

Potential bleaching effects on coral reproduction
Mary Hagedorn A B C , Virginia L. Carter A B , Claire Lager B , Julio F. Camperio Ciani A B , Alison N. Dygert A B , Reuben D. Schleiger A B and E. Michael Henley A BA Department of Reproductive Sciences, Smithsonian Conservation Biology Institute – National Zoological Park, Front Royal, VA 22630, USA.
B Hawai’i Institute of Marine Biology, University of Hawaii, Kaneohe, HI 96744, USA.
C Corresponding author. Email: hagedornm@si.edu
Reproduction, Fertility and Development 28(8) 1061-1071 https://doi.org/10.1071/RD15526
Submitted: 12 December 2015 Accepted: 5 April 2016 Published: 17 June 2016
Journal Compilation © CSIRO Publishing 2016 Open Access CC BY-NC-ND
Abstract
Bleaching profoundly impacts coral reproduction, often for years after an event. However, detailed reproductive characteristics of coral after bleaching have not been broadly described, especially as they relate to cryopreservation. Therefore, in the present study we measured several reproductive characteristics in coral in Kaneohe Bay, Hawaii, for two species, namely Fungia scutaria and Montipora capitata, during the bleaching period of 2014 and 2015. We examined spawning periods, egg morphometry, sperm concentration, fresh and cryopreserved sperm motility exposed to different concentrations of dimethyl sulfoxide, time of first cleavage, larval survival with fresh and cryopreserved spermatozoa, infection success and settlement success. Many of these reproductive parameters were reduced in 2015, especially sperm motility. Once the reduced-motility spermatozoa from 2015 post-bleach were cryopreserved, there was a steep decline in post-thaw viability and this would prevent any substantive further use of these samples in reproduction for conservation benefit. Worldwide, as bleaching events become more frequent, the ability to bank and conserve coral ex situ may be significantly reduced. Thus, it is imperative that while genetic diversity is still high in these populations, intensive efforts are made to bank coral species during non-bleaching periods.
Additional keywords: cryopreservation, Fungia scutaria, Montipora capitata, sperm, Symbiodinium.
Introduction
Symbiotic partnerships are essential for the health and function of biological systems, ranging from individuals to ecosystems (Relman 2008). Most frequently, the basis for the symbiosis is the exchange of nutrients between partners. Symbiotic relationships allow animals and plants to grow in nutrient-poor environments by using the rich metabolic proficiencies of micro-organisms. In return, the microbe in these mutualistic partnerships usually gains nutrients and/or a protected environment. Many tropical corals host intracellular dinoflagellate symbionts, called Symbiodinium, that are critical for their survival (Kawaguti 1944; McLaughlin and Zahl 1959). These symbionts are characterised by their clades, and each clade has certain geographical distribution and temperature tolerance, beyond which they begin to stress (Baker 2003; LaJeunesse et al. 2004).
When the coral holobiont begins to stress, it can loose its life-giving symbionts and bleach. Bleaching is a physiological response to many environmental variables, including, among others, temperature, water chemistry, solar irradiance and bacterial infections. Stress responses of coral have been described for many years (Vaughan 1914), but mass bleaching phenomena were first noted around the world in the mid-1970s and 1980s, leading to subsequent coral mortality and concern for the long-term survival of reefs (for a review, see Baker et al. 2008). These mass bleaching events correlate with El Niño Southern Oscillation events, which produce elevated sustained (10–12 weeks) sea surface temperatures approximately 1°C above the summer maximum, and in time can result in coral mortality (Glynn and D’Crox 1990; Glynn 1996). As these bleaching events became more common, it was realised that they were tightly linked to the production of greenhouse gases by our overuse of fossil fuels (Jokiel and Coles 1990; Glynn 1991, 1993; Goreau 1992; Fitt et al. 2001; Baker et al. 2008). The report from the Intergovernmental Panel on Climate Change (IPCC 2007) has predicted that, during the period 2012–40, coral reefs will experience more frequent bleaching events closer together in time and has identified these bleaching events as the greatest threat to the world’s reef systems.
Bleaching profoundly affects reproduction in coral often for more than one season (Ward et al. 2002). In that study, Ward et al. (2002) determined that although the global bleaching event in March 1998 affected Heron Island coral extensively, some individuals bleached whereas others did not. By November 1998 the ‘recovered’ coral had fewer polyps containing eggs and testes and the existing eggs were smaller than the unbleached colonies (Ward et al. 2002). Moreover, the subsequent bleaching event the following year wiped out all the reproduction in the ‘recovered’ coral (Cox and Ward 2002). An 11-year study on marked coral individuals in Panama was conducted to determine how likely coral are to recover from bleaching (Levitan et al. 2014). The results were extremely disturbing because they clearly demonstrated the stressful long-term effects of warming leading to the reduced reproduction on coral that visibly bleached, as well as on those that did not bleach (Levitan et al. 2014). So, even though corals were not conspicuously loosing their Symbiodinium, the long-term stress was found to disrupt reproduction.
It is clear that bleaching has profound effects on the reproduction of coral; however, detailed reproductive characteristics of coral after bleaching are limited (Szmant and Gassman 1990; Randall and Szmant 2009). Therefore, we were interested in detailing the reproductive profile of Fungia scutaria and Montipora capitata coral populations in Kaneohe Bay (most of which bleached in October 2014), 9 months after their recovery. We compared the details of their post-bleaching reproductive profile to the more than 30 years of reproductive data on F. scutaria in Kaneohe Bay (Krupp 1983; Schwarz et al. 1999; Hagedorn et al. 2006, 2010, 2012, 2015a) and the almost 20 years of data for M. capitata reproduction (Field 1998; Cox 2007; Padilla-Gamiño et al. 2011, 2014; Padilla-Gamiño and Gates 2012; Hagedorn et al. 2015b). Nevertheless, without consistent yearly reproductive profiles assessed in a similar manner, these historical comparisons can only be suggestive at best. However, this reproductive profile, presented here, may lay the fundamental groundwork to examine future reproductive changes.
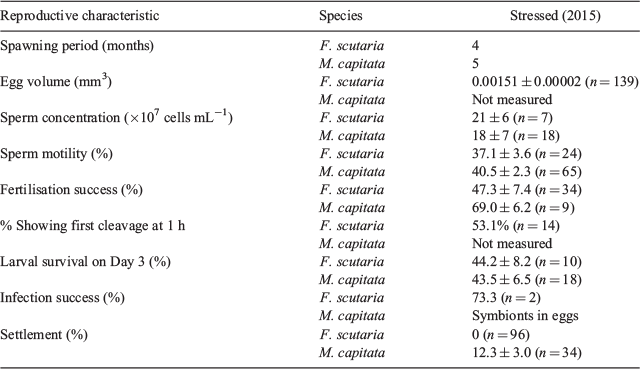
When coral experience a 1°C increase in temperature for ≥6 weeks, they begin to bleach (Glynn and D’Crox 1990). The 2014 warming resulted in widespread bleaching of most species in Kaneohe Bay, with 82%– 87% of the M. capitata, Porites compressa and F. scutaria exhibiting bleaching (Nielson 2014). In the present study, during the 2015 bleaching event, we examined spawning periods, egg morphometry, sperm concentration, fresh and cryopreserved sperm motility exposed to different concentrations of dimethyl sulfoxide (DMSO), time of first cleavage, larval survival with fresh and cryopreserved spermatozoa, infection success and settlement success for both F. scutaria and M. capita. In addition, we investigated how cryostress might affect fertilisation success, the time of first cleavage and larval survival.
