

An angled rack with a bypass and a nature-like fishway pass Atlantic salmon smolts downstream at a hydropower dam
D. Nyqvist A , J. Elghagen A B , M. Heiss C and O. Calles A DA River Ecology and Management (RivEM), Department of Environmental and Life Sciences, Karlstad University, SE-651 88 Karlstad, Sweden.
B Elghagen Fiskevård, Långgatan 10, SE-265 31 Åstorp, Sweden.
C BNGF GmbH – Büro für Naturschutz-, Gewässer- und Fischereifragen, D-82396 Pähl, Germany.
D Corresponding author. Email: olle.calles@kau.se
Marine and Freshwater Research 69(12) 1894-1904 https://doi.org/10.1071/MF18065
Submitted: 28 February 2018 Accepted: 21 June 2018 Published: 27 September 2018
Journal Compilation © CSIRO 2018 Open Access CC BY-NC-ND
Abstract
Hydropower dams disrupt longitudinal connectivity and cause fragmentation of river systems, which has led to declines in migratory fish species. Atlantic salmon smolts rely on intact longitudinal connectivity to move downstream from rearing habitats in freshwater to feeding grounds at sea. Smolts often suffer increased mortality and delays when they encounter hydropower plants during their downstream migration. Currently, there are few examples of downstream passage solutions that allow safe and timely passage. We assessed the performance of two passage solutions at a hydropower dam, namely, an angled 15-mm rack with a bypass and a large nature-like fishway. The performance of these new fish passage solutions was evaluated by tracking radio-tagged Atlantic salmon smolts as they encountered the facilities. The radio-tagged smolts passed the dam 9.5 h after release (median) and exhibited a dam-passage efficiency of 84%, with passage rates increasing with body length. Fish passage occurred through both the rack bypass and the nature-like fishway. The passage efficiencies were 70–95% for the rack bypass and 47% for the nature-like fisway. The new fish passage facilities resulted in improved passage conditions at the site, confirming that angled racks with bypasses as best-practise solutions for downstream passage, but also that large nature-like fishways may act as downstream passage routes for salmon.
Additional keywords: downstream passage, fish passage solution, migration, passage efficiency.
Introduction
Fish migrate for feeding, reproduction and refuge, often in response to temporal or ontogenetic changes (Lucas et al. 2001). In regulated rivers, dams block migratory routes and hinder fish movements among habitats, which has caused fragmentation, declines and sometimes even local extinction of migratory species (Northcote 1998; Jonsson et al. 1999; Marmulla 2001).
The need to offer migratory fish a safe route past dams has been acknowledged for hundreds of years (Montgomery 2004). Despite this, fish passage solutions are lacking at many dams (Calles et al. 2013a; Nieminen et al. 2017), and where solutions exist, they typically target upstream passage of strong salmonid swimmers (Katopodis and Williams 2012). However, during the past decade, downstream passage solutions have gained more attention, both in research and management (Whitney and Council 1997; Calles et al. 2013c).
Downstream passage solutions typically pass migrating fish via bypasses, spill gates or fishways (Johnson and Dauble 2006; Colotelo et al. 2012; Calles et al. 2013b). Many downstream-migrating fish typically follow bulk flow (Coutant and Whitney 2000) and where little water is assigned to passing fish, structural guidance devices are often needed to guide fish to preferred passage routes (Calles et al. 2013a, 2013b; Nyqvist et al. 2017a). Low-sloping turbine intake racks (or screens) use the existing water current to guide downstream-migrating fish towards one or several bypass entrances, and can either be arranged with a low slope from the bottom to the surface (inclined rack) or with a low slope from one side of the intake channel to the other (angled rack; DWA 2005; Calles et al. 2013b). Inclined and angled racks have been applied to pass several fish species with variable success (Nettles and Gloss 1987; EPRI 2001; Gosset et al. 2005; Tomanova et al. 2017), and are considered as best practice in Sweden (Calles et al. 2013a). However, as with most fish passage solutions, the efficiencies with which inclined and angled racks guide different fish species and life-stages have been little researched, and there are no published evaluations of Atlantic salmon smolt downstream passage efficiencies at angled racks with bypasses.
Even at dams with fish passage solutions, migrating fish often experience migratory failure. Downstream-migrating fish might suffer direct or delayed mortality as an effect of spill, bypass, or turbine passage (Muir et al. 2001; Ferguson 2005; Ferguson et al. 2006; Serrano et al. 2009; Nyqvist et al. 2017b). Delay can also be an important cause of migratory failure in relation to dams; delayed migrating fish may suffer predation, accumulation of stress, loss of migration motivation, elevated energetic costs, and mistimed arrival to feeding or spawning sites (McCormick et al. 1999; Muir et al. 2006; Kemp and Williams 2008). As the effectiveness varies greatly among individual fish passage solutions and sites, evaluating such remedial measures is needed to ensure that the goal is achieved, i.e. the restoration of longitudinal connectivity (Roscoe and Hinch 2010; Bunt et al. 2012; Noonan et al. 2012), and to learn from successes and failures.
Atlantic salmon (Salmo salar) is a socially and economically important migrating fish species. Its life-cycle typically consists of a juvenile stage in the river, migration to feeding areas at sea, and a return migration to its river of origin to spawn (Jonsson and Jonsson 2011). Juvenile salmon, before leaving the river, go through a series of behavioural, physiological and morphological changes, called smoltification. The smolts become silvery, more streamlined, lose positive rheotaxis and territoriality, begin shoaling, change their visual pigments, and increase their salinity tolerance, preparing them for a life in the marine environment (McCormick et al. 1998). Smolt status is temporally constrained. If smolts are delayed during their migration, loss of smolt characters, including salinity tolerance and migratory urge, occurs (McCormick et al. 1998). Atlantic salmon smolts are considered obligatory migrants, making them suitable and important study objects for dam passage performance.
Here, we evaluate the functionality of two widely promoted but rarely studied fish passage solutions, namely an angled turbine intake rack and a large nature-like fishway, using radio-telemetry. The angled rack and bypass is considered as a state-of-the-art downstream passage solution, whereas the large nature-like fishway mainly constitutes an upstream passage solution (Johnson and Dauble 2006; Calles et al. 2013a). We present dam downstream passage efficiency and time to passage (delay), as well as the route-specific passage efficiencies of both the angled-rack bypass and the nature-like fishway. Further, fish passage performance is typically affected by fish characteristics and environmental conditions (Roscoe and Hinch 2010; Castro-Santos 2012). We hypothesise that smolt passage behaviour might vary with hydraulic environment, fish size, illumination and the progression of spring (Hesthagen and Garnås 1986; McCormick et al. 1998; Nyqvist et al. 2017b). Hence, we take these factors into account when analysing rates of approach and passage through the rack bypass and the nature-like fishway (Castro-Santos and Perry 2012).
