably parts of its biota reflect a dynamic history of ancient Gondwanan origin, long-term isolation from other continental landmasses, marine inundation during the Oligocene, glaciation during the Pleistocene, and evolutionary radiations that have produced a spectacular proportion of endemic species (Gibbs 2006). New Zealand has received considerable attention from biogeographers, beginning with Ernst Dieffenbach, a German naturalist who lived there from 1839–1841 and corresponded with Charles Darwin about the biogeographical affinities of the islands’ birds. Researchers have focused on New Zealand’s biogeography with particular vigour over the past two decades as molecular systematics has provided new tools with which to approach evolutionary questions. Molecular systematists and biogeographers have addressed topics such as the number and location of Pleistocene refugia (e.g. Marske et al. 2009; Buckley et al. 2010), the Alpine Fault Hypothesis (e.g. Heads and Craw 2004) and, most contentiously, a vicariance- v. dispersal-based origin of New Zealand’s terrestrial biota (e.g. Trewick et al. 2007; Phillips et al. 2010).
Although researchers have long recognised that land area was drastically reduced (i.e. to less than 15% of its current size) during the marine incursions of the Oligocene (e.g. Cooper and Cooper 1995), more recently Waters and Craw (2006) suggested that there is no strong evidence for continuously emergent land through the period (see also Landis et al. 2008). Trewick et al. (2007) and Wallis and Trewick (2009) asserted that the preponderance of biogeographical evidence favours a scenario of complete submergence during the Oligocene, and some authors have suggested that the entire terrestrial biota arrived via dispersal during the last 22 million years (Landis et al. 2008).
Even if a large fraction of the terrestrial biota presently inhabiting New Zealand arrived via dispersal, there is compelling evidence for an archaic Gondwanan origin for some of New Zealand’s invertebrates – i.e. lineages that have been continuously present in New Zealand since its connection to other Gondwanan landmasses. Here, we review the debate and make our case for the importance of vicariance in the establishment of at least some of this unique fauna.
Moa’s Ark and Goodbye Gondwana
New Zealand originated as part of the former supercontinent Gondwana, which formed the southern portion of Pangaea during the Triassic. Gondwana broke into fragments starting ~165–160 Mya when the continental block formed by Madagascar and India began to rift from both Australia and Africa; 135–130 Mya the South Atlantic Ocean opened at the far southern tip of Africa and South America. It was not until 165–110 Mya that Africa and Antarctica finally separated. Approximately 88–84 Mya India and Madagascar rifted from one another, with India eventually colliding with Asia around 50 Ma. New Zealand, South America and Australia were originally connected across Antarctica; 80 Mya New Zealand began to drift away from West Antarctica, opening the Tasman Sea. Final terrestrial connections between New Zealand and Australia were broken by 60 Mya, although Australia and South America remained in contact across Antarctica until at least the early Eocene. At this time Antarctica had a temperate climate and was home to Nothofagus forests, but by 46 Mya cold conditions were severe enough that overland dispersal by temperate-adapted organisms would have been impaired (e.g. Scotese et al. 1988; reviewed in Sanmartín 2002). Southern South America and Antarctica still remained in contact through the Antarctic Peninsula until the opening of the Drake Passage at the Eocene–Oligocene boundary (30–28 Mya). This led to the establishment of the Antarctic Circumpolar Current and the onset of the first Antarctic glaciation (reviewed in Sanmartín 2002; Sanmartín and Ronquist 2004), which has influenced the New Zealand biota to certain extent.
New Zealand has long been considered to host a large number of lineages of Gondwanan origin, an idea captured by the phrase ‘Moa’s Ark’, the title of a popular documentary on the country’s natural history (Bellamy et al. 1990). The ‘Moa’s Ark’ paradigm is based on charismatic examples of supposed vicariance in ratite birds, chironomid midges, and the Southern beech, or on the presence of ancient relicts such as the tuatara and leiopelmatid frogs, with no close extant relatives outside of New Zealand (Trewick et al. 2007). However, recent reinterpretations of New Zealand’s geology and its fauna have led to very different conclusions, framed as the ‘Goodbye Gondwana’ paradigm (McGlone 2005), which questions the antiquity of the land surface and thus of its terrestrial biota, and postulates that virtually every terrestrial group has reached New Zealand through fortuitous over-water dispersal during the past 22 million years (Landis et al. 2008), with subsequent diversification (Waters and Craw 2006; Trewick et al. 2007; Goldberg et al. 2008).
