

Hyperglycaemia and lipid differentially impair mouse oocyte developmental competence
Siew L. Wong A , Linda L. Wu A , Rebecca L. Robker A , Jeremy G. Thompson A B and Melanie L. Sutton McDowall A B CA Robinson Research Institute, School of Paediatrics and Reproductive Health, The University of Adelaide, Medical School, Frome Road, Adelaide, SA 5005, Australia.
B Australian Research Council (ARC) Centre of Excellence for Nanoscale BioPhotonics, Institute of Photonics and Advanced Sensing.
C Corresponding author. Email: melanie.mcdowall@adelaide.edu.au
Reproduction, Fertility and Development 27(4) 583-592 https://doi.org/10.1071/RD14328
Submitted: 4 September 2014 Accepted: 6 January 2015 Published: 26 February 2015
Abstract
Maternal diabetes and obesity are characterised by elevated blood glucose, insulin and lipids, resulting in upregulation of specific fuel-sensing and stress signalling pathways. Previously, we demonstrated that, separately, upregulation of the hexosamine biosynthetic pathway (HBP; under hyperglycaemic conditions) and endoplasmic reticulum (ER) stress (due to hyperlipidaemia) pathways reduce blastocyst development and alter oocyte metabolism. In order to begin to understand how both glucose and lipid metabolic disruptions influence oocyte developmental competence, in the present study we exposed mouse cumulus–oocyte complexes to hyperglycaemia (30 mM) and/or lipid (40 μM) and examined the effects on embryo development. The presence of glucosamine (GlcN; a hyperglycaemic mimetic) or increased lipid during in vitro maturation severely perturbed blastocyst development (P < 0.05). Hyperglycaemia, GlcN and hyperglycaemia + lipid treatments significantly increased HBP activity, increasing total O-linked glycosylation (O-GlcNAcylation) of proteins (P < 0.0001). All treatments also induced ER stress pathways, indicated by the expression of specific ER stress genes. The expression of genes encoding the HBP enzymes glutamine:fructose-6-phosphate amidotransferase 2 (Gfpt2) and O-linked β-N-acetylglucosaminyltransferase (Ogt) was repressed following lipid treatment (P < 0.001). These findings partially implicate the mechanism of O-GlcNAcylation and ER stress as likely contributors to compromised fertility of obese women.
Additional keywords: cumulus–oocyte complex, embryo, endoplasmic reticulum stress, hexosamine biosynthesis pathway, hyperlipidemia.
Introduction
Maternal obesity, a condition associated with elevated plasma insulin, glucose and lipid, is a global health problem that affects an increasing number of women of reproductive age (Australian Institute of Health and Welfare (AIHW) 2012). Obesity is also a contributing factor to Type 2 diabetes, characterised by hyperglycaemia (elevated blood glucose level) and hyperlipidemia. These conditions have been associated with a higher risk of subfertility and pregnancy complications, including an increased risk of anovulation, pre-eclampsia, miscarriage and spontaneous abortion (Jungheim and Moley 2010). They are also associated with an increased incidence of congenital anomalies, macrosomia and stillborns (Sirimi and Goulis 2010). Moreover, these complications extend beyond neonatal health, into childhood and adulthood (Heerwagen et al. 2010; Dabelea and Crume 2011; O’Reilly and Reynolds 2013).
The periconception period (including the final stages of oocyte development and fertilisation) is particularly sensitive to the maternal metabolic environment. For example, zygotes collected from oviducts of diabetic mice and subsequently transferred to normoglycaemic recipients have retarded fetal growth and increased fetal abnormalities (Wyman et al. 2008). Hyperglycaemia during oocyte maturation and early development is associated with altered intracellular parameters, such as perturbed meiotic maturation and disrupted mitochondrial distribution (Colton et al. 2002; Chang et al. 2005), as well as apoptosis in follicular and granulosa cells (Chang et al. 2005).
The detrimental effects of hyperglycaemia on oocyte developmental competence are mediated, in part, by upregulation of the hexosamine biosynthesis pathway (HBP; Sutton-McDowall et al. 2006; Schelbach et al. 2010, 2012), a fuel-sensing pathway that metabolises glucose to uridine diphosphate-N-acetylglucosamine (UDP-GlcNAc). Single UDP-GlcNAc molecules modify serine or threonine amino acids of proteins, a process known as β-O-linked glycosylation (O-GlcNAcylation), which acts in an analogous manner to phosphorylation to regulate protein function (Butkinaree et al. 2010). Altered O-GlcNAcylation is one of the primary pathologies of diabetes in somatic cells and notably the primary mechanism behind the development of insulin resistance in Type 2 diabetes (Marshall et al. 1991; Yang et al. 2008).
Glucosamine (GlcN), a known hyperglycaemic mimetic, can be metabolised via the HBP by bypassing the rate-limiting enzyme of the HBP, namely glutamine:fructose-6-phosphate amidotransferase (GFPT; Nelson et al. 2000; Uldry et al. 2002); hence, it is a potent stimulator of HBP pathway activity. Previously, we demonstrated that GlcN supplementation during IVM results in increased O-GlcNAcylation in mouse COCs (Frank et al. 2014a) and perturbed oocyte developmental competence in cow, pig and mouse and decreased cleavage rates in the mouse (Sutton-McDowall et al. 2006; Kimura et al. 2008; Schelbach et al. 2012; Frank et al. 2013).
Hyperlipidaemia is also known to cause numerous perturbations in oocyte structure and developmental competence. Female mice fed a high-fat diet (HFD) have higher rates of anovulation, smaller and fewer mature oocytes, increased lipid accumulation, altered mitochondrial activity and decreased rates of oocyte nuclear maturation and fertilisation compared with oocytes derived from control mice (Igosheva et al. 2010; Jungheim et al. 2010; Wu et al. 2010; Luzzo et al. 2012). Increased lipid accumulation induces lipotoxicity, which causes damage to cellular organelles, particularly mitochondria and endoplasmic reticulum (ER). A biomarker of lipotoxicity is ER stress, characterised by the accumulation of misfolded proteins and consequently triggering the unfolded protein response (UPR; Alhusaini et al. 2010; Wu et al. 2012a).
UPR is an attempt by the cell to slow protein production and improve protein folding, characterised by the induction of several genes, including the markers activating transcription factor 4 (Atf4), activating transcription factor 6 (Atf6) and glucose-regulated protein 78 (Grp78; Malhotra and Kaufman 2007). In the absence of the UPR, cellular stress will likely culminate in apoptotic cell death. Cumulus–oocyte complexes (COCs) from mice fed an HFD were shown to have increased expression of ER stress marker genes Atf4 and Grp78, similar to those matured in lipid-rich follicular fluid (Wu et al. 2010; Yang et al. 2012). Similar events happen in women, with increased Atf4 expression observed in granulosa cells of obese women (Wu et al. 2010).