Materials and methods
Study river
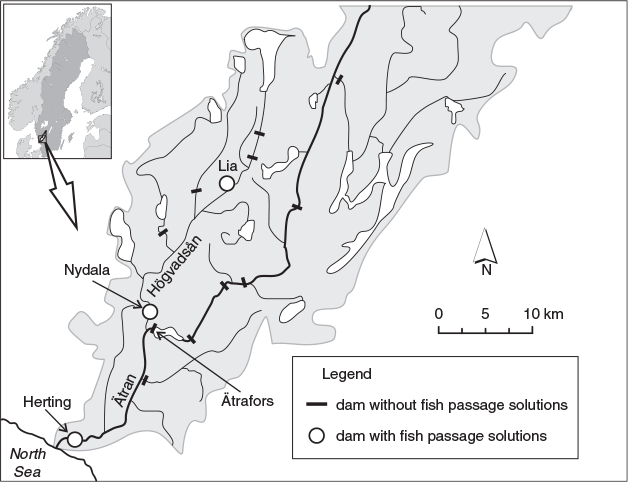
The River Ätran (56°52′55″N, 12°28′46′E′) runs through south-western Sweden and enters the North Sea (Kattegatt) in the city of Falkenberg. The river is 243 km long and has a mean annual discharge of 57 m3 s–1 (1990–2011, range 20–319 m3 s–1; Olofsson 2013). The study dam, Herting, situated ~3 km upstream from the sea, is the first of many hydropower dams in the river system, and the only dam in the main stem with upstream and downstream fish passage solutions. Atlantic salmon have access to spawning and rearing grounds within 24 km of the river up to the second hydropower dam (Ätrafors), and within 34 km of the tributary River Högvadsån (Fig. 1; Calles et al. 2010, 2012, 2013b).
|
Herting hydropower dam
The Herting dam consists of two powerhouses (Fig. 2), Herting 1 (H1) and Herting 2 (H2). Both powerhouses were built during the first half of the 20th century (H1 1903; H2 1945) and are equipped with Kaplan turbines. H1 is equipped with two turbines and has an intake capacity of 40 m3 s–1 (#1: 250 rpm, 15.0 m3 s–1; #2: 187 rpm, 25.0 m3 s–1), whereas H2 has one turbine (187 rpm, 25.0 m3 s–1).

|
Several fish passage solutions have been implemented at the dam (Fig. 2; Hebrand 2012). In the turbine intake channel (H1), an angled bar rack with a gap width of 15 mm was installed to guide fish to a bypass with two entrances. The rack consists of horizontally arranged hydrodynamically shaped composite bars (CompRack, Halmstad, Sweden), and is angled 30° to the main direction of flow. The rack has a wet area of 80 m2 at the minimum allowed water level, resulting in a maximum normal velocity vector of 0.5 m s–1 (40 m3 s–1/80 m2). A full-depth bypass, with a total intake capacity of 3 m3 s–1, is located at the downstream end of the rack. The bypass is equiped with a fish trap, where fish can be caught for monitoring purposes. During normal operation, 0.6 m3 s–1 is discharged through an orifice close to the bottom (200 × 200 mm; W × H) and a slot at the surface (300 × 650 mm). The downstream passage facility is a modified version of the design by Ebel (2013), and is considered as best practice for Sweden (Calles et al. 2013a).
A large nature-like fishway, constructed on the old riverbed, has a required minimum discharge of 11 m3 s–1 (or the current total discharge in the river). Water is spilled into the nature-like fishway through the hydraulic fishway entrance (5.4 m3 s–1) and over two adjacent weirs (5.6 m3 s–1). When the total discharge is higher than 51.6 m3 s–1, i.e. the sum of the base flow in the fishway and bypass and the turbine intake capacity, the water level in the forebay increases and more water spills over the weirs into the nature-like fishway. To facilitate passage, and direct more water to the nature-like fishway, the H2 hydropower plant is operated only when few fish are expected to migrate, i.e. December–February. In addition, water can also be spilled into the tail-race of H2 via upward opening spill-gates at a 90 m3 s–1 capacity.
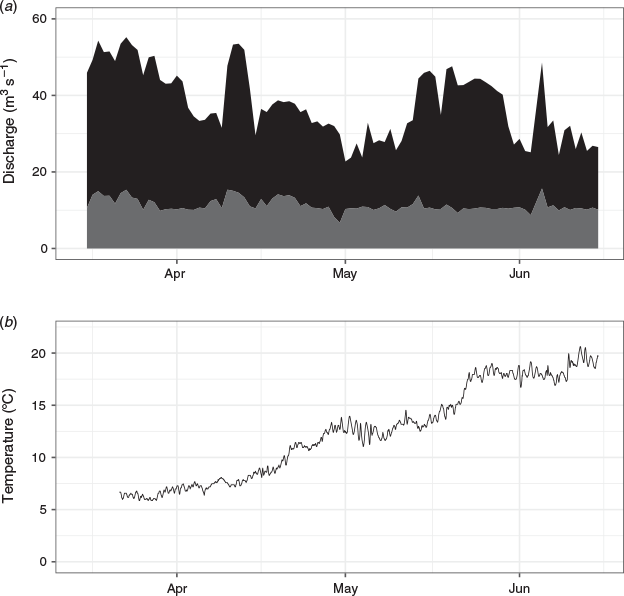
During the study period, total average river discharge was 37 m3 s–1. On average, 24.5 m3 s–1 (range 19.1–30.7 m3 s–1) were used for electricity production in H1, with the bypass passing, on average, 2.4% (range 2.0–3.1%) of the discharge in the turbine intake channel. This resulted in a normal velocity range of 0.24–0.38 m s–1 at the rack, which was well below the maximum normal velocity of 0.5 m s–1. On average, 31% (range = 21–35%) of total discharge passed via the nature-like fishway (Fig. 3). Downstream-migrating smolts could, therefore, theoretically pass via the nature-like fishway or, after having entered the turbine intake channel, via the bypass or the rack and turbines. Small quantities of water (<1 m3 s–1) leaked through the spill gates and H2 during the study period.
|
Radio-telemetry
Wild downstream-migrating Atlantic salmon smolts were caught in a Wolf-trap at the Nydala dam in River Högvadsån (Fig. 1), transported in aerated tanks, radio-tagged (Model ATS F1525, weight 0.65 g, life 21 days; 40–42 pulses min–1; Advanced Telemetry Systems (ATS), Isanti, MN, USA) and released ~400 m upstream (n = 40) and 100 m downstream (control; n = 9) of the Herting H1 powerhouse where the angled rack and bypass facility is situated (~500 m between release sites; Fig. 2). Before release, fish were observed for an average of 4.25 h in a 70-L tank with constant supply of river water, to ensure recovery after the tagging procedure. Smolts released upstream of the dam had to pass the dam to be able to continue their downstream migration, whereas the smolts released downstream of the dam allow for a simple control for handling and tagging effects. Fish were anesthetised (Benzocaine, Sigma–Aldrich, Saint Louis, MO, USA) before surgery, and tagging followed the standard procedure for surgical implants of trailing-whip antenna radio-transmitters (Jepsen et al. 2002; Liedtke and Rub 2012; Thorstad et al. 2013). Fish were measured for length (mm), weight (0.1 g), height (immediately posterior of the dorsal fin; mm) and degree of smoltification (Tanguy et al. 1994). The total procedure, including anaesthesia and surgery, lasted from 3.53–7.13 min (mean 4.96 min). The tagged fish were observed in a 70-L holding tank with constant supply of river water to ensure full recovery after the tagging procedure. Only apparently healthy and fully recovered fish were used in the study.
Yagi-antennas with stationary automatic receivers (N = 14, Model R4500S, ATS) tracked fish movement in the river, with particular focus on the area immediately upstream of the Herting dam (for details, see Heiß 2015). A dropper antenna (a striped coaxial cable) was placed in the bypass fish-collection trap, to detect fish present in the trap. A Yagi-antenna was located ~1.1 km downstream of the H1 powerhouse, and 1.0 km downstream of the release site of the control group, to detect fish continuing their migration towards the sea. To verify automatic-generated data, fish were manually positioned daily, using a manual receiver (R2000, ATS) and a three-element Yagi-antenna. Fish were tracked from release until they migrated to sea, or were considered dead in the dam area.