Materials and methods
Coral collection and husbandry
Whole individuals of F. scutaria (n = 60) and fragments of M. capitata (n = 34) were collected from the shallow patch reefs in Kaneohe Bay, Hawaii, in May and June of 2015. The n-values reported throughout the paper indicate the number of individuals used; this is complicated by the fact that M. capitata is a hermaphrodite, but the eggs and spermatozoa of the same individual were never crossed. In addition, to ensure genetic diversity, corals were collected from different patch reefs throughout the entire bay. Coral were maintained in flowing seawater tables connected directly to Kaneohe Bay with natural temperature and lower light exposure throughout the spawning season. The F. scutaria colonies were kept in the same tanks after spawning, but M. capitata were returned to the reefs. Collection was performed with the appropriate permits from the state of Hawaii’s Department of Land and Natural Resources (Special Activity Permits #SAP 2011: 2011–1; 2012: 2012–63, 2013: 2013–47; 2014: 2015–17). No institutional ethics approval was required for any of the experimental research described herein.
We had collected consistent data for spawning periods, sperm motility and sperm concentration and post-thaw motility for the two species between 2010 and 2015, and then used data from other time periods for comparison.
Gamete collection
Gametes were collected using different approaches for each species, as described previously (Hagedorn et al. 2012, 2015b). Briefly, whole F. scutaria were collected on over 16 patch reefs throughout Kaneohe Bay in June 2015. Two to four days after the full moon in July, August and September, individuals were isolated in bowls and allowed to spawn. Spawning was observed to occur between 1630 and 1830 hours. Once spawning was observed, our research team monitored each individual to check for the release of either eggs or spermatozoa. Eggs were allowed to remain in the bowl until spawning ceased, and were then collected for experiments described below. Spermatozoa were collected as they were released and immediately transferred to a 50-mL centrifuge tube using a transfer pipette. Every effort was made to keep the spermatozoa as concentrated as possible for use in the experiments described below. Eggs and spermatozoa from each individual were kept separate until assessed, and then IVF was conducted.
In contrast, M. capitata spawned 0–2 nights after the new moon. To ensure genetic diversity, fragments (~10 cm × 10 cm) were collected from intact colonies located on at least 20 patch reefs throughout Kaneohe Bay. The M. capitata gametes were collected from both individuals identified in our tanks and multiple individuals from the surface of the south-facing lagoon at the Hawaii Institute of Marine Biology. To collect egg–sperm bundles from individuals, fragments were isolated in floating pots, at 1930 hours and spawning commenced around 2100 hours. Bundles floated to the surface and were collected with 200-µL pipettes or transfer pipettes and were placed in 50-mL centrifuge tubes with approximately 5 mL eggs added to 10 mL seawater, which maintained a sperm concentration of 109 cells mL–1. Because this species has a toxin in the eggs that can affect the spermatozoa (Hagedorn et al. 2015b), the egg–sperm bundles were allowed to break apart without agitation. After breaking apart naturally, concentrated spermatozoa from each individual were removed from the bottom of each tube and eggs were gently washed with filtered seawater (FSW) by carefully refilling the tube, allowing the eggs to float to the surface and then removing the FSW below the eggs four to five times. For egg–sperm bundles from multiple individuals, bundles were collected as they were released to the surface of the lagoon with scoops. Approximately, 5 mL eggs was placed in 5 mL seawater in a 50-mL conical plastic tube to maintain the sperm concentration at 109 cells mL–1, as above. For the multiple individual collections in the lagoon, only spermatozoa were used for further testing.
Temperature measurements
Coral and larvae were maintained in an open system with sufficient flow (~16 L min–1) to maintain temperatures similar those on the reef. To understand the differences in the temperature profiles between our tanks and the bay that may contribute to warming stress, bay temperature was measured by an in situ weather station on Coconut Island and in the tanks by Hobo loggers (HOBO Pendant© Temperature/Light 64K Data Logger, Onset, Bourne, MA, USA) taking measurements every 2 min or single static measurements at 1200 or 1500 hours everyday with an Omega HH147U Data Logger Thermocouple (Stamford, CT, USA). Warming stress that can lead to bleaching has two key components: the temperature of the water and the intensity of the light (Lesser and Shick 1989; Ferrier-Pagès et al. 2007). To reduce irradiance, the tanks were covered with shade cloth.
Egg morphometry and assessment
After spawning, approximately 2–30 eggs from individual F. scutaria (n = 18) were assessed, using computer-aided morphometry (Olympus BX41, Center Valley, PA, USA, QImaging Retiga 200R camera, Surrey, BC, Canada and NIH Image J software, Bethesda, MD, USA), and a mean diameter and volume calculated. If any individual’s eggs were irregularly shaped, they were not used in further reproductive experiments. These data were compared with previous F. scutaria data taken in 2005 and 2007 using the same methods (Hagedorn et al. 2010).
In addition, during the imaging above, F. scutaria (n = 14) eggs (~100) were also scored for debris and mucus in the water and overall per cent abnormality (irregular shapes or sizes) using a scale from 0%, being no abnormal eggs, to 25%, 50%, 75% and 100% indicating progressively more abnormal eggs.
Sperm motility and concentration assessments
Sperm motility was assessed for each individual, colony or wild-caught pooled sample before deciding whether to include those spermatozoa in subsequent experiments as reported in previous studies (Hagedorn et al. 2012). Each night, a low threshold of motility was determined and spermatozoa below that threshold were not used further. From previous studies (Hagedorn et al. 2012), we have determined some guidelines for rapid assessments once the spermatozoa are collected. This was important because the time available to work with coral spermatozoa was limited to 2–5 h, depending on the species. Generally, the low threshold for coral sperm motility is approximately 50%, but during the studies in 2015 we accepted a threshold as low as 30% motility. In terms of sperm concentration, the low threshold depends on how the spermatozoa will be used after thawing. For fresh spermatozoa, concentrations ranging from 104 to 106 cells mL–1 in the in vitro culture are successful, depending on the species (Hagedorn et al. 2012). Generally, we used twice to three times this concentration in the in vitro culture with cryopreserved spermatozoa because its post-thaw motility is often 50% less than its starting value. An additional consideration was that after assessment only spermatozoa meeting both the concentration and motility requirements were pooled for use in subsequent experiments, because including inferior samples would result in little or no reproduction.
Spermatozoa were assessed for each individual F. scutaria or M. capitata fragment or wild-caught pooled sample (M. capitata) subjectively by eye following previously reported methods (Hagedorn et al. 2012).
During spawning, efforts were made to collect spermatozoa consistently from spawn to spawn. With F. scutaria, spermatozoa were collected as they were expelled from the mouth. In the case of M. capitata, 5 mL of egg–sperm bundles were collected with 10 mL FSW in a 50-mL centrifuge tube. Sperm concentration was measured using computer-aided sperm analysis (CASA; Hamilton Thorne, Ceros II System, Olympus BX41 with a ×10 objective and a green filter) system. A 3-µL sperm sample was loaded onto a specialised Standard Count 4 Chamber Slide (20 µm; REF SC20-01-04-B; Leja, The Netherlands) and three to five unique video fields for each sperm sample were acquired. Some replicate concentrations were taken using a haemocytometer to verify accuracy of the CASA system to assess concentration.