A link between the HBP and ER stress has been suggested in somatic cells (Srinivasan et al. 2009; Sage et al. 2010; Lombardi et al. 2012), with increasing activity through the HBP increasing the transcript and translocation of ER stress markers. However, little is known about how hyperglycaemia and lipid conditions impact reproductive function, in particular preimplantation embryo development. Furthermore, it is unclear whether these metabolic perturbations operate through the same or different mechanisms. The aim of the present study was to examine the impact of hyperglycaemia and lipid supplementation on mouse oocyte developmental competence. We hypothesised that combination of lipid and high glucose leads to the accumulation of the HBP product UDP-GlcNAc by increasing O-GlcNAcylation and activating the ER stress pathway.
Materials and methods
Mice
Female CBA/F1 mice were obtained at 21 days of age from the University of Adelaide (Waite campus) and kept in the Animal House at the Medical School, the University of Adelaide (North Terrace campus) under a 14 : 10-h light–dark cycle with ad libitum access to food and water. All animal experiments were approved by the University of Adelaide's Animal Ethics Committee (Medical) and were conducted in accordance with the Australian Code of Practice for the Care and Use of Animals for Scientific Purposes.
Isolation and in vitro maturation of mouse COCs
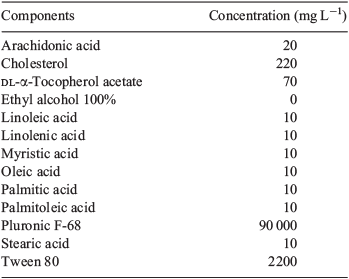
Immature, unexpanded COCs were isolated by puncturing the antral follicles of ovaries collected 46 h after i.p. injection of 5 IU pregnant mare's serum gonadotropin (PMSG; Folligon; Intervet, Boxmeer, The Netherlands). All COCs were collected in HEPES-buffered α-minimum essential medium (MEM) handling media (Life Technologies, Invitrogen, Carlsbad, CA, USA) supplemented with 4 mg mL–1 fatty acid-free bovine serum albumin (MP Biomedicals, Solon, OH, USA). Immature COCs were cultured in groups of 30 in 1.5 mL pre-equilibrated in vitro maturation (IVM) medium, overlaid with paraffin oil (Merck, Darmstadt, Germany). The base IVM medium (control) was bicarbonate-buffered α-MEM supplemented with 1% fetal bovine serum (FBS; Invitrogen), 50 mIU mL–1 recombinant human FSH (Puregon-Organon, Oss, The Netherlands) and 10 ng mL–1 recombinant human epidermal growth factor (EGF; R&D Systems, Minneapolis, MN, USA). Experimental treatments were as follows: (1) control (5.56 mM glucose); (2) high glucose (30 mM glucose); (3) glucosamine (2.5 mM GlcN plus 5.56 mM glucose); (4) lipid (40 μM of a commercially available lipid concentrate; GIBCO, Invitrogen); and (5) lipid and high glucose (40 μM lipid concentrate plus 30 mM glucose). COCs were cultured at 37°C in an atmosphere of 6% CO2, 5% O2 and 89% N2 for 8 or 16 h. We have previously demonstrated that large culture volumes are required to maintain hyperglycaemic concentrations of glucose due to the high metabolic rate of COCs (Frank et al. 2013). In the present study, 30 mM glucose was used to avoid depletion to more normoglycaemic levels during the course of maturation (Sutton-McDowall et al. 2010). The commercially available lipid concentrate was a solution composed of 1 mM of 10 lipids, dissolved in ethyl alcohol and Tween 80 (see Table 1 for the composition of the lipid concentrate).
|
Development of the lipid accumulation model
A dose–response experiment was performed to determine the concentration of the lipid concentrate (0, 5, 10 20, 40 μM) that resulted in increased lipid accumulation within the oocyte. After 16 h culture, COCs were fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS) for 1 h, washed in PBS and transferred to 1 μg mL–1 of the neutral lipid stain BODIPY 493/503 (Life Technologies, Invitrogen) in PBS for 1 h in the dark at room temperature. COCs were washed in PBS for 5 min and mounted on coverslips in 3 μL DAKO fluorescent mounting medium (Dako, Glostrup, Denmark). Images were captured using a Fluoview FV10i confocal microscope (Olympus, Tokyo, Japan) using a green laser (excitation 480 nm, emission 515 nm); identical magnification, image and laser settings were used throughout experiments (see Fig. S1 available as Supplementary Material to this paper). Two replicates were performed.
Assessment of cumulus expansion index
Cumulus expansion was assessed after 16 h culture by an independent assessor, blinded to treatments, using a scale described previously (Vanderhyden et al. 1990), as follows: 0, no expansion of cumulus cells; +1, the outer most layers of cumulus cells expanded; +2, expansion of the entire outer half of cumulus cells; +3, all layers expanded except the corona radiatae; and +4, maximal expansion of all layers of cumulus cells. For each treatment group, a mean cumulus expansion index (CEI; over the range 0.0–4.0) was calculated. Three replicates were performed, averaging 25 COCs per treatment group and replicate.
IVF and assessment of embryo development
Following 16 h maturation, COCs were washed once in fertilisation medium (VitroFert; Cook Australia, Brisbane, Qld, Australia) and COCs were transferred to pre-equilibrated fertilisation drops overlaid with paraffin oil. Male mice, which had previously been assessed for mating ability (not less than 3 days prior), were used as sperm donors for IVF. Mice were killed by cervical dislocation and the epididymides and vasa deferentia were collected into warm (37°C) wash medium (VitroWash; Cook Australia), cleaned of excess fat and tissue and transferred into 1 mL fertilisation medium. Spermatozoa were extracted into the medium and allowed to capacitate at 37°C in an atmosphere of 6% CO2, 5% O2 and 89% N2 for 1 h. Capacitated spermatozoa (10 μL) were added to 90-μL fertilisation drops and COCs and spermatozoa were coincubated for 4 h at 37°C in an atmosphere of 6% CO2, 5% O2 and 89% N2. COCs were transferred to wash medium and cumulus cells were removed mechanically by repeat pipetting. Presumptive zygotes were washed in embryo culture medium (VitroCleave; Cook Australia) and placed into culture drops (four to seven per 10-μL drop) at 37°C in an atmosphere of 6% CO2, 5% O2 and 89% N2. Twenty-four hours after IVF (Day 2), the fertilisation rate was assessed and 2-cell embryos were transferred onto a fresh 20-μL drop of embryo culture medium. Embryo morphology was assessed on Day 5 (the end of the culture period, 96–100 h after fertilisation). Embryo development was assessed on Day 2 (expected 2-cell stage) and Day 5 (blastocysts or hatching blastocysts). Seven replicates were performed, with 50 COCs per treatment group and replicate.
