

Geostatistical tools to assess shifts in recreational fishing
Shannon J. Burchert A * , Glenn A. Hyndes A , Karina L. Ryan B and Ute Mueller A
A * , Glenn A. Hyndes A , Karina L. Ryan B and Ute Mueller A
A School of Science, Edith Cowan University, 270 Joondalup Drive, Joondalup, WA 6027, Australia.
B Department of Primary Industries and Regional Development, Northside Drive, Hillarys, WA 6025, Australia.
Marine and Freshwater Research 73(5) 588-604 https://doi.org/10.1071/MF21131
Submitted: 7 May 2021 Accepted: 13 December 2021 Published: 9 February 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
The use of geostatistical indices to examine fishery-dependent data over time is novel. We assess the value of a range of these indices for describing trends in boat-based recreational fishing in Western Australia and areas of high effort and catches for two demersal species with varying biological characteristics, life histories and distributions. The number of blocks (10- × 10-nautical mile (~19- × 19-km) area) visited decreased by 40%, while the number of fishing trips with demersal species catch increased by 15%. Spatial indices showed a south-easterly shift in demersal catch per unit effort (CPUE) across survey years. The southerly shift of West Australian dhufish (Glaucosoma herbraicum) CPUE and subsequent return to the north likely to reflect the response of this temperate species to a marine heatwave. In comparison, CPUE of retained Baldchin groper (Choerodon rubescens) shifted north, whereas released catches shifted south with high probabilities of hotspots present associated with habitat structure. We identified hotspots of retained and released CPUE, highlighting potential areas where species experience high post-release mortality through barotrauma, and targeted spatial management strategies. Spatial indices allow quantitative definition of the structure of the fishery, identify high catch areas, and assess changes over time at a broader scale to sustainably manage multispecies fisheries.
Keywords: angling, catch rate, centre of gravity, fisheries management, fishery-dependent data, geostatistics, hotspot, index of collocation.
Introduction
Fishery-dependent data are useful for the monitoring and management of a multi-sector fishery, where resource utilisation is based on stock assessments and sustainability modelling (McPhee et al. 2002; Griffiths and Fay 2015). Catch rate estimates, or catch per unit effort (CPUE), from commercial, charter and private boat-based recreational fishing have been used to determine the status of a fishery, evaluate historical trends and, in some cases, estimate population abundance (Morgan and Burgess 2004; Gaughan and Santoro 2020). Whereas these types of stock assessments are vital for the sustainability of the fishery resource, understanding the spatial and temporal structure of the stock is invaluable because CPUE changes through time and space in response to various environmental and anthropogenic factors (Perry et al. 2005; Booth et al. 2011; van Putten et al. 2017; Humphries et al. 2019).
Fishing and its impacts are known to change spatially and temporally (Petitgas et al. 2003; Kleisner et al. 2010); however, this has been documented mainly for commercial fisheries. Spatial modelling approaches using recreational fishing data were initially investigated in 2010 (Parnell et al. 2010); however, spatio-temporal modelling has been limited, with few studies exploring survey data (Tao et al. 2012; Aidoo et al. 2015, 2016; Winfield 2016; Petitgas et al. 2018; Polansky et al. 2018; Navarro et al. 2020). Understanding these spatial and temporal shifts is important because sharing of the stock and fishing grounds in a multi-sector fishery could potentially cause a particular species or area to be overexploited. Also, species within multi-species fisheries are often managed collectively; however, they have unique biological characteristics, life histories and geographic distributions (Newman et al. 2018). Characterising locations for catch, effort and CPUE not only provides the ability to assess trends and patterns within key fisheries at a resource level but also provides insights into fisher behaviour in terms of CPUE hotspots and exploitation of bag and boat limits in a multi-species fishery.
The identification of areas of high concentration of effort or CPUE, termed hotspots, is fundamental for the development of management strategies in terms of resource conservation (Petitgas et al. 2016). Spatial management approaches, such as the introduction of marine parks or ‘no-take’ zones, are becoming a more viable option to reduce targeted effort and, consequently, catch (Lynch 2006, 2014; Smallwood and Beckley 2012; Lynch et al. 2020). The concept of spatial predictability of hotspots has been applied to biodiversity indices, trophic interaction areas, fish distributions and single species distributions by using long-term data (Myers et al. 2000; Santora et al. 2011; Stuart-Smith et al. 2013; Yasuda et al. 2014; Petitgas et al. 2016). Monitoring and management of spatially complex resources are primarily undertaken using commercial catch data (fishery-dependent) and fishery-independent surveys of key species to inform policies and implement restrictions such as size at capture, gear type, effort, and seasonal and spatial closures. The use of spatial information from private boat-based recreational fishing (not including charter) is novel in terms of adoption into management, because these large datasets are collected infrequently and are limited in terms of catches, scope, and temporal and spatial scale (Henry and Lyle 2003; Roa-Ureta and Niklitschek 2007; Hartill et al. 2016; Santos et al. 2019). Although spatial information is captured within these fishery-dependent surveys, these data have not been fully explored with respect to developing an understanding of the recreational fishery in space through time, by using geostatistical techniques (Woillez et al. 2005; Santos et al. 2019). Techniques include co-kriging hotspots and various descriptive spatial indices such as the centre of gravity (CG) which is the mean location weighted by the variable of interest, inertia (I) which is the square distance between data locations and the CG, but also global and local index of collocation, positive area and equivalent area.
In the past two decades, spatial indices have been explored with fishery-independent trawl and dredge data, and also acoustic survey data to demonstrate their ability to capture and detect changes in spatial distributions of various marine species (Petitgas 2001; Woillez et al. 2005; Petitgas et al. 2017, 2018). Spatial indexes such as CG and I have been applied in many studies to assess the mean location of a population and the dispersion of that population around its CG across a time series (Woillez et al. 2005; Mueller et al. 2012; Petitgas et al. 2017; Currie et al. 2019). The global and local indexes of collocation, which are not as commonly applied in marine science, are used to measure the spatial overlap of two populations, and the similarity of the populations at locations in two given periods (Kulka et al. 2003; Aidoo 2016).
Adapting these techniques to specialised areas of fisheries science has the potential to inform resource management in terms of fine-scale trends relative to catch and CPUE, so as to determine areas of high activity. These approaches also form valuable tools in examining shifts using fishery-dependent data. This study aims to assess the utility of geostatistical spatial indices to describe private boat-based recreational fishing in terms of temporal shifts and areas of high fishing activity, to inform fisheries management. To achieve this, multiple-years of phone-diary surveys on recreational boat-based catches (retained and released) of two key Western Australian demersal species will be used. These two species were selected because they form a substantial proportion of catch for the Western Australian demersal fishery, have different life-history characteristics and distributions in a multi-species fishery, which will allow the methodological approach to be generalised for a wider application. Retained and released catches were analysed separately to determine general compliance levels of bag and boat limit regulations, and to identify areas of high release rates, which can result in post-release mortality (Cooke and Schramm 2007; Post and Parkinson 2012). High release areas with targeted effort are potential candidates for spatial closures, where spatial indices and hotspot mapping can provide managers with additional information to introduce relevant policies.