Support for the ‘Goodbye Gondwana’ paradigm comes from different sources of evidence, including the fossil record (e.g. Pole 2001), but chiefly from molecular systematics and estimation of divergence times that for example provides evidence for trans-oceanic dispersal of the Southern beech (e.g. Cook and Crisp 2005b; Knapp et al. 2005), formerly the textbook example of a classic Gondwanan taxon (e.g. Swenson et al. 2001). More surprisingly, the monophyly of the flightless ratite birds (the moas, kiwis and their relatives, the emus, rheas, cassowaries and ostriches) has recently been questioned through a new analysis of mitochondrial genomic data, including New Zealand’s now-extinct moas, whose DNA was sequenced from exceptionally well-preserved bones in museum collections (Phillips et al. 2010). This analysis identified tinamous, which can fly and have not traditionally been included within the ratites, as the sister group to the flightless moas. If the most recent common ancestor of moas and tinamous could fly, it may no longer be necessary to invoke Gondwanan vicariance to explain their arrival on New Zealand – although the fact that an animal’s ancestor had the ability to fly does not necessarily indicate that vicariance has not played a role in shaping current distributions, as shown for example in forest dwelling birds (Weir et al. 2009).
More broadly, a recent compilation of molecular phylogenetic analyses of ~100 plant and animal groups suggests that only 10% of these are even plausibly of archaic origin predating the vicariant split of Zealandia from Gondwana (Wallis and Trewick 2009). They summarise current opinion on the extent of the Oligocene drowning episode thusly: ‘If further paleontological data reveal evidence of uninterrupted fossil records through the Oligocene, complete inundation is made less likely (Winkworth et al. 1999; Lee et al. 2009). Notwithstanding these gaps in our knowledge, many commentators have already debunked the concept of NZ as a Gondwanan relictual ark (McGlone 2005; Waters & Craw 2006; Trewick et al. 2007; Landis et al. 2008)’ (Wallis and Trewick 2009: p. 3565). The same authors do concede (p. 3565) that ‘[m]ore work on arthropods is needed; they are likely to include recent and ancient dispersers as well as Gondwanan relicts (Gressitt 1961, 1970; Boyer & Giribet 2007)’. In another paper Goldberg et al. (2008) review the molecular evidence for several New Zealand terrestrial animals, concluding that the New Zealand biota is ‘in most respects, more like that of an oceanic archipelago than a continent’ (p. 3319). Few have questioned this new trend in interpretation of New Zealand biogeography (Knapp et al. 2007; Edgecombe and Giribet 2008; Boyer and Giribet 2009; Allwood et al. 2010).
Interestingly, a similar argument has emerged for New Caledonia (Murienne et al. 2005; Grandcolas et al. 2008), also considered part of Zealandia, although in this case there is significant geological evidence supporting the total submergence of New Caledonia under the adjacent oceanic plate, followed by ophiolithic obduction in the Eocene. By contrast, the evidence of complete submersion for New Zealand is not as strong. Murienne (2009) recently summarised the three existing models for New Caledonia, also applicable to New Zealand. The ‘museum model’ implies that the island has acted as a museum and the origin and diversification of the New Caledonia lineages precedes a supposed drowning event. The two other models suppose that the diversification initiates after the land re-emerged; one with a series of ‘mountain refugia, island refugia and long-distance dispersal’ and another without refugia and explaining all the current diversity by long-distance dispersal. The author concluded that the three models might render identical phylogenetic patterns, making these models difficult to choose from, but molecular dating could at least discern between the first and last models. An additional model: a cryptic mass extinction, leaving a phylogenetic pattern indistinguishable from that of an evolutionary radiation, has been recently proposed as an alternative (Crisp and Cook 2009).
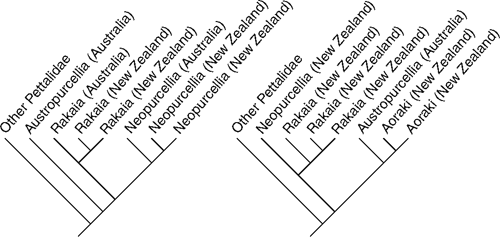
Dispersal hypotheses are difficult, if not impossible, to falsify, but easy to corroborate with a phylogenetic pattern that results in paraphyly of the source group (Edgecombe and Giribet 2008) (see Fig. 1). Hence, if one postulates that an endemic clade from New Zealand has originated in Australia, it should render its Australian counterparts paraphyletic, with the New Zealand lineage nested within (but see Cook and Crisp 2005a for a different interpretation). But more relevant, perhaps, is the recent use of molecular dating in phylogenetic analyses. Given enough data and even considering broad errors in dating techniques, it is now feasible to distinguish whether a lineage has originated before or after a postulated geological event. In the context of New Zealand, 22 Mya is the age that would distinguish the competing ‘Moa’s Ark’ and ‘Goodbye Gondwana’ scenarios. Therefore, in this article we revisit some cases of our ‘favourite’ organisms, arthropods and other related terrestrial invertebrates, to test the 22 Mya barrier. We are not arguing that Gondwanan vicariance is the mechanism explaining all or even the majority of the New Zealand biota. Rather, we are interested in identifying cases that firmly establish that New Zealand has acted, even if in few instances, as a biodiversity ark for Gondwanan taxa, and that some of them may even be considered relictual (e.g. Craterostigmomorpha, Sphenodontida).