Immunocytochemistry
Following IVM (16 h), COCs were fixed in 4% paraformaldehyde in PBS overnight. Whole COCs were adhered on Cell-Tak (BD Biosciences, San Jose, CA, USA)-coated slides and stained immunohistochemically using the primary antibody CTD110.6 (anti-O-GlcNAc antibody; Covance, Princeton, NJ, USA) for O-GlcNAc and propidium iodide (PI) for nuclear staining. Briefly, COCs were permeabilised for 30 min in 0.25% Triton X-100 (United States Biochemical, Salem, MA, USA), blocked for 2 h using 10% goat serum in PBS (Jackson Immuno Research Labs, West Grove, PA, USA) and incubated overnight at 4°C with 1 : 250 CTD110.6 in blocking solution. On Day 2, COCs were washed and incubated for 2 h at room temperature with 1 : 250 Alexa Fluor 488 goat anti-mouse IgM (Invitrogen) in blocking solution, washed twice in PBS–polyvinylpolypyrrolidone (PVP) and 30 min with PI before being mounted under a coverslip in 3 μL DAKO fluorescent mounting medium (Dako). A negative control with no primary antibody was also included and no autofluorescence was observed with the microscope settings used. Fluorescence intensity and localisation were examined using a Fluoview FV10i confocal microscope (Olympus) using the Alexa Fluor 488 filter for CTD110.6 (excitation 488 nm, emission 519 nm) and the PI filter (excitation 488 nm, emission 617 nm). Single optical sections were acquired from a median plane of zona in the oocytes. Three replicates were performed, with 10 COCs per treatment group. Localisation and intensity of CTD110.6-positive staining across the COC were determined using ImageJ software (NIH; http://imagej.nih.gov/ij/) by placing a box across the oocyte and cumulus cell image and measuring pixel intensity. The mean fluorescence intensity in each pixel column was reported and the mean ± s.e.m. calculated. Data are represented graphically as the intensity of fluorescence over pixel widths (Wu et al. 2010). Total O-linked-glycosylated protein was determined as the sum of total fluorescence in the boxed area.
RNA isolation and real-time reverse transcription–polymerase chain reaction
Following 8 or 16 h IVM, groups of 90–100 COCs were collected in a minimal volume of medium, snap frozen in liquid nitrogen and stored at –80°C. Total RNA was isolated from COC using the TRIzol reagent (Invitrogen) method, followed by DNase treatment using an Ambion kit (Invitrogen) as per the manufacturer's instructions. RNA concentration and purity were quantified using a Nanodrop ND-1000 Spectrophotometer (Biolab, Carmel, IN, USA) before reverse transcribing 600 ng RNA using random primers (Invitrogen) and Superscript III Reverse Transcriptase (Invitrogen) according to the manufacturer's instructions. Ribosomal protein RPl19 (QIAGEN, Venlo, Limburg, The Netherlands) was used as a validated internal control for every sample. The X-box binding protein 1 (Xbp1s) primers were 5′-AGG CTT GGT GTA TAC ATG G-3′ (reverse) and 5′-GGT CTG CTG AGT CCG CAG GAG G-3′ (forward; Ozcan et al. 2009); the other primers were Quantitect Primer assays (QIAGEN). Real-time reverse transcription–polymerase chain reaction (RT-PCR) was performed in quadruplicate using SYBR green PCR Master Mix (Applied Biosystems, Foster City, CA, USA) and a Rotor-Gene 6000 (Corbett, Valencia, CA, USA) real-time rotary analyser. Real-time RT-PCR data were analysed using the 2–(ΔCT) method and are expressed as the fold change relative to a calibrator sample, which was included in each run. Eight replicates were performed for the control treatment and four replicates were performed for all other treatments, with 90–100 COCs per treatment group and replicate.
Statistical analysis
Statistical analysis was performed using Graph Pad Prism version 5.0 (GraphPad Software, La Jolla, CA, USA). Differences between treatment groups were tested using a general linear model, followed by Bonferroni post hoc tests. Cleavage and blastocyst rates were arcsine transformed before analyses. Outcome parameters are reported as the mean ± s.e.m. Two-sided P < 0.05 was considered significant.
Results
Experiment 1: lipid and GlcN impair oocyte developmental competence
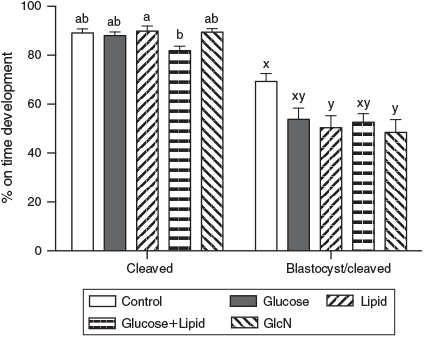
CEIs were examined before IVF. Hyperglycaemia and lipid treatments during IVM had no effect on cumulus expansion (P > 0.05; Table 2). On Day 2, the cleavage rate of COCs treated with high glucose + lipid was significantly lower than lipid treatment alone (81.86 ± 1.88% vs 89.86 ± 2.06%, respectively; P < 0.05). There were no significant differences in cleavage rates between the other treatment groups. The fertilised oocytes from COCs treated with GlcN or lipid yielded significantly fewer blastocysts on Day 5 than the control (48.49 ± 5.19% and 50.35 ± 4.93% vs 69.28 ± 3.29%, respectively; P < 0.05). The blastocyst rate in the high glucose + lipid treatment group tended to be lower than in the control group (Fig. 1; P = 0.08).

|
|
Experiment 2: O-GlcNAcylation localisation
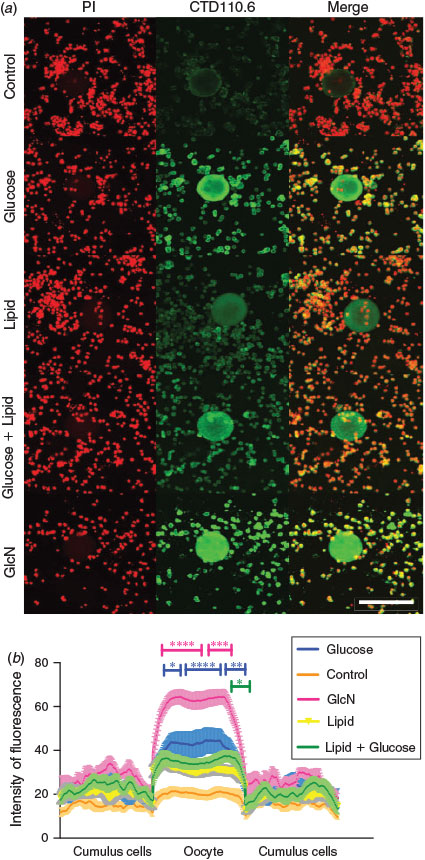
Immunohistochemical localisation of O-GlcNAc with CTD110.6 revealed positive staining in the oocytes, with higher intensity in COCs treated with high glucose, lipid, GlcN and high glucose + lipid compared with control (Fig. 2a). Densitometry analysis of CTD110.6 revealed similar results, where GlcN treatment significantly increased O-GlcNAcylation-positive staining within the oocyte compared with the control group (Fig. 2b; P < 0.05). Increased O-GlcNAcylation was also observed in oocytes treated with high glucose. In contrast, the addition of lipid to high glucose diminished the extent of staining relative to glucose alone. No significant differences were observed in cumulus cells among treatment groups (Fig. 2b).
