

Abrupt historical change in the nature of a Ramsar Wetland: Seaford Swamp, greater Melbourne, Australia
Peter A. Gell A B C * , Emmy Frost D , Cameron Gell A and Rebekah Kurpiel D
A B C * , Emmy Frost D , Cameron Gell A and Rebekah Kurpiel D
A
B
C
D
Abstract
Seaford Swamp is a back-of-dune wetland on the eastern side of Port Phillip Bay in greater urban Melbourne. It is a fresh marsh dominated by the reed Phragmites australis that is expanding into open water.
Three sediment cores were collected to determine the ecological history of the wetland.
The cores were analysed for diatoms and pollen and dated using 14C and 210Pb.
The longest core showed an abrupt change from marine–brackish to saline and ultimately the present fresh condition. Saltmarsh declined and was replaced by reed, sedge and rush species. Cores from the centre and south of the wetland showed a similar history, albeit more variable in the south, yet still reflecting a sustained change from marine–estuarine conditions to fresh–brackish. The 7000-year record showed considerable stability despite known climatic and sea-level changes.
The abrupt change in condition appeared to correspond with hydrological engineering of the site from 1870 for agricultural and urban development. Increased catchment runoff, including from urban areas, are likely to have sustained the freshwater state.
The marine–estuarine state represents an alternate condition of the swamp that may broaden options for management under future sea-level rise.
Keywords: diatoms, palaeoenvironmental reconstruction, Phragmites australis, Ramsar Wetland, saltmarsh, Seaford Swamp, urban runoff, wetland change.
Introduction
Wetlands change in response to natural drivers such as climate and hydrology (Reidinger-Whitmore et al. 2023), as well as a result of pressures from humans (Mills et al. 2023). In many instances, the first impacts of people are ancient (Dubois et al. 2018) and so the natural state of the wetland is obscured by time. Long-term data, historical records and palaeoecological information can expand our understanding of the response of ecosystems to natural or anthropogenic impacts (Manzano et al. 2020). The knowledge derived from these approaches provides evidence for the condition pathway through which a wetland has passed. This evidence may be critical for current management because it provides evidence for variability, present transience or stability, and the primary drivers of condition. Although human recollection may provide evidence for past condition, it can be punctuated, subjective and dominated by recent experience (Tibby et al. 2008). Palaeoecological approaches often provide continuous records of change, and while resolution diminishes with sediment depth, can provide evidence of state and transition beyond memory and documentary records.
The extraction of evidence of past condition is available through the exhumation of continuous sediment sequences and the extraction from the sediment layers of biological and chemical indicators of wetland state. These palaeolimnological approaches have now been widely applied to wetlands across south-eastern Australia (e.g. Head 1987; Gell et al. 2005a; MacGregor et al. 2005; Reid et al. 2007; Cook et al. 2016), often showing natural character states that have been obscured with time.
The Ramsar Convention on Wetlands came into effect in 1975, with the aim of mitigating the loss and degradation of the world’s wetlands (Stroud et al. 2022). It was widely recognised that, by then, many wetlands had been lost or changed from their historic condition (Gell et al. 2023). This trend of loss and degradation has continued (Davidson et al. 2023), but it is believed that the Convention has acted to slow its progress. The ecological character of Ramsar sites (wetlands listed under the Convention as internationally important) are described at the time-of-listing, but signatory nations are not obliged to use the ‘time-of-listing’ state as the natural ‘ecological character’ (Pritchard 2022). Where evidence shows a past condition, then restoration to that target may be appropriate, particularly if that represents a ‘positive change’. Importantly, site managers otherwise seek to retain the ecological character, including the biota and ecosystem services, of the wetlands, as well as the biodiversity criteria used to list the site as internationally important (Pittock et al. 2010).
In wetlands that are on a trajectory of degradation, management interventions that reinstate a past condition (or ecological character) and retain at least one criterion for listing may be preferred to options that seek to retain time-of-listing conditions. Such a decision was undertaken in Chilika Lake in India that suffered from high sedimentation and a transition to primarily freshwater (Kumar et al. 2020a). Here palaeoecological evidence (Pandey et al. 2014) showed greater connection to the ocean in the past and management invested in establishing an opening to the sea to recover lost fisheries production as well as species more dependent on brackish–saline conditions, such as mangrove communities (Kumar et al. 2020a, 2020b).
The Seaford and Edithvale Wetlands are located on Bunurong Country and were originally part of a continuous ‘Carrum Carrum Swamp’ that ran parallel to the coast behind the foredune systems, from near the modern-day Melbourne suburbs Mordialloc to Frankston (Department of Sustainability and Environment 2012). From 1866, this wetland system was subjected to considerable deliberate change for the purpose of establishing, first, market gardening and dairy farming, and ultimately, residential communities within the developing Melbourne metropolitan area (Table 1). Today the hydrological system of Seaford Swamp has substantially changed in that it no longer receives water from Carrum Swamp and is now fed from a pump on a drain and via tidal inflows from Kananook Creek via another drain (Lane 2011). Quin et al. (2016) noted that tall marsh vegetation, dominated by common reed (Phragmites australis), has been expanding into other local wetland vegetation types reducing floristic diversity and faunal habitat suitability. This is presently managed by grooming at considerable cost to the local managers.
| Date | Activity | |
|---|---|---|
| 1866 | Carrum Swamp surveyed | |
| 1871 | Carrum Swamp opened for selection | |
| 1873 | First contracts let for Drainage | |
| 1879 | Construction of the ‘Patterson Cut’, a 10-m-wide drainage channel | |
| 1882 | Railway extended from Caulfield to Frankston | |
| 1901 | Small blocks sold near Carrum railway station | |
| 1910 | Local road upgraded as McLeod Road | |
| 1974 | Development of Patterson Lakes | |
| 2001 (29 Aug) | Ramsar site designation |
There are many biological indicator groups that can be extracted from sediments to reconstruct the past condition of wetlands. Diatoms are a form of algae that are sensitive to water quality (Reid et al. 1995). They have a skeleton of silica and so they are often preserved as fossils in sediments. The availability of databases of diatom species and water-quality data from studies across Australia (e.g. Gell 1997; Philibert et al. 2006) and internationally (e.g. Hall and Smol 1992; Gasse et al. 1995; Witkowski et al. 2000) provide a means of interpreting water quality from the fossil diatom record. The extraction of diatoms from swamp and estuarine sediment sequences can, therefore, provide a record of changes in water quality over time (Taffs et al. 2017a). Similarly, pollen can be preserved in wetland sediments and the identification and enumeration of these can yield evidence for past terrestrial and aquatic vegetation (Ellison 2017). Important to understanding ecological change from these fossil assemblages is the establishment of a timeline. The chronology of sediment cores can be estimated using biological (exotic pollen markers) and geochemical (radiometric isotopes) means (Last and Smol 2001).