|
Examples of supposed Gondwanan relicts in New Zealand
Many groups of terrestrial New Zealand animals show Gondwanan distributions, as summarised in previous reviews (e.g. Sanmartín and Ronquist 2004; Trewick et al. 2007; Goldberg et al. 2008), but a Gondwanan distribution is not sufficient for accepting a hypothesis of vicariance for New Zealand’s fauna (Trewick et al. 2007). Here, we focus on four groups of terrestrial invertebrates that plausibly arrived in New Zealand before the 22 Mya Oligocene drowning barrier: mite harvestmen, Craterostigmus centipedes, stone centipedes, and peripatus or velvet worms.
The reasons for choosing these examples are the existence of (1) resolved phylogenies and (2) molecular dating. These are also taxa that fall in the category of short-range endemics, characterised by Harvey (2002) as taxa that lack active dispersal mechanisms and rely on permanently moist micro-habitats, features that may preclude them from trans-oceanic dispersal. These characteristics may make the selected examples useful candidates for biogeographic study.
Mite harvestmen: Pettalidae (Fig. 2a–d)

|
Mite harvestmen (Arachnida, Opiliones, Cyphophthalmi), and the family Pettalidae in particular, have played a role in the study of Gondwanan and New Zealand biogeography. The family has a typical temperate Gondwanan distribution, with species in Chile, South Africa, Madagascar, Sri Lanka, Australia and New Zealand (but not in New Caledonia, which hosts a member of a tropical Gondwanan clade, the family Troglosironidae) (Boyer et al. 2007b; Boyer and Giribet 2007). Originally, New Zealand’s mite harvestmen were described in two genera, Rakaia and Neopurcellia, each of which also included species in Queensland, Australia, implying a shared evolutionary history spanning the Tasman Sea (Fig. 1). However, recent taxonomic revision by Boyer and Giribet (2007), based on a combined analysis of morphological and molecular data, refutes that hypothesis. The mite harvestmen of Queensland were found to be monophyletic and are now grouped in a single genus (Austropurcellia) with no New Zealand representatives. The New Zealand species are grouped into three genera: Aoraki (Fig. 2a) in the North and South islands, Rakaia (Fig. 2c, d) in the North, South and Stewart islands, and Neopurcellia (Fig. 2b) in the south-west coast of the South Island (Boyer and Giribet 2007, 2009). The natural history of these slow-moving leaf-litter dwellers suggests extremely poor dispersal capabilities, with species ranges typically measuring 100 km in diameter or less. Population genetic data generated from New Zealand’s only ‘widespread’ mite harvestman, found throughout the north-west quadrant of the South Island, indicate deep structure associated with geography, corroborating this point of view (Boyer et al. 2007a).
Pettalids have been a centre of discussion on the age of New Zealand’s terrestrial fauna. For example, Boyer and Giribet (2009: 1095) state that: ‘If New Zealand has indeed been submerged, and the terrestrial biota of the archipelago has arrived solely through transoceanic dispersal, we would expect to see close relationships between the pettalid fauna of New Zealand and species from other nearby land masses.’ However, Goldberg et al. (2008: 3323) postulate that: ‘the levels of molecular divergence among the pettalid lineages are too low to be consistent with an ancient origin, unless there has been a very substantial taxon specific change in mutation rate. Diversity and spatial structuring on South Island mountains that are ca. 5 Ma old suggest an arrival within the past few million years.’ This latter statement is interesting, as no dating had been performed in the analyses of Boyer and Giribet (2007, 2009). A more recent article on broader Opiliones phylogeny incorporated samples of the three New Zealand pettalid genera and the two Australian genera, and used a parametric rate-smoothing approach to estimate the origin of Aoraki at 55 Mya (±21), Rakaia at 118 Mya (±22), a divergence between Neopurcellia and Rakaia at 150 Mya (±25), and a divergence of Austropurcellia and Aoraki at 167 Mya (±27) (Giribet et al. in press). All these ages are much older than 22 Mya, contradicting the suggestion by Goldberg et al. (2008) that the New Zealand pettalids arrived within the past 5 million years.