|
Experiment 3: expression of HBP enzymes and ER stress genes
The mRNA expression for HBP enzymes, namely Gfpt1, Gfpt2, O-linked β-N-acetylglucosaminyltransferase (Ogt) and hyaluronan synthase 2 (Has2), was determined following exposure to high glucose and lipid. After 8 h maturation, GlcN treatment resulted in 2.2-, 2.8- and 4.0-fold increases in Gfpt1, Gfpt2 and Has2 expression, respectively, compared with control (Fig. 3a, b, d). There were no other significant differences in gene expression between the other treatment groups (Fig. 3). After 16 h, there was a corresponding increased in Gfpt1 (1.8-fold) and Gfpt2 (1.4-fold) expression (Fig. 3a, b) following the GlcN treatment. Interestingly, the addition of lipid to the culture medium significantly decreased the expression of Ogt (an enzyme that adds the O-GlcNAcylation to proteins) and Gfpt2 compared with control (Fig. 3b, c).

|
After 8 h maturation, supplementation of GlcN in the culture medium resulted in increased expression of the ER stress gene Grp78 (threefold) and Atf4 (Fig. 4a–c). After 16 h, a similar trend was observed in gene expression levels, with increased Xbp1 (1.8-fold) and Grp78 (2.6-fold) expression following GlcN treatment (Figs 4a, c). Lipid- and high glucose + lipid-treated COC also exhibited significantly increased expression of Xbp1 (1.8-fold). In addition, after 16 h, Atf4 expression was increased in COCs treated with GlcN (2.3-fold) and high glucose (1.5-fold). Interestingly, Atf4 expression was significantly reduced when lipid was added to the high-glucose culture medium. There were no significant differences in Atf6 gene expression between treatment groups (Fig. 4d).

|
Discussion
There is a significant body of evidence that demonstrates poor preimplantation embryo development outcomes following hyperglycaemic or lipidaemic exposure during in vivo or in vitro development. The present study elucidates some of the cellular mechanisms by which high glucose and lipid conditions contribute to reduced oocyte quality and developmental competence. In the present study we used a chemically defined lipid concentrate that contains various non-esterified fatty acids (NEFAs) as a substitute for using single lipids, such as palmitic acid or lipid-rich follicular fluid, which have been used in other studies (Aardema et al. 2011; Wu et al. 2012b; Yang et al. 2012). This was because the lipid concentrate more closely resembles the NEFA detected in women with increased body mass index (Robker et al. 2009; Valckx et al. 2012) and follicular fluid contains hormones, growth factors and undefined proteins that may compromise development. Furthermore, we have investigated the role of high glucose and lipid on O-GlcNAcylation and ER stress during and at the end of IVM. Notably, hyperglycaemic conditions increase aberrant O-GlcNAcylation in oocytes and induce the expression of ER stress genes. Surprisingly, the presence of lipid in the culture medium represses the expression of genes encoding HBP enzymes, such as the rate-limiting enzymes Gfpt2 and Ogt.
The cleavage rate of COCs treated with the combination of high glucose and lipid was lower than treatment with lipid alone, whereas blastocyst development was significantly reduced following GlcN and lipid supplementation, but only tended to decrease following high glucose treatment. This discrepancy with high glucose treatment may be explained by the difference in culture media used: a much simpler medium was used in our previous study (Frank et al. 2013). The result with lipid treatment is consistent with several other studies demonstrating that lipid level is critical during embryo development (Leroy et al. 2005; Van Hoeck et al. 2011; Wu et al. 2012b; Yang et al. 2012). Interestingly, there was a tendency for decreased blastocyst development following culture with high glucose + lipid, suggesting that high levels of glucose do not have an additive effect in impairing oocyte developmental competence.
Increased O-GlcNAcylation of proteins is a hallmark of increased HBP activity, induced by either hyper- or hypoglycaemia. Following IVM, high glucose or GlcN significantly increase O-GlcNAcylation levels, in line with GlcN being a potent stimulator of HBP UDP-GlcNAc production (~40-fold, as measured in adipocytes; Marshall et al. 1991). Excess flux with either glucose or GlcN through HBP has been shown previously to reduce embryo development and this phenomenon was reversed using an O-linked N-acetylglucosamine trasnferase (OGT) inhibitor, reflecting their relative potential to stimulate HBP and UDP-GlcNAc production (Pantaleon et al. 2010; Frank et al. 2014a, 2014b). The addition of lipid to the medium did not increase O-GlcNAcylation levels. This phenomenon could be explained by the fact that opposing interactions between unsaturated fatty acids (such as oleic acid) are able to inhibit the pro-apoptotic effect of their counterpart-saturated palmitic and stearic acid (Nĕmcová-Fürstová et al. 2011).
There were no additive effects of high glucose and lipid on β-O-GlcNAcylation. Indeed, intensity within the oocyte the in lipid and high glucose group was reduced overall compared with high glucose-treated COCs. Under the in vitro conditions used herein, lipid appears to play a role in downregulating cellular responses to hyperglycaemia. Hyperglycaemia is a characteristic of both types of diabetes, yet hyperlipidaemia is only associated with Type 2 diabetes. Although the present study used an in vitro model, we have also found that systemic administration of GlcN during the pericompaction period has significant effects on fetal survival and abnormalities, which were most evident in lean rather than obese mice, indicating the possibility that high lipid and glucose levels may interact and regulate fuel-sensing pathways, specifically the HBP (Schelbach et al. 2013).
Compared with the control group, GlcN treatment significantly increased mRNA expression of the rate-limiting enzymes of the HBP, namely Gfpt1 and Gfpt2, after 8 and 16 h culture. Although high glucose treatment did not affect the gene expression of HBP enzymes (possibly due to glucose regulation of these enzymes being at the substrate availability and post-translational level), expression of Gfpt2 and Ogt (the enzymes that modify proteins with GlcNAc) was downregulated in the lipid-treated compared with control group, supporting the notion that lipids interact with glucose-sensing pathways. Elevated fatty acids have been shown to interfere with glucose by inhibiting glucose-induced insulin secretion and β-cell oxidation in rat pancreatic islets (Sako and Grill 1990; Zhou and Grill 1994). Moreover, free fatty acid supplementation decreases the expression of solute carrier family 2 (facilitated glucose transporter), member 2 (SLC2A2) and glucokinase (Gremlich et al. 1997). Therefore, downregulation of HBP enzymes in the presence of lipids may be due to impaired glucose transporter activities, reducing glucose transportation down the pathway and decreasing the production of UDP-GlcNAc, the end-product of HBP. Alternatively, the level of UDP-GlcNAc may also be limited by the known feedback inhibition of Gfpt by UDP-GlcNAc (Kornfeld 1967). All these interactions of lipid with glucose further support the idea that nutrient-sensing pathways are interconnected in the ovary, COC and other ovarian cells. Although O-GlcNAcylation staining revealed maximal staining in the oocyte, the differences in gene expression are likely to be within the oocyte and surrounding cumulus cells. Oocytes exhibit low transcriptional activity; however, bidirectional communication between the oocyte and cumulus cells is essential for oocyte viability (Eppig 1991; Albertini et al. 2001). So, changes in cumulus cells should be reflected in the oocytes.