Materials and methods
Seaford Swamp was visited on 22 July 2022. Three sites were selected for coring on the basis of ease-of-access, and the anecdotal record of permanency of inundation. The sites selected were in the south (SFS), central (SFC) and northern (SFN) sections of the Swamp (Fig. 1). The central points in sites with open water were accessed on foot with a support canoe (Fig. 2). The initial cores, extracted using a d-section hand-corer (Jowsey 1966), were short. Maximum core lengths were 48 cm (SFS), 60 cm (SFC) and 85 cm (SFN). Preliminary analysis of the sediments showed that diatoms were preserved and that broad changes in the assemblages were evident down core. This outcome justified a detailed analysis combining diatom analysis, with analysis of preserved plant pollen (palynology) and radiometric dating (210Pb; 14C). Seaford Swamp was visited again for sediment coring on 2 February 2024. In this instance, a 2.0-m core was extracted from the northern site (SFN; Fig. 3) and cores of 50 and 64 cm were extracted from the central (SFC) and southern locations (SFS) shown on Fig. 1. All cores were wrapped with plastic and stored in a refrigerator until they were subsampled.
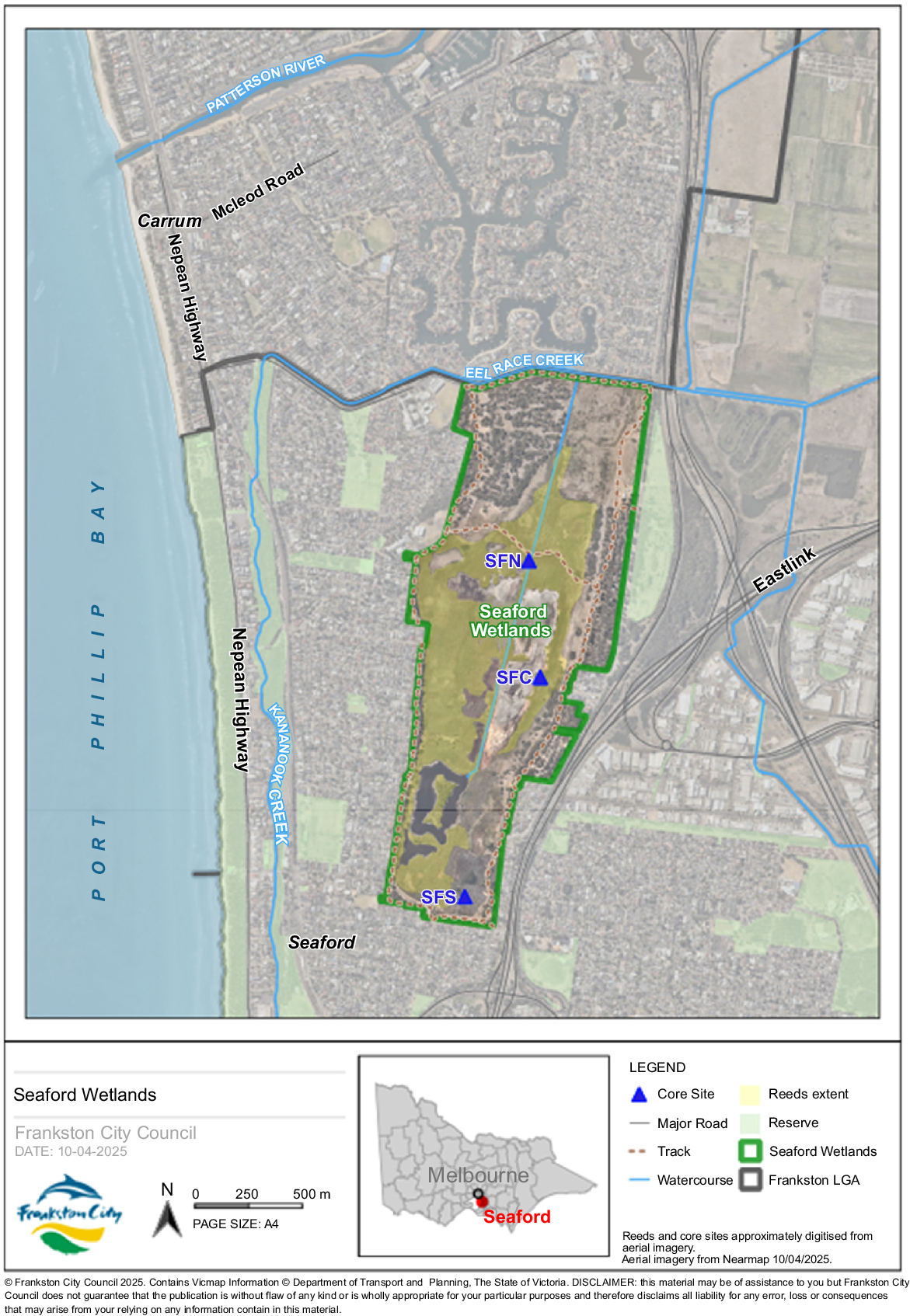
Location of Core SFN showing open water conditions and stands of common reed in the distance.


The longer core (SFN) was selected as a focus for detailed analysis and dating. In total, 30 samples were taken from the 2.0-m core for pollen and diatom analysis at increasing intervals down core, and 20 samples were taken for supplementary diatom analysis of the SFC and SFS cores.
For diatom analysis, the samples were treated with HCl and H2O2 and mounted on slides with Naphrax, following standard procedures (Battarbee et al. 2001). The coverslips were scanned with a Zeiss Axiolab microscope at 1000× magnification under phase contrast. At least 100 diatoms, and as many as 260, were identified from each sample, by reference to Krammer and Lange-Bertalot (1986, 1988, 1991a, 1991b), Foged (1978) and Witkowski et al. (2000), and enumerated. Species counts were expressed as a percentage of the total and presented as a stratigraphic diagram by using the program C2 (ver. 1.8.0, see https://www.staff.ncl.ac.uk/stephen.juggins/software/C2Home.htm; Juggins 2007). Species were classified as marine, mesosaline, fresh–oligosaline and eutrophic on the basis of the above flora and a range of national and international databases. The diatoms were grouped into zones delineated by sustained changes in the main flora.
Samples for pollen analysis were prepared following standard acetolysis approaches following Gosling et al. (2022). Usually at least 300 pollen grains were identified following the Australian pollen database. Pollen counts are expressed as a percentage of total dryland pollen and are presented as a stratigraphic diagram by using C2.