Centipedes: Craterostigmus (Fig. 2e)
Centipedes are ubiquitous terrestrial predators currently classified in five extant orders (Edgecombe and Giribet 2007). One of these orders, Craterostigmomorpha, remained monotypic until the recent formal description of a second species from New Zealand (Edgecombe and Giribet 2008), although the presence of Craterostigmus in New Zealand had been known for quite some time (Archey 1917; R. E. Crabill, Jr, pers. comm. in Lewis 1981) considered at least two species to be distinguishable within Craterostigmus based on morphology, but his arguments in favour of the distinctness of a New Zealand species were never published, and only recently some minor morphological differences between these two species have been proposed (Prunescu and Prunescu 2006; Edgecombe and Giribet 2008). Edgecombe and Giribet (2008) focused on genetic distances within and among the two species to postulate an ancient split of these two lineages, despite the minimal morphological variation between the two, and Giribet et al. (2009) further tested the reciprocal monophyly of the species. However, the lack of Craterostigmus fossils prevented the authors from calibrating this molecular tree (Edgecombe and Giribet 2008). In a more recent analysis of the whole centipede order, Murienne, Edgecombe and Giribet (unpubl. data) used several calibrations in the centipede tree to conclude that the two species of Craterostigmus diverged from each other 270 Mya (s.d. = 26.30, min = 179.75, max = 317.79), at least by the Jurassic. Certainly the ages for Craterostigmus divergences, even if perhaps inflated owing to methodological issues (see Murienne, Edgecombe and Giribet, unpubl. data), contradict the possibility of C. crabilli being a geologically young introduction to New Zealand, as postulated by some.
Stone centipedes: Paralamyctes (Fig. 2f)
Other centipede groups that have been proposed to show Gondwanan patterns involving New Zealand are found in the lithobiomorph family Henicopidae (Edgecombe et al. 2002; Edgecombe and Giribet 2003). Among henicopids, the best-studied genus is Paralamyctes, found in Chile, South Africa, Madagascar, India, Australia (including Tasmania) and New Zealand (Giribet and Edgecombe 2006), a distribution paralleling those of Pettalidae and Peripatopsidae (see below). However, in these other groups, none or just a few genera have a vicariant distribution across multiple land masses, whereas Paralamyctes is widespread and distributed across the former temperate Gondwana, suggesting either an ancient age for the genus or a much higher dispersal capability than the one observed in the groups discussed above. No Paralamyctes fossil is known for calibrating the trees; however, the recent dating of a densely sampled centipede tree (Murienne, Edgecombe and Giribet, unpubl. data) suggests that divergences within the eight sampled species in the genus range between 137 and 253 Mya, hence suggesting that the multiple trans-Tasman clades are a result of old vicariant events, and not due to recent dispersal.
Velvet worms or ‘Peripatus’: the family Peripatopsidae (Fig. 2g, h)
Onychophorans famously require very specific environmental conditions to avoid desiccation. Of the two extant families, Peripatopsidae has a temperate Gondwanan distribution paralleling those of Pettalidae and Paralamyctes with some minor differences. Members of Peripatopsidae are known from Chile, South Africa, Australia (including Tasmania), New Guinea, and New Zealand (Reid 1996; Mayer 2007) (pettalids are also known from Sri Lanka and Madagascar, but they are not known from Tasmania or New Guinea; Paralamyctes is missing from New Guinea but present in the Indian subcontinent). The New Zealand Onychophora has been studied in detail (Gleeson 1996; Trewick 1998, 1999, 2000; Allwood et al. 2010). Two independent genera are recognised in New Zealand, Peripatoides and Ooperipatellus, both of which find their sister clade in Tasmania (Ooperipatellus is a trans-Tasman genus), or Tasmania and/or mainland Australia (Peripatoides) (Gleeson et al. 1998; Allwood et al. 2010). The presence of deep genetic structure within Peripatoides has been interpreted to indicate an ancient origin with local survival of populations through climate cycles (Goldberg et al. 2008). Allwood et al. (2010) estimated divergence times of the New Zealand clades, suggesting that the New Zealand and Australian genera probably diversified during the Cretaceous, but all of the New Zealand Ooperipatellus divergences had a large error and overlapped both with the Oligocene Drowning and the rifting of New Zealand from Australia. However, the diversification of Peripatoides pre-dated the peak of the Oligocene Drowning (Allwood et al. 2010).