Sperm cryopreservation
Using methods described previously (Hagedorn et al. 2012), 0.5-mL samples of F. scutaria spermatozoa were mixed 1 : 1 with 20% DMSO in FSW in 1.8-mL cryovials, held for 20 min at 24–26°C, cooled at 20°C min–1 to –80°C, quenched in liquid nitrogen for at least 10 min, thawed in 30°C FSW and then post-thaw viability was assessed.
Fertilisation success
Although sperm concentration and motility are important, fertilisation success is the best indication that both spermatozoa and eggs are healthy. To examine this, we added either fresh or cryopreserved spermatozoa to fresh F. scutaria eggs. For F. scutaria, approximately 100 eggs from eight female F. scutaria were loaded into two parallel sets of 15-mL scintillation vials filled with 4.995 mL FSW, then 5 µL of either fresh (1 × 108 cells mL–1, resulting in 1 × 105 cells mL–1) or 5 µL cryopreserved spermatozoa (2 × 108 cells mL–1, resulting in 2 × 105 cells mL–1) were added to each vial and their fertilised eggs counted 4 h later. The two sets of eight vials were fertilised with two different sperm pools (A and B) of male spermatozoa (n = 3 males per pool at 55% and 36% fresh motility, respectively).
For M. capitata, only fresh spermatozoa were used to assess fertilisation success. IVF trials were conducted by adding one to two egg–sperm bundles from two different individuals (n = 9) into 5 mL FSW in 15-mL glass scintillation vials. After 60 min, the bundles fell apart and egg numbers were recorded. Spermatozoa in each vial yielded a final working concentration of 106 cells mL–1 and fertilised larvae were counted.
After 12 h, fertilisation success was assessed by counting the number of developing larvae in each vial under a dissecting microscope at ×10 magnification.
Development
To determine the effect of bleaching on embryo development, 50 F. scutaria eggs from each individual harvested after bleaching were placed into 5 mL FSW in a 15-mL scintillation vial after fertilisation and the time to first cleavage in F. scutaria individuals (n = 14) was determined. Seven treatments were used to examine the effects not only of sperm concentration, but also increasing concentrations of DMSO. When we use cryopreserved spermatozoa for fertilising fresh eggs, generally we try to freeze the spermatozoa at a concentration for F. scutaria at 108 cells mL–1 with 10% DMSO (Hagedorn et al. 2012). These samples were diluted 1 : 1000 for fertilisation, producing a concentration of 105 cells mL–1 and 0.01% DMSO in the vial. Too high a concentration of DMSO can cause developmental issues. To fully explore this, we used seven treatments in the present study: (1) samples with no spermatozoa; (2) spermatozoa at 104 cells mL–1; (3) spermatozoa at 104 cells mL–1 and 0.01% DMSO; (4) sperm at 105 cells mL–1; (5) sperm at 105 cells mL–1 and 0.1% DMSO; (6) spermatozoa at 106 cells mL–1; and (7) spermatozoa at 106 cells mL–1 and 1% DMSO. The time of first cleavage in the samples was examined on a Wild dissecting microscope at ×10 magnification 1, 2 and 4 h after spermatozoa were added to the scintillation vial.
Larval survival
To assess larval survival, we distributed eggs from 10 F. scutraria females equally into two 3-L bowls, each containing 500 000 eggs, with seawater reduced to 100 mL. Eggs were then fertilised with 1 mL fresh or cryopreserved spermatozoa (fresh motility 54%; n = 6 males), resulting in 5 × 105 cells mL–1 in the bowls with fresh spermatozoa and 1 × 106 cells mL–1 for the cryopreserved spermatozoa with an average post-thaw motility of 13%. Each bowl was gently swirled periodically for an hour, and then topped up to 3 L with FSW. All larvae in all bowls were cleaned within 12 h using a series of sieves and FSW and this was repeated every 24 h for 3 days. To assess survival, each bowl was mixed well and 10 subsamples of larvae (20 µL–1 mL) were taken, counted on a microscope (SZX7; Olympus) at a magnification of ×1.25 and then the samples replaced. If the sample volume was <1 mL, the counts were multiplied to equal a 1 mL volume and the mean number of larvae per bowl was reported as larvae mL–1.
In the present study, M. capitata egg–sperm fertilisation and survival were assessed by IVF. This entailed crossing one bundle each from two different individuals, allowing the bundles to fall apart and cross-fertilise in 5 mL FSW in a 15-mL glass scintillation vial. Over three nights, 34 unique crosses were completed representing 14 individuals, and their developing larvae were counted in each vial under a dissecting microscope at ×10 magnification 1 to 2 h after the bundles fell apart and at the end of 3 days. The number of eggs in each vial was maintained and assessed through larval survival and settlement.
Infection success
The ability of F. scutaria larvae to take up their Symbiodinium after bleaching was examined on Day 4 of their development. The surviving F. scutaria larvae were cleaned and divided into small bowls of either 100 or 50 mL FSW to yield 1000–2500 larvae mL–1. Then, 300 µL brine shrimp essence (frozen brine shrimp were crushed into FSW; the brine shrimp were then filtered out and only the supernatant used) was added to each small bowl of larvae 5 min before the addition of Symbiodinium to initiate a searching and feeding response from the larvae (Schwarz et al. 1999). Symbiodinium were extracted and cleaned as described previously (Schwarz et al. 1999; Hagedorn et al. 2010) and concentrated to a density of 1 × 107 mL–1 before being added to the small bowls of larvae to result in a final concentration of 1 × 106 mL–1 Symbiodinium in each bowl.
After infection, larvae were observed carefully over the 4-h infection period to make sure they were swimming. At 4 h after infection, larvae were cleaned several times by placing them in a 40 µm mesh sieve, rinsing with FSW until all excess Symbiodinium had been removed, and then transferred into larger bowls and returned to 3 L FSW. Larvae were cleaned by removing and replacing 90% of the FSW each morning for 2 days, and then infection success was assessed. At least 100 larvae from each bowl were examined using an Olympus SZX7 microscope and scored as infected if they contained at least one Symbiodinium.
Settlement success
The F. scutaria larvae from the two bowls (described above) that produced effective infections were used in settlement experiments on Day 6, as described previously (Schwarz et al. 1999). Briefly, infected larvae (n = ~100 per well) were placed in 12-well plastic plates (n = 4 per bowl) that had a 2- to 3-day biofilm. These plates were left in running water for 2 days, the larvae were counted and fresh FSW was renewed each day.
To get the Day 3 M. capitata larvae to settle (n = 14), they, were placed into 10 mL FSW in 10-mL petri dishes with 1 to 2 cm chips of crustose coralline algae. Over three nights, their settlement was assessed by counting the developing larvae in each vial under a dissecting microscope at ×10.
Statistical analysis
All data analyses were performed using Prism 6.0 (GraphPad, San Diego, CA, USA) and Excel (Microsoft, Redmond, WA, USA). All percentage data were log transformed before statistical analyses. For multiple group comparisons, normality was tested graphically and data analysed. All analyses and post-tests are specified in the Results, and all data are expressed as the mean ± s.e.m. For integer data, Chi-squared analysis was performed.