Statistical analysis and definitions
The area upstream of the dam was partitioned into an approach zone and an intake zone (Fig. 2). Radio detections defined presence in the zones. Within the approach zone, visits to the vicinity of the fishway were also recorded, but used only for calculations of passage efficiency for the fishway (Fig. 2). Consequently, fish present in the approach zone could transition into thintake zone, pass via the nature-like fishway or reject the approach zone in an upstream direction. Fish present in the intake zone could pass via the bypass, the rack and turbines or return upstream. Periods of >2 h without radio detection were defined as departure from the approach zone, and periods of >30 min without detection departure from the intake channel. Departed individuals returning to the respective zones were considered new visits.
We used Cox-regression, a type of time-to-event analysis, to model effects of fixed and time-varying covariates on rates (proportion or probability over time) of intake approach and fishway passage for fish present in the approach zone, and bypass passage rates for fish present in the intake zone. In time-to-event analysis, delay and the proportion of fish passed are analysed simultaneously. The analysis also takes into account fish that do not pass or approach, but were available to do so, when analysing event rates. (Castro-Santos and Haro 2003; Hosmer et al. 2008; Castro-Santos and Perry 2012). All tagged fish present in the respective zones were considered available to pass, and were included in the analyses. Fish that left the zone through an alternative route (i.e. were no longer available to pass) were included as censored observations. The censored fish were considered available to pass, and were included in the analyses until the censoring event occurred. Fish that left the zone but returned were again included in the analysis (considered available to pass). Some fish visited the zones (approach zone and intake channel) several times. To avoid pseudoreplication, we considered a new visit to be dependent on previous visits, and so all models were stratified by visit number to each respective zone (Allison 2010).
Individual body length, Julian day (as a proxy for the progression of spring), day or night, and relative discharge (fishway discharge/total discharge) were included as covariates in the candidate models for rate of approach (entry) to the intake channel and fishway passage. The same set of covariates were used to construct the candidate models for the analysis of bypass-passage rate for fish present in the intake channel, but relative discharge was substituted with discharge in the intake channel (≈generation in H1). All combinations of three or fewer covariates and interactions were used as candidate models. Minimisation of Akaike information criterion (AIC) was used to select the best model among all candidate models (Burnham and Anderson 2003). Models with an AIC-value of –2 or lower from the null (ΔAICNull < –2) model and within 2 AIC units from the best model (ΔAICmin > 2) were considered good models (Burnham and Anderson 2003). When more than one competing model were good, and constituting derivates of each other, only the best model with the fewest parameters was chosen (Richards 2008; Schwinn et al. 2017). The assumption of proportionality of hazard was explicitly tested for all good models (Fox 2002).
Dam passage efficiency was defined as the proportion of fish successfully negotiating the hydropower plant, by any route, after visiting the area upstream of the dam. Passage time was defined as the time from release to passage of the dam. Tagged fish that were recaptured in the bypass trap were assumed to survive de-tagging and release.
Route-specific passage efficiency (commonly referred to as fish guidance efficiency, FGE, Scruton et al. 2008) was defined as the proportion of fish attempting to pass the dam via a specific fish passage solution that found, entered and successfully negotiated the fish passage solution. Fish passing via the intake channel but not confirmed as bypass passages by the dropper antenna or the fish-collection trap are considered potential turbine passages or bypass passages (intake passage). Hence, the number of fish passing via the bypass is reported as a range, ranging from ‘confirmed’ to ‘confirmed + potential’ bypass passages. Route-specific passage efficiencies were estimated for the two fish passage solutions, i.e. the rack bypass and the nature-like fishway. The route-specific passage efficiencies are, henceforth, referred to as ‘bypass passage efficiency’ and ‘fishway passage efficiency’. Fish were considered attempting to pass the dam when entering the approach zone (Fig. 2). Similarly, fish were considered attempting to pass the dam via the rack bypass when entering the intake zone and via the fishway when entering the area in the vicinity of the fishway (Fig. 2). In addition, route-specific passage efficiency during the first visit was defined as the number of fish passing via the rack bypass on their first visit, divided by the number of fish visiting the turbine intake channel (Bunt et al. 2012).
All statistical analyses were performed using R (R Foundation for Statistical Computing, Vienna, Austria). The ‘survival’ package (ver. 2.38; T. M. Therneau and T. Lumley, see https://CRAN.R-project.org/package=survival) was used for time-to-event analysis, data were plotted with ggplot2 (Wickham 2016), and dplyr (ver. 0.7.4, H. Wickham, R. Francois, L. Henry and K. Müller, see https://CRAN.R-project.org/package=dplyr), and sqldf (ver. 0.4-11, G. Grothendieck, see https://CRAN.R-project.org/package=sqldf) was used for data management.
Fish-collection trap
The fish-collection trap was emptied every morning during the entire study period for monitoring purposes. All caught fish were identified to species level. Fish were checked for injuries, dead fish were noted, and total length was measured to the closest millimetre (because of high abundance of salmonid smolts, only subsamples of randomly picked individuals were measured). All fish caught in the fish trap were then released downstream of the bypass channel. For tagged fish, the transmitter was first removed (i.e. fish were de-tagged) before they were released.
Results
In total, 49 smolts were tagged and released upstream and downstream of the Herting dam 14–23 April 2014. The tagged smolts had a mean length (±1 s.d.) of 146 ± 6 mm, with no difference between fish released upstream and downstream of the dam (Student’s t-test, P = 0.41). The tagged fish were, on average, 24 ± 2 mm high. Fish were released in the evening, at sunset (1950–2120 hours) in batches of four to eight fish and tracked from first release until 1 May, when all fish considered alive had continued towards the sea.
All control fish (n = 9) migrated downstream and were detected by the downstream receiver, 1.0 km downstream of the release site, and continued towards the river mouth (i.e. 100% survival). Median duration from release to arrival at the downstream receiver was 8.3 h, corresponding to a speed of ~109 m h–1 (Table 1).

|
In total, 37 of the 40 fish released upstream of the dam were detected in the forebay (Fig. 2, ‘approach zone’) almost immediately after release (median = 3 min, IQR = 1–5 min, range: 1–101 min). The three fish not detected after release were omitted from the dataset. From the 37 fish detected in the forebay, 34 passed the dam, of which 31 where considered as successful passages, resulting in a dam passage efficiency of 84% (Table 1). Fifteen fish (44%) passed via the nature-like fishway and 19 fish (56%) via the turbine intake channel. Of the fish that passed via the turbine intake channel, 14 were confirmed to have passed via the bypass (recaptured), whereas five fish were not collected in the bypass and, thus, had an unclear passage route (i.e. passed via the turbines or the bypass and escaping capture; Table 1). The complete opening of the bypass gate did not seem to induce passage because no radio-tagged smolts entered the trap within 4 h before or after such spill events. Median time from release to passage was 10.5 h, corresponding to ~40 m h–1 for fish passing the dam via the intake channel and 9.6 h or ~21 m h–1 for fish passing the dam via the fishway (Table 1). The three fish failing to pass the dam took 2 days (n = 2) and 16 days (n = 1) from first arrival to last departure. Duration from release to passage (distance was 200 m to the fishway and 400 m to the rack bypass) for fish released upstream of the dam was not different from the time from release to continued downstream migration (~1 km) for control fish (Wilcoxon signed rank test, P = 0.5; Table 1).