Conclusions
Evaluating the hypothesis of New Zealand’s total submersion during the Oligocene requires the strictest tests, nowadays perhaps satisfactorily addressed only by rigorous phylogenetic analysis (the phylogenetic pattern must be congruent with the hypothesis tested; e.g. recent dispersal likely results in paraphyly of the source landmass) and dating of the phylogenies examined. The era of invoking vicariance without phylogenetic analysis has long passed and, likewise, phylogenetic dating must be incorporated to discern between contrasting hypotheses. The four cases discussed above suggest, to us, a pattern that contradicts the recent idea of total submersion of New Zealand. This is evidenced both by the reciprocal monophyly of the New Zealand clades and their sister taxa in the case of Pettalidae, and by the dating of the cladogenetic events that led to the current diversity. Such dating indicates an origin much older than the proposed episode of total submersion. The case of the onychophoran clades is more contentious, as molecular dating results remain ambiguous.
In addition to the further testing of candidate groups such as velvet worms, there may be many other examples to be studied, such as the harvestmen (Arachnida, Opiliones) of the family Monoscutidae (Fig. 2i), restricted to Australia, Tasmania, and New Zealand (including the subantarctic islands) (Cokendolpher and Taylor: 118–121, in Pinto-da-Rocha and Giribet 2007; Taylor and Hunt 2009); the harvestman families Triaenonychidae (Fig. 2j) and Caddidae, with lineages well represented in New Zealand (Forster 1947, 1954, 1962, 1963; Giribet and Kury 2007); and the New Zealand endemic springtail genus Holacanthella (Hexapoda, Collembola, Neanuridae) with close relatives in Australia, New Caledonia, India and nearby non-Gondwanan landmasses (Stevens et al. 2007). Sound phylogenetic relationships need to be obtained before being able to add these taxa to the list of possible relictual candidates of ‘Moa’s Ark’. Certain families of Acari and Pseudoscorpiones have also been proposed for biogeographic studies of Gondwanan patterns (Harvey 1996a, b), but a recent phylogenetic analysis of pseudoscorpions yielded little broad biogeographic structure (Murienne et al. 2008), perhaps owing to the presence of phoresy in many pseudoscorpion species. Minute soil-dwelling spiders (e.g. Micropholcommatidae: Rix et al. 2008; Rix and Harvey 2010) and certain trap-door spiders (Mygalomorphae) (Forster and Wilton 1968; Griswold and Ledford 2001) could be excellent candidates for more detailed biogeographic research.
Although many, or even the overwhelming majority, of the terrestrial animal lineages represented in New Zealand today might have arrived via dispersal following submergence of land during the Oligocene, or at least show patterns of recent diversification (e.g. Brown et al. 1999; Buckley and Simon 2007), the cases reviewed here indicate that, for at least a few groups, vicariance is still a plausible biogeographical scenario. In fact, the combination of a few ancient divergences with more recent diversifications is not very different from the model proposed for the leaf beetle diversity of the Neotropics (McKenna and Farrell 2006), a combination of ‘evolutionary cradles’ and ‘museums of diversity’. Having an unbalanced mix of old species-poor lineages and a larger proportion of recent radiations may actually be expected even in stable parts of our planet. It is not yet time to bid New Zealand’s Gondwanan heritage goodbye.
Acknowledgments
Prashant Sharma, Ron Clouse, Greg Edgecombe, Mark Harvey and two anonymous reviewers provided comments which helped to improve upon earlier versions of this article.
Allwood J.,
Gleeson D.,
Mayer G.,
Daniels S.,
Beggs J. R., Buckley T. R.
(2010) Support for vicariant origins of the New Zealand Onychophora. Journal of Biogeography 37, 669–681.
| Crossref | GoogleScholarGoogle Scholar |

Archey G.
(1917) The occurrence in New Zealand of Craterostigmus tasmanianus Pocock (Chilopoda). Transactions and Proceedings of the New Zealand Institute 49, 319–320.
Boyer S. L., Giribet G.
(2007) A new model Gondwanan taxon: systematics and biogeography of the harvestman family Pettalidae (Arachnida, Opiliones, Cyphophthalmi), with a taxonomic revision of genera from Australia and New Zealand. Cladistics 23, 337–361.
| Crossref | GoogleScholarGoogle Scholar |
Boyer S. L., Giribet G.