Results
Spawning period
From 2010 to 2015, the spawning of the two species was monitored each month (Table 1). F. scutaria spawned from June to October, depending on whether the full moon was early in the month. If it was later in the month, then usually there were only four spawns in a summer, whereas an early full moon might produce five spawns in a summer. M. capitata spawned three times in a summer, usually June through August, but in 2014 it spawned May–July.

|
Temperature measures
From assessing the meteorological records collected from the weather station on Coconut Island, Kaneohe Bay experienced year-long warming starting in the summer of 2014 through 2015, whereby the average sea surface temperature in August and October was 1–1.8°C and 2.5–2.6°C, respectively, above average August and October sea surface temperatures for the previous 3 years. The temperatures in our tanks tracked the temperatures on the reef ±1°C in mean monthly values from 2010 to 2013 and 2015 (see Fig. 1). In 2015, Kaneohe Bay was 2°C warmer in July and August than 2010–13. The mean monthly photosynthetically active radiation (PAR) values for Kaneohe Bay in July and August for 2015 were 1110 ± 63 and 910 ± 74 µE m–1 s–1, respectively. However, the mean irradiance levels in our tanks in 2015 were 1.1% or less of these values.
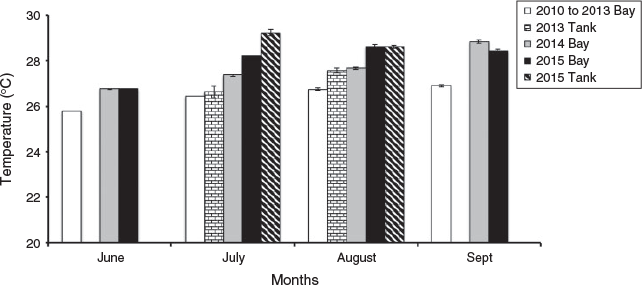
|
Egg morphometry
Measurements of mean F. scutaria egg volumes were compared with previously recorded data (Hagedorn et al. 2010) from 2007 and 2005 (Fig. 2; Table 2), demonstrating differences in the mean egg volumes for all years, with 2015 having the smallest overall mean volume (P < 0.05, analysis of variance (ANOVA); F = 154, Tukey’s multiple comparison test). The 2015 mean egg volume exhibited a –50.9% (2007) and –10% (2005) difference in volume with comparison years.
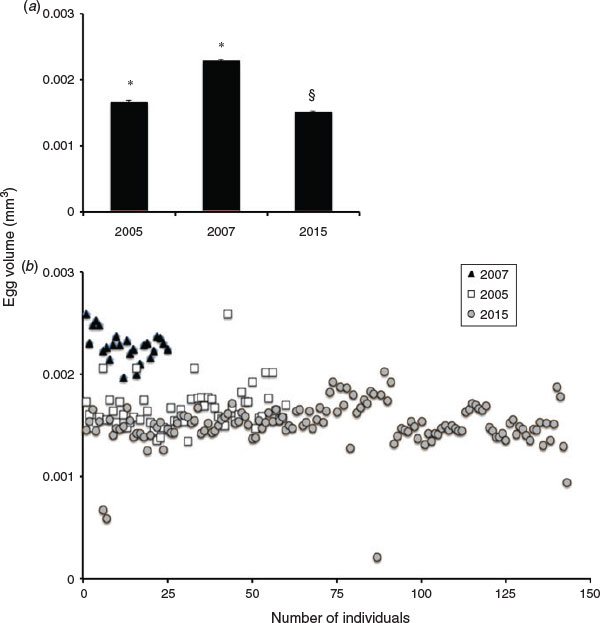
|
|
Generally, in non-bleaching years, F. scutaria eggs were extremely translucent white in colour and uniform in size, with no debris or mucus surrounding the eggs. In 2015, all the egg samples from the 14 stressed individuals had debris and mucus surrounding the eggs, which was unusual. Moreover, 5 ± 2% of the eggs were abnormal in shape, ranging from 0% to 50% abnormal.
Sperm concentration
There was no difference in the mean sperm concentration for F. scutaria between 2015 and 2010 and for M. capitata between 2015 and 2011 (Table 3; P > 0.05, Kruskal–Wallis test, Dunn’s multiple comparison test). For F. scutaria, the sperm concentrations produced in July and August were averaged for each year, whereas those for June and July were averaged for M. capitata. These months were chosen because these are the peak breeding months for these species during each summer.

|
Sperm motility
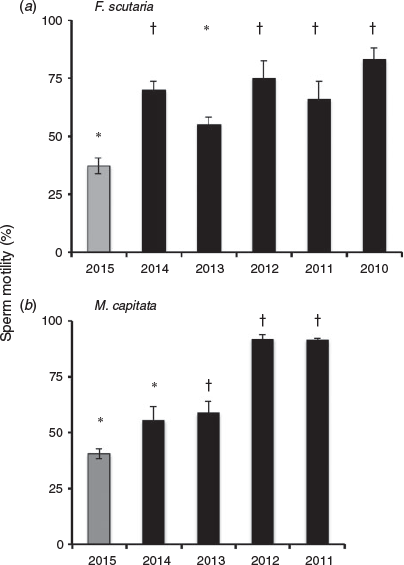
We examined total progressive motility for both F. scutaria and M. capitata spermatozoa over multiple spawning events during the summer of 2015 and compared them with previous motility data from 2014 to 2010 (unpubl. data, Fig. 3). In 2015, F. scutaria had an average total progressive motility of 37.1 ± 3.6% (n = 24), whereas M. capitata had total progressive motility of 40.5 ± 2.3% (n = 65). These mean total progressive percentage motilities were one of lowest of the four or five comparison years (F. scutaria: P < 0.05, ANOVA, F = 7.2, Dunnett’s multiple comparison test; M. capitata: P < 0.05, ANOVA, F = 26.2, Dunnett’s multiple comparison test).
|
Fertilisation success
The fertilisation success of stressed F. scutaria eggs and fresh spermatozoa (105 cells mL–1; n = 22) was examined. Fertilisation success in 2015 was 47.3 ± 7.4% (n = 34), and reduced compared with a previous year (2009), when fertilisation success was 82.5 ± 4.8% (n = 14; P < 0.05, Student’s t-test). This reflected a percentage change of 42.8% in the fertilisation success between the two years. In contrast, M. capitata had a fertilisation success of 69.0 ± 6.2% (Table 2), but we did not have any previous data to compare this with.
Time of first cleavage
We examined the time of first cleavage of the eggs from 2015 F. scutaria individuals (n = 14) exposed to different concentrations of fresh spermatozoa (104, 105 and 106 cells mL–1). All the eggs from each individual were subdivided into seven treatments (see below for additional treatments). We were not concerned about the fertilisation success within each treatment, only when the first developmental cleavage occurred within the treatment. Only 53.3% of eggs from stressed individuals (n = 14) exposed to fresh spermatozoa cleaved at 1 h, with the remainder cleaving after 2 h (35.6%) or 4 h (11.1%; n = 45 samples).