Materials and methods
Study area
The study area follows the western coast of Australia from 27°S, 114°E to 34°S, 115°30ʹE (Supplementary Fig. S1). Our focus is on boat-based recreational fishing in the West Coast Bioregion, a management region for fisheries in Western Australia. This study focuses on the demersal scalefish species that reside within 20–250-m water depth (Department of Primary Industries and Regional Development 2021). Boat-based recreational fishing (which requires a licence) was considered for analysis where fishers target, retain or release a demersal species during a given survey period. The analyses focused on the combined demersal species that are targeted and caught by recreational fishers, as well as two key species (West Australian dhufish, Glaucosoma herbraicum; and Baldchin groper, Choerodon rubescens) within that group of fishes. These two species are long-lived, endemic to Western Australia, have a recovering fisheries status and are targeted across multiple sectors (commercial, charter and recreational); however, they have varying life histories, unique biological characteristics and geographic distributions. West Australian dhufish is gonochoristic, has sex-dependent growth rates that also show geographic differences and is a more temperate species, where its distribution is widespread within the study region (Smallwood et al. 2013). Baldchin groper is protogynous, and its catches are concentrated in the mid-west of the West Coast Bioregion (Hutchins 2001). These commonalities and differences make them ideal candidates to explore spatial and temporal statistical methods. For the recreational sector, management of these species is currently through boat, bag and size limits at a bioregional scale, and a 2-month seasonal closure to reduce catch. In addition to this, Baldchin groper at the Abrolhos Islands is currently protected during its spawning period, to provide additional protection from exploitation (Newman et al. 2018; Gaughan and Santoro 2020).
Survey data
Data used in this study were collected as part of the state-wide recreational fishing surveys conducted on a biennial basis (March 2011–February 2012, May 2013–April 2014, September 2015–August 2016 and September 2017–August 2018; Ryan et al. 2019). All components of these surveys were completed by recreational boat fishing licence holders. The phone-diary survey data consist of deidentified numeric and categorical response variables in relation to fishing events (records) for individual licence holders randomly selected from the recreational boat fishing licence database for each survey period. In the most recent survey year (2017–18), 2931 recreational boat fishing licence holders participated, with response rates being consistent across survey years (Ryan et al. 2019). The phone-diary survey data are recorded on an event basis across the designated year for each survey period. A fishing event is defined as an attempt to catch a fish species, irrespective of success, using a line from a boat at any one point in time. For this study, fishing records were aggregated by block for individual survey years. Variables used for spatial analyses include fishing duration (time spent fishing in hours), the number of fishing records, retained and released catch numbers (survey data), and retained CPUE and released CPUE for each block across species of interest. Positive area and equivalent area were calculated with catch data because these spatial indices are used to describe and summarise species catch distributions in terms of population density for a given area.
For each 10- × 10-nautical mile (~19- × 19-km) block b, CPUE (Řb retained or released) was calculated using the ratio of means estimator (Hoenig et al. 1997), where Nb denotes the number of events in block b, ci,b retained or released catch for event i in block b, fishing effort hi,b = ti,b × ni,b, with ti,b time (h) and ni,b number of licence and non-licence holders whom participated in the fishing activity. Because non-licence holders on board with a licence holder are able to contribute to the bag limit for that licence holder, and therefore putting effort into the fishery, the total number of fishers was used to calculate effort, as follows.

For this study, all blocks or fishing records in marine waters where demersal species were retained or released were considered. Data for non-demersal fishing locations, predominantly 50 nautical miles (~93 km) from the coastline and inland river systems, were excluded (8.4% of records in 2011–12, 7.3% in 2013–14, 3.8% in 2015–16 and 3.2% in 2017–18).
Geostatistical spatial indices
This study applied geostatistical spatial indices described within the ICES Cooperative Research Report (Petitgas et al. 2017) to capture the spatial structure of boat-based recreational fishing and they are listed in Supplementary Table S1, with references to their application. Indices, including total area (TA) in square nautical miles, total records, total catch, percentage of records with catches >0 (% positive records), percentage of records with catches = 0 (% zeroes), exploitation (Exp%) or the percentage of records for a given survey year with catches equating to or exceeding the mixed demersal species boat limit, overall CPUE inertia (I, nautical miles2), CPUE inertia in the north–south (N–S) direction (nautical miles2), CPUE inertia in the east–west (E–W) direction (nautical miles2), positive area (PA, nautical miles2) covered by records with catches >0 using catch data, and equivalent area (EA, nautical miles2) using catch data, are used to describe spatial patterns within a given survey year to characterise boat-based recreational fishing targeting or catching demersal finfish species in the study region (Table 1). Polygons were created on the basis of the spatial extent of fishing in each survey year, delineating the maximal sampled area. The latitudinal range of data extends to ~450 nautical miles (~833 km), where the longitudinal range varied from 10 to 50 nautical miles (~19–93 km).

|
Centre of gravity and associated inertia
Spatial shifts were identified within the study region across survey years by calculating a series of spatial indices including the centre of gravity (CG), associated inertia (I) and inertia in the north–south and east–west directions for a range of variables including, retained CPUE, released CPUE, number of records and mean effort. The CG = (xCG, yCG) represents the mean location of aggregated data (zi) weighted by the variable of interest (Woillez et al. 2005) and I describes the mean square distance (nautical miles) between the block locations ui = (xi, yi), i = 1,…N (as measured by the coordinates of the block centres) and the CG weighted by the variable selected.
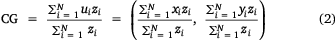
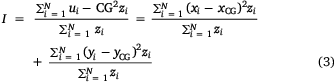
To test whether the elongated shape of the study region influenced the various CGs, the CG was calculated for locations weighted with a value of 1, which showed no shape effect on the centres of gravity for the variables of interest.
Global and local index of collocation
To determine whether data are geographically distinct across survey years, the global and local indices of collocation were both calculated for variables including retained CPUE, released CPUE, number of records and mean effort (Woillez et al. 2005; Petitgas et al. 2017). The global index of collocation (GIC) measures the overlap of regions with records for different survey years by comparing the distance between the CG from 2 years (ΔCG) and mean distance between block locations (I1 and I2). The local index of collocation (LIC) then provides a measure of the similarity of the selected variables z1 and z2 at locations where records are available in both survey years. Owing to the nature of boat-based recreational fishing data, NA values are present in the data because some block locations were not visited in each survey year.
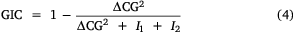
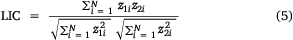
Positive area
The total area (nautical miles2) of the study region with a catch record (zi) greater than zero is referred to as positive area (PA). Here, it was calculated separately for West Australian dhufish, Baldchin groper and the demersal species as the sum of the areas of all blocks, where a fisher had retained or released at least one fish, as follows:
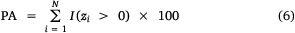
where I(zi > 0) = 1 if (zi > 0) and 0 otherwise and 100 (nautical miles2) is the area of a fishing block.
Equivalent area
To determine the area that would be covered if retained or released catch from boat-based recreational fishing was constant, the equivalent area (EA, nautical miles2) was calculated as
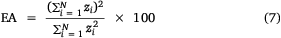
Boat limit
To determine the level of exploitation, the number of recreational fishers and retained catches for individual records were assessed. For each individual record, there are management restrictions on what can be retained from the demersal fishery to avoid exploitation of resources. Bag limits for the mixed demersal fishery in the West Coast Bioregion are restricted to two fish per licence holder, with no maximum number of retained fish per vessel. The percentage of individual fishing records per 10- × 10-nautical mile (~19- × 19-km) block with retained catches zi ≥ bag limit × number of licence holders was calculated and aggregated across survey years for the West Coast Bioregion.
Hotspot definition
A hotspot was defined as a region where retained CPUE and released at block locations within the study area were above a cut-off value and spatially uncorrelated with the remainder of the distribution below the cut-off value (Petitgas et al. 2016). A sequence of cut-off values were chosen on the basis of the distribution of the retained CPUE and released CPUE for each species across survey years increasing in value 0.001 < 0.02 < 0.08 < 0.20 < 0.40 < 0.80 and were then coded as ‘hard’ indicator values.