One fish was lost while passing via the nature-like fishway (7%), remaining in the nature-like fishway until the end of the study, whereas all fish passing via the turbine intake channel, and not collected in the bypass, successfully continued their downstream migration. Fish collected in the bypass were detagged, so as to recover the radio-transmitter, and released to continue their downstream migration. Two fish (14%, n = 14) were found stranded and dead on the dewatering rack in the collection facility. The fish collected in the bypass trap spent a median of 11.6 h (IQR = 10.9–12.2; range = 1.9–19.3 h) in the trap between passage and release.
Most fish passed on their first visit to the dam (median number of visits = 1), but individual fish visited the dam one to three times before passage. The three fish that did not pass the dam also visited the approach zone one to three times. All 15 fish passing via the nature-like fishway did so on their first visit to the approach zone, after a median duration of 9.6 h (Table 1). In total, 32 of the 37 smolts entered the vicinity of the fishway (Fig. 2) at least once and so the 15 individuals passing via the nature-like fishway corresponded to a fishway passage efficiency of 47% (Table 1). Longer fish passed via the nature-like fishway at higher rates than did shorter fish, and fishway-passage rate was higher later in spring (Table 2a). Fish approached and entered the turbine intake channel one to five times. Median time from release to first approach or entry was 77 min (IQR = 30 min to 26 h, range = 6 min to 7 days). For all approaches, i.e. including also fish having returned to the approach zone from the intake channel, median time to entry into the intake channel was 98 min (IQR = 38 min to 6.6 h, range = 6 min to 7 days). No good model was found for rate of intake channel approach (Table 2b), but sample size was low and this should not rule out any potential effects existing in nature.

|
Twenty fish visited the turbine intake channel. Of these fish, 19 passed via a turbine intake channel route, 14 were confirmed bypass passages and five potential bypass passages (Table 1). This resulted in a bypass passage efficiency of 70–95%. The corresponding numbers for the first visit to the intake channel were 11 confirmed and four potential bypass passages, a first-attempt bypass-passage efficiency of 55–75%. Although most fish visited the intake channel only once before passage, individual fish made one to five visits to the intake channel. The median duration of visits to the intake channel resulting in passage was 5.5 min (IQR = 2.3–44 min, range = 1 min to 16.8 h), whereas visits resulting in upstream rejection of the intake zone lasted for a median of 182 min (IQR = 22–475 min, range = 6 min to 21.7 h). Bypass passage rate was higher for longer fish (Table 2c). For the time-to-event analysis of bypass passage rate, non-confirmed bypass passages were excluded (however, a similar result was obtained when running the test with these fish included as bypass passed fish).
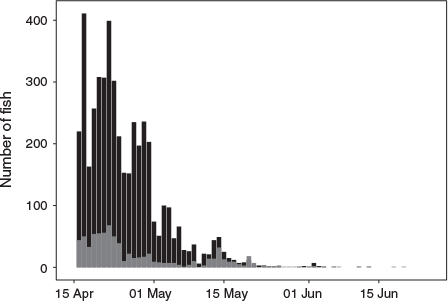
In total, 5904 individual fish, belonging to 19 different fish species were caught in the bypass fish-collection trap between 15 April and 29 June 2014. Salmonids dominated catches, with 4747 Atlantic salmon smolts and 798 brown trout smolts being recorded caught in the trap during the study period (Fig. 4). Half of the trapped Atlantic salmon smolts had been caught by 22 April, 75% by 29 April and the last Atlantic salmon smolt was caught on 25 June. For brown trout, the corresponding dates were 22 April (50%), 1 May (75%) and 20 June (last fish; Fig. 4). In addition, 50 kelts (12 salmon, 38 trout) and 15 fallback spawners (10 salmon, 5 trout) were caught in the trap.
|
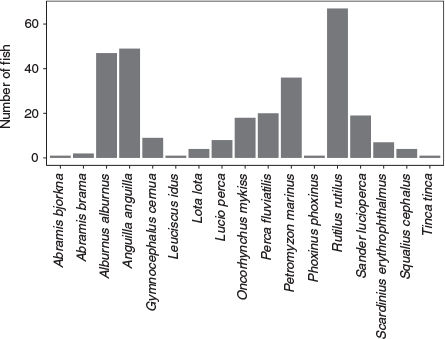
Other species frequently caught in the trap were roach (Rutilus rutilus), eel (Anguilla Anguilla) and common bleak (Alburnus alburnus; Fig. 5). A total number of 133 individual fish (2.2%) was found dead in the trap, either in the collection box (n = 115) or stuck on the dewatering rack (n = 18). It was not known to what extent untagged fish drifted dead to the trap, or died after being caught.
|
Discussion
Salmon smolt dam passage efficiency at the Herting dam was reasonably high as 84% of the radio-tagged fish detected in the forebay of the dam successfully passed the dam. Fish passing the dam used the nature-like fishway and the bypass to almost the same extent. Most had passed the dam within 2 days of release, which was comparable to the duration from release to continued downstream migration for fish released downstream of the dam. Unexpectedly, the mortality rate was higher in the bypass (14%) than in the reservoir (8%; failed to pass and were likely predated), which means that the dam passage success would have been >90% if the unforeseen bypass-induced mortality could be avoided.
From a methodological point of view, survival in the present study is defined as successful migration (including passage for fish released upstream of the dam) towards the sea, past the most downstream receiver, 1 km downstream of the dam. This does not take into account potential drifting of dead fish (Calles et al. 2013b; Havn et al. 2017), nor delayed mortality from injuries or stress acquired in dam passage (Muir et al. 2001; Ferguson et al. 2006); however, a previous study in which dead radio-tagged eels were released at the same site showed that no eels drifted to sea in spite of the average discharge being almost twice as high as in this study (Calles et al. 2012). The smolt turbine passage data from the same study showed that smolts killed in the turbines ended up close to the tail-race area of the Herting power plant. Also, three fish failed to pass the dam despite being present in the area upstream of the dam for a prolonged period of time. Failure to pass is often as important in limiting migratory success as is route-specific mortality (Nettles and Gloss 1987; Nyqvist et al. 2017b). In this particular case, fish were released in the proximity of the dam and part of the observed passage failure could be due to some individuals not continuing migration after release, i.e. losses caused by fish handling rather than owing to suboptimal passage facility performance. Extensive failure to initiate migration after release has been reported for both hatchery and wild smolts (Spicer et al. 1995; Larsen et al. 2016; Nyqvist et al. 2017c). However, among the control fish, all fish successfully migrated downstream. The lentic, potentially predator-rich environment upstream of the dam, may have had a negative effect on both initiation of migration and survival of initial disoriented fish (Calles and Greenberg 2009; Schwinn et al. 2017).
The majority of the fish released upstream of the dam where detected in the forebay (approach zone) almost immediately on release, and, hence, we could not distinguish delay caused by the dam itself (passage delay) from delay caused by fish handling (post-release delay; Nyqvist et al. 2017c). To allow this, fish should have been released further upstream. However, time from release to passage for fish released upstream of the dam was not different from the time from release to continued downstream migration for control fish released downstream of the dam, indicating little delay at the dam.
Longer fish passed at higher passage rates, both via the nature-like fishway and via the bypass. This might be due to generally higher swimming speeds in larger fish (Peake and McKinley 1998), but could, at least for bypass passage rates, indicate better fish-guiding effects of the rack on larger fish (Nyqvist et al. 2017d). Fish also passed via the nature-like fishway at higher rates later in the season, perhaps because of faster movements at higher temperatures or increased migratory urge as the migration season progressed (Dingle 2006; Jonsson and Jonsson 2011; Nyqvist et al. 2017c). Surprisingly, relative discharge did not affect fishway passage or approach to the intake channel, as was seen for Atlantic salmon kelts at the same dam (Nyqvist et al. 2017a). This is likely to be the result of low sample size and low variation in relative discharge, and should not be interpreted as movements being independent of discharge.