Larval survival
During 2015, F. scutaria larvae survival was measured in a pair-matched set of bowls (n = 10) where eggs from the same female were evenly divided into the bowls. One-half of the bowls was fertilised with a pooled fresh sperm sample (n = 6 males, 54% motility) and the other half was fertilised with the same spermatozoa, but cryopreserved (see below). In addition, M. capitata larvae were counted in their vials every day.
Larval survival from Day 1 to Day 3 was relatively similar for both F. scutaria (44.2 ± 8.4%) and M. capitata (43.5 ± 6.5%; Table 2).
Infection success
In addition to these previously described reproductive characteristics reported here, the successful uptake or ‘infection’ of Symbiodinium for F. scutaria was assessed in our in vitro system. At the end of 6 days, two bowls of larvae remained with 114 and 39 larvae mL–1. These two bowls had reasonable infection success of 82.9% and 63.6%, respectively. However, before the introduction of the symbionts, the larvae were examined and 20%–40% of the larvae in the bowls were misshapen and often fused into large associations. Nevertheless, even these developmentally abnormal larvae took up Symbiodinium. Infection success in M. capitata was not examined because of horizontal transmission of symbionts from adult to egg.
Settlement success
One day after placement into the settlement trays, the F. scutaria larvae were counted and were still actively swimming. However, the following day most were dead, with a few lone settlers in two wells. The M. capitata settlement was counted 14 days after fertilisation. Preliminary observations in June 2015 showed that larvae were still swimming 10 days after development with no settlement. Therefore, settlement was checked daily from 10 days after development until settlement was observed on Day 14; average settlement was 12.3 ± 3.0%.
Cryopreservation success
During the 2015 bleaching year, several reproductive factors were affected even further when freezing and/or cryoprotectants were involved. First, the fertilisation success of F. scutaria using cryopreserved spermatozoa was reduced during the 2015 bleaching season (Fig. 4). Specifically, the fresh Pools A and B had a fertilisation success of 48.8 ± 8.4% and 42.3 ± 10.3%, respectively, whereas their cryopreserved counterparts had a fertilisation success of 3.6 ± 1.5% and 2.8 ± 1.0% s.e.m., respectively (P < 0.05, ANOVA, F = 15.8). Overall, there was an approximate tenfold loss of fertilisation success with the use of cryopreservation in this stressed system.
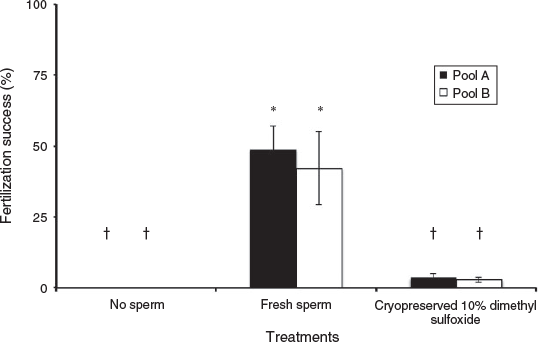
|
As described above, almost half the embryos produced from fresh eggs and fresh spermatozoa suffered a developmental delay. In parallel subsamples, we examined the time of first cleavage of the eggs exposed to fresh sperm concentrations of 104, 105 and 106 cells mL–1 and increasing concentrations of DMSO (0.01%, 0.1% and 1%) in the vial. The cryoprotectant concentration increased with the increasing concentration of spermatozoa. Only 18.8% of eggs from individuals exposed to fresh spermatozoa with cryoprotectants cleaved at 1 h, with the bulk of samples cleaving at 2 h (50.0%) and less at 4 h (31.3%; n = 16 samples). In general, the addition of DMSO delayed development considerably from 1 to 2 h compared with developing larvae from eggs and spermatozoa with no added cryoprotectant (P < 0.05, Chi-squared, d.f. 3.709).
Finally, compared with their fresh counterparts, the larvae fertilised from cryopreserved spermatozoa were severely compromised, with an average percentage loss of larvae from Day 1 to Day 3 >90% (see Fig. 5). To determine whether the larvae produced from fresh spermatozoa and cryopreserved spermatozoa survived similarly in 2015, their larval survival curves were compared and found to be different (P < 0.05, linear regression, F = 7.5).
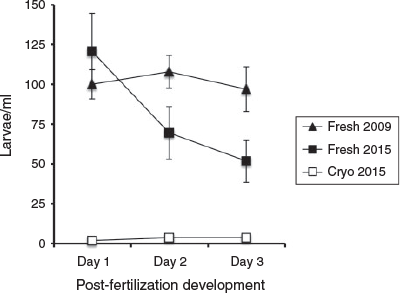
|
Discussion
During the ongoing bleaching event of 2015, we investigated the reproductive characteristics of two coral species, namely F. scutaria and M. capitata, because approximately 85% of their populations suffered bleaching in the previous autumn of 2014 in Kaneohe Bay, Hawaii (Nielson 2014). We examined and compared our observed measurements with previous data to determine whether bleaching may have an effect on a specific reproductive characteristic, such as sperm motility or sperm concentration. In addition, we were interested in how the addition of cryostress might further affect fertilisation success, the time of first cleavage and larval survival. Our goal was to determine the effects of bleaching on coral reproduction; however, much greater efforts are needed to examine natural variation and to help tease out the changes directly related to bleaching stress. Nevertheless, our data (Table 2) form the foundation for further reproductive studies.
Because our coral were held in tanks in an open seawater system, one of our concerns was that we were studying a ‘tank temperature effect’ and not the physiological changes due to the bleaching coral in the bay. To understand this, Fig. 1 charts the summer temperature in the bay from 2010 to 2015 and in our tanks in 2013 and 2015. In August of 2013, compared with the temperature in the bay, the temperature in the tanks was approximately 1°C greater, producing normal spawning and no bleaching. This same 1°C elevated difference was observed in July 2015 in the tanks, but because this 1°C difference in 2013 did not affect the spawning or physiology of the coral we assumed that this held true for 2015 as well. Thus, we felt confidant that temperature variations produced by our tanks and seawater system were not unduly affecting the physiological changes of the coral.
In terms of egg volume (see Fig. 2), this was reduced for F. scutaria during 2015 compared with the previous year’s measurements (Hagedorn et al. 2010). Although the mean size of eggs for 2005 was only slightly larger than 2015, there was a 10% difference in mean egg volume. If you compare 2015 and 2007, there was a 51% difference in mean egg volume. In 2005, there were a few eggs that were as large as the ones in 2007, but no eggs in the other years matched the small eggs in 2015 (Fig. 2). These shifts in yearly mean volume could reflect natural variation or possibly gradations in recovery. In 2002, there was a bleaching event in Kaneohe Bay (Jokiel and Brown 2004), and it is clear that bleaching can have multiple year effects on reproduction (Levitan et al. 2014). Perhaps, 2005 was still a recovery year and in 2007 the eggs were completely recovered.
In addition, the time of first cleavage appeared to be delayed in 2015. The first cleavage in F. scutaria occurs at 1 h (Krupp 1983), but only 53.3% of eggs from 2015 individuals cleaved at this time, with the remaining 46.7% of treatments cleaving at 2 or 4 h. Krupp’s (1983) study did not mention any variation in the time of first cleavage, so again these two observation could be just natural variation or it could be that the stress of warming-reduced egg volume extended the time of first cleavage in F. scutaria.