Ten subsamples across the upper 50 cm of Core SFN were sent to the laboratories at the Australian Nuclear Science and Technology Organisation (ANSTO) in Menai (Sydney) for radiometric analysis. Samples were prepared following standard procedures and activated in an alpha (small) counter. Initially, four samples were analysed (0, 10, 20, 30 cm) and the results of these were used to select the other two samples for analysis (5, 15 cm). The basal sediments of Core SFN held many small shells and these were extracted, wrapped in alfoil, and sent to the Chronos Radiocarbon Laboratory at UNSW for AMS 14C dating.
Results
Sediment stratigraphy and dating
The 200-cm SFN core showed a clear stratigraphic change, with marl-like grey clays from the base to 25 cm (Fig. 3), above which were organic sediments with plant remains. Similar grey clays were evident at the base of the 50-cm core from SFC and the 64-cm core from SFS. Generally, there was a sharp transition to organic surface sediments, although at SFS this varied with alternating grey clay and organic sediments.
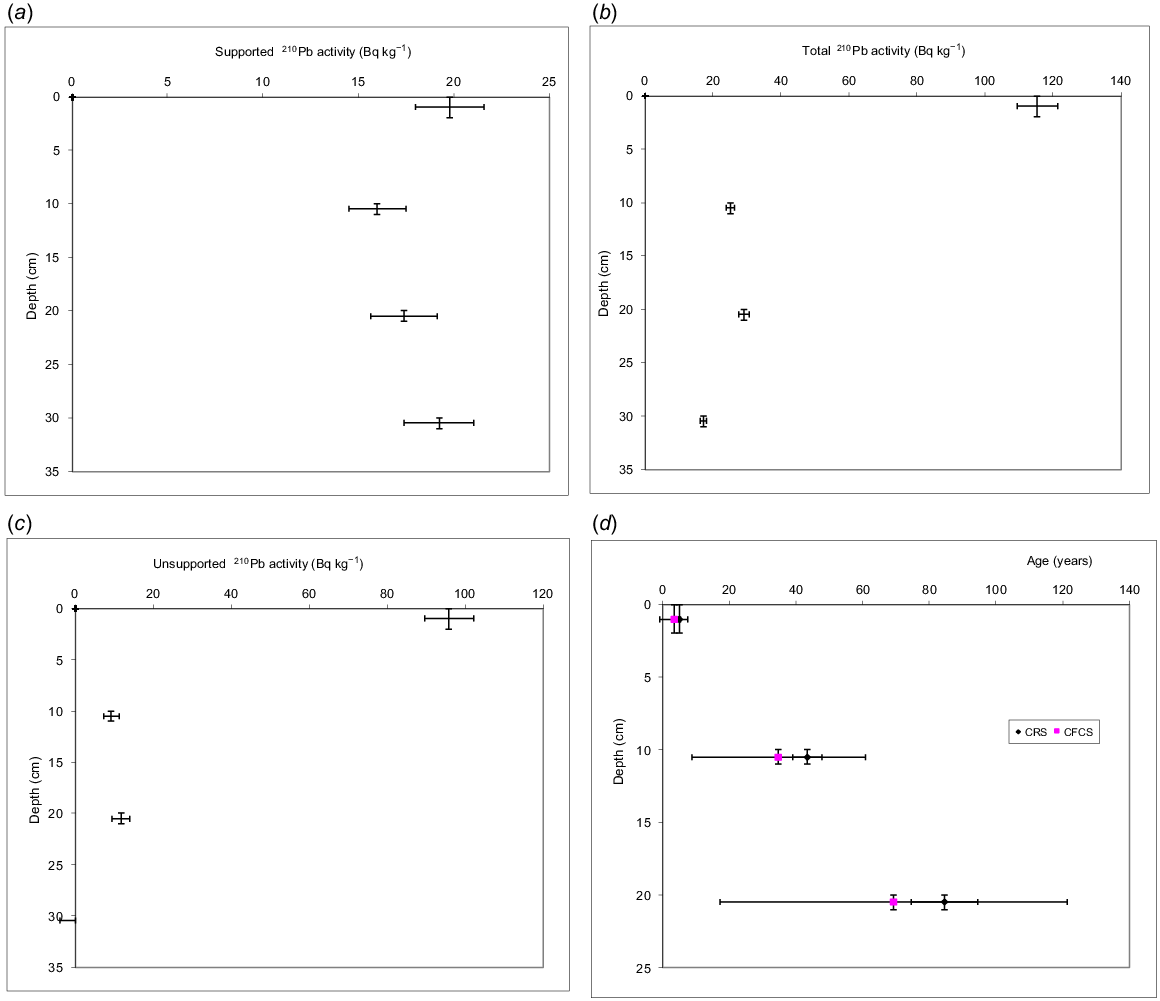
The preliminary 210Pb data from the initial four samples are provided as Fig. 4. Here, the sample at 30 cm is fully depleted (−2 ± 2 Bq kg−1), suggesting an age of >> 80 years. The unsupported activity at 20 cm suggests an age of 72 or 90 years, depending on whether the Constant Flux Constant Sedimentation (CFCS) or Constant Rate of Supply (CRS) models are applied (see Appleby 2001). This would suggest that the sediments at 20 cm were deposited in c. 1943 CE, yielding an average sedimentation rate of ~2.5 mm year−1. The sample at 15 cm yielded background results (Table 2), confounding the initial age–depth curve attempted from the initial samples. Further, the unsupported 210Pb at 5–6 cm was lower than at 10–11 cm. With these additional results, the low values at 5 cm are now considered ‘background’, even though an age was initially inferred for 20–21 cm. 210Pb analyses can be affected by low sedimentation rates, variable sources of sediment, and salinity (Gell et al. 2005b). It may also be affected by bioturbation; however, the clear diatom stratigraphy questions that prospect. Here, we tentatively accept the sedimentation rates inferred from the first four samples; however, clearly the timeline requires support from additional analyses.