If Ai represents the set of all block locations u with Z(u) ≥ zi, where zi is a cut-off value, then the indicator function associated with it can be defined as
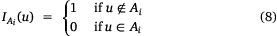
The indicator variogram is then defined as
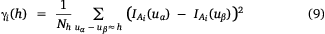
where uα and uβ represent block locations separated by a distance h. The indicator variograms γi(h) measures the likelihood that a vector of length h has one extremity inside and one outside Ai.
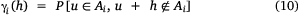
The indicator cross-variogram is then defined as
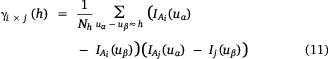
where Aj and Ai are the sets associated with cut-offs zj and zi respectively. It can be interpreted as a measure of the likelihood that a vector of length h has one extremity inside Aj and one outside Ai (Petitgas et al. 2016).
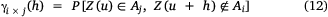
Consequently, the quotient of γi × j (h; zi) and γi (h; zi; Eqn 13) then measures the probability of being inside of Aj when entering Ai from separation distance (h; Petitgas et al. 2016).
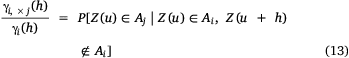
When the likelihood of moving from Ai to Aj at a separation distance of h is no longer dependent on h, suggesting that higher values are spatially uncorrelated (variogram showing pure nugget, which means that variance does not depend on distance; Supplementary Fig. S7, S8) and therefore cannot be predicted within Ai, zi represents the ‘top’ cut-off value and Ai represents the hotspot (Petitgas et al. 2016).
Indicator variograms were fitted using the ‘model.auto( )’ function implemented in the RGeostats package (ver. 12.0.0, MINES ParisTech & ARMINES, see http://rgeostats.free.fr/) with two structured spherical models and a nugget effect. In total, 15 lags were used at a distance of 10 nautical miles (~19 km). A cut-off was sufficiently high when the associated variogram showed a loss in structure, i.e. it could be modelled as pure nugget.
Co-kriging hotspots
To produce probability maps of the retained and released CPUE hotspots occurring, indicators 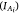 were co-kriged (multivariate form of kriging, Petitgas et al. 2016). The experimental indicator and cross-variograms were calculated using 15 lags at a distance of 10 nautical miles (~19 km). A linear model of co-regionalisation with two structured spherical models and a nugget effect was then fitted. For co-kriging, a moving neighbourhood was used with a minimum of four and a maximum of 20 samples at a radius of 120 nautical miles (~222 km). To allow for co-kriging among survey years, indicators of hotspots were selected at the same sample locations and interpolated on a 5- × 5-nautical mile (~9- × 9-km) grid (Bivand et al. 2008; Petitgas et al. 2017).
were co-kriged (multivariate form of kriging, Petitgas et al. 2016). The experimental indicator and cross-variograms were calculated using 15 lags at a distance of 10 nautical miles (~19 km). A linear model of co-regionalisation with two structured spherical models and a nugget effect was then fitted. For co-kriging, a moving neighbourhood was used with a minimum of four and a maximum of 20 samples at a radius of 120 nautical miles (~222 km). To allow for co-kriging among survey years, indicators of hotspots were selected at the same sample locations and interpolated on a 5- × 5-nautical mile (~9- × 9-km) grid (Bivand et al. 2008; Petitgas et al. 2017).
Software and assumptions
RStudio was the computing environment utilised for this study, with the package RGeostats to compute all spatial indices (ver. 12.0.0, MINES ParisTech & ARMINES, see http://rgeostats.free.fr/) and ggplot2 for producing the maps (ver. 3.3.5, H. Whickham, see https://ggplot2.tidyverse.org). Within the geostatistical modelling framework and throughout this study, there is an assumption that observations of CPUE are spatially autocorrelated, where space is characterised by patterns rather than randomness (Cressie 1991; Griffith 2007). This assumption follows Tobler’s first law of geography, where ‘everything is related to everything else, but near things are more related than distant things’ (Miller 2004).
Results
Maps of the records of fishers targeting demersal species showed a contraction of fishing events towards the coastline, with high recreational fishing effort (>200 records) being concentrated adjacent to population centres (e.g. adjacent to Perth and Geraldton; Fig. 1), and fishery access points being within the Perth metropolitan region. Similarly, fishing effort contracted towards the coast after 2011–12, with high effort being situated offshore rather than adjacent to population centres in 2011–12 and, subsequently, closer to the coast across most of the bioregion (Fig. 1). High effort was also observed adjacent to population centres and access points. Maps of retained CPUE for all demersal species (Fig. 2) showed high values at the Abrolhos Islands and Geraldton in survey years 2011–12, 2015–16 and 2017–18. Retained CPUE for all demersal species were highest south of Perth in 2013–14, off the coast of Bunbury in 2015–16 and between Perth and Geraldton in 2017–18. Likewise, released CPUE for all demersal species were highest at the Abrolhos Islands in all survey years, near Perth and Mandurah in 2011–12 and 2013–14, and near Bunbury in 2015–16 and 2017–18. Released CPUE of West Australian dhufish across survey years were often higher than were retained CPUE in several areas of the study region (Fig. 3). Released CPUE of West Australian dhufish was widely distributed across the West Coast Bioregion, with retained CPUE being high near Geraldton and Bunbury. Similarly, the released CPUE of Baldchin groper was often higher than was retained CPUE, with CPUE at the Abrolhos Islands being high (Fig. 4). Unlike West Australian dhufish, both retained and released CPUE of Baldchin groper showed similar localised distributions in the West Coast Bioregion.

|

|

|

|
The CG for the number of records, mean effort targeting demersal species (Fig. 1), retained CPUE (Fig. 2) and released CPUE (Fig. 2) all showed a southerly latitudinal shift from 2011–12 to 2013–14 and a slight northerly shift in subsequent years. A southerly shift in the CG was evident for retained and released CPUE of West Australian dhufish, where latitudinal shifts were similar in 2015–16 and 2017–18 (Fig. 3). However, Baldchin groper showed minor fluctuations among survey years (Fig. 4).
The total area (TA) with recorded demersal fishing activity decreased by 40% across the survey years (Table 1). Although there was a decrease in total area, the total number of records alongside total catch and the percentage of blocks with positive (non-zero) catch records for retained catches increased slightly across survey years (Table 2). The percentage of records with a positive released catch of all demersal species showed no trend, with values ranging between 91.3% in 2013–14 and 96.6% in 2015–16 (Table 1). Owing to the elongation of the study region, as described by the overall inertia across species and CPUE (Table 1), anisotropy was present where inertia along the two principal axes was greater in the N–S direction than in the E–W direction. Inertia in the N–S direction increased slightly across years for retained and released CPUE. On a species level, retained and released CPUE for West Australian dhufish showed an increase in overall inertia and inertia in N–S and E–W directions between 2011–12 and the remaining survey years. However, retained and released CPUE for Baldchin groper increased in overall inertia and, in the N–S and E–W directions for released CPUE, but not for retained CPUE.

|
Similarly to TA, positive area (PA) showed a contraction across survey years, where 2011–12 displayed a greater spread of retained and released catch for all demersal species than did other survey years (Table 1). Overall, the positive area for all demersal species has decreased across survey years, with the greatest change between 2011–12 and 2013–14 (Table 1). Positive area for retained and released CPUE of West Australian dhufish followed the same pattern as did those of all demersal species, except equivalent area for retained CPUE of West Australian dhufish, which increased overall. However, retained and released CPUE of Baldchin groper showed spatial patterns regarding equivalent area different from those of West Australian dhufish. The equivalent area for Baldchin groper increased across survey years for retained and released CPUE.