We did not measure egg volume or the number of eggs per bundle in M. capitata because handling of the eggs releases a toxin in the eggs that has a negative effect on reproduction (Hagedorn et al. 2015b). However, a previous study suggested that heterotrophic feeding can maintain the number of eggs in the egg–sperm bundles during bleaching for M. capitata (Cox 2007), suggesting that nutritional augmentation, through additional feeding or supplied food, may have some capacity to off-set some reproductive effects of bleaching.
Although sperm concentration did not appear to change much over the 5-year analysis, the mean sperm motility for F. scutaria was reduced by 47% in 2015 compared with mean motility for previous 4 years, and M. capitata had a reduced mean motility of 44% in 2015 compared with the previous 3 years. Sperm motilities recorded in 2015 are significantly different than many of the previous years, nevertheless, there is at least one year in the sequence that it is not.
There was a further loss of reproductive capacity using cryopreservation, as the result of a cascade of stressors. With nearly a 50% decrease in mean fresh sperm motility in 2015, this loss has a profound effect on the applicability of cryopreservation to systems that have suffered bleaching. Once fresh, stressed spermatozoa was cryopreserved in 2015, there was a further loss of motility after thawing and reduced fertilisation success. In non-bleaching years, the fertilisation success of F. scutaria can be as high as 82% with fresh spermatozoa and 60% with cryopreserved spermatozoa (Hagedorn et al. 2012). In 2015, fertilisation success for F. scutaria was approximately 46% with fresh spermatozoa and approximately 3% with cryopreserved spermatozoa, suggesting a tenfold loss of fertilisation success for cryopreserved spermatozoa with bleaching stress. Moreover, the cryoprotectant solutes altered development, causing a delay in the timing of first cleavage. These extraordinary losses with both temperature and cryopreservation stressors on the reproductive system make this conservation method almost untenable during and after bleaching years.
In vitro larval survival for F. scutaria can vary. Occasionally, a whole 3-L bowl will be lost, but this is rare. In non-bleaching years, our larval survival is excellent. For example, F. scutaria reared in bowls containing approximately 98 larvae mL–1 (n = 8 bowls) exhibited 3% loss from Day 1 to Day 3 (Hagedorn et al. 2009). However, in 2015, this was not the case. Stocking densities varied from 32 to 136 larvae mL–1 and were dependent on the number of eggs a particular female produced, but there was a 48% reduction in larval survival for F. scutaria by Day 3. To determine whether the larvae produced from fresh spermatozoa survived the same as in a previous year, the larval survival curves between 2015 and 2009 were compared and found to be different (P < 0.05, linear regression, F = 4.9). When the larval survival of M. capitata was compared with historical data (Vermeij et al. 2009), a 31% reduction in survival was noted. Clearly more data are needed on larval survival to identify whether this is a natural variation in spawning years or effects due to bleaching.
Infection and settlement success may have also negatively affected F. scutaria in 2015. These data suggest that the mean infection success (73.3%) for F. scutaria was reduced compared with previous reports of 96.8% (Schwarz et al. 1999). In addition, in 2015, M. capitata settlement was reduced by over 50% compared with 1997 numbers (Field 1998) and 100% in F. scutaria. This complete loss of F. scutaria settlement contrasted with the 70% settlement success observed at Day 8 in 1999 (Schwarz et al. 1999). This could have been due to the multiple effects on reproduction over the 8 days before settlement, especially the larval malformations we observed in 2015, or it could also have been due to our inexperience with settling F. scutaria. However, we settle many other species quite effectively.
Rising water temperature has already greatly affected coral populations around the world, and bleaching stress may affect species differently. Recently, a long-term study on Orbicella in the Caribbean (Levitan et al. 2014) suggested that, at the population level, the negative effects of bleaching persist for many years, causing a reduction in spawning; however, these authors did not find the timing of spawning or coral survivorship or tissue loss to be altered. Our studies corroborate some of the findings of Levitan et al. (2014) in that the spawning period was not altered in M. capitata or in F. scutaria during bleaching. During the 2015 F. scutaria spawning season, due to elevated temperatures and bleaching we observed a reduction in egg volume, delayed development and increased larval abnormalities, which match well with previous observations. In Acropora palmata, under bleaching conditions development was accelerated, developmental abnormalities increased and larval survival was reduced (Randall and Szmant 2009).
A recent report summarised the long history of anthropogenic changes in Kaneohe Bay over the past seven centuries, including agricultural and human effects resulting in near extinction of the coral in the bay (Bahr et al. 2015). These authors suggested that these stressors have made Kaneohe Bay coral ‘super resilient’, so the reproductive changes observed in 2015 may be far worse in other less-resilient areas. Moreover, the IPCC (2007) has predicted that coral bleaching will occur more frequently around the globe. So, regardless of whether the coral in Kaneohe Bay live through these and future bleaching events, their reproductive biology may have been affected by bleaching, suggesting that future adaptation may be limited by this reduction in reproduction.
Long-term studies of coral reproduction suggest dire consequences for coral living in areas of high bleaching incidence because the long-term negative reproductive effects may ‘bridge the interval between subsequent bleaching events’ (Levitan et al. 2014), as was observed on Heron Island (Ward et al. 2002), thus compounding these reproductive issues. Given the extraordinary energy expenditure needed for coral spawning and the potentially terrible reproductive results before settlement (noted here), we wonder whether a coral might have a greater reproductive advantage in future years, if it does not spawn after a bleaching event (Anthony et al. 2009; Schoepf et al. 2013). In fact, this is the effective reproductive strategy of many species (Ward et al. 2002).
The IPCC (2007) considers warming oceans and their related and compounding threats, producing severely reduced reproduction, recruitment and adaptation of coral, as one of the greatest threats to the survival of coral reefs worldwide. As bleaching events become more frequent, the ability to bank and conserve coral to maintain the biodiversity and genetic diversity ex situ will become significantly reduced. Thus, it is imperative that while genetic diversity is still high in these populations and bleaching events not present in an area for that year, intensive efforts should be made to bank coral species during non-bleaching periods. This may require very flexible conservation strategies to move to reefs with optimal, non-bleaching climates in a particular year to maximise cryopreservation benefits.
Acknowledgements
These experiments were supported through funds from the Smithsonian Institution, the Hawaii Institute of Marine Biology, the Roddenberry Foundation, the Matthew Frank Foundation, the Anela Kolohe Foundation and the Cedarhill Foundation. The authors thank Dr Budhan Pukazhenthi for comments on the manuscript and Evan Phillipson for his technical assistance. This manuscript was approved for publication by the Hawaii Institute of Marine Biology as Contribution #1642.