Fish passing via the bypass are caught in a trap for monitoring purposes. Interestingly, the bypass trap was the location where the tagged fish experienced the highest relative mortality. Of the fish caught in the trap, 14% (2 of 14 fish) died on the dewatering rack connected to it. Among all fish collected in the trap during spring, 2% were collected dead. The tagged fish caught in the trap and then released spent on average 12 h in the trap. Prolonged confinement and handling can have post-release effects on migratory survival. Fish collection for monitoring provides important information about migration periods, fish abundance and length relationships, but is also time-consuming, and comes with a cost (mortality, stress, delay) for the migrating fish. In addition to optimising the bypass for fish passage, i.e. minimising the negative effect, adapting the period and intensity of collection to the information needed are upfront ways to improve the survival of fish passing the bypass. Improvements were, in fact, made to the bypass dewatering rack and the trap after the completion of this study, resulting in a reduced numbers of dead fish. Nevertheless, carrying out monitoring of fish passage solutions without negatively affecting fish passage success remains a challenge.
The dam passage efficiency for smolts at Herting after modifications was reasonably high (84%), especially when taking into account that this is the only hydroelectric dam where smolts pass in River Ätran and the average passage efficiency recorded for downstream passage solutions (68.5%, Noonan et al. 2012). Nettles and Gloss (1987) reported that turbine entrainment of land-locked Atlantic salmon was reduced after the installation of an angled 25-mm trash rack on the Boquet River, but because of substantial losses recorded upstream of the plant, passage efficiencies were not reported. Havn et al. (2018) reported a dam passage efficiency of 75–84% for Atlantic salmon smolts for a facility with inclined 10-mm racks and multiple bypass entrances on the River Sieg; the losses mainly occurred in the reservoir and inside the bypass facility, i.e. similar to our findings at Herting. Tomanova et al. (2017) observed that 85% of PIT-tagged hatchery salmon smolts passed via safe routes, i.e. bypass passage efficiency was 85%, at a small-scale plant on Gave d’Oloron River after an inclined (26°) 20-mm rack was installed. Several slat spacings were evaluated for Atlantic salmon smolts at a Louver facility at Hadley Falls, Connecticut River, which showed an increasing bypass passage efficiency with a decreasing slat spacing, from 80% at 305-mm spacing to 91% (86–97%) at 76-mm spacing (Harza Engineering Company and RMC Environmental Services Inc. 1992, 1993; Stira and Robinson 1997). Another well studied Louver facility with a 100-mm slat spacing can be found at Grand-Falls Windsor on the Exploits River, where Atlantic salmon smolt bypass efficiency was repeatedly studied for different settings, with the results ranging 23–73% (Scruton et al. 2002, 2003, 2004, 2007, 2008). The highest downstream passage efficiencies reported for juvenile salmonids originate from the T. W. Sullivan project on the Willamette River, where an angled 38-mm bar rack and a high-flow bypass had a 100% bypass-passage efficiency both for juvenile chinook and steelhead (Karchesky et al. 2008).
Unexpectedly, the observed dam passage efficiency for smolts at Herting was even higher before the installation of the low-sloping rack and the associated modifications of the bypass (90%; Calles et al. 2012). However, the passage success recorded before modifications may have been inflated, as most of the smolts (69%) passed through the racks and turbines on their way downstream, and the extent of delayed mortality could not be quantified. The current bypass passage efficiency recorded at Herting could be as high as 95%, but also as low as 70%, because some fish passed the H1 powerhouse without being caught or detected in the bypass trap and because the resolution of the telemetry data was not high enough for us to be able to rule out that some fish passed through the racks and turbines. Regardless of the exact efficiency, the recorded rack bypass passage efficiency of 70–95% was considerably higher than the 17% recorded for the old bypass, lacking a guiding rack, at the exact same location (Calles et al. 2012). The current passage conditions at Herting, with most fish passing the dam via the fishway and rack bypass instead of through the turbines, has likely reduced the risk of diffuse delayed mortality. Especially, the current passage conditions, together with later modifications of the trap and a shift to limited periods of monitoring during smolt migration, has further contributed to increased dam-passage efficiency at the dam.
Overall, fish passed the dam with little delay, and with a reasonably high survival, using both the nature-like fishway and the bypass, guided by the angled rack. An even higher passage performance at the same dam was observed for Atlantic salmon kelts (Nyqvist et al. 2017a) and European eel (Anguilla anguilla, O. Calles, J. Elghagen, D. Nyqvist and P. A. Nilsson, unpubl. data). The Herting fish passage facility is one of few examples with documented high passage efficiencies with limited delays for both salmon (smolts and kelts) and silver eels. Moreover, the bypass trap data showed that the angled rack also guides several additional fish species to the bypass, albeit at unknown efficiencies. Last, the study also highlighted the need to consider potential fish mortality in the monitoring process, both when constructing traps and designing monitoring programs.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Acknowledgements
Thanks go to Peter Olsson and Simon Karlsson for hard work in the field, and to Sven-Erik and Berit Möller for allowing us to collect smolts from the Wolf-trap at Nydala mill. The study was funded by the Swedish Agency for Marine and Water Management and Falkenberg Municipality. The study was performed under ethical permission from the Swedish Board of Agriculture (85-2013).
References
Allison, P. D. (2010). ‘Survival Analysis using SAS: a Practical Guide.’ (SAS Institute: Cary, NC, USA.)Bunt, C., Castro‐Santos, T., and Haro, A. (2012). Performance of fish passage structures at upstream barriers to migration. River Research and Applications 28, 457–478.
| Performance of fish passage structures at upstream barriers to migration.Crossref | GoogleScholarGoogle Scholar |
Burnham, K. P., and Anderson, D. R. (2003). ‘Model Selection and Multimodel Inference: a Practical Information–Theoretic Approach.’ (Springer Science & Business Media: New York, NY, USA.)
Calles, O., and Greenberg, L. (2009). Connectivity is a two-way street: the need for a holistic approach to fish passage problems in regulated rivers. River Research and Applications 25, 1268–1286.
| Connectivity is a two-way street: the need for a holistic approach to fish passage problems in regulated rivers.Crossref | GoogleScholarGoogle Scholar |
Calles, O., Olsson, I. C., Comoglio, C., Kemp, P. S., Blunden, L., Schmitz, M., and Greenberg, L. A. (2010). Size-dependent mortality of migratorymigratory silver eels at a hydropower plant, and implications for escapement to the sea. Freshwater Biology 55, 2167–2180.
| Size-dependent mortality of migratorymigratory silver eels at a hydropower plant, and implications for escapement to the sea.Crossref | GoogleScholarGoogle Scholar |
Calles, O., Karlsson, S., Hebrand, M., and Comoglio, C. (2012). Evaluating technical improvements for downstream migrating diadromous fish at a hydroelectric plant. Ecological Engineering 48, 30–37.
| Evaluating technical improvements for downstream migrating diadromous fish at a hydroelectric plant.Crossref | GoogleScholarGoogle Scholar |
Calles, O., E. Degerman, H. Wickström, J. Christiansson, S. Gustafsson, and I. Näslund. (2013a). Anordningar för upp-och nedströmspassage av fisk vid vattenanläggningar. Underlag till vägledning om lämpliga försiktighetsmått och bästa möjliga teknik för vattenkraft. Havs-och vattenmyndighetens rapport 14:114, Havs- och vattenmyndigheten (HaV): Göteborg, Sweden. [In Swedish with an English summary].