210Pb dating output from the initial four samples. The difference between the supported 210Pb (a) and the total 210Pb (b) provides the unsupported 210Pb values (c). The depletion of the unsupported 210Pb is modelled to yield the age of the samples (d) relative to the surface.
| ANSTO ID | Depth (cm) | Supported 210Pb | Unsupported 210Pb | |
|---|---|---|---|---|
| Z605 | 0–2 | 20 ± 2 | 96 ± 6 | |
| Z606 | 5–6 | 19 ± 2 | 8 ± 2 | |
| Z607 | 10–11 | 16 ± 1 | 9 ± 2 | |
| Z608 | 15–16 | 22 ± 2 | −11 ± 2 | |
| Z609 | 20–21 | 17 ± 2 | 12 ± 2 | |
| Z611 | 30–31 | 19 ± 2 | −2 ± 2 |
One AMS 14C age was sought from shells collected from sediments between 190 and 195 cm. The raw result from the analysis was 6330 ± 21 years BP (UNSW-3375), which was converted to 6713 ± 83 years BP (Table 3) after the marine reservoir effect was factored in, following Reimer and Reimer (2001).
| UNSW lab code | Sample label | Date 14C year BP | Date ± 14C year BP | Calibrated age | Age± | |
|---|---|---|---|---|---|---|
| UNSW-3375 | SFN (190–195 cm) | 6330 | 21 | 6713 | 83 |
The arrival of Pinus pollen at 12 cm confirms a post-European age supported by increases in exotic plantain and daisies at ~30 cm (Fig. 5). The arrival of pine pollen has been used to mark 1850 AD, but Bickford et al. (2008) showed that it first appeared in some sediment records much later.
Diatoms
Most diatom valves were readily identified to species. The identification of some types within the grey clay sediments were identified with less confidence because the valves were often fractured, and in places partly dissolved. Here, depending on species shape, the central area was counted as one, whereas ends were counted as a half. So, the more heavily silicified species were identified more readily and others were mostly recognised where large fragments or entire valves were seen.
The diatom analysis of 30 samples from the SFN core yielded 146 diatom types. The C2 stratigraphic diagram of the diatom assemblages across the 2.0 m of the SFN sediment core is provided as Fig. 6.
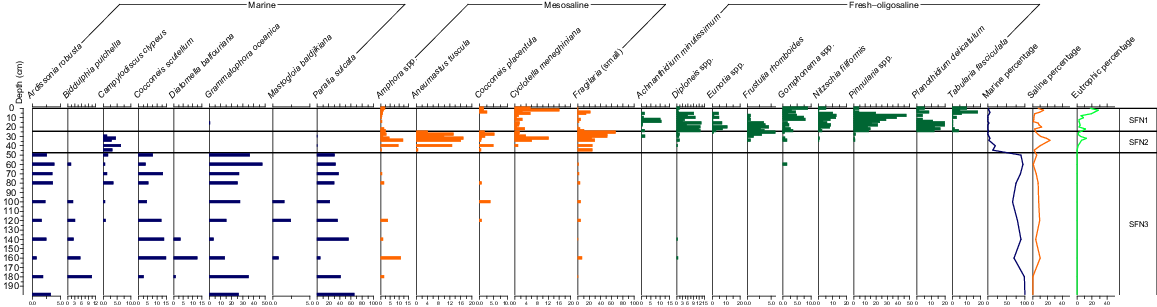
The diatoms of the lower 150 cm are dominated by marine–estuarine taxa, in particular Grammatophora oceanica and Paralia sulcata. Other species typical of coastal waters recorded were Ardissonia robusta, Biddulphia pulchella, Campylodiscus clypeus, Cocconeis scutellum, Cocconeis heteroidea, Diatomella balfouriana, Mastogloia baldjikiana, Navicula (syn. Pinnuavis) yarrensis, Opephora olsenii, Plagiogramma staurophorum, Rhaphoneis amphiceros and Trachyneis australis. Within these sediments, there is a transition from Biddulphia pulchella, Tryblionella panduriformis and Trachyneis australis below 1 m to Campylodiscus clypeus above 1 m. There were very few records of species that prefer fresh to oligosaline waters in those samples, with the proportion of marine taxa ranging from 60 to 100% of valves.
Above 50 cm, there is an abrupt decline in the proportion of marine species from 80% at 50 cm to 12% at 45 cm. The main coastal species that persisted into this zone was Campylodiscus clypeus, a species that has been recorded in inland saline systems (Gell 1997). These species were mostly replaced by mesosaline taxa, principally Aneumastus tuscula and Amphora spp., whereas euryhaline species Cocconeis placentula, Cyclotella meneghiniana and small species within the Fragilariaceae (e.g. Staurosira venter) also were common. The upper part of this zone marked the arrival of freshwater taxa, notably Frustulia rhomboides and the epiphytic Gomphonema spp. No marine species were recorded in the uppermost sample of this zone.
This zone is marked by the decline in mesosaline species and their replacement by fresh to oligosaline, and aerophilous forms. No marine taxa were recorded in this zone. Frustulia rhomboides and Gomphonema spp. continue to be well represented into this zone, with the latter decreasing towards the surface. Most notably, the often-acidophilous genera Eunotia spp. and Pinnularia spp. enter the record towards the top of SFN-2 and dominate in this zone. Other taxa occur more sporadically, with Planothidium delicatulum, Achnanthidium minutissimum and, towards the surface, Tabularia fasciculata becoming common. The latter, along with Nitzschia spp. (incl. N. filiformis) and Cyclotella meneghiniana, mark an increase in eutrophication, particularly above 10 cm.
There are several eurytopic (widely tolerant) species present through the record, including Cocconeis placentula and small species in the genera Staurosira and Staurosirella (ex-Fragilaria). The latter typically predominate in disturbed situations where there is a loss of habitat suitable for other species (Fluin et al. 2007).
The diatom analysis of the 20 samples from Core SFC yielded 114 taxa. The C2 stratigraphic diagram of the diatom assemblages across the 49 cm of the SFC sediment core is provided as Fig. 7.
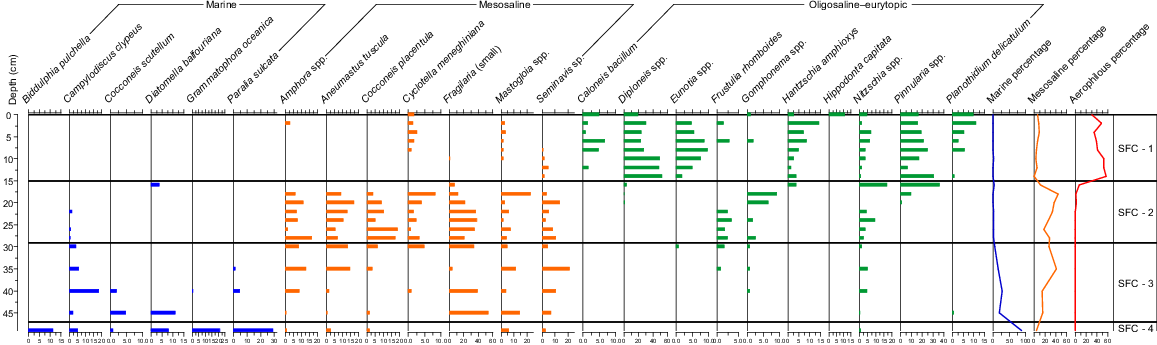
The lowermost sample was dominated by the marine forms Biddulphia pulchella, Grammatophora oceanica and Paralia sulcata, along with Diatomella balfouriana and Cocconeis scutellum. Ardossonia robusta, Diploneis didyma, Mastogloia baldjikiana, Navicula yarrensis and Rhaphoneis australis were also recorded in this lowermost sample. Marine forms represented over 80% of the total.