The global index of collocation (GIC) for effort shows stability across survey years for locations where fishing is occurring. The GIC highlights minor differences between 2011–12 and subsequent years for retained and released CPUE for all demersal species as well as West Australian dhufish and Baldchin groper (Table 2). The local index of collocation (LIC) showed some local variability for mean effort, where 2011–12 showed a difference from all other survey years (Table 2). The LIC for West Australian dhufish and Baldchin groper both indicated strong differences among years for released CPUE and moderate differences for retained CPUE (Table 2). Comparing retained against released CPUE, all demersal species showed minimal local variability across survey years. However, there were strong differences for retained and released CPUE of West Australian dhufish in 2013–14, and Baldchin groper in 2011–12 and 2017–18.
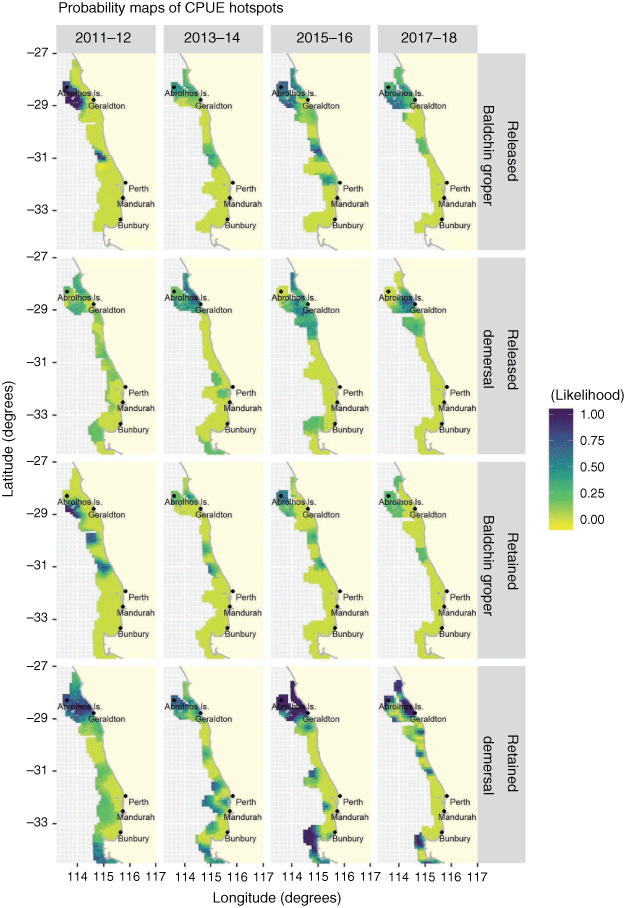
The appropriate cut-offs to define the likelihood of an area being a hotspot across species were selected on the basis of the variogram of the indicators (Supplementary Fig. S3–S8). Cut-off values were applied for all demersal species (retained CPUE = 0.08 and released CPUE = 0.20), West Australian dhufish (retained CPUE = 0.20 and released CPUE = 0.40), and Baldchin groper (retained CPUE = 0.20 and released CPUE = 0.08; Supplementary Table S3). The likelihood of a retained CPUE hotspot for all demersal species was high in the northern region at the Abrolhos Islands and in the southern region, south of Bunbury (Fig. 5). The likelihood of a hotspot for released CPUE of all demersal species was higher in the northern region, near Geraldton and the Abrolhos Islands. Variogram ratios for retained and released CPUE of West Australian dhufish (Supplementary Fig. S5, S6) showed no evidence of pure nugget, suggesting instability where no definitive hotspots were found in the study region. Probability maps of retained and released CPUE of Baldchin groper identified a high likelihood of a hotspot at the Abrolhos in 2011–12, which dissipated in consecutive years for retained CPUE, but remained present in 2015–16 and 2017–18 for released CPUE. When a cut-off of >0.5 was applied to the probability maps (Fig. 5), areas outlined in darker colours (purple) presented as hotspots.
|
Discussion
Geostatistical spatial indices (such as the CG, PA, GIC, LIC and EA), in conjunction with hotspot analysis, have successfully been used on fishery-dependent data from a boat-based recreational fishery to show shifts in effort and CPUE over time that may correspond to changes in management, socio-economic or environmental conditions. In previous studies, these spatial indices have been utilised to detect changes and patterns in spatial distributions of marine populations in terms of fish egg and larvae distributions in marine waters (Woillez et al. 2005; Woillez et al. 2016). However, they have not been applied to data in a management context such as recreational fishing data (Woillez et al. 2005). Recreational fishing data are often collected infrequently, zero-inflated with sporadic high catches, and comprise unsampled locations. Despite these issues, spatial indices were useful in characterising the boat-based recreational fishery in our study region through time and space and are likely to form a valuable set of tools to characterise other fisheries on the basis of fishery-dependent data.
The spatial footprint of the boat-based recreational fishers, on the basis of fishing events and effort, had a greater spread, and extended further into more offshore areas in 2011–12 than in any other subsequent survey year. This pattern was supported by the decreasing total area, positive area, equivalent area and the interannual variability of GIC and LIC. Such temporal variation in fishing events and effort potentially reflects shifts in fisher behaviour in response to changing abundances of target species and catch compositions within the area as a result of trophic-level changes (Humphries et al. 2019), but also may be due to various socio-economic or cultural factors (Lai et al. 2019). From 1998 onward in Western Australia, progressive management changes included reduced bag limits and size limits, that aimed to restrict catch because of declines in the CPUE of key target species (Jackson et al. 2016). These management regulations were followed by the introduction of a demersal finfish closure in the West Coast Bioregion (from 27°S, 114°E to 34°S, 115°30ʹE) between 2007 and 2010 (Fairclough et al. 2014). These regulations resulted in demonstrable stock recovery by 2011 (Newman et al. 2018), suggesting that fishers did not need to travel as far to gain their catch limits post 2011–12.
The CG for retained and released CPUE of all demersal species showed a southerly shift between 2011–12 and subsequent years, with a gradual northerly shift after 2013–14, alongside a positive area contraction of fishing events and effort. Similarly, the CG for retained and released CPUE of West Australian dhufish both shifted ~1° south between 2011–12 and 2013–14, then proceeded to migrate north in subsequent survey years. In comparison, the CG for retained and released CPUE of Baldchin groper showed only slight shifts, which is likely to reflect the fact that the species preferred habitat at the Abrolhos Islands and fewer habitats in more southerly, temperate waters (Newman et al. 2018). The southerly shift in CPUE of all demersal species and West Australian dhufish may reflect the response of target species to the 2010–11 marine heatwave that occurred off Western Australia, increasing sea surface temperatures by 4–5°C, and reaching depths of 100 m (Caputi et al. 2014).
The marine heatwave of 2010 appears to have had a short-term impact with a long-term recovery, because distributional shifts in catches were recorded. A southerly latitudinal shift was noted by commercial and recreational fishers for subtropical and temperate species, which evidentially influenced the recruitment and survival of species in the West Coast Bioregion (Caputi et al. 2014). Temperate species, such as West Australian dhufish, were more vulnerable to temperature changes where a shift in community structure can be attributed to habitat loss and relative tropicalisation of fish assemblages, where West Australian dhufish migrated to deeper waters buffering the effects of the increasing temperatures (Caputi et al. 2014). This southerly shift between 2011–12 and 2013–14, in conjunction with contraction of recreational fishing effort towards the coastline, was highlighted by the decrease in the positive area, suggesting that species are redistributing following this major disturbance, gradually migrating back north and returning to a pre-perturbed state (Caputi et al. 2014).