The induction of ER stress is a well-characterised response of lipotoxicity (Borradaile et al. 2006; Malhi and Gores 2008). Consistent with previous studies, Xbp1 and Atf4 were specifically upregulated in COC following lipid treatment (Wu et al. 2012b; Yang et al. 2012). GlcN treatment significantly increased Xbp1, Atf4 and Grp78 expression, suggesting that the COC is undergoing a distinct UPR response or that the presence of GlcN influences the stress response. Interestingly, high glucose significantly increased the expression of Atf4 after 16 h culture, which was downregulated in the high glucose and lipid treatment group, further emphasising that lipid appears to inhibit the effects of hyperglycaemia. In contrast, Atf6 expression remained unaffected by all treatments at both time points. Activation of Atf6 requires a dissociation step from its inhibitory regulator, binding immunoglobulin protein (BiP). Binding of Atf6 to BiP was reported to be very stable (Shen et al. 2005) and could explain why there was no activation of this pathway in response to the treatments.
In summary, the findings of the present study demonstrate that lipid at levels of 40 μM (and GlcN) induce ER stress and that high glucose (and GlcN) increase O-GlcNAcylation but that the two treatments do not potentiate each other in these pathways. Furthermore, the present study suggests the possibility of lipid downregulating the detrimental consequences of hyperglycaemia on oocyte health. This study also indicates O-GlcNAcylation and ER stress as likely contributors to the reduced fertility observed in obese women. This provides new leads for further investigations into possible treatment strategies and interventions that may improve pregnancy and fetal outcomes in obese women and those with associated co-morbidities (e.g. diabetes).
Acknowledgements
The authors thank Dr Laura Frank for expert technical assistance, Lesley Ritter for assistance with cumulus expansion scoring and Dr Xiaoqian Wang for embryo development scoring. MSM was funded by the Australian Research Council Centre of Excellence for Nanoscale BioPhotonics. The Fluoview FV10i confocal microscope was purchased as part of the Sensing Technologies for Advanced Reproductive Research (STARR) facility, funded by the South Australia's Premier's Science and Research Fund.
References
Aardema, H., Vos, P. L., Lolicato, F., Roelen, B. A., Knijn, H. M., Vaandrager, A. B., Helms, J. B., and Gadella, B. M. (2011). Oleic acid prevents detrimental effects of saturated fatty acids on bovine oocyte developmental competence. Biol. Reprod. 85, 62–69.| Oleic acid prevents detrimental effects of saturated fatty acids on bovine oocyte developmental competence.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXotFOls74%3D&md5=91c35c1502e6480acc58b76f6a2faff4CAS | 21311036PubMed |
Australian Institute of Health and Welfare (AIHW) (2012). ‘Australia's Health 2012. Australia's Health Series No. 13.’ Catalogue no. AUS 156. (AIHW: Canberra.)
Albertini, D. F., Combelles, C. M., Benecchi, E., and Carabatsos, M. J. (2001). Cellular basis for paracrine regulation of ovarian follicle development. Reproduction 121, 647–653.
| Cellular basis for paracrine regulation of ovarian follicle development.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXktVaisbY%3D&md5=deb9f901bb90e46c53ca80b16045c08aCAS | 11427152PubMed |
Alhusaini, S., McGee, K., Schisano, B., Harte, A., McTernan, P., Kumar, S., and Tripathi, G. (2010). Lipopolysaccharide, high glucose and saturated fatty acids induce endoplasmic reticulum stress in cultured primary human adipocytes: salicylate alleviates this stress. Biochem. Biophys. Res. Commun. 397, 472–478.
| Lipopolysaccharide, high glucose and saturated fatty acids induce endoplasmic reticulum stress in cultured primary human adipocytes: salicylate alleviates this stress.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXosV2lsrY%3D&md5=59c60396f7675d91fb74a3c5fa2fa239CAS | 20515657PubMed |
Borradaile, N. M., Han, X., Harp, J. D., Gale, S. E., Ory, D. S., and Schaffer, J. E. (2006). Disruption of endoplasmic reticulum structure and integrity in lipotoxic cell death. J. Lipid Res. 47, 2726–2737.
| Disruption of endoplasmic reticulum structure and integrity in lipotoxic cell death.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhtlSit7fE&md5=49e2c2cbd4e6ef9c872610be85c089d5CAS | 16960261PubMed |
Butkinaree, C., Park, K., and Hart, G. W. (2010). O-Linked beta-N-acetylglucosamine (O-GlcNAc): extensive crosstalk with phosphorylation to regulate signaling and transcription in response to nutrients and stress. Biochim. Biophys. Acta 1800, 96–106.
| O-Linked beta-N-acetylglucosamine (O-GlcNAc): extensive crosstalk with phosphorylation to regulate signaling and transcription in response to nutrients and stress.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXotVWitQ%3D%3D&md5=06a6f73a8a58823a340330048d5e36c8CAS | 19647786PubMed |
Chang, A. S., Dale, A. N., and Moley, K. H. (2005). Maternal diabetes adversely affects preovulatory oocyte maturation, development, and granulosa cell apoptosis. Endocrinology 146, 2445–2453.
| Maternal diabetes adversely affects preovulatory oocyte maturation, development, and granulosa cell apoptosis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXjslamtbk%3D&md5=97323c6060e6b276acf7d7e0f224b9c3CAS | 15718275PubMed |
Colton, S. A., Pieper, G. M., and Downs, S. M. (2002). Altered meiotic regulation in oocytes from diabetic mice. Biol. Reprod. 67, 220–231.
| Altered meiotic regulation in oocytes from diabetic mice.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XkvV2itb8%3D&md5=14d0e8885a8fabd40da870f4bf97f7f2CAS | 12080021PubMed |
Dabelea, D., and Crume, T. (2011). Maternal environment and the transgenerational cycle of obesity and diabetes. Diabetes 60, 1849–1855.
| Maternal environment and the transgenerational cycle of obesity and diabetes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXptVChtLc%3D&md5=a47355214a76548a4ca2e0ae477ace08CAS | 21709280PubMed |
Eppig, J. J. (1991). Intercommunication between mammalian oocytes and companion somatic cells. BioEssays 13, 569–574.
| Intercommunication between mammalian oocytes and companion somatic cells.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK387is1ShsA%3D%3D&md5=94f7607b92d428edb9bccf8dade3a11dCAS | 1772412PubMed |
Frank, L. A., Sutton-McDowall, M. L., Russell, D. L., Wang, X., Feil, D. K., Gilchrist, R. B., and Thompson, J. G. (2013). Effect of varying glucose and glucosamine concentration in vitro on mouse oocyte maturation and developmental competence. Reprod. Fertil. Dev. 25, 1095–1104.