This zone is marked by the loss of marine forms and the persistence of the estuarine diatoms Cocconeis scutellum, Diatomella balfouriana and then Campylodicsus clypeus. The estuarine forms were largely lost to the record by 30 cm, with only Campylodiscus clypeus persisting to the top of the zone. Estuarine forms were increasingly replaced by mesosaline types, including Amphora spp., Aneumastus tuscula, Mastogloia spp. and Seminavis sp.
From 30 to 16 cm, the mesosaline types that entered in Zone SFC-3 dominated the assemblage, along with Aneumastus tuscula, Cocconeis placentula, Cyclotella meneghiniana and Seminavis sp. Frustulia rhomboides entered the record from before this zone. Species common in freshwater, including those in the genera Gomphonema and Pinnularia, increased from 20 cm. Small species in the Fragilariaceae were common from 45 cm, declined through the next zone and were largely absent in SFC-1. The epiphytes Cocconeis placentula and Gomphonema spp. were common.
By 12 cm, Eunotia spp. and Pinnularia spp. dominated. Aerophilous species, including Caloneis sp., Diploneis spp., and Hantzschia amphioxys, increased dramatically from 16 cm to become common from 14 cm. Planothidium delicatulum increased from 8 cm and Hippodonta capitata was common at the surface.
The diatom analysis of the 20 samples from Core SFS yielded 114 taxa. The C2 stratigraphic diagram of the diatom assemblages across the 64 cm of the SFS sediment core is provided as Fig. 8.
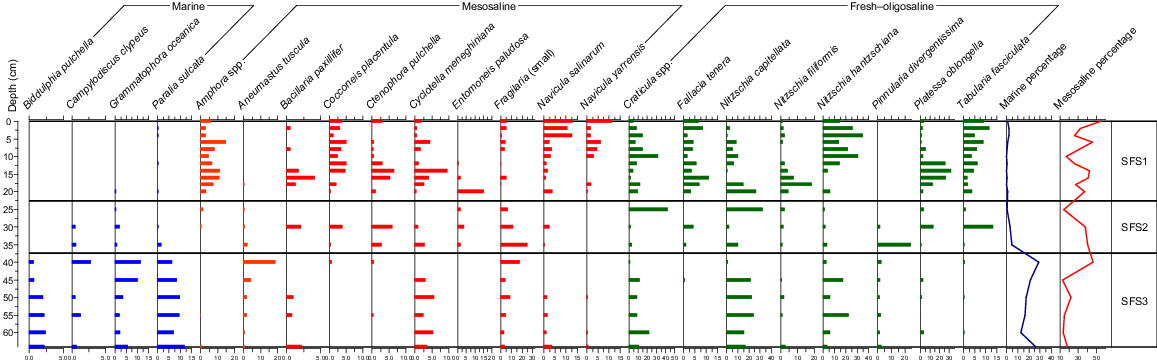
Like in Cores SFN and SFC, the basal sediments (here 64–40 cm) were dominated by the marine forms Biddulphia pulchella, Grammatophora oceanica and Paralia sulcata, along with Campylodiscus clypeus. Ardissonia robusta, Cocconeis scutellum, Lyrella lyra, Mastogloia baldjikiana and Navicula yarrensis were also recorded in Zone SFS-3. The marine sum was lower than for the other cores, at ~30%. Here, the marine dominance was shared with mesosaline and oligosaline forms, including Aneumastus tuscula, Cyclotella meneghiniana, Craticula spp. and Nitzschia spp.
The assemblage between 35 and 25 cm was represented by a variable mix of mesosaline and oligosaline taxa. Species that were common in only one or few samples included Bacillaria paxillifer, Craticula spp., Ctenophora pulchella, Nitzschia capitellata, Pinnularia divergentissima and Tabularia fasciculata. Small forms of the Fragilariaceae became common from the top of SFS-3 and declined through this zone.
Above 25 cm, the mesosaline Amphora spp. increased, along with indictors of oligosaline to fresh conditions, including Tabularia fasciculata. Other species commonly observed through this zone included Cocconeis placentula, Navicula salinarum, Craticula spp. and Platessa oblongella. Several common taxa recorded in Zone SFS-1 are known as aerophilous types (e.g. Fallacia tenera), thriving in mudflat habitats. Nutrient indicators (e.g. Nitzschia spp.) are common in SFS-1, but several were evident throughout the core (e.g. Cyclotella meneghiniana).
Wetland diatom summary
The longer core (SFN) provided evidence for the dominance of marine species over c. 6800 years. These species are evident in the lower sections of the shorter cores, showing that the marine forms were persistent throughout the wetland in pre-European times. The three cores showed that most species (e.g. Ardissonia robusta, Biddulphia pulchella, Grammatophora oceanica, Paralia sulcata) were common throughout the site. Also common to all cores was their gradual replacement by Campylodiscus clypeus and then a range of mesosaline species such as Aneumastus tuscula and the euryhaline Cocconeis placentula. Amphora spp. increased in all cores, but arrived later in Core SFS. The rise in fresh, acidophilous and epiphytic taxa are common throughout, but again less so in the upper parts of Core SFS. Aerophilous species and nutrient indicators became common in the upper sections of all three cores.