Areas with a greater likelihood of a hotspot for retained CPUE for all demersal species showed spatial consistencies across survey years, with the Abrolhos Islands and south of Bunbury being identified as high-activity areas over the cut-off. The likelihood of a hotspot for released CPUE for all demersal species was also high just offshore of Geraldton, adjacent to the Abrolhos Islands, in 2015–16 and 2017–18. Baldchin groper presented similar results, which was likely to reflect their aggregating nature to preferred habitat types, making the species particularly vulnerable to fishers and susceptible to overexploitation (Jennings et al. 2001; Clark and Dunn 2012; Robinson et al. 2014; Bouchet et al. 2017). Recreational fishers target demersal species with the use of global positioning systems and echo-sounder technology or prior knowledge of fish aggregations, and much like commercial fishers, fish a patch until bag and boat limits are achieved and return to those successful locations (Marriott et al. 2011; Lambert et al. 2014; Robinson et al. 2014; Hilborn et al. 2015). Identifying the likelihood of hotspot locations is of interest in a conservation approach to the spatial management of fisheries, but also may help inform on the drivers and long-term impacts of fishing in a region (Petitgas et al. 2016).
With developments in gear, advancements in fishing technology (sounders and global positioning satellites) that are widely available and the increasing efficiency of vessels (technological creep), the catchability of targeted species is increasing, especially for recreational fishers (Tidd 2013). Recreational fishing, if not managed accordingly, can potentially lead to high stock exploitation, selective harvest of ‘trophy fish’, causing shifts in population structures, and unwanted post-release mortality (FAO 2017). Although we did not account for technological creep in effort of the recreational fishery, we identified hotspots for released CPUE within the West Coast Bioregion, which is concerning because of the likelihood of barotrauma (expansion of gases within fish owing to rapid decompression) and post-release mortality (Cooke and Schramm 2007; Wise et al. 2012). Owing to bag and size limits, released CPUE are generally higher than retained CPUE for all demersal species and West Australian dhufish, which is consistent with estimated released catches being nearly twice those for estimated retained catches (Ryan et al. 2019). Recorded in the most recent 2017–18 survey, 60% of respondents released West Australian dhufish and Baldchin groper because of their being undersize (Ryan et al. 2019). Yet, since West Australian dhufish is assumed to experience a 50% post-release mortality (Department of Primary Industries and Regional Development 2021), the released CPUE would account for additional mortality to that shown by retained CPUE. High released CPUE across survey years coincide with areas of high retained CPUE supported by the LIC (0.9), suggesting fishers are releasing catches in these high-activity areas. Although current fishery management regulations are notably effective where spatial indices suggest a fishery returning to a pre-perturbed state, there is potential for alternative targeted spatial management strategies such as no-take zones for areas with high released CPUE. Because a range of targeted recreational species around the world experience high post-release mortality (Arlinghaus et al. 2007; Post and Parkinson 2012), the use of indices such as LIC is likely to assist with identifying areas of high activity and release mortality for spatial management strategies across the globe.
The distribution of retained and released CPUE for all demersal species, as well as West Australian dhufish and Baldchin groper, may have been influenced by external factors, including their unique spatial and sex-dependent growth rates, and fishing pressure from multiple sectors (commercial, charter and recreational; Morgan and Burgess 2004; Pennino et al. 2016; Maunder and Thorson 2019). Areas with high released CPUE dominate the northern region for the demersal fishery, in particular, released CPUE of Baldchin groper at the Abrolhos Islands. This archipelago is noted as an area with a few blocks of concentrated effort and is known to be a highly abundant small proportion of area (Hutchins 2001). These spatial indices, along with probability maps of released CPUE hotspots, suggest that the Abrolhos Islands are an isolated case where there is potential scope for targeted spatial management, given individual reporting of access to the fishery at the Abrolhos Islands is mandatory. An example of this type of management challenge can be seen in the Gulf of Mexico, where the declining health of the red snapper fishery is attributed to the derby-style behaviour of fishers, where they compete to attain high harvest levels before seasonal closures (Hackett et al. 2005; Smith et al. 2009; Farmer et al. 2020). Bag limits and seasonal closures alone have proven ineffective for a derby-style fishery, as effort compression can lead to higher CPUE, resulting in high released CPUE. Management restrictions for the mixed demersal fishery in the West Coast Bioregion have been effective, with <4% of annual recorded fishing events being non-compliant with bag limits exceeded (Supplementary Fig. S2). Although compliance levels in conjunction with spatial patterns suggest that the demersal fishery is returning to a pre-perturbed state, spatially targeted effort restrictions may become necessary to manage areas with high released catches, alongside bag and size limits and seasonal closures. Spatio-temporal analyses, such as those adopted in the current study, provide an effective tool to identify areas that could be targeted for spatial management.
Conclusions
The spatial behaviour of the recreational fishery is linked to fisher behaviour, unprecedented climatic events, and species biology, where patterns of retained and released CPUE were unique on a species level within the study region. This study demonstrated the value of utilising geostatistical spatial indices, including the CG and positive area, collectively with a time series of fishery-dependent data to delineate and understand potential drivers of spatial shifts in effort and CPUE distributions. Furthermore, local index of collocation helped confirm whether CPUE hotspots or areas of high fishing activity were consistent through time. These indices provide a set of tools to allow fisheries managers to adapt and apply appropriate short- to long-term management strategies, such as targeted spatial closures, to improve fish stocks. As suggested in this study, current restrictions for the demersal fishery appear effective, because effort and CPUE remained stable after 2011–12, when there was a contraction of fishing effort towards the coast and a gradual northerly shift of CPUE following the 2010–11 marine heatwave. However, there is the potential for spatially targeted effort restrictions in areas of high released CPUE, alongside existing regulations, to minimise the likelihood of barotrauma and post-release mortality. The spatial indices and hotspots used in this study are likely to assist fisheries scientists and managers to identify high fishing-activity areas and assess their potential impacts, to manage a fishery returning to a pre-perturbed state, especially in multispecies and multisector fisheries.
Data availability
The data that support this study were obtained from the Department of Primary Industries and Regional Development Western Australia by permission. Data may be made available upon approval to researchers who meet the criteria for access to confidential data (contact datarequest@dpird.wa.gov.au).
Conflicts of interest
The authors declare that they have no conflicts of interest.
Declaration of funding
This study was funded and supported by Edith Cowan University.
Supplementary material
Supplementary material is available online.
Acknowledgements
The authors acknowledge the Department of Primary Industries and Regional Development staff and Edith Cowan University’s Survey Research Centre for the collection of survey data and the recreational fishers who voluntarily participated in the surveys.
References
Aidoo EN (2016) Geostatistical modelling of recreational fishing data: a fine-scale spatial analysis. PhD Thesis, Edith Cowan University. Available at https://ro.ecu.edu.au/theses/1813/Aidoo, EN, Mueller, U, Goovaerts, P, and Hyndes, GA (2015). Evaluation of geostatistical estimators and their applicability to characterise the spatial patterns of recreational fishing catch rates. Fisheries Research 168, 20–32.
| Evaluation of geostatistical estimators and their applicability to characterise the spatial patterns of recreational fishing catch rates.Crossref | GoogleScholarGoogle Scholar | 26120221PubMed |
Aidoo, EN, Mueller, U, Hyndes, GA, and Ryan, KL (2016). The effects of measurement uncertainty on spatial characterisation of recreational fishing catch rates. Fisheries Research 181, 1–13.
| The effects of measurement uncertainty on spatial characterisation of recreational fishing catch rates.Crossref | GoogleScholarGoogle Scholar |
Arlinghaus, R, Cooke, SJ, Lyman, J, Policansky, D, Schwab, A, Suski, C, Sutton, SG, and Thorstad, EB (2007). Understanding the complexity of catch-and-release in recreational fishing: an integrative synthesis of global knowledge from historical, ethical, social, and biological perspectives. Reviews in Fisheries Science 15, 75–167.