Calles, O., Karlsson, S., Vezza, P., Comoglio, C., and Tielman, J. (2013b). Success of a low‐sloping rack for improving downstream passage of silver eels at a hydroelectric plant. Freshwater Biology 58, 2168–2179.
| Success of a low‐sloping rack for improving downstream passage of silver eels at a hydroelectric plant.Crossref | GoogleScholarGoogle Scholar |
Calles, O., Rivinoja, P., and Greenberg, L. (2013c). A historical perspective on downstream passage at hydroelectric plants in Swedish rivers. In ‘Ecohydraulics: an Integrated Approach’. (Eds I. Maddock, A. Harby, P. Kemp, and P. Wood.) pp. 309–322. (Wiley: Chichester, UK.)
Castro-Santos, T. (2012). Adaptive fishway design: a framework and rationale for effective evaluations. In ‘Monitoring, Funktionskontrollen und Qualitätssicherung an Fischaufstiegsanlagen’. pp. 76–90. (Bundesanstalt für Gewässerkunde: Koblenz, Germany) Available at https://henry.baw.de/handle/20.500.11970/102404 [Verified 17 August 2018].
Castro-Santos, T., and Haro, A. (2003). Quantifying migratory delay: a new application of survival analysis methods. Canadian Journal of Fisheries and Aquatic Sciences 60, 986–996.
| Quantifying migratory delay: a new application of survival analysis methods.Crossref | GoogleScholarGoogle Scholar |
Castro-Santos, T., and Perry, R. (2012). Time-to-event analysis as a framework for quantifying fish passage performance. In ‘Telemetry Techniques: a User Guide for Fisheries Research’. (Eds N. S. Adams, J. W. Beeman, and J. H. Eiler.) pp. 427–452. (America Fisheries Society: Bethesda, MD, USA.)
Colotelo, A. H., Jones, B. W., Harnish, R. A., McMichael, G. A., Ham, K. D., Deng, Z. D., Squeochs, G. M., Brown, R. S., Weiland, M. A., Ploskey, G. R., Li, X., and Fu, T. (2012). Steelhead kelt passage distributions and Federal Columbia River Power System survival for fish tagged above and at Lower Granite Dam draft final report. Batelle, Pacific Northwest Division, Richland, WA, USA.
Coutant, C. C., and Whitney, R. R. (2000). Fish behavior in relation to passage through hydropower turbines: a review. Transactions of the American Fisheries Society 129, 351–380.
| Fish behavior in relation to passage through hydropower turbines: a review.Crossref | GoogleScholarGoogle Scholar |
Dingle, H. (2006). Animal migration: is there a common migratory syndrome? Journal of Ornithology 147, 212–220.
| Animal migration: is there a common migratory syndrome?Crossref | GoogleScholarGoogle Scholar |
DWA (2005). ‘Fish Protection Technologies and Downstream Fishways. Dimensioning, Design, Effectiveness Inspection.’ (German Association for Water, Wastewater and Waste (DWA): Hennef, Germany).
Ebel, G. (2013). ‘Fischschutz und Fischabstieg an Wasserkraftanlagen. Handbuch Rechen-und Bypasssysteme. Ingenieurbiologische Grundlagen, Modellierung und Prognose, Bemessung und Gestaltung.’ (Büro für Gewässerökologie und Fischereibiologie: Halle (Saale), Germany.)
EPRI (2001). Evaluation of angled bar racks and louvers for guiding fish at water intakes (1005193). EPRI, Palo Alto, CA, USA, and Dominion Millstone Laboratories: Waterford, CT, USA.
Ferguson, J. W. (2005). The behavior and ecology of downstream migrating Atlantic salmon (Salmo salar. L.) and brown trout (Salmo trutta L.) in regulated rivers in northern Sweden. Report 44, Vattenbruksinstitutionen, Umeå, Sweden.
Ferguson, J. W., Absolon, R. F., Carlson, T. J., and Sandford, B. P. (2006). Evidence of delayed mortality on juvenile Pacific salmon passing through turbines at Columbia River dams. Transactions of the American Fisheries Society 135, 139–150.
| Evidence of delayed mortality on juvenile Pacific salmon passing through turbines at Columbia River dams.Crossref | GoogleScholarGoogle Scholar |
Fox, J. (2002). Cox proportional-hazards regression for survival data. In ‘An R and S-PLUS Companion to Applied Regression’. pp. 1–18. (Sage: London, UK.)
Gosset, C., Travade, F., Durif, C., Rives, J., and Elie, P. (2005). Tests of two types of bypass for downstream migration of eels at a small hydroelectric power plant. River Research and Applications 21, 1095–1105.
| Tests of two types of bypass for downstream migration of eels at a small hydroelectric power plant.Crossref | GoogleScholarGoogle Scholar |
Harza Engineering Company and RMC Environmental Services Inc (1992). Response of Atlantic salmon smolts to louvers in the Holyoke Canal, Spring 1992. New England Power Company.
Harza Engineering Company and RMC Environmental Services Inc (1993). Response of juvenile clupeids to louvers in the Holyoke Canal, Fall 1992. New England Power Company.
Havn, T. B., Økland, F., Teichert, M. A., Heermann, L., Borcherding, J., Sæther, S. A., Tambets, M., Diserud, O. H., and Thorstad, E. B. (2017). Movements of dead fish in rivers. Animal Biotelemetry 5, 7.
| Movements of dead fish in rivers.Crossref | GoogleScholarGoogle Scholar |
Havn, T. B., Thorstad, E. B., Teichert, M. A. K., Sther, S. A., Heermann, L., Hedger, R. D., Tambets, M., Diserud, O. H., Borcherding, J., and Okland, F. (2018). Hydropower-related mortality and behaviour of Atlantic salmon smolts in the River Sieg, a German tributary to the Rhine. Hydrobiologia 805, 273–290.
| Hydropower-related mortality and behaviour of Atlantic salmon smolts in the River Sieg, a German tributary to the Rhine.Crossref | GoogleScholarGoogle Scholar |
Hebrand, M. (2012). Falkenbergs kommun. Hertings kraftverk, Ätran. Återställning av vandringsväg för fisk. Teknisk beskrivning. Rivning och nybyggnad av damm, återställning av åfåra, kontrollstationer samt fiskgaller. Fiskevårdsteknik AB, Lund, Sweden.
Heiß, M. (2015) Evaluation of innovative rehabilitation measures targeting downstream migrating Atlantic salmon smolt (Salmo salar) at a hydroelectric power plant in southern Sweden. M.Sc. Thesis, Ludwig-Maximilians-Universität, München, Germany.
Hesthagen, T., and Garnås, E. (1986). Migration of Atlantic salmon smolts in River Orkla of central Norway in relation to management of a hydroelectric station. North American Journal of Fisheries Management 6, 376–382.
| Migration of Atlantic salmon smolts in River Orkla of central Norway in relation to management of a hydroelectric station.Crossref | GoogleScholarGoogle Scholar |
Hosmer, D. W., Lemeshow, S., and May, S. (2008). ‘Applied Survival Analysis: Regression Modeling of Time-to-Event Data’, 2nd edn. (Wiley: Hoboken, NJ, USA.)