| Effect of varying glucose and glucosamine concentration in vitro on mouse oocyte maturation and developmental competence.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXhsF2ltLzK&md5=fe1501d6fc79d34eb0e9cbf8e33ee16dCAS | 23131421PubMed |
Frank, L. A., Sutton-McDowall, M. L., Brown, H. M., Russell, D. L., Gilchrist, R. B., and Thompson, J. G. (2014a). Hyperglycaemic conditions perturb mouse oocyte in vitro developmental competence via beta-O-linked glycosylation of heat shock protein 90. Hum. Reprod. 29, 1292–1303.
| Hyperglycaemic conditions perturb mouse oocyte in vitro developmental competence via beta-O-linked glycosylation of heat shock protein 90.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2cXotF2ku70%3D&md5=e6e1830938ebaae750f6d079a05ed90fCAS | 24713123PubMed |
Frank, L. A., Sutton-McDowall, M. L., Gilchrist, R. B., and Thompson, J. G. (2014b). The effect of peri-conception hyperglycaemia and the involvement of the hexosamine biosynthesis pathway in mediating oocyte and embryo developmental competence. Mol. Reprod. Dev. 81, 391–408.
| The effect of peri-conception hyperglycaemia and the involvement of the hexosamine biosynthesis pathway in mediating oocyte and embryo developmental competence.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2cXht1ymsrc%3D&md5=c3e74d22e529db22a135164a4bba4881CAS | 24415135PubMed |
Gremlich, S., Bonny, C., Waeber, G., and Thorens, B. (1997). Fatty acids decrease IDX-1 expression in rat pancreatic islets and reduce GLUT2, glucokinase, insulin, and somatostatin levels. J. Biol. Chem. 272, 30 261–30 269.
| Fatty acids decrease IDX-1 expression in rat pancreatic islets and reduce GLUT2, glucokinase, insulin, and somatostatin levels.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXnslGhurk%3D&md5=849e3696ae1d6470bd63c2d74a5976d1CAS |
Heerwagen, M. J., Miller, M. R., Barbour, L. A., and Friedman, J. E. (2010). Maternal obesity and fetal metabolic programming: a fertile epigenetic soil. Am. J. Physiol. Regul. Integr. Comp. Physiol. 299, R711–R722.
| Maternal obesity and fetal metabolic programming: a fertile epigenetic soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXht1WgurvI&md5=fca89c9445a1dab76a3a98b2ce1f4c7cCAS | 20631295PubMed |
Igosheva, N., Abramov, A. Y., Poston, L., Eckert, J. J., Fleming, T. P., Duchen, M. R., and McConnell, J. (2010). Maternal diet-induced obesity alters mitochondrial activity and redox status in mouse oocytes and zygotes. PLoS ONE 5, e10074.
| Maternal diet-induced obesity alters mitochondrial activity and redox status in mouse oocytes and zygotes.Crossref | GoogleScholarGoogle Scholar | 20404917PubMed |
Jungheim, E. S., and Moley, K. H. (2010). Current knowledge of obesity's effects in the pre- and periconceptional periods and avenues for future research. Am. J. Obstet. Gynecol. 203, 525–530.
| Current knowledge of obesity's effects in the pre- and periconceptional periods and avenues for future research.Crossref | GoogleScholarGoogle Scholar | 20739012PubMed |
Jungheim, E. S., Schoeller, E. L., Marquard, K. L., Louden, E. D., Schaffer, J. E., and Moley, K. H. (2010). Diet-induced obesity model: abnormal oocytes and persistent growth abnormalities in the offspring. Endocrinology 151, 4039–4046.
| Diet-induced obesity model: abnormal oocytes and persistent growth abnormalities in the offspring.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXht1Oju7rN&md5=3ae385c2afcbf3ae82f5263f60e39232CAS | 20573727PubMed |
Kimura, K., Iwata, H., and Thompson, J. G. (2008). The effect of glucosamine concentration on the development and sex ratio of bovine embryos. Anim. Reprod. Sci. 103, 228–238.
| The effect of glucosamine concentration on the development and sex ratio of bovine embryos.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhsVSmtLfL&md5=116dbfa7b9a43f0b2576e5f5791e2de3CAS | 17198747PubMed |
Kornfeld, R. (1967). Studies on l-glutamine d-fructose 6-phosphate amidotransferase. I. Feedback inhibition by uridine diphosphate-N-acetylglucosamine. J. Biol. Chem. 242, 3135–3141.
| 1:CAS:528:DyaF2sXktFyjtbw%3D&md5=8d9581eafb23607b64f49ab1bf70a8ffCAS | 4961641PubMed |
Leroy, J. L., Vanholder, T., Mateusen, B., Christophe, A., Opsomer, G., de Kruif, A., Genicot, G., and Van Soom, A. (2005). Non-esterified fatty acids in follicular fluid of dairy cows and their effect on developmental capacity of bovine oocytes in vitro. Reproduction 130, 485–495.
| Non-esterified fatty acids in follicular fluid of dairy cows and their effect on developmental capacity of bovine oocytes in vitro.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXhtFent7rJ&md5=16ddfb8af1de3a55f051d7e88b361557CAS | 16183866PubMed |
Lombardi, A., Ulianich, L., Treglia, A. S., Nigro, C., Parrillo, L., Lofrumento, D. D., Nicolardi, G., Garbi, C., Beguinot, F., Miele, C., and Di Jeso, B. (2012). Increased hexosamine biosynthetic pathway flux dedifferentiates INS-1E cells and murine islets by an extracellular signal-regulated kinase (ERK)1/2-mediated signal transmission pathway. Diabetologia 55, 141–153.
| Increased hexosamine biosynthetic pathway flux dedifferentiates INS-1E cells and murine islets by an extracellular signal-regulated kinase (ERK)1/2-mediated signal transmission pathway.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhsFOqu73J&md5=22cab6313e5eea24bb3f85bad0dbdff8CAS | 22006246PubMed |
Luzzo, K. M., Wang, Q., Purcell, S. H., Chi, M., Jimenez, P. T., Grindler, N., Schedl, T., and Moley, K. H. (2012). High fat diet induced developmental defects in the mouse: oocyte meiotic aneuploidy and fetal growth retardation/brain defects. PLoS ONE 7, e49217.
| High fat diet induced developmental defects in the mouse: oocyte meiotic aneuploidy and fetal growth retardation/brain defects.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhslKjtrvN&md5=9bc77d7bb0f30a2e41cdb8f2da55d9a2CAS | 23152876PubMed |
Malhi, H., and Gores, G. J. (2008). Molecular mechanisms of lipotoxicity in nonalcoholic fatty liver disease. Semin. Liver Dis. 28, 360–369.