Pollen
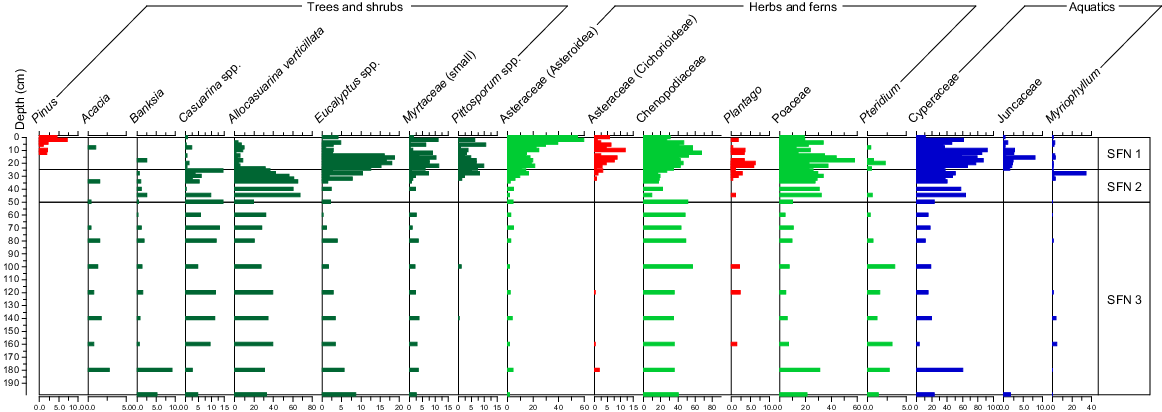
The pollen analysis of the 30 samples from Core SFN yielded 81 pollen and spore types. The C2 stratigraphic diagram of the pollen assemblages across the 200 cm of the SFN sediment core is provided as Fig. 5.
The most commonly-observed dryland pollen types from the SFN core were Banksia sp., Casuarinaceae (sheoak), Chenopodiaceae (saltbush), Eucalyptus spp. and small Myrtaceae (likely to be Leptospermum and Melaleuca spp.). The most common herbs were aquatic plants represented by Poaceae (grass, incl. Phragmites), Plantago (plantain), Cyperaceae (sedge), Pteridium (bracken) and Myriophyllum (milfoil). Pinus, plantain and Asteraceae (Cichorioidea; thistles etc.) were the common exotic forms (Fig. 5).
From 199 to 50 cm, the most common pollen forms were Allocasuarina (local species include A. verticillata and Allocasuarina pusilla–paludosa) and Chenopodiaceae, with Eucalyptus and Poaceae being less common. Cyperaceae were moderately common, along with Myriophyllum. Acacia and Pteridium were frequently recorded.
From 50 to 25 cm, Allocasuarina remained common, whereas Banksia and saltbush declined. Eucalyptus spp., Myrtaceae, Pittosporum spp. and Asteraceae increased from the top of this zone. Myriophyllum was common at 28 cm.
From 25 cm, Allocasuarina and Banksia declined and Eucalyptus, small Myrtaceae, Asteraceae, Pittosporum increased along with Cyperaceae and Juncaceae (rushes). Poaceae numbers remained high. Pinus pollen entered the record at 12 cm. Chenopodiaceae numbers increased to levels equal to those below 50 cm.
Discussion
Diatom assemblages can be used to infer past water quality and habitat availability on the basis of the known ecology of living species. Because many diatom species have a cosmopolitan distribution, their known ecological preferences can be derived from works published elsewhere (e.g. Witkowski et al. 2000). In Australia, many diatom data bases have been established (e.g. Gell 1997; Philibert et al. 2006; Chessman et al. 2007) that allow for conditions to be deduced from inland waterways. Although there has been work on the modern and fossil diatom assemblages from Australian estuaries (Taffs et al. 2017b), relatively little work has been undertaken on the diatoms of Australian marine environments. Here, the observations of Foged (1978) during a tour of eastern Australia in 1966, combined with the global treatise of Witkowski et al. (2000), form a basis for understanding the ecological preferences of the thalassic (marine) species observed in the lower sediments of the three cores.
The distinct diatom assemblages allow for a confident interpretation of the environmental history to be made. Clearly, Seaford Swamp was, for millennia prior to European settlement, an open tidal system with mesosaline to eusaline waters. The high numbers of the estuarine plankton form Paralia sulcata suggest that the inflows of sea water to Seaford Swamp were substantial. The main marine taxa were observed at the base of all three cores, suggesting that, although being more variable in SFS, tidal conditions existed throughout the Seaford Swamp for millennia. The existence of a saltmarsh environment is inferred from the abundance of saltbush pollen and low numbers of aquatic plants such as grass, sedge and milfoil. Away from the swamp, the vegetation was dominated by sheoak and Banksia.
The 14C dating suggests that the wetland was established soon after sea levels stabilised at 7000 years ago. The period from 3500 to 2200 years ago was dry across western Victoria, relative to the mid-Holocene (Jones 2001) and probably also dry in the Port Philip Bay region. Dry conditions are implicated in a hiatus, from 2800 to 1000 years BP, in the deposition of delta deposits at the mouth of the Yarra River suggesting low sea levels in Port Phillip Bay (Holdgate and Norvick 2017). The loss of thalassic diatoms by 45 cm may have resulted from the receding Bay at 2800 years BP. However, such a timeline would invoke extremely low sedimentation rates from 45 to 30 cm, spanning the resumption of sea levels at 1000 years to the arrival of Europeans. In lieu of more detailed dating, the abrupt change at 45 cm is considered here to have occurred in early European settlement. So, the stability in the diatom assemblage at SFN between 200 and 45 cm suggests that the Bunurong people witnessed relatively stable wetland conditions leading into European settlement.
In the recent past, the hydrology of the system changed progressively from brackish-saline to mostly fresh. Although this may occur with gradual geomorphic changes at the coast, the abrupt changes recorded here suggest that this was probably due to urban and rural development, land drainage and hydrological changes associated with the construction of the Patterson Cut, a 10-m-wide drainage channel, in 1879 (Ross n.d.). The 210Pb dating places this change at before 1943, and possibly as early as the late 19th Century. This has resulted in the blocking of sea water inflow to the wetland. The replacement of marine species with brackish-saline suggests the wetland lost tidal input but continued to be influenced by the residual salts for some time. From the late 19th Century, much of the catchment was cleared for agriculture and there has been substantial urban development. This has increased fresh catchment and urban runoff that has been diverted into the system as part of the development of the region. In Cores SFN and SFC, this has seen the complete replacement of the original diatom flora with fresh-to-oligosaline taxa, many with affinities for organic acids. This pattern is less clear in Core SFS, which was more variable and transitioned to mesosaline, eutrophic and aerophilous species. The pollen record shows a decline in saltbush (albeit temporary) and a rise in aquatic plants such as grass (probably reed), rush, milfoil and sedge that typically prefer fresher conditions. This appears to be most evident in the north of the swamp as reflected in the greater abundance of epiphytic diatoms (Cocconeis spp., Gomphonema spp.) and those preferring acid–neutral conditions (Eunotia spp., Pinnularia spp.) consistent with organic accumulation. The transition was abrupt in the north and more gradual in the south, implicating catchment and urban drainage entering the system from the north as the cause of the change. The changes in the diatom flora also suggest post-European increases in nutrient and sediment flux, again probably owing to changes in land uses in the catchment. Settlement has seen the decline in Banksia and Allocasuarina and increase in Eucalyptus.