Jepsen, N., Koed, A., Thorstad, E. B., and Baras, E. (2002). Surgical implantation of telemetry transmitters in fish: how much have we learned? Hydrobiologia 483, 239–248.
| Surgical implantation of telemetry transmitters in fish: how much have we learned?Crossref | GoogleScholarGoogle Scholar |
Johnson, G. E., and Dauble, D. D. (2006). Surface flow outlets to protect juvenile salmonids passing through hydropower dams. Reviews in Fisheries Science 14, 213–244.
| Surface flow outlets to protect juvenile salmonids passing through hydropower dams.Crossref | GoogleScholarGoogle Scholar |
Jonsson, B., and Jonsson, N. (2011). ‘Ecology of Atlantic Salmon and Brown Trout: Habitat as a Template for Life Histories.’ (Springer.)
Jonsson, B., Waples, R., and Friedland, K. (1999). Extinction considerations for diadromous fishes. ICES Journal of Marine Science: Journal du Conseil 56, 405–409.
| Extinction considerations for diadromous fishes.Crossref | GoogleScholarGoogle Scholar |
Karchesky, C. M., Hanks, M. E., and McDonald, R. D. (2008). Annual report: T.W. Sullivan Powerhouse Evaluation of Bypass System Fish Guidance Efficiency (FGE). Normandeau Associates, Bedford, NH, USA.
Katopodis, C., and Williams, J. G. (2012). The development of fish passage research in a historical context. Ecological Engineering 48, 8–18.
| The development of fish passage research in a historical context.Crossref | GoogleScholarGoogle Scholar |
Kemp, P. S., and Williams, J. G. (2008). Response of migrating Chinook salmon (Oncorhynchus tshawytscha) smolts to in‐stream structure associated with culverts. River Research and Applications 24, 571–579.
| Response of migrating Chinook salmon (Oncorhynchus tshawytscha) smolts to in‐stream structure associated with culverts.Crossref | GoogleScholarGoogle Scholar |
Larsen, M. H., Johnsson, J. I., Näslund, J., Thomassen, S. T., and Aarestrup, K. (2016). Reduced rearing density increases postrelease migration success of Atlantic salmon (Salmo salar) smolts. Canadian Journal of Fisheries and Aquatic Sciences 73, 804–810.
| Reduced rearing density increases postrelease migration success of Atlantic salmon (Salmo salar) smolts.Crossref | GoogleScholarGoogle Scholar |
Liedtke, T. L., and Rub, A. M. W. (2012). Techniques for telemetry transmitter attachment and evaluation of transmitter effects on fish performance. In ‘Telemetry Techniques: a User Guide for Fisheries Research’. (Eds N. S. Adams, J. W. Beeman, and J. H. Eiler.) pp. 48–87. (American Fisheries Society: Bethesda, MD, USA.)
Lucas, M. C., Baras, E., Thom, T. J., Duncan, A., and Slavík, O. (2001). ‘Migration of Freshwater Fishes.’ (Blackwell Science, Malden, MA, USA.)
Marmulla, G. (2001). ‘Dams, Fish and Fisheries: Opportunities, Challenges and Conflict Resolution.’ (Food & Agriculture Organisation: Rome, Italy.)
McCormick, S. D., Hansen, L. P., Quinn, T. P., and Saunders, R. L. (1998). Movement, migration, and smolting of Atlantic salmon (Salmo salar). Canadian Journal of Fisheries and Aquatic Sciences 55, 77–92.
| Movement, migration, and smolting of Atlantic salmon (Salmo salar).Crossref | GoogleScholarGoogle Scholar |
McCormick, S. D., Cunjak, R. A., Dempson, B., O’Dea, M. F., and Carey, J. B. (1999). Temperature-related loss of smolt characteristics in Atlantic salmon (Salmo salar) in the wild. Canadian Journal of Fisheries and Aquatic Sciences 56, 1649–1667.
| Temperature-related loss of smolt characteristics in Atlantic salmon (Salmo salar) in the wild.Crossref | GoogleScholarGoogle Scholar |
Montgomery, D. R. (2004). ‘King of Fish: the Thousand-year Run of Salmon.’ (Basic Books: Cambridge, MA, USA.)
Muir, W. D., Smith, S. G., Williams, J. G., and Sandford, B. P. (2001). Survival of juvenile salmonids passing through bypass systems, turbines, and spillways with and without flow deflectors at Snake River dams. North American Journal of Fisheries Management 21, 135–146.
| Survival of juvenile salmonids passing through bypass systems, turbines, and spillways with and without flow deflectors at Snake River dams.Crossref | GoogleScholarGoogle Scholar |
Muir, W. D., Marsh, D. M., Sandford, B. P., Smith, S. G., and Williams, J. G. (2006). Post-hydropower system delayed mortality of transported Snake River stream-type Chinook salmon: unraveling the mystery. Transactions of the American Fisheries Society 135, 1523–1534.
| Post-hydropower system delayed mortality of transported Snake River stream-type Chinook salmon: unraveling the mystery.Crossref | GoogleScholarGoogle Scholar |
Nettles, D., and Gloss, S. (1987). Migration of landlocked Atlantic salmon smolts and effectiveness of a fish bypass structure at a small-scale hydroelectric facility. North American Journal of Fisheries Management 7, 562–568.
| Migration of landlocked Atlantic salmon smolts and effectiveness of a fish bypass structure at a small-scale hydroelectric facility.Crossref | GoogleScholarGoogle Scholar |
Nieminen, E., Hyytiäinen, K., and Lindroos, M. (2017). Economic and policy considerations regarding hydropower and migratory fish. Fish and Fisheries 18, 54–78.
| Economic and policy considerations regarding hydropower and migratory fish.Crossref | GoogleScholarGoogle Scholar |
Noonan, M. J., Grant, J. W., and Jackson, C. D. (2012). A quantitative assessment of fish passage efficiency. Fish and Fisheries 13, 450–464.
| A quantitative assessment of fish passage efficiency.Crossref | GoogleScholarGoogle Scholar |
Northcote, T. (1998). Migratory behaviour of fish and its significance to movement through riverine fish passage facilities. In ‘Migration and Fish Bypasses’. (Eds M. Jungwirth, S. Schmutz, and S. Weiss.) pp. 3–18. (Fishing News Books: Oxford, UK.)
Nyqvist, D., Nilsson, P. A., Alenäs, I., Elghagen, J., Hebrand, M., Karlsson, S., Kläppe, S., and Calles, O. (2017a). Upstream and downstream passage of migrating adult Atlantic salmon: remedial measures improve passage performance at a hydropower dam. Ecological Engineering 102, 331–343.
| Upstream and downstream passage of migrating adult Atlantic salmon: remedial measures improve passage performance at a hydropower dam.Crossref | GoogleScholarGoogle Scholar |
Nyqvist, D., Greenberg, L. A., Goerig, E., Calles, O., Bergman, E., Ardren, W. R., and Castro‐Santos, T. (2017b). Migratory delay leads to reduced passage success of Atlantic salmon smolts at a hydroelectric dam. Ecology Freshwater Fish 26, 707–718.
| Migratory delay leads to reduced passage success of Atlantic salmon smolts at a hydroelectric dam.Crossref | GoogleScholarGoogle Scholar |
Nyqvist, D., McCormick, S. D., Greenberg, L., Ardren, W. R., Bergman, E., Calles, O., and Castro-Santos, T. (2017c). Downstream migration and multiple dam passage by Atlantic salmon smolts. North American Journal of Fisheries Management 37, 816–828.
| Downstream migration and multiple dam passage by Atlantic salmon smolts.Crossref | GoogleScholarGoogle Scholar |
Nyqvist, D., Bergman, E., Calles, O., and Greenberg, L. (2017d). Intake approach and dam passage by downstream‐migrating Atlantic salmon kelts. River Research and Applications 33, 697–706.
| Intake approach and dam passage by downstream‐migrating Atlantic salmon kelts.Crossref | GoogleScholarGoogle Scholar |
Olofsson, H. (2013). ‘Recipientkontrollen i Ätran 2012.’ (Ätrans Vattenråd & ALcontrol AB: Linköping, Sweden.)