References
Anthony, K. R. N., Hoogenboom, M. O., Maynard, J. F., Grottoli, A. G., and Middlebrook, R. (2009). Energetics approach to predicting mortality risk from environmental stress: a case study of coral bleaching. Funct. Ecol. 23, 539–550.| Energetics approach to predicting mortality risk from environmental stress: a case study of coral bleaching.Crossref | GoogleScholarGoogle Scholar |
Bahr, K. D., Jokiel, P. L., and Toonen, R. J. (2015). The unnatural history of Kāne’ohe Bay: coral reef resilience in the face of centuries of anthropogenic impacts. PeerJ 3, e950.
| The unnatural history of Kāne’ohe Bay: coral reef resilience in the face of centuries of anthropogenic impacts.Crossref | GoogleScholarGoogle Scholar | 26020007PubMed |
Baker, A. C. (2003). Flexibility and specificity in coral–algal symbiosis: diversity, ecology, and biogeography of Symbiodinium. Annu. Rev. Ecol. Evol. Syst. 34, 661–689.
| Flexibility and specificity in coral–algal symbiosis: diversity, ecology, and biogeography of Symbiodinium.Crossref | GoogleScholarGoogle Scholar |
Baker, A. C., Glynn, P. W., and Riegl, B. (2008). Climate change and coral reef bleaching: an ecological assessment of long-term impacts, recovery trends and future outlook. Estuar. Coast. Shelf Sci. 80, 435–471.
| Climate change and coral reef bleaching: an ecological assessment of long-term impacts, recovery trends and future outlook.Crossref | GoogleScholarGoogle Scholar |
Cox, E. F. (2007). Continuation of sexual reproduction in Montipora capitata following bleaching. Coral Reefs 26, 721–724.
| Continuation of sexual reproduction in Montipora capitata following bleaching.Crossref | GoogleScholarGoogle Scholar |
Cox, E. F., and Ward, S. (2002). Impact of elevated ammonium on reproduction in two Hawaiian scleractinian corals with different life history patterns. Mar. Pollut. Bull. 44, 1230–1235.
| Impact of elevated ammonium on reproduction in two Hawaiian scleractinian corals with different life history patterns.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XovFeiur4%3D&md5=83036450cb016fea54ebc9e6ac6d7894CAS | 12523521PubMed |
Ferrier-Pagès, C., Richard, C., Forcioli, D., Allemand, D., Pichon, M., and Shick, J. M. (2007). Effects of temperature and UV radiation increases on the photosynthetic efficiency in four scleractinian coral species. Biol. Bull. 213, 76–87.
| Effects of temperature and UV radiation increases on the photosynthetic efficiency in four scleractinian coral species.Crossref | GoogleScholarGoogle Scholar | 17679722PubMed |
Field, S. (1998) Settlement Biology of Larvae of Montipora verrucosa and Porites lobata in Hawaii. In: ‘Reproduction in Reef Corals Results of the 1997 Edwin W. Pauley Summer Program in Marine Biology, Technical report No. 42’. (Eds E. F. Cox, D. A. Krupp, P. L. Jokiel) pp. 111–119. (University of Hawaii: Honolulu.)
Fitt, K., Brown, B. E., Warner, M. E., and Dunne, R. P. (2001). Coral bleaching interpretation of thermal tolerance limits and thermal thresholds in tropical corals. Coral Reefs 20, 51–65.
| Coral bleaching interpretation of thermal tolerance limits and thermal thresholds in tropical corals.Crossref | GoogleScholarGoogle Scholar |
Glynn, P. W. (1991). Coral reef bleaching in the 1980s and possible connections with global warming. Trends Ecol. Evol. 6, 175–179.
| Coral reef bleaching in the 1980s and possible connections with global warming.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3M7hsVegtw%3D%3D&md5=3675aa344053b98e92a3139cb713722eCAS | 21232450PubMed |
Glynn, P. W. (1993). Coral reef bleaching: ecological perspectives. Coral Reefs 12, 1–17.
| Coral reef bleaching: ecological perspectives.Crossref | GoogleScholarGoogle Scholar |
Glynn, P. W. (1996). Coral reef bleaching: facts, hypotheses and implications. Glob. Change Biol. 2, 495–509.
| Coral reef bleaching: facts, hypotheses and implications.Crossref | GoogleScholarGoogle Scholar |
Glynn, P. W., and D’Crox, L. (1990). Experimental evidence for high temperature stress as the cause of El Niño-coincident coral mortality. Coral Reefs 8, 181–191.
| Experimental evidence for high temperature stress as the cause of El Niño-coincident coral mortality.Crossref | GoogleScholarGoogle Scholar |
Goreau, T. J. (1992). Bleaching and reef community change in Jamaica: 1951–1991. Am. Zool. 32, 683–695.
| Bleaching and reef community change in Jamaica: 1951–1991.Crossref | GoogleScholarGoogle Scholar |
Hagedorn, M., Pan, R., Cox, E. F., Hollingsworth, L., Krupp, D., Lewis, T. D., Leong, J. C., Mazur, P., Rall, W. F., MacFarlane, D. R., Fahy, G., and Kleinhans, F. W. (2006). Coral larvae conservation: physiology and reproduction. Cryobiology 52, 33–47.
| Coral larvae conservation: physiology and reproduction.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD28%2FosFOgsw%3D%3D&md5=f973d8db01d574914abf49cc0510151bCAS | 16337183PubMed |
Hagedorn, M., Carter, V. L., Hollingsworth, L., Leong, J. C., Kanno, R., Borneman, E. H., Petersen, D., Laterveer, M., Brittsan, M., and Schick, M. (2009). Ex situ culture of Caribbean and Pacific coral larvae comparing various flow-through chambers. In ‘Proceedings of the Smithsonian Marine Science Symposium’. (Eds M. A. Lang, I. G. Macintyre, K. Rutzler) pp. 259–268. (Smithsonian Institution Press: Washington DC.)
Hagedorn, M., Carter, V. L., Ly, S., Andrell, R. M., Yancey, P. H., Leong, J. A., and Kleinhans, F. W. (2010). Analysis of internal osmolality in developing coral larvae, Fungia scutaria. Physiol. Biochem. Zool. 83, 157–166.
| Analysis of internal osmolality in developing coral larvae, Fungia scutaria.Crossref | GoogleScholarGoogle Scholar | 19938981PubMed |
Hagedorn, M., Carter, V., Martorana, K., Paresa, M. K., Acker, J., Baums, I. B., Borneman, E., Brittsan, M., Byers, M., Henley, M., Laterveer, M., Leong, J. A., McCarthy, M., Meyers, S., Nelson, B. D., Petersen, D., Tiersch, T., Uribe, R. C., Woods, E., and Wildt, D. (2012). Preserving and using germplasm and dissociated embryonic cells for conserving Caribbean and Pacific coral. PLoS One 7, e33354.
| Preserving and using germplasm and dissociated embryonic cells for conserving Caribbean and Pacific coral.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XktF2it7Y%3D&md5=af5f090fc61fe99829dea0d422c6a042CAS | 22413020PubMed |
Hagedorn, M., Carter, V., Zuchowicz, N., Phillips, M., Penfield, C., Shamenek, B., Vallen, E. A., Kleinhans, F. W., Peterson, K., White, M., and Yancey, P. H. (2015a). Trehalose is a chemical attractant in the establishment of coral symbiosis. PLoS One 10, e0117087.
| Trehalose is a chemical attractant in the establishment of coral symbiosis.Crossref | GoogleScholarGoogle Scholar | 25629699PubMed |
Hagedorn, M., Farrell, A., Carter, V., Zuchowicz, N., Johnston, E., Padilla-Gamino, J., Gunasekera, S., and Paul, V. (2015b). Effects of toxic compounds in Montipora capitata on exogenous and endogenous zooxanthellae performance and fertilization success. PLoS One 10, e0118364.
| Effects of toxic compounds in Montipora capitata on exogenous and endogenous zooxanthellae performance and fertilization success.Crossref | GoogleScholarGoogle Scholar | 25714606PubMed |
Intergovernment Panel on Climate Change (IPCC) (2007) Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. (Eds S. Solomon, D. Qin, M. Manning, Z. Chen, M. Marquis, K. B. Averyt, M. Tignor, H. L. Miller.) (Cambridge University Press: Cambridge.)