| Understanding the complexity of catch-and-release in recreational fishing: an integrative synthesis of global knowledge from historical, ethical, social, and biological perspectives.Crossref | GoogleScholarGoogle Scholar |
Bivand RS, Pebesma EJ, Gómez-Rubio V (2008) ‘Applied spatial analysis with R.’ (Springer Media)
| Crossref |
Booth, DJ, Bond, N, and MacReadie, P (2011). Detecting range shifts among Australian fishes in response to climate change. Marine and Freshwater Research 62, 1027–1042.
| Detecting range shifts among Australian fishes in response to climate change.Crossref | GoogleScholarGoogle Scholar |
Bouchet, PJ, Meeuwig, JJ, Huang, Z, Letessier, TB, Nichol, SL, Caley, MJ, and Watson, RA (2017). Continental-scale hotspots of pelagic fish abundance inferred from commercial catch records. Global Ecology and Biogeography 26, 1098–1111.
| Continental-scale hotspots of pelagic fish abundance inferred from commercial catch records.Crossref | GoogleScholarGoogle Scholar |
Caputi N, Jackson G, Pearce A (2014) The marine heat wave off Western Australia during the summer of 2010/11 – 2 years on management implications of climate change effects on fisheries in WA: an example of an extreme event. Fisheries research report 250’. (Department of Fisheries, Western Australia: Perth, WA, Australia) Available at https://fish.wa.gov.au/Documents/research_reports/frr250.pdf
Clark, MR, and Dunn, MR (2012). Spatial management of deep-sea seamount fisheries: balancing sustainable exploitation and habitat conservation. Environmental Conservation 39, 204–214.
| Spatial management of deep-sea seamount fisheries: balancing sustainable exploitation and habitat conservation.Crossref | GoogleScholarGoogle Scholar |
Cooke, SJ, and Schramm, HL (2007). Catch-and-release science and its application to conservation and management of recreational fisheries. Fisheries Management and Ecology 14, 73–79.
| Catch-and-release science and its application to conservation and management of recreational fisheries.Crossref | GoogleScholarGoogle Scholar |
Cressie N (1991) ‘Statistics for spatial data.’ (Wiley)
Currie, JC, Thorson, JT, Sink, KJ, Atkinson, LJ, Fairweather, TP, and Winker, H (2019). A novel approach to assess distribution trends from fisheries survey data. Fisheries Research 214, 98–109.
| A novel approach to assess distribution trends from fisheries survey data.Crossref | GoogleScholarGoogle Scholar |
Department of Primary Industries and Regional Development (2021) ‘Fisheries management paper number 305, West Coast demersal scalefish resource harvest strategy 2021–2025.’ (State of Western Australia: Perth, WA, Australia). Available at http://www.fish.wa.gov.au/Documents/management_papers/fmp305.pdf
Fairclough DV, Molony BW, Crisafulli BM, Keay IS, Hesp SA, Marriott RJ (2014) ‘Status of demersal finfish stocks on the west coast of Australia. Fisheries research report number 253’. (Department of Fisheries, Western Australia: Perth, WA, Australia) Available at http://www.fish.wa.gov.au/Documents/research_reports/frr253.pdf
(2017). The role of recreational fisheries in the sustainable management of marine resources. GLOBEFISH Highlights 2, 65–68.
Farmer, NA, Froeschke, JT, and Records, DL (2020). Forecasting for recreational fisheries management: a derby fishery case study with Gulf of Mexico red snapper. ICES Journal of Marine Science 77, 2265–2284.
| Forecasting for recreational fisheries management: a derby fishery case study with Gulf of Mexico red snapper.Crossref | GoogleScholarGoogle Scholar |
Gaughan DJ, Santoro K (Eds) (2020) Status reports of the fisheries and aquatic resources of Western Australia 2018/19: state of the fisheries. (Department of Primary Industries and Regional Development, WA) Available at http://www.fish.wa.gov.au/Documents/sofar/status_reports_of_the_fisheries_and_aquatic_resources_2018-19.pdf
Griffith, D (2007). Spatial structure and spatial interaction: 25 years later. The Review of Regional Studies 37, 28–38.
| Spatial structure and spatial interaction: 25 years later.Crossref | GoogleScholarGoogle Scholar |
Griffiths, S, and Fay, G (2015). Integrating recreational fisheries data into stock assessment: implications for model performance and subsequent harvest strategies. Fisheries Management and Ecology 22, 197–212.
| Integrating recreational fisheries data into stock assessment: implications for model performance and subsequent harvest strategies.Crossref | GoogleScholarGoogle Scholar |
Hackett, SC, Krachey, MJ, Brown, S, and Hankin, D (2005). Derby fisheries, individual quotas, and transition in the fish processing industry. Marine Resource Economics 20, 47–60.
| Derby fisheries, individual quotas, and transition in the fish processing industry.Crossref | GoogleScholarGoogle Scholar |
Hartill, BW, Payne, GW, Rush, N, and Bian, R (2016). Bridging the temporal gap: continuous and cost-effective monitoring of dynamic recreational fisheries by web cameras and creel surveys. Fisheries Research 183, 488–497.
| Bridging the temporal gap: continuous and cost-effective monitoring of dynamic recreational fisheries by web cameras and creel surveys.Crossref | GoogleScholarGoogle Scholar |
Henry GW, Lyle JM (Eds) (2003) The National Recreational and Indigenous Fishing survey. FRDC project number 99/158. (Australian Government Department of Agriculture, Fisheries and Forestry: Canberra, ACT, Australia) Available at https://eprints.utas.edu.au/2526/1/Henry_Lyle_Nationalsurvey.pdf
Hilborn, R, Fulton, EA, Green, BS, Hartmann, K, Tracey, SR, and Watson, RA (2015). When is a fishery sustainable? Canadian Journal of Fisheries and Aquatic Sciences 72, 1433–1441.
| When is a fishery sustainable?Crossref | GoogleScholarGoogle Scholar |
Hoenig, JM, Jones, CM, Pollock, KH, Robson, DS, and Wade, DL (1997). Calculation of catch rate and total catch in roving surveys of anglers. Biometrics 53, 306–317.
| Calculation of catch rate and total catch in roving surveys of anglers.Crossref | GoogleScholarGoogle Scholar |
Humphries, AT, Gorospe, KD, Carvalho, PG, Yulianto, I, Kartawijaya, T, and Campbell, SJ (2019). Catch composition and selectivity of fishing gears in a multi-species Indonesian coral reef fishery. Frontiers in Marine Science 6, 378.
| Catch composition and selectivity of fishing gears in a multi-species Indonesian coral reef fishery.Crossref | GoogleScholarGoogle Scholar |
Hutchins, JB (2001). Biodiversity of shallow reef fish assemblages in Western Australia using a rapid censusing technique. Records of the Western Australian Museum 20, 247–270.
Jackson, G, Ryan, KL, Green, TJ, Pollock, KH, and Lyle, JM (2016). Assessing the effectiveness of harvest tags in the management of a small-scale, iconic marine recreational fishery in Western Australia. ICES Journal of Marine Science 73, 2666–2676.
| Assessing the effectiveness of harvest tags in the management of a small-scale, iconic marine recreational fishery in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Jennings S, Kaiser MJ, Reynolds JD (2001) ‘Marine fisheries ecology.’ (Blackwell Science Ltd)
Kleisner, KM, Walter, JF, Diamond, SL, and Die, DJ (2010). Modeling the spatial autocorrelation of pelagic fish abundance. Marine Ecology Progress Series 411, 203–213.
| Modeling the spatial autocorrelation of pelagic fish abundance.Crossref | GoogleScholarGoogle Scholar |
Kulka, DW, Hendrickson, L, Bez, N, Schlitzer, R, Black, GAP, and Simpson, MR (2003). Report of the workshop on mapping and geostatistical methods for fisheries stock assessment. Northwest Atlantic Fisheries Organization Scientific Council Studies 39, 1–43.