| Molecular mechanisms of lipotoxicity in nonalcoholic fatty liver disease.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhsVKju7fF&md5=02e20718fb244faeb888a9ea2823b35dCAS | 18956292PubMed |
Malhotra, J. D., and Kaufman, R. J. (2007). Endoplasmic reticulum stress and oxidative stress: a vicious cycle or a double-edged sword? Antioxid. Redox Signal. 9, 2277–2294.
| Endoplasmic reticulum stress and oxidative stress: a vicious cycle or a double-edged sword?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXht1GrsLzL&md5=78f168cd01d100829dd049d688d3c7f2CAS | 17979528PubMed |
Marshall, S., Bacote, V., and Traxinger, R. R. (1991). Discovery of a metabolic pathway mediating glucose-induced desensitization of the glucose transport system. Role of hexosamine biosynthesis in the induction of insulin resistance. J. Biol. Chem. 266, 4706–4712.
| 1:CAS:528:DyaK3MXhs1WltL0%3D&md5=2b41f197d868161b85ac85f21827718dCAS | 2002019PubMed |
Nelson, B. A., Robinson, K. A., and Buse, M. G. (2000). High glucose and glucosamine induce insulin resistance via different mechanisms in 3T3-L1 adipocytes. Diabetes 49, 981–991.
| High glucose and glucosamine induce insulin resistance via different mechanisms in 3T3-L1 adipocytes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXjvFylu74%3D&md5=fae39c6bdb2ebfc4d23eb7fe95d3ca80CAS | 10866051PubMed |
Nĕmcová-Fürstová, V., James, R. F., and Kovář, J. (2011). Inhibitory effect of unsaturated fatty acids on saturated fatty acid-induced apoptosis in human pancreatic beta-cells: activation of caspases and ER stress induction. Cell. Physiol. Biochem. 27, 525–538.
| Inhibitory effect of unsaturated fatty acids on saturated fatty acid-induced apoptosis in human pancreatic beta-cells: activation of caspases and ER stress induction.Crossref | GoogleScholarGoogle Scholar | 21691070PubMed |
O’Reilly, J. R., and Reynolds, R. M. (2013). The risk of maternal obesity to the long-term health of the offspring. Clin. Endocrinol. (Oxf) 78, 9–16.
| The risk of maternal obesity to the long-term health of the offspring.Crossref | GoogleScholarGoogle Scholar | 23009645PubMed |
Ozcan, L., Ergin, A. S., Lu, A., Chung, J., Sarkar, S., Nie, D., Myers, M. G., and Ozcan, U. (2009). Endoplasmic reticulum stress plays a central role in development of leptin resistance. Cell Metab. 9, 35–51.
| Endoplasmic reticulum stress plays a central role in development of leptin resistance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXptFWquw%3D%3D&md5=4337533202542028650ead559962b940CAS | 19117545PubMed |
Pantaleon, M., Tan, H. Y., Kafer, G. R., and Kaye, P. L. (2010). Toxic effects of hyperglycemia are mediated by the hexosamine signaling pathway and O-linked glycosylation in early mouse embryos. Biol. Reprod. 82, 751–758.
| Toxic effects of hyperglycemia are mediated by the hexosamine signaling pathway and O-linked glycosylation in early mouse embryos.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXjslelt7g%3D&md5=ce8d63049509a4bb361a109f3d00c0e6CAS | 20032283PubMed |
Robker, R. L., Akison, L. K., Bennett, B. D., Thrupp, P. N., Chura, L. R., Russell, D. L., Lane, M., and Norman, R. J. (2009). Obese women exhibit differences in ovarian metabolites, hormones, and gene expression compared with moderate-weight women. J. Clin. Endocrinol. Metab. 94, 1533–1540.
| Obese women exhibit differences in ovarian metabolites, hormones, and gene expression compared with moderate-weight women.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXlvVCqtrw%3D&md5=ad9fc9f21f34fed2964925dd65571412CAS | 19223519PubMed |
Sage, A. T., Walter, L. A., Shi, Y., Khan, M. I., Kaneto, H., Capretta, A., and Werstuck, G. H. (2010). Hexosamine biosynthesis pathway flux promotes endoplasmic reticulum stress, lipid accumulation, and inflammatory gene expression in hepatic cells. Am. J. Physiol. Endocrinol. Metab. 298, E499–E511.
| Hexosamine biosynthesis pathway flux promotes endoplasmic reticulum stress, lipid accumulation, and inflammatory gene expression in hepatic cells.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXjsFGgsL0%3D&md5=0ce3310bfcde4aaef8afd28ef9e6f20aCAS | 19952345PubMed |
Sako, Y., and Grill, V. E. (1990). A 48-hour lipid infusion in the rat time-dependently inhibits glucose-induced insulin secretion and B cell oxidation through a process likely coupled to fatty acid oxidation. Endocrinology 127, 1580–1589.
| A 48-hour lipid infusion in the rat time-dependently inhibits glucose-induced insulin secretion and B cell oxidation through a process likely coupled to fatty acid oxidation.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3cXlvVGisrs%3D&md5=01fb4e23b61feedc856b72691820c928CAS | 1698143PubMed |
Schelbach, C. J., Kind, K. L., Lane, M., and Thompson, J. G. (2010). Mechanisms contributing to the reduced developmental competence of glucosamine-exposed mouse oocytes. Reprod. Fertil. Dev. 22, 771–779.
| Mechanisms contributing to the reduced developmental competence of glucosamine-exposed mouse oocytes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXlsVCqt7k%3D&md5=a158e2ff505ba5c1ffa41b002fe845ebCAS | 20450829PubMed |
Schelbach, C. J., Robker, R. L., Bennett, B. D., Gauld, A. D., Thompson, J. G., and Kind, K. L. (2013). Altered pregnancy outcomes in mice following treatment with the hyperglycaemia mimetic, glucosamine, during the periconception period. Reprod. Fertil. Dev. 25, 405–416.
| Altered pregnancy outcomes in mice following treatment with the hyperglycaemia mimetic, glucosamine, during the periconception period.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXhslart7k%3D&md5=c349f9a24deb00fb777d5260bb475cd7CAS | 23445817PubMed |
Shen, J., Snapp, E. L., Lippincott-Schwartz, J., and Prywes, R. (2005). Stable binding of ATF6 to BiP in the endoplasmic reticulum stress response. Mol. Cell. Biol. 25, 921–932.
| Stable binding of ATF6 to BiP in the endoplasmic reticulum stress response.Crossref | GoogleScholarGoogle Scholar | 15657421PubMed |
Sirimi, N., and Goulis, D. G. (2010). Obesity in pregnancy. Hormones 9, 299–306.
| Obesity in pregnancy.Crossref | GoogleScholarGoogle Scholar | 21112860PubMed |
Srinivasan, V., Tatu, U., Mohan, V., and Balasubramanyam, M. (2009). Molecular convergence of hexosamine biosynthetic pathway and ER stress leading to insulin resistance in L6 skeletal muscle cells. Mol. Cell. Biochem. 328, 217–224.