Jokiel, P. L., and Brown, E. K. (2004). Global warming, regional trends and inshore environmental conditions influence coral bleaching in Hawaii. Glob. Change Biol. 10, 1627–1641.
| Global warming, regional trends and inshore environmental conditions influence coral bleaching in Hawaii.Crossref | GoogleScholarGoogle Scholar |
Jokiel, P. L., and Coles, S. L. (1990). Response of Hawaiian and other Indo-Pacific reef corals to elevated temperature. Coral Reefs 8, 155–162.
| Response of Hawaiian and other Indo-Pacific reef corals to elevated temperature.Crossref | GoogleScholarGoogle Scholar |
Kawaguti, S. (1944). On the physiology of reef corals VI. Study on the pigments. Palao Trop. Biol. Stn. Stud. 2, 617–674.
Krupp, D. A. (1983). Sexual reproduction and early development of the solitary coral Fungia scutaria (Anthozoa: Scleractinia). Coral Reefs 2, 159–164.
| Sexual reproduction and early development of the solitary coral Fungia scutaria (Anthozoa: Scleractinia).Crossref | GoogleScholarGoogle Scholar |
LaJeunesse, T. C., Thornhill, D. J., Cox, E. F., Stanton, F. G., Fitt, W. K., and Schmidt, G. W. (2004). High diversity and host specificity observed among symbiotic dinoflagellates in reef coral communities from Hawaii. Coral Reefs 23, 596–603.
Lesser, M. P., and Shick, M. J. (1989). Effects of irradiance and ultra-violet radiation on photoadaptation in the zooxanthellae of Aiptasia pallida:primary production, photoinhibition, and enzymatic defenses against oxygen toxicity. Mar. Biol. 102, 243–255.
| Effects of irradiance and ultra-violet radiation on photoadaptation in the zooxanthellae of Aiptasia pallida:primary production, photoinhibition, and enzymatic defenses against oxygen toxicity.Crossref | GoogleScholarGoogle Scholar |
Levitan, D. R., Boudreau, W., Jara, J., and Knowlton, N. (2014). Long-term reduced spawning in Orbicella coral species due to temperature stress. Mar. Ecol. Prog. Ser. 515, 1–10.
| Long-term reduced spawning in Orbicella coral species due to temperature stress.Crossref | GoogleScholarGoogle Scholar |
McLaughlin, J. J. A., and Zahl, P. A. (1959). Axenic Zooxanthellae from various invertebrate hosts. Ann. N. Y. Acad. Sci. 77, 55–72.
| Axenic Zooxanthellae from various invertebrate hosts.Crossref | GoogleScholarGoogle Scholar |
Nielson, B. (2014). ‘DAR Coral Bleaching Rapid Response Surveys September–October 2014.’ Available at http://dlnr.hawaii.gov/reefresponse/files/2014/10/DARCoralBleachingSrvy_Results_10.28.2014.pdf
Padilla-Gamiño, J. L., and Gates, R. D. (2012). Spawning dynamics in the Hawaiian reef-building coral Montipora capitata. Mar. Ecol. Prog. Ser. 449, 145–160.
| Spawning dynamics in the Hawaiian reef-building coral Montipora capitata.Crossref | GoogleScholarGoogle Scholar |
Padilla-Gamiño, J. L., Weatherby, T. M., Waller, R. G., and Gates, R. D. (2011). Formation and structural organization of the egg–sperm bundle of the scleractinian coral Montipora capitata. Coral Reefs 30, 371–380.
| Formation and structural organization of the egg–sperm bundle of the scleractinian coral Montipora capitata.Crossref | GoogleScholarGoogle Scholar |
Padilla-Gamiño, J. L., Hedouin, L., Waller, R. G., Smith, D., Truong, W., and Gates, R. D. (2014). Sedimentation and the reproductive biology of the Hawaiian reef-building coral Montipora capitata. Biol. Bull. 226, 8–18.
| 24648203PubMed |
Randall, C. J., and Szmant, A. M. (2009). Elevated temperature affects development, survivorship, and settlement of the elkhorn coral, Acropora palmata (Lamarck 1816). Biol. Bull. 217, 269–282.
| 20040751PubMed |
Relman, D. A. (2008). ‘Til death do us part’: coming to terms with symbiotic relationships. Nat. Rev. Microbiol. 6, 721–724.
| ‘Til death do us part’: coming to terms with symbiotic relationships.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhtFWitL3M&md5=5eea378441c1e4ceb9894878bd990144CAS | 19086265PubMed |
Schoepf, V., Grottoli Aé, G., Warner, M. E., Cai, W. J., Melman, T. F., Hoadley, K. D., Pettay, D. T., Hu, X., Li, Q., Xu, H., Wang, Y., Matsui, Y., and Baumann, J. H. (2013). Coral energy reserves and calcification in a high-CO2 world at two temperatures. PLoS One 8, e75049.
| Coral energy reserves and calcification in a high-CO2 world at two temperatures.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXhs1Cns7zM&md5=55824d3badc2f5ced4b483917a590000CAS | 24146747PubMed |
Schwarz, J. A., Krupp, D. A., and Weis, V. M. (1999). Late larval development and onset of symbiosis in the scleractinian coral Fungia scutaria. Biol. Bull. 196, 70–79.
| Late larval development and onset of symbiosis in the scleractinian coral Fungia scutaria.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC2Mvls1OntA%3D%3D&md5=d135093880959ffbef894132996471ebCAS | 25575388PubMed |
Szmant, A. M., and Gassman, N. J. (1990). The effects of prolonged ‘bleaching’ on the tissue biomass and reproduction of the reef coral Montastrea annularis. Coral Reefs 8, 217–224.
| The effects of prolonged ‘bleaching’ on the tissue biomass and reproduction of the reef coral Montastrea annularis.Crossref | GoogleScholarGoogle Scholar |
Vaughan, T. W. (1914). Sketch of the geologic history of the Florida coral reef tract and comparisons with other coral reef areas. J. Wash. Acad. Sci. 4, 26–34.
Vermeij, M. J., Smith, J. E., Smith, C. M., Vega Thurber, R., and Sandin, S. A. (2009). Survival and settlement success of coral planulae: independent and synergistic effects of macroalgae and microbes. Oecologia 159, 325–336.
| Survival and settlement success of coral planulae: independent and synergistic effects of macroalgae and microbes.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD1M7ksVeiug%3D%3D&md5=25aed2dbabf660cce9093016381aeeebCAS | 19050932PubMed |
Ward, S., Harrison, P., and Hoegh-Guldberg, O. (2002) Coral bleaching reduces reproduction of scleractinian corals and increases susceptibility to future stress. In ‘Proceedings of the Ninth International Coral Reef Symposium’, Volume 2, 23–27 October 2000, Bali. (Eds M. K. Moosa, S. Soemodihardjo, A. Soegiarto, K. Romimohtarto, A. Nontji, S. Suharsono.) pp. 1123–1128. (Ministry of Environment, Indonesian Institute of Sciences, International Society for Reef Studies: Bali.)