Lai, EKM, Mueller, U, Hyndes, GA, and Ryan, KL (2019). Comparing estimates of catch and effort for boat-based recreational fishing from aperiodic access-point surveys. Fisheries Research 219, 105305.
| Comparing estimates of catch and effort for boat-based recreational fishing from aperiodic access-point surveys.Crossref | GoogleScholarGoogle Scholar |
Lambert, GI, Jennings, S, Kaiser, MJ, Davies, TW, and Hiddink, JG (2014). Quantifying recovery rates and resilience of seabed habitats impacted by bottom fishing. Journal of Applied Ecology 51, 1326–1336.
| Quantifying recovery rates and resilience of seabed habitats impacted by bottom fishing.Crossref | GoogleScholarGoogle Scholar |
Lynch, TP (2006). Incorporation of recreational fishing effort into design of marine protected areas. Conservation Biology 20, 1466–1476.
| Incorporation of recreational fishing effort into design of marine protected areas.Crossref | GoogleScholarGoogle Scholar | 17002764PubMed |
Lynch, TP (2014). A decadal time-series of recreational fishing effort collected during and after implementation of a multiple use marine park shows high inter-annual but low spatial variability. Fisheries Research 151, 85–90.
| A decadal time-series of recreational fishing effort collected during and after implementation of a multiple use marine park shows high inter-annual but low spatial variability.Crossref | GoogleScholarGoogle Scholar |
Lynch, TP, Smallwood, CB, Ochwada-Doyle, FA, Lyle, J, Williams, J, Ryan, KL, Devine, C, Gibson, B, and Jordan, A (2020). A cross continental scale comparison of Australian offshore recreational fisheries research and its applications to marine park and fisheries management. ICES Journal of Marine Science 77, 1190–1205.
| A cross continental scale comparison of Australian offshore recreational fisheries research and its applications to marine park and fisheries management.Crossref | GoogleScholarGoogle Scholar |
Marriott, RJ, Wise, B, and St John, J (2011). Historical changes in fishing efficiency in the west coast demersal scalefish fishery, Western Australia: implications for assessment and management. ICES Journal of Marine Science 68, 76–86.
| Historical changes in fishing efficiency in the west coast demersal scalefish fishery, Western Australia: implications for assessment and management.Crossref | GoogleScholarGoogle Scholar |
Maunder, MN, and Thorson, JT (2019). Modeling temporal variation in recruitment in fisheries stock assessment: a review of theory and practice. Fisheries Research 217, 71–86.
| Modeling temporal variation in recruitment in fisheries stock assessment: a review of theory and practice.Crossref | GoogleScholarGoogle Scholar |
McPhee, DP, Leadbitter, D, and Skilleter, GA (2002). Swallowing the bait: is recreational fishing in Australia ecologically sustainable? Pacific Conservation Biology 8, 40–51.
| Swallowing the bait: is recreational fishing in Australia ecologically sustainable?Crossref | GoogleScholarGoogle Scholar |
Miller, HJ (2004). Tobler’s first law and spatial analysis. Annals of the Association of American Geographers 94, 284–289.
| Tobler’s first law and spatial analysis.Crossref | GoogleScholarGoogle Scholar |
Morgan AC, Burgess GH (2004) 11. Fishery-dependent sampling: total catch, effort and catch composition. In ‘Management techniques for elasmobranch fisheries’. (Eds JA Musick, R Bonfil) FAO Fisheries Technical Paper 474, pp. 182–200. (Food and Agriculture Organization of the United Nations: Rome, Italy) Available at https://www.fao.org/3/a0212e/A0212E15.htm#ch11
Mueller, U, Kangas, M, Sporer, E, and Caputi, N (2012). Variability in the spatial and temporal distribution of the saucer scallop, Amusium balloti, in Shark Bay – management implications. Marine and Freshwater Research 63, 1152–1164.
| Variability in the spatial and temporal distribution of the saucer scallop, Amusium balloti, in Shark Bay – management implications.Crossref | GoogleScholarGoogle Scholar |
Myers, N, Mittermeier, RA, Mittermeier, CG, da Fonseca, GAB, and Kent, J (2000). Biodiversity hotspots for conservation priorities. Nature 403, 853–858.
| Biodiversity hotspots for conservation priorities.Crossref | GoogleScholarGoogle Scholar | 10706275PubMed |
Navarro, M, Hailu, A, Langlois, T, Ryan, KL, and Kragt, ME (2020). Determining spatial patterns in recreational catch data: a comparison of generalized additive mixed models and boosted regression trees. ICES Journal of Marine Science 77, 2316–2225.
| Determining spatial patterns in recreational catch data: a comparison of generalized additive mixed models and boosted regression trees.Crossref | GoogleScholarGoogle Scholar |
Newman, SJ, Brown, JI, Fairclough, DV, Wise, BS, Bellchambers, LM, Molony, BW, Lenanton, RCJ, Jackson, G, Smith, KA, Gaughan, DJ, Fletcher, WJ, McAuley, RB, and Wakefield, CB (2018). A risk assessment and prioritisation approach to the selection of indicator species for the assessment of multi-species, multi-gear, multi-sector fishery resources. Marine Policy 88, 11–22.
| A risk assessment and prioritisation approach to the selection of indicator species for the assessment of multi-species, multi-gear, multi-sector fishery resources.Crossref | GoogleScholarGoogle Scholar |
Parnell, PE, Dayton, PK, Fisher, RA, Loarie, CC, and Darrow, RD (2010). Spatial patterns of fishing effort off San Diego: implications for zonal management and ecosystem function. Ecological Applications 20, 2203–2222.
| Spatial patterns of fishing effort off San Diego: implications for zonal management and ecosystem function.Crossref | GoogleScholarGoogle Scholar | 21265452PubMed |
Pennino, MG, Conesa, D, López-Quílez, A, Muñoz, F, Fernández, A, and Bellido, JM (2016). Fishery-dependent and -independent data lead to consistent estimations of essential habitats. ICES Journal of Marine Science 73, 2302–2310.
| Fishery-dependent and -independent data lead to consistent estimations of essential habitats.Crossref | GoogleScholarGoogle Scholar |
Perry, AL, Low, PJ, Ellis, JR, and Reynolds, JD (2005). Ecology: climate change and distribution shifts in marine fishes. Science 308, 1912–1915.
| Ecology: climate change and distribution shifts in marine fishes.Crossref | GoogleScholarGoogle Scholar | 15890845PubMed |
Petitgas, P (2001). Geostatistics in fisheries survey design and stock assessment: models, variances and applications. Fish and Fisheries 2, 231–249.
| Geostatistics in fisheries survey design and stock assessment: models, variances and applications.Crossref | GoogleScholarGoogle Scholar |
Petitgas, P, Poulard, JC, and Biseau, A (2003). Comparing commercial and research survey catch per unit of effort: Megrim in the Celtic Sea. ICES Journal of Marine Science 60, 66–76.
| Comparing commercial and research survey catch per unit of effort: Megrim in the Celtic Sea.Crossref | GoogleScholarGoogle Scholar |
Petitgas, P, Woillez, M, Doray, M, and Rivoirard, J (2016). A geostatistical definition of hotspots for fish spatial distributions. Mathematical Geosciences 48, 65–77.
| A geostatistical definition of hotspots for fish spatial distributions.Crossref | GoogleScholarGoogle Scholar |
Petitgas P, Woillez M, Rivoirard J, Renard D, Bez N (2017) Handbook of geostatistics in R for fisheries and marine ecology. ICES cooperative research report 338,. (International Council for the Exploration of the Sea: Copenhagen, Denmark)
| Crossref |
Petitgas, P, Woillez, M, Doray, M, and Rivoirard, J (2018). Indicator-based geostatistical models for mapping fish survey data. Mathematical Geosciences 50, 187–208.