(2009) Welcome back New Zealand: regional biogeography and Gondwanan origin of three endemic genera of mite harvestmen (Arachnida, Opiliones, Cyphophthalmi). Journal of Biogeography 36, 1084–1099.
| Crossref | GoogleScholarGoogle Scholar |
Boyer S. L.,
Baker J. M., Giribet G.
(2007a) Deep genetic divergences in Aoraki denticulata (Arachnida, Opiliones, Cyphophthalmi): a widespread ‘mite harvestman’ defies DNA taxonomy. Molecular Ecology 16, 4999–5016.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Boyer S. L.,
Clouse R. M.,
Benavides L. R.,
Sharma P.,
Schwendinger P. J.,
Karunarathna I., Giribet G.
(2007b) Biogeography of the world: a case study from cyphophthalmid Opiliones, a globally distributed group of arachnids. Journal of Biogeography 34, 2070–2085.
| Crossref | GoogleScholarGoogle Scholar |
Brown B.,
Emberson R. M., Paterson A. M.
(1999) Phylogeny of “Oxycanus” lineages of hepialid moths from New Zealand inferred from sequence variation in the mtDNA COI and II gene regions. Molecular Phylogenetics and Evolution 13, 463–473.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Buckley T. R., Simon C.
(2007) Evolutionary radiation of the cicada genus Maoricicada Dugdale (Hemiptera : Cicadoidea) and the origins of the New Zealand alpine biota. Biological Journal of the Linnean Society. Linnean Society of London 91, 419–435.
| Crossref | GoogleScholarGoogle Scholar |
Buckley T. R.,
Marske K., Attanayake D.
(2010) Phylogeography and ecological niche modelling of the New Zealand stick insect Clitarchus hookeri (White) support survival in multiple coastal refugia. Journal of Biogeography 37, 682–695.
| Crossref | GoogleScholarGoogle Scholar |
Cook L. G., Crisp M. D.
(2005a) Directional asymmetry of long-distance dispersal and colonization could mislead reconstructions of biogeography. Journal of Biogeography 32, 741–754.
| Crossref | GoogleScholarGoogle Scholar |
Cook L. G., Crisp M. D.
(2005b) Not so ancient: the extant crown group of Nothofagus represents a post-Gondwanan radiation. Proceedings of the Royal Society of London. Series B. Biological Sciences 272, 2535–2544.
| Crossref | GoogleScholarGoogle Scholar |
Cooper A., Cooper R. A.
(1995) The Oligocene bottleneck and New Zealand biota: genetic record of a past environmental crisis. Proceedings of the Royal Society of London. Series B. Biological Sciences 261, 293–302.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
Crisp M. D., Cook L. G.
(2009) Explosive radiation or cryptic mass extinction? Interpreting signatures in molecular phylogenies. Evolution 63, 2257–2265.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Edgecombe G. D., Giribet G.
(2003) Relationships of Henicopidae (Chilopoda : Lithobiomorpha): New molecular data, classification and biogeography. African Invertebrates 44, 13–38.
Edgecombe G. D., Giribet G.
(2007) Evolutionary biology of centipedes (Myriapoda : Chilopoda). Annual Review of Entomology 52, 151–170.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Edgecombe G. D., Giribet G.
(2008) A New Zealand species of the trans-Tasman centipede order Craterostigmomorpha (Arthropoda : Chilopoda) corroborated by molecular evidence. Invertebrate Systematics 22, 1–15.
| Crossref | GoogleScholarGoogle Scholar |
Edgecombe G. D.,
Giribet G., Wheeler W. C.
(2002) Phylogeny of Henicopidae (Chilopoda : Lithobiomorpha): A combined analysis of morphology and five molecular loci. Systematic Entomology 27, 31–64.
| Crossref | GoogleScholarGoogle Scholar |
Forster R. R.
(1947) The zoogeographical relationships of the New Zealand Opiliones. The New Zealand Science Congress 1947, 233–235.
Forster R. R.
(1954) The New Zealand harvestmen (Sub-order Laniatores). Canterbury Museum Bulletin 2, 1–329.
Forster R. R.
(1962) A key to the New Zealand harvestmen. Tuatara: Journal of the Biological Society 10, 129–137.
Forster R. R.
(1963) A key to the New Zealand harvestmen – Part 2. Tuatara: Journal of the Biological Society 11, 28–40.
Forster R. R., Wilton C. L.
(1968) The spiders of New Zealand. Part II. Otago Museum Bulletin 2, 1–180.
Giribet G., Edgecombe G. D.