Peake, S., and McKinley, R. (1998). A re-evaluation of swimming performance in juvenile salmonids relative to downstream migration. Canadian Journal of Fisheries and Aquatic Sciences 55, 682–687.
| A re-evaluation of swimming performance in juvenile salmonids relative to downstream migration.Crossref | GoogleScholarGoogle Scholar |
Richards, S. A. (2008). Dealing with overdispersed count data in applied ecology. Journal of Applied Ecology 45, 218–227.
| Dealing with overdispersed count data in applied ecology.Crossref | GoogleScholarGoogle Scholar |
Roscoe, D. W., and Hinch, S. G. (2010). Effectiveness monitoring of fish passage facilities: historical trends, geographic patterns and future directions. Fish and Fisheries 11, 12–33.
| Effectiveness monitoring of fish passage facilities: historical trends, geographic patterns and future directions.Crossref | GoogleScholarGoogle Scholar |
Schwinn, M., Baktoft, H., Aarestrup, K., and Koed, A. (2017). A comparison of the survival and migration of wild and F1-hatchery-reared brown trout (Salmo trutta) smolts traversing an artificial lake. Fisheries Research 196, 47–55.
| A comparison of the survival and migration of wild and F1-hatchery-reared brown trout (Salmo trutta) smolts traversing an artificial lake.Crossref | GoogleScholarGoogle Scholar |
Scruton, D., McKinley, R., Kouwen, N., Eddy, W., and Booth, R. (2002). Use of telemetry and hydraulic modeling to evaluate and improve fish guidance efficiency at a louver and bypass system for downstream-migrating Atlantic salmon (Salmo salar) smolts and kelts. Hydrobiologia 483, 83–94.
| Use of telemetry and hydraulic modeling to evaluate and improve fish guidance efficiency at a louver and bypass system for downstream-migrating Atlantic salmon (Salmo salar) smolts and kelts.Crossref | GoogleScholarGoogle Scholar |
Scruton, D., McKinley, R., Kouwen, N., Eddy, W., and Booth, R. (2003). Improvement and optimization of fish guidance efficiency (FGE) at a behavioural fish protection system for downstream migrating Atlantic salmon (Salmo salar) smolts. River Research and Applications 19, 605–617.
| Improvement and optimization of fish guidance efficiency (FGE) at a behavioural fish protection system for downstream migrating Atlantic salmon (Salmo salar) smolts.Crossref | GoogleScholarGoogle Scholar |
Scruton, D. A., Pennell, C. J., Robertson, M. J., Clarke, K. D., Eddy, W., and McKinley, R. S. (2004). Telemetry studies of the passage route and entrainment of downstream migrating wild Atlantic salmon (Salmo salar) smolts at two hydroelectric installations on the Exploits River, Newfoundland, Canada. In ‘Aquatic Telemetry: Advances and Applications’. (Eds M. T. Spedicato, G. Lembo, and G. Marmulla.) pp. 91–101. (FAO/COISPA: Rome, Italy.)
Scruton, D., Pennell, C., Bourgeois, C., Goosney, R., Porter, T., and Clarke, K. (2007). Assessment of a retrofitted downstream fish bypass system for wild Atlantic salmon (Salmo salar) smolts and kelts at a hydroelectric facility on the Exploits River, Newfoundland, Canada. Hydrobiologia 582, 155–169.
| Assessment of a retrofitted downstream fish bypass system for wild Atlantic salmon (Salmo salar) smolts and kelts at a hydroelectric facility on the Exploits River, Newfoundland, Canada.Crossref | GoogleScholarGoogle Scholar |
Scruton, D., Pennell, C., Bourgeois, C., Goosney, R., King, L., Booth, R., Eddy, W., Porter, T., Ollerhead, L., and Clarke, K. (2008). Hydroelectricity and fish: a synopsis of comprehensive studies of upstream and downstream passage of anadromous wild Atlantic salmon, Salmo salar, on the Exploits River, Canada. Hydrobiologia 609, 225–239.
| Hydroelectricity and fish: a synopsis of comprehensive studies of upstream and downstream passage of anadromous wild Atlantic salmon, Salmo salar, on the Exploits River, Canada.Crossref | GoogleScholarGoogle Scholar |
Serrano, I., Rivinoja, P., Karlsson, L., and Larsson, S. (2009). Riverine and early marine survival of stocked salmon smolts, Salmo salar L., descending the Testebo River, Sweden. Fisheries Management and Ecology 16, 386–394.
| Riverine and early marine survival of stocked salmon smolts, Salmo salar L., descending the Testebo River, Sweden.Crossref | GoogleScholarGoogle Scholar |
Spicer, A. V., Moring, J. R., and Trial, J. G. (1995). Downstream migratory behavior of hatchery-reared, radio-tagged Atlantic salmon (Salmo salar) smolts in the Penobscot River, Maine, USA. Fisheries Research 23, 255–266.
| Downstream migratory behavior of hatchery-reared, radio-tagged Atlantic salmon (Salmo salar) smolts in the Penobscot River, Maine, USA.Crossref | GoogleScholarGoogle Scholar |
Stira, R. J., and Robinson, D. A. (1997). Effectiveness of a louver bypass system for downstream passage of Atlantic salmon smolts and juvenile clupeids in the Holyoke Canal, Connecticut River, Holyoke, Massachusetts. In ‘EPRI Fish Passage Workshop’, 6–8 May 1997, Milwaukee, WI, USA. (Electric Power Research Institute: Palo Alto, CA, USA.)
Tanguy, J., Ombredane, D., Bagliniere, J., and Prunet, P. (1994). Aspects of parr-smolt transformation in anadromous and resident forms of brown trout (Salmo trutta) in comparison with Atlantic salmon (Salmo salar). Aquaculture 121, 51–63.
| Aspects of parr-smolt transformation in anadromous and resident forms of brown trout (Salmo trutta) in comparison with Atlantic salmon (Salmo salar).Crossref | GoogleScholarGoogle Scholar |
Thorstad, E. B., Rikardsen, A. H., Alp, A., and Okland, F. (2013). The use of electronic tags in fish research: an overview of fish telemetry methods. Turkish Journal of Fisheries and Aquatic Sciences 13, 881–896.
Tomanova, S., Courret, D., and Alric, A. (2017). Protecting fish from entering turbines: the efficiency of a low-sloping rack for downstream migration of Atlantic salmon smolts. Houille Blanche 1, 11–13.
| Protecting fish from entering turbines: the efficiency of a low-sloping rack for downstream migration of Atlantic salmon smolts.Crossref | GoogleScholarGoogle Scholar |
Whitney, R. R., and Council, N. P. P. (1997). ‘Downstream Passage for Salmon at Hydroelectric Projects in the Columbia River Basin: Development, Installation, and Evaluation.’ (Northwest Power Planning Council: Portland, OR, USA.)
Wickham, H. (2016). ‘ggplot2: Elegant Graphics for Data Analysis.’ (Springer: New York, NY, USA.)