

19 OPTIMIZATION OF ENUCLEATION TIME AFTER IVM FOR SOMATIC CELL NUCLEAR TRANSFER IN GOAT
G. S. Ajithkumar A , B. Krishnamohan A and B. C. Sarkhel AJawaharlal Nehru Krishi Vishwa Vidyalaya, Jabalpur, Madhya Pradesh, India
Reproduction, Fertility and Development 21(1) 109-110 https://doi.org/10.1071/RDv21n1Ab19
Published: 9 December 2008
Abstract
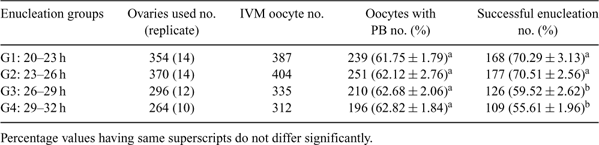
For cloning by somatic cell nuclear transfer (SCNT) in goat, there are conflicting reports about the proper time of enucleation after IVM of oocytes, which varied from 20 to 27 h (Das SK et al. 2003 Small Rumin. Res. 48, 217–225; Keefer CL et al. 2002 Biol. Reprod. 66, 199–203; Daniel SM et al. 2007 Small Rumin. Res. 77, 45–50). The present investigation has been undertaken to standardize the optimum time of enucleation after IVM of oocytes. The hypothesis behind the study was that enucleation performed during early stages of maturation maintains the MII plate and polar body (PB) in a closer position and therefore makes it easy to enucleate. To test this hypothesis, caprine COCs were aspirated from slaughterhouse ovaries of goats and good quality oocytes were matured in TCM 199 containing 7.5% FBS supplemented with FSH, LH, and estradiol. Enucleation was performed in four different interval groups after IVM (20–23 h, 23–26 h, 26–29 h and 29–32 h). The enucleation of oocytes was conducted as per method described by (Du F et al. 2006 Theriogenology 65, 642–665). Briefly, IVM oocytes were enucleated by squashing and compressing out the first PB along with 10 to 15% of its surrounding cytoplasm with an enucleation needle through a slit made on the zona pellucida. Successful enucleation was confirmed by fluorescent microscopy of removed ooplasm after staining with Hoechst 33342. The enucleation percentage values after arcsine transformation was analyzed by completely randomized design ANOVA. The result of enucleation at different intervals has been summarized in Table 1. There was no significant difference (P > 0.05) in number of PB observed among the four enucleation groups, however the enucleation percentage decreased significantly (P < 0.05) with increase in enucleation time (70.29% and 70.51% in G1 and G2 v. 59.52% and 55.61% in G3 and G4 respectively). With increase in time of enucleation after maturation the size of perivitelline space increases, causing deviation of PB from spindle, thus the success rate of enucleation is reduced (Song K et al. 2007 Repro. Fertil. Dev. 19, 293–294). In G1 and G2 groups the PB and MII chromosomes are located close together with stronger spindle force that requires minimum ooplasm to be removed with higher percentage of successful enucleation. Hence, it was concluded that G1 and G2 groups may be considered as most efficient for enucleation but the developmental potential of reconstructed oocytes after nuclear transfer in each group needs to be tested (study under progress).
|