| Indicator-based geostatistical models for mapping fish survey data.Crossref | GoogleScholarGoogle Scholar |
Polansky, L, Newman, KB, Nobriga, ML, and Mitchell, L (2018). Spatiotemporal models of an estuarine fish species to identify patterns and factors impacting their distribution and abundance. Estuaries and Coasts 41, 572–581.
| Spatiotemporal models of an estuarine fish species to identify patterns and factors impacting their distribution and abundance.Crossref | GoogleScholarGoogle Scholar |
Post, JR, and Parkinson, EA (2012). Temporal and spatial patterns of angler effort across lake districts and policy options to sustain recreational fisheries. Canadian Journal of Fisheries and Aquatic Sciences 69, 321–329.
| Temporal and spatial patterns of angler effort across lake districts and policy options to sustain recreational fisheries.Crossref | GoogleScholarGoogle Scholar |
Roa-Ureta, R, and Niklitschek, E (2007). Biomass estimation from surveys with likelihood-based geostatistics. ICES Journal of Marine Science 64, 1723–1734.
| Biomass estimation from surveys with likelihood-based geostatistics.Crossref | GoogleScholarGoogle Scholar |
Robinson, J, Cinner, JE, and Graham, NAJ (2014). The influence of fisher knowledge on the susceptibility of reef fish aggregations to fishing. PLoS One 9, e91296.
| The influence of fisher knowledge on the susceptibility of reef fish aggregations to fishing.Crossref | GoogleScholarGoogle Scholar | 24646910PubMed |
Ryan KL, Hall NG, La EK, Smallwood CB, Tate A, Taylor SM, Wise BS (2019) Statewide survey of boat-based recreational fishing in Western Australia 2017/18. Fisheries research report number 297, Department of Primary Industries and Regional Development, Western Australia. Available at http://www.fish.wa.gov.au/Documents/research_reports/frr297.pdf
Santora, JA, Sydeman, WJ, Schroeder, ID, Wells, BK, and Field, JC (2011). Mesoscale structure and oceanographic determinants of krill hotspots in the California Current: implications for trophic transfer and conservation. Progress in Oceanography 91, 397–409.
| Mesoscale structure and oceanographic determinants of krill hotspots in the California Current: implications for trophic transfer and conservation.Crossref | GoogleScholarGoogle Scholar |
Santos, RO, Rehage, JS, Kroloff, EKN, Heinen, JE, and Adams, AJ (2019). Combining data sources to elucidate spatial patterns in recreational catch and effort: fisheries-dependent data and local ecological knowledge applied to the South Florida bonefish fishery. Environmental Biology of Fishes 102, 299–317.
| Combining data sources to elucidate spatial patterns in recreational catch and effort: fisheries-dependent data and local ecological knowledge applied to the South Florida bonefish fishery.Crossref | GoogleScholarGoogle Scholar |
Smallwood, CB, and Beckley, LE (2012). Spatial distribution and zoning compliance of recreational fishing in Ningaloo Marine Park, north-western Australia. Fisheries Research 125–126, 40–50.
| Spatial distribution and zoning compliance of recreational fishing in Ningaloo Marine Park, north-western Australia.Crossref | GoogleScholarGoogle Scholar |
Smallwood C, Hesp S, Beckley L (2013) Biology, stock status and management summaries for selected fish species in south-western Australia. Fisheries research report 242. (Department of Fisheries, Western Australia: Perth, WA, Australia) Available at https://fish.wa.gov.au/Documents/research_reports/frr242.pdf
Smith, JN, Snyder, SM, Berkson, J, Murphy, BR, and McMullin, SL (2009). Fisheries management of red snapper in the gulf of Mexico: a case study. Journal of Natural Resources and Life Sciences Education 38, 115–127.
| Fisheries management of red snapper in the gulf of Mexico: a case study.Crossref | GoogleScholarGoogle Scholar |
Stuart-Smith, RD, Bates, AE, Lefcheck, JS, Duffy, JE, Baker, SC, Thomson, RJ, Stuart-Smith, JF, Hill, NA, Kininmonth, SJ, Airoldi, L, Becerro, MA, Campbell, SJ, Dawson, TP, Navarrete, SA, Soler, GA, Strain, EMA, Willis, TJ, and Edgar, GJ (2013). Integrating abundance and functional traits reveals new global hotspots of fish diversity. Nature 501, 539–542.
| Integrating abundance and functional traits reveals new global hotspots of fish diversity.Crossref | GoogleScholarGoogle Scholar | 24067714PubMed |
Tao, JP, Gong, YT, Tan, XC, Yang, Z, and Chang, JB (2012). Spatiotemporal patterns of the fish assemblages downstream of the Gezhouba Dam on the Yangtze River. Science China. Life Sciences 55, 626–636.
| Spatiotemporal patterns of the fish assemblages downstream of the Gezhouba Dam on the Yangtze River.Crossref | GoogleScholarGoogle Scholar |
Tidd, AN (2013). Effective fishing effort indicators and their application to spatial management of mixed demersal fisheries. Fisheries Management and Ecology 20, 377–389.
| Effective fishing effort indicators and their application to spatial management of mixed demersal fisheries.Crossref | GoogleScholarGoogle Scholar |
van Putten, IE, Jennings, S, Hobday, AJ, Bustamante, RH, Dutra, LXC, Frusher, S, Fulton, EA, Haward, M, Plagányi, É, Thomas, L, and Pecl, G (2017). Recreational fishing in a time of rapid ocean change. Marine Policy 76, 169–177.
| Recreational fishing in a time of rapid ocean change.Crossref | GoogleScholarGoogle Scholar |
Winfield, IJ (2016). Recreational fisheries in the UK: natural capital, ecosystem services, threats, and management. Fisheries Science 82, 203–212.
| Recreational fisheries in the UK: natural capital, ecosystem services, threats, and management.Crossref | GoogleScholarGoogle Scholar |
Wise, BS, Telfer, CF, Lai, EKM, Hall, NG, and Jackson, G (2012). Long-term monitoring of boat-based recreational fishing in Shark Bay, Western Australia: providing scientific advice for sustainable management in a World Heritage Area. Marine and Freshwater Research 63, 1129–1141.
| Long-term monitoring of boat-based recreational fishing in Shark Bay, Western Australia: providing scientific advice for sustainable management in a World Heritage Area.Crossref | GoogleScholarGoogle Scholar |
Woillez M, Petitgas P, Rivoirard J, Poulard, J (2005) Indices for capturing spatial pattern and change across years of a fish population: an application on European Hake (Merluccius merluccius) in the Bay of Biscay. ICES Communication 33. Available at https://www.ices.dk/sites/pub/CM%20Doccuments/2005/L/L1605.pdf
Woillez, M, Walline, PD, Ianelli, JN, Dorn, MW, Wilson, CD, and Punt, AE (2016). Evaluating total uncertainty for biomass- and abundance-at-age estimates from eastern Bering Sea walleye pollock acoustic-trawl surveys. ICES Journal of Marine Science 73, 2208–2226.
| Evaluating total uncertainty for biomass- and abundance-at-age estimates from eastern Bering Sea walleye pollock acoustic-trawl surveys.Crossref | GoogleScholarGoogle Scholar |
Yasuda, T, Yukami, R, and Ohshimo, S (2014). Fishing ground hotspots reveal long-term variation in chub mackerel Scomber japonicus habitat in the East China Sea. Marine Ecology Progress Series 501, 239–250.
| Fishing ground hotspots reveal long-term variation in chub mackerel Scomber japonicus habitat in the East China Sea.Crossref | GoogleScholarGoogle Scholar |