(2006) The importance of looking at small-scale patterns when inferring Gondwanan biogeography: a case study of the centipede Paralamyctes (Chilopoda, Lithobiomorpha, Henicopidae). Biological Journal of the Linnean Society. Linnean Society of London 89, 65–78.
| Crossref | GoogleScholarGoogle Scholar |
Giribet G.,
Guzmán Cuéllar A., Edgecombe G. D.
(2009) Further use of molecular data in studying biogeographic patterns within the centipede genus Craterostigmus: the case for a monophyletic New Zealand species. Soil Organisms 81, 557–563.
Giribet G.,
Vogt L.,
Pérez González A.,
Sharma P., Kury A. B.
(in press) A multilocus approach to harvestman (Arachnida : Opiliones) phylogeny with emphasis on biogeography and the systematics of Laniatores. Cladistics 26,
| Crossref | GoogleScholarGoogle Scholar |
Gleeson D. M.
(1996) Onychophora of New Zealand: Past, present and future. New Zealand Entomologist 19, 51–55.
Gleeson D. M.,
Rowell D. M.,
Tait N. N.,
Briscoe D. A., Higgins A. V.
(1998) Phylogenetic relationships among onychophora from Australasia inferred from the mitochondrial cytochrome oxidase subunit I gene. Molecular Phylogenetics and Evolution 10, 237–248.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Goldberg J.,
Trewick S. A., Paterson A. M.
(2008) Evolution of New Zealand’s terrestrial fauna: a review of molecular evidence. Philosophical Transactions of the Royal Society B. Biological Sciences 363, 3319–3334.
| Crossref | GoogleScholarGoogle Scholar |
Grandcolas P.,
Murienne J.,
Robillard T.,
DeSutter-Grandcolas L.,
Jourdan H., Guilbert E.
(2008) New Caledonia: a very old Darwinian island? Philosophical Transactions of the Royal Society B. Biological Sciences 363, 3309–3317.
| Crossref | GoogleScholarGoogle Scholar |
Griswold C. E., Ledford J.
(2001) A monograph of the migid trap door spiders of Madagascar and review of the world genera (Araneae, Mygalomorphae, Migidae). Occasional Papers of the California Academy of Sciences 151, 1–120.
Harvey M. S.
(1996b) The biogeography of Gondwanan pseudoscorpions (Arachnida). Revue suisse de Zoologie hors série 1 , 255–264.
|
CAS |
Harvey M. S.
(2002) Short-range endemism among the Australian fauna: some examples from non-marine environments. Invertebrate Systematics 16, 555–570.
| Crossref | GoogleScholarGoogle Scholar |
Heads M., Craw R.
(2004) The Alpine Fault biogeographic hypothesis revisited. Cladistics 20, 184–190.
| Crossref | GoogleScholarGoogle Scholar |
Knapp M.,
Stöckler K.,
Havell D.,
Delsuc F.,
Sebastiani F., Lockhart P. J.
(2005) Relaxed molecular clock provides evidence for long-distance dispersal of Nothofagus (southern beech). PLoS Biology 3, e14.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Knapp M.,
Mudaliar R.,
Havell D.,
Wagstaff S. J., Lockhart P. J.
(2007) The drowning of New Zealand and the problem of Agathis. Systematic Biology 56, 862–870.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Landis C. A.,
Campbell H. J.,
Begg J. G.,
Mildenhall D. C.,
Paterson A. M., Trewick S. A.
(2008) The Waipounamu Erosion Surface: questioning the antiquity of the New Zealand land surface and terrestrial fauna and flora. Geological Magazine 145, 173–197.
| Crossref | GoogleScholarGoogle Scholar |
Marske K. A.,
Leschen R. A. B.,
Barker G. M., Buckley T. R.
(2009) Phylogeography and ecological niche modelling implicate coastal refugia and trans-alpine dispersal of a New Zealand fungus beetle. Molecular Ecology 18, 5126–5142.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Mayer G.
(2007) Metaperipatus inae sp. nov. (Onychophora : Peripatopsidae) from Chile with a novel ovarian type and dermal insemination. Zootaxa 1440, 21–37.
McGlone M. S.
(2005) Goodbye Gondwana. Journal of Biogeography 32, 739–740.
| Crossref | GoogleScholarGoogle Scholar |
McKenna D. D., Farrell B. D.
(2006) Tropical forests are both evolutionary cradles and museums of leaf beetle diversity. Proceedings of the National Academy of Sciences of the United States of America 103, 10 947–10 951.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Murienne J.