| Molecular convergence of hexosamine biosynthetic pathway and ER stress leading to insulin resistance in L6 skeletal muscle cells.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXnvFKgtro%3D&md5=870201fed9f6a8d64e60e31c05f9aef2CAS | 19370316PubMed |
Sutton-McDowall, M. L., Mitchell, M., Cetica, P., Dalvit, G., Pantaleon, M., Lane, M., Gilchrist, R. B., and Thompson, J. G. (2006). Glucosamine supplementation during in vitro maturation inhibits subsequent embryo development: possible role of the hexosamine pathway as a regulator of developmental competence. Biol. Reprod. 74, 881–888.
| Glucosamine supplementation during in vitro maturation inhibits subsequent embryo development: possible role of the hexosamine pathway as a regulator of developmental competence.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28Xjsl2jur8%3D&md5=380109c0be67107cef1de50ca8963396CAS | 16436527PubMed |
Sutton-McDowall, M. L., Gilchrist, R. B., and Thompson, J. G. (2010). The pivotal role of glucose metabolism in determining oocyte developmental competence. Reproduction 139, 685–695.
| The pivotal role of glucose metabolism in determining oocyte developmental competence.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXltFajtr0%3D&md5=72e1531d06ee8b60f71467945f41f3c0CAS | 20089664PubMed |
Uldry, M., Ibberson, M., Hosokawa, M., and Thorens, B. (2002). GLUT2 is a high affinity glucosamine transporter. FEBS Lett. 524, 199–203.
| GLUT2 is a high affinity glucosamine transporter.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38Xls1WksrY%3D&md5=6ac69b3e5f7ea1f3eae72b29deab02ceCAS | 12135767PubMed |
Valckx, S. D., De Pauw, I., De Neubourg, D., Inion, I., Berth, M., Fransen, E., Bols, P. E., and Leroy, J. L. (2012). BMI-related metabolic composition of the follicular fluid of women undergoing assisted reproductive treatment and the consequences for oocyte and embryo quality. Hum. Reprod. 27, 3531–3539.
| BMI-related metabolic composition of the follicular fluid of women undergoing assisted reproductive treatment and the consequences for oocyte and embryo quality.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhslKrsLrL&md5=36e8e2c48e7445163a84e1ab94e8a470CAS | 23019302PubMed |
Van Hoeck, V., Sturmey, R. G., Bermejo-Alvarez, P., Rizos, D., Gutierrez-Adan, A., Leese, H. J., Bols, P. E., and Leroy, J. L. (2011). Elevated non-esterified fatty acid concentrations during bovine oocyte maturation compromise early embryo physiology. PLoS ONE 6, e23183.
| Elevated non-esterified fatty acid concentrations during bovine oocyte maturation compromise early embryo physiology.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhtFKmsLjM&md5=b6eb69e3072d782b967fdaef51eb2625CAS | 21858021PubMed |
Vanderhyden, B. C., Caron, P. J., Buccione, R., and Eppig, J. J. (1990). Developmental pattern of the secretion of cumulus expansion-enabling factor by mouse oocytes and the role of oocytes in promoting granulosa cell differentiation. Dev. Biol. 140, 307–317.
| Developmental pattern of the secretion of cumulus expansion-enabling factor by mouse oocytes and the role of oocytes in promoting granulosa cell differentiation.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK3czitlersw%3D%3D&md5=d254635a3cf3e261bb5b149a24fd28baCAS | 2115479PubMed |
Wu, L. L., Dunning, K. R., Yang, X., Russell, D. L., Lane, M., Norman, R. J., and Robker, R. L. (2010). High-fat diet causes lipotoxicity responses in cumulus–oocyte complexes and decreased fertilization rates. Endocrinology 151, 5438–5445.
| High-fat diet causes lipotoxicity responses in cumulus–oocyte complexes and decreased fertilization rates.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhsVKks7bL&md5=eefe18c6b932287de00f82f7d18fd452CAS | 20861227PubMed |
Wu, L. L., Norman, R. J., and Robker, R. L. (2012a). The impact of obesity on oocytes: evidence for lipotoxicity mechanisms. Reprod. Fertil. Dev. 24, 29–34.
| The impact of obesity on oocytes: evidence for lipotoxicity mechanisms.Crossref | GoogleScholarGoogle Scholar |
Wu, L. L., Russell, D. L., Norman, R.J., and Robker, R. L. (2012b). Endoplasmic reticulum (ER) stress in cumulus–oocyte complexes impairs pentraxin-3 secretion, mitochondrial membrane potential (ΔΨm), and embryo development. Mol. Endocrinol. 26, 562–573.
| Endoplasmic reticulum (ER) stress in cumulus–oocyte complexes impairs pentraxin-3 secretion, mitochondrial membrane potential (ΔΨm), and embryo development.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xlt1ensrs%3D&md5=6b717d7aea88f13eb2febbd3718bd7c1CAS | 22383462PubMed |
Wyman, A., Pinto, A. B., Sheridan, R., and Moley, K. H. (2008). One-cell zygote transfer from diabetic to nondiabetic mouse results in congenital malformations and growth retardation in offspring. Endocrinology 149, 466–469.
| One-cell zygote transfer from diabetic to nondiabetic mouse results in congenital malformations and growth retardation in offspring.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXht1yqsLc%3D&md5=37be794dcb93716892545e6ec3867ec8CAS | 18039778PubMed |
Yang, X., Ongusaha, P. P., Miles, P. D., Havstad, J. C., Zhang, F., So, W. V., Kudlow, J. E., Michell, R. H., Olefsky, J. M., Field, S. J., and Evans, R. M. (2008). Phosphoinositide signalling links O-GlcNAc transferase to insulin resistance. Nature 451, 964–969.
| Phosphoinositide signalling links O-GlcNAc transferase to insulin resistance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXit1yns74%3D&md5=566b4edad3367006d5069c51d6a298f8CAS | 18288188PubMed |
Yang, X., Wu, L. L., Chura, L. R., Liang, X., Lane, M., Norman, R. J., and Robker, R. L. (2012). Exposure to lipid-rich follicular fluid is associated with endoplasmic reticulum stress and impaired oocyte maturation in cumulus–oocyte complexes. Fertil. Steril. 97, 1438–1443.
| Exposure to lipid-rich follicular fluid is associated with endoplasmic reticulum stress and impaired oocyte maturation in cumulus–oocyte complexes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XktlChsbs%3D&md5=86e359977edd7ce09a76e8239914f8f8CAS | 22440252PubMed |
Zhou, Y. P., and Grill, V. E. (1994). Long-term exposure of rat pancreatic islets to fatty acids inhibits glucose-induced insulin secretion and biosynthesis through a glucose fatty acid cycle. J. Clin. Invest. 93, 870–876.
| Long-term exposure of rat pancreatic islets to fatty acids inhibits glucose-induced insulin secretion and biosynthesis through a glucose fatty acid cycle.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2cXktVWntb4%3D&md5=2b7a69f0cb5de7d55238665e67a14235CAS | 8113418PubMed |