The shift post-European settlement from a saline state to fresh is not unusual with coastal lagoons elsewhere in southern and eastern Australia, such as in northern and central New South Wales (Tibby et al. 2008; Leyden et al. 2011), south-eastern South Australia (Haynes et al. 2007) and the lower Barwon River in central Victoria (Reeves et al. 2016; Gell et al. 2022) experiencing considerable freshening.
The terrestrial pollen shows a decline in Allocasuarina and Banksia. Such a decline has been documented elsewhere on the Mornington Peninsula (Calder 1975), owing to their vulnerability to disease and preference as a source of fuel and timber. Eucalyptus has been advantaged by this disturbance, as has Pittosporum, which may have been widely planted or invaded unaided (e.g. P. undulatum). Reed (Phragmites australis) is intolerant of high salinity (Hart et al. 1991) and so has been advantaged by the increase in fresh water. This poses a management problem because it is now dominating other aquatic habitats.
Seaford Swamp has been identified as a system that has been affected by human development and hydrological management over recent centuries. This record attests to that change and identifies the natural state of the system enjoyed by the original inhabitants prior to this intensive disturbance. The long-term state was one strongly influenced by tidal inflows and was probably a saline marsh. Most of this habitat has been lost and the system is now largely fresh-oligosaline, allowing for the colonisation of aquatic plants typical of freshwater such as reed. The growth of reed now poses an issue for the management authority that now invests resources in controlling its growth.
Seaford Swamp was listed as an internationally important wetland under the Ramsar Convention in 2001 where it was broadly recognised as a predominantly freshwater marsh. The identification here, of a prior estuarine state, and the timing and nature of its transition to fresh water, is clear evidence that the ecological character of the wetland had changed before being listed as internationally important. That is, the ecological character (or condition) described when it was listed does not represent its natural ‘ecological character’, but rather it represents a character that was the result of land and water use and development following European settlement of the area.
The extended record of change in the Seaford Swamp identified in this investigation ‘opens up’ the suite of target states that management may consider. This could include the present freshwater state or a more saline one that might reduce the proliferation of reeds and reflect that witnessed by Bunurong People in the past. The pre-European estuarine state identified here also provides a future option under scenarios of inundation under sea-level rise (DEECA 2024). Whichever, it remains that management needs to also consider the listing criteria that underpin its original nomination as well as the obligation to maintain the ecological character, which includes the key features of the biodiversity, the ecological processes that sustain the wetlands, and the ecosystem services that it provides for people.
Data availability
The original data are available from the corresponding author subject to the consent of the funding body.
Declaration of funding
This project was funded by the Frankston City Council under a contract to Diatoma.
Acknowledgements
We acknowledge the Bunurong People on whose Country this research was conducted. The interest in wetland past condition shown by David Fairbridge and others at Frankston City Council is recognised and welcomed. The team appreciated the assistance in the field from Matt Green and Alex Milicic. Sabika Maizma (ANSTO) managed the 210PB analyses and Tim Barrows oversaw the 14C date provided by The University of New South Wales. Max Finlayson provided comments on an earlier draft. Frankston City Council drafted Fig. 1.
References
Battarbee RW, Jones VJ, Flower RJ, Cameron NG, Bennion H, Carvalho L, Juggins S (2001) Diatoms. In ‘Tracking environmental change using lake sediments. Vol. 3: terrestrial, algal and siliceous indicators’. (Eds EF Stoermer, HJB Birks, WM Last) pp. 155–202. (Kluwer Academic Publishers: Dordrecht, Netherlands)
Bickford S, Gell P, Hancock GJ (2008) Wetland and terrestrial vegetation change since European settlement on the Fleurieu Peninsula, South Australia. The Holocene 18, 425-436.
| Crossref | Google Scholar |
Chessman BC, Bate N, Gell PA, Newall P (2007) A diatom species index for bioassessment of Australian rivers. Marine and Freshwater Research 58(6), 542-557.
| Crossref | Google Scholar |
Cook PLM, Jennings M, Holland DP, Beardall J, Briles C, Zawadzki A, Doan P, Mills K, Gell P (2016) Blooms of cyanobacteria in a temperate Australian lagoon system post and prior to European settlement. Biogeosciences 13, 3677-3686.
| Crossref | Google Scholar |
Department of Energy, Environment and Climate Action (2024) Port Phillip Bay Coastal Hazard Assessment – Summary #4. Inundation hazard assessment. (DEECA: Melbourne, Vic., Australia ) Available at https://www.marineandcoasts.vic.gov.au/__data/assets/pdf_file/0038/693398/PPBCHA-communications-summary4-inundation-hazard-assessment.pdf
Department of Sustainability and Environment (2012) Description of the ecological character of the Edithvale–Seaford wetlands Ramsar site. (DSE, Victorian Government: Melbourne, Vic., Australia ) Available at https://www.water.vic.gov.au/__data/assets/pdf_file/0028/672805/Edithvale_Seaford_Wetlands_Ecological_Character_Description.pdf
Dubois N, Saulnier-Talbot É, Mills K, Gell P, Battarbee R, Bennion H, Chawchai S, Dong X, Francus P, Flower R, Gomes DF, Gregory-Eaves I, Humane S, Kattel G, Jenny JP, Langdon P, Massaferro J, McGowan S, Mikomägi A, Ngoc NTM, Ratnayake AS, Reid M, Rose N, Saros J, Schillereff D, Tolotti M, Valero-Garcés B (2018) First human impacts and responses of aquatic systems: a review of palaeolimnological records from around the world. The Anthropocene Review 5, 28-68.
| Crossref | Google Scholar |
Fluin J, Gell P, Haynes D, Tibby J, Hancock G (2007) Paleolimnological evidence for the independent evolution of neighbouring terminal lakes, the Murray Darling Basin, Australia. Hydrobiologia 591, 117-134.
| Crossref | Google Scholar |
Gasse F, Juggins S, Khelifa LB (1995) Diatom-based transfer functions for inferring past hydrochemical characteristics of African lakes. Palaeogeography, Palaeoclimatology, Palaeoecology 117, 31-54.
| Crossref | Google Scholar |
Gell PA (1997) The development of a diatom database for inferring lake salinity, Western Victoria, Australia: towards a quantitative approach for reconstructing past climates. Australian Journal of Botany 45, 389-423.
| Crossref | Google Scholar |
Gell P, Tibby J, Fluin J, Leahy P, Reid M, Adamson K, Bulpin S, MacGregor A, Wallbrink P, Hancock G, Walsh B (2005a) Accessing limnological change and variability using fossil diatom assemblages, south-east Australia. River Research and Applications 21, 257-269.
| Crossref | Google Scholar |
Gell PA, Bulpin S, Wallbrink P, Hancock G, Bickford S (2005b) Tareena Billabong – a palaeolimnological history of an ever-changing wetland, Chowilla Floodplain, lower Murray–Darling Basin. Marine and Freshwater Research 56, 441-456.
| Crossref | Google Scholar |
Gosling W, Lawson I, Roucoux K, Boreham S (2022) Pollen Preparation Protocol for Fossil Pollen: fossil pollen preparation protocol (V. 1-1). (Cambridge University: Cambridge, UK) Available at https://www.geog.cam.ac.uk/facilities/laboratories/techniques/pollen/
Hall RI, Smol JP (1992) A weighted-averaging regression and calibration model for inferring total phosphorus concentration from diatoms in British Columbia (Canada) lakes. Freshwater Biology 27, 417-434.
| Crossref | Google Scholar |
Hart BT, Bailey P, Edwards R, Hortle K, James K, McMahon A, Meredith C, Swadling K (1991) A review of the salt sensitivity of the Australian freshwater biota. Hydrobiologia 210, 105-144.
| Crossref | Google Scholar |
Haynes D, Gell P, Tibby J, Hancock G, Goonan P (2007) Against the tide: the freshening of naturally saline coastal lakes, southeastern South Australia. Hydrobiologia 591, 165-183.
| Crossref | Google Scholar |
Head L (1987) The Holocene prehistory of a coastal wetland system: discovery Bay, southeastern Australia. Human Ecology 15, 435-462.
| Crossref | Google Scholar |
Holdgate G, Norvick M (2017) Geological evolution of the Holocene Yarra Delta and its relationship with Port Phillip Bay. Australian Journal of Earth Sciences 64, 301-318.
| Crossref | Google Scholar |
Jowsey PC (1966) An improved peat sampler. New Phytologist 65(2), 245-248.
| Crossref | Google Scholar |
Kumar R, Finlayson CM, Pattnaik AK (2020a) Ecological characterization of Chilika: defining strategies and management needs for wise use. In ‘Ecology, conservation, and restoration of Chilika Lagoon, India’. (Eds CM Finlayson, G Rastogi, D Mishra, A Pattnaik) pp. 23–61. (Springer: Dordrecht, Netherlands)
Leyden E, Tibby J, García A, Zawadzki A (2011) Unexpected post-settlement freshening and increase in charophytes in Bombah Broadwater (Myall Lakes, New South Wales, Australia). Journal of Paleolimnology 46, 637-647.
| Crossref | Google Scholar |
MacGregor AJ, Gell PA, Wallbrink PJ, Hancock G (2005) Natural and post-European settlement variability in water quality of the lower Snowy River floodplain, eastern Victoria, Australia. River Research and Applications 21, 201-213.
| Crossref | Google Scholar |
Manzano S, Julier ACM, Dirk CJ, Razafimanantsoa AHI, Samuels I, Petersen H, Gell P, Hoffman MT, Gillson L (2020) Using the past to manage the future: the role of palaeoecological and long-term data in ecological restoration. Restoration Ecology 28(6), 1335-1342.
| Crossref | Google Scholar |
Pandey S, Scharf BW, Mohanti M (2014) Palynological studies on mangrove ecosystem of the Chilka Lagoon, east coast of India during the last 4165 yrs BP. Quaternary International 325, 126-135.
| Crossref | Google Scholar |
Philibert A, Gell P, Newall P, Chessman B, Bate N (2006) Development of diatom-based tools for assessing stream water quality in south-eastern Australia: assessment of environmental transfer functions. Hydrobiologia 572, 103-114.
| Crossref | Google Scholar |
Pittock J, Finlayson CM, Gardner A, McKay C (2010) Changing character: the Ramsar Convention on wetlands and climate change in the Murray–Darling Basin, Australia. Environmental and Planning Law Journal 27(6), 401-425.
| Google Scholar |
Pritchard D (2022) The ‘ecological character’ of wetlands: a foundational concept in the Ramsar Convention, yet still cause for debate 50 years later. Marine and Freshwater Research 73(9–10), 1127-1133.
| Crossref | Google Scholar |
Reeves JM, Gell PA, Reichman SM, Trewarn AJ, Zawadzki A (2016) Industrial past, urban future: using palaeo-studies to determine the industrial legacy of the Barwon estuary, Victoria, Australia. Marine and Freshwater Research 67, 837-849.
| Crossref | Google Scholar |
Reid MA, Tibby JC, Penny D, Gell PA (1995) The use of diatoms to assess past and present water quality. Australian Journal of Ecology 20, 57-64.
| Crossref | Google Scholar |
Reid MA, Sayer CD, Kershaw AP, Heijnis H (2007) Palaeolimnological evidence for submerged plant loss in a floodplain lake associated with accelerated catchment soil erosion (Murray River, Australia). Journal of Paleolimnology 38, 191-208.
| Crossref | Google Scholar |
Reimer PJ, Reimer RW (2001) A marine reservoir correction database and on-line interface. Radiocarbon 43, 461-463.
| Crossref | Google Scholar |
Ross C (n.d.) Carrum – a brief local history. Available at https://sites.google.com/site/originalcarrumcowboys/history [Verified 19 April 2024]
Stroud DA, Davidson NC, Finlayson CM, Gardner RC (2022) Development of the text of the Ramsar Convention: 1965–1971. Marine and Freshwater Research 73, 1107-1126.
| Crossref | Google Scholar |
Taffs KH, Saunders KM, Weckstrom K, Gell PA, Skilbeck CG (2017a) Introduction to the application of palaeoecological techniques in estuaries. In ‘Applications of paleoenvironmental techniques in estuarine studies, developments in paleoenvironmental research’. (Eds K Weckstrom, P Gell, K Saunders, G Skilbeck) pp. 1–6. (Springer: Dordrecht, Netherlands)
Taffs KH, Saunders KM, Logan B (2017b) Diatoms as indicators of environmental change in estuaries. In ‘Applications of paleoenvironmental techniques in estuarine studies, developments in paleoenvironmental research’. (Eds K Weckstrom, P Gell, K Saunders, G Skilbeck) pp. 277–294. (Springer: Dordrecht, Netherlands)
Tibby J, Lane MB, Gell PA (2008) Local knowledge and environmental management: a cautionary tale from Lake Ainsworth, New South Wales, Australia. Environmental Conservation 34, 334-341.
| Crossref | Google Scholar |