(2009) Testing biodiversity hypotheses in New Caledonia using phylogenetics. Journal of Biogeography 36, 1433–1434.
| Crossref | GoogleScholarGoogle Scholar |
Murienne J.,
Grandcolas P.,
Piulachs M. D.,
Bellés X.,
D’Haese C.,
Legendre F.,
Pellens R., Guilbert E.
(2005) Evolution of a shaky piece of Gondwana: is local endemism recent in New Caledonia? Cladistics 21, 2–7.
Murienne J.,
Harvey M. S., Giribet G.
(2008) First molecular phylogeny of the major clades of Pseudoscorpiones (Arthropoda : Chelicerata). Molecular Phylogenetics and Evolution 49, 170–184.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Phillips M. J.,
Gibb G. C.,
Crimp E. A., Penny D.
(2010) Tinamous and moa flock together: Mitochondrial genome sequence analysis reveals independent losses of flight among ratites. Systematic Biology 59, 90–107.
| Crossref | GoogleScholarGoogle Scholar |
Pole M. S.
(2001) Can long-distance dispersal be inferred from the New Zealand plant fossil record? Australian Journal of Botany 49, 357–366.
| Crossref |
Prunescu C.-C., Prunescu P.
(2006) Rudimentary supernumerary Malpighian tubules in the order Craterostigmomorpha Pocock 1902. Norwegian Journal of Entomology 53, 113–118.
Reid A. L.
(1996) A review of the Peripatopsidae (Onychophora) in Australia, with descriptions of new genera and species, and comments on Peripatopsid relationships. Invertebrate Taxonomy 10, 663–936.
| Crossref | GoogleScholarGoogle Scholar |
Rix M. G., Harvey M. S.
(2010) The spider family Micropholcommatidae (Arachnida, Araneae, Araneoidea): a relimitation and revision at the generic level. Zookeys 36, 1–321.
| Crossref | GoogleScholarGoogle Scholar |
Rix M. G.,
Harvey M. S., Roberts J. D.
(2008) Molecular phylogenetics of the spider family Micropholcommatidae (Arachnida: Araneae) using nuclear rRNA genes (18S and 28S). Molecular Phylogenetics and Evolution 46, 1031–1048.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Sanmartín I., Ronquist F.
(2004) Southern hemisphere biogeography inferred by event-based models: Plant versus animal patterns. Systematic Biology 53, 216–243.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Scotese C. R.,
Gahagan L. M., Larson R. L.
(1988) Plate tectonic reconstructions of the Cretaceous and Cenozoic ocean basins. Tectonophysics 155, 27–48.
| Crossref |
Stevens M. I.,
Winter D. J.,
Morris R.,
McCartney J., Greenslade P.
(2007) New Zealand’s giant Collembola: new information on distribution and morphology for Holacanthella Börner, 1906 (Neanuridae: Uchidanurinae). New Zealand Journal of Zoology 34, 63–78.
Swenson U.,
Backlund A.,
McLoughlin S., Hill R. S.
(2001) Nothofagus biogeography revisited with special emphasis on the enigmatic distribution of subgenus Brassospora in New Caledonia. Cladistics 17, 28–47.
| Crossref | GoogleScholarGoogle Scholar |
Taylor C. K., Hunt G. S.
(2009) New genus of Megalopsalidinae (Arachnida: Opiliones: Monoscutidae) from north-eastern Australia. Zootaxa 2130, 41–59.
Trewick S. A.
(1998) Sympatric cryptic species in New Zealand Onychophora. Biological Journal of the Linnean Society. Linnean Society of London 63, 307–329.
| Crossref | GoogleScholarGoogle Scholar |
Trewick S. A.
(1999) Molecular diversity of Dunedin peripatus (Onychophora: Peripatopsidae). New Zealand Journal of Zoology 26, 381–393.
Trewick S. A.
(2000) Mitochondrial DNA sequences support allozyme evidence for cryptic radiation of New Zealand Peripatoides (Onychophora). Molecular Ecology 9, 269–281.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Trewick S. A.,
Paterson A. M., Campbell H. J.
(2007) Hello New Zealand. Journal of Biogeography 34, 1–6.
| Crossref | GoogleScholarGoogle Scholar |
Wallis G. P., Trewick S. A.
(2009) New Zealand phylogeography: evolution on a small continent. Molecular Ecology 18, 3548–3580.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Waters J. M., Craw D.
(2006) Goodbye Gondwana? New Zealand biogeography, geology, and the problem of circularity. Systematic Biology 55, 351–356.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Weir J. T.,
Bermingham E., Schluter D.
(2009) The Great American Biotic Interchange in birds. Proceedings of the National Academy of Sciences of the United States of America 106, 21 737–21 742.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |