

A continuum of stomatal responses to water deficits among 17 wine grape cultivars (Vitis vinifera)
Alexander D. Levin A B E , Larry E. Williams C D and Mark A. Matthews C
A B E , Larry E. Williams C D and Mark A. Matthews C
A Department of Horticulture, Oregon State University, Corvallis, OR 97331, USA.
B Southern Oregon Research and Extension Center, 569 Hanley Road., Central Point, OR 97502, USA.
C Department of Viticulture and Enology, University of California, Davis, CA 95616, USA.
D Kearney Agricultural Research and Extension Center, 9240 S. Riverbend Avenue., Parlier, CA 93648, USA.
E Corresponding author. Email: alexander.levin@oregonstate.edu.
Functional Plant Biology 47(1) 11-25 https://doi.org/10.1071/FP19073
Submitted: 15 March 2019 Accepted: 19 July 2019 Published: 16 October 2019
Journal Compilation © CSIRO 2019 Open Access CC BY-NC-ND
Abstract
Vitis vinifera L. cultivars have been previously classified as isohydric, near-isohydric, anisohydric or isohydrodynamic, depending on the study. To test the hypothesis that V. vinifera cultivars’ stomatal behaviour can be separated into distinct classes, 17 cultivars grown in a replicated field trial were subjected to three irrigation treatments to manipulate vine water status across multiple years. Predawn (ΨPD) and midday (Ψl) leaf water potential and midday stomatal conductance (gs) were measured regularly throughout several seasons. The relationship of gs to Ψl was best modelled as a sigmoidal function and maximum stomatal conductance (gmax), water status at the onset of stomatal closure (Ψl95), sensitivity of closure (gsensitivity) and water status at the end of closure (Ψl25) were compared. There were no significant differences in gmax among cultivars. Cultivar-specific responses of gs to Ψl were broadly distributed along a continuum based on the relationship between Ψl95 and gsensitivity. Season-long cultivar mean Ψl values were positively related to Ψl25. In general, cultivars responded similarly to one another at high and low water status, but their stomatal behaviour differed at moderate water deficits. The results show that V. vinifera cultivars possess both iso- and anisohydric stomatal behaviours that depend on the intensity of water deficits, and call into question previous classifications assuming a single behaviour.
Additional keywords: genotypic variation, grapevine physiology, stomatal conductance, water potential.
Introduction
Increasing demand for fresh water in the face of finite resources is intensifying pressure to improve water use efficiency in agriculture in general, and in crop production in particular. Because regulation of stomatal aperture plays a major role in plant control of water loss in dry environments, it has been the focus of intensive research directed towards improved drought tolerance and water use efficiency. For example, reduced maximum transpiration rate (Sinclair et al. 2005) and early stomatal closure (Martin-St. Paul et al. 2017) have been proposed as traits of drought avoidance. In both cases, the putative benefit in yield stability arises from postponing dehydration. Indeed, the sensitivity of stomata to water deficits could be viewed as a measure of drought avoidance (Jones 1974; Bates and Hall 1981) and is sometimes described in terms of ‘isohydric’ vs ‘anisohydric’ behaviour (Tardieu and Simonneau 1998).
In addition to species variation, it has long been known that genetic variation in stomatal response to water deficits is present within crop species (e.g. in sorghum, Henzell et al. 1976; e.g. in rice, Dingkuhn et al. 1989). Schultz (2003) provided some evidence of such genetic variation within grapevine, Vitis vinifera L., but there have since been contradictory reports of cultivar-specific responses (Chaves et al. 2010; Costa et al. 2012). Ambiguity over the precise definition of what constitutes an iso- vs anisohydric response (Hochberg et al. 2018) has caused difficulties in categorisation of either cultivars within a species (Lavoie-Lamoureux et al. 2017) or across many species (Klein 2014; Martinez-Vilalta and Garcia-Forner 2017). However, in general, isohydric species/cultivars will maintain water status similar between droughted and well-watered plants (Tardieu and Simonneau 1998) or that a measure of plant water status (such as leaf water potential) will not decrease below a minimum value on a diurnal or seasonal basis (Schultz 2003). Such a response by a crop species may minimise the usefulness of assessing plant water status via the measurement of leaf water potential. Cifre et al. (2005) concluded that if grapevine cultivars respond to soil water deficits in an isohydric manner, then midday leaf water potential cannot be used in an irrigation management program or as one would suspect the validation of other means to assess plant water status (Williams 2017).
Based on observed responses of plant water relations and gas exchange, some grapevine cultivars such as Grenache, have been classified as isohydric (Schultz 2003; Cifre et al. 2005) whereas others, such as Syrah and Thompson Seedless have been classified as anisohydric (Schultz 2003; Soar et al. 2006b; Williams and Baeza 2007; Williams 2012). However, the discrepancy among cultivars is not entirely clear. This has led some researchers to modify the binary categorisation (i.e. isohydric vs anisohydric) and propose that cultivars may modulate their stomatal response (Domec and Johnson 2012) based on changes in soil moisture (Collins et al. 2010), or differences in leaf to air vapour pressure deficit (VPD) (Soar et al. 2006b). In addition, acclimation of stomatal response to water deficits has been shown in grapevine through a modification of leaf hydraulic conductivity and osmotic adjustment (Martorell et al. 2015; Hochberg et al. 2017a). Accordingly, alternative classifications such as ‘near-isohydric’ and ‘isohydrodynamic’ (a relatively constant difference between predawn (ΨPD) and midday leaf water potential (Ψl) throughout seasonal moisture cycles) have been offered to account for differences in stomatal response to drought among cultivars and the resulting effects on plant water status (Schultz 2003; Franks et al. 2007; Chaves et al. 2010; Shellie and Bowen 2014).
Putative cultivar differences in stomatal response would be important for water-conserving vineyard management strategies because gs is highly correlated with daily water use (Williams et al. 2012), and because moderate water deficits have positive effects on fruit quality (Roby et al. 2004). Furthermore, relative to vegetative growth, reproductive growth of irrigated grapevines is less affected when applied water amounts are reduced by up to 40% of crop evapotranspiration (Williams et al. 2010a, 2010b). As such, some water deficit is beneficial in production of horticultural crops like wine grapes in contrast to production of agronomic crops like rice or sorghum.
The relationship between gs and Ψl in grapevine is affected by numerous factors; predominate among them are rootstock and cultivar, accounting for 19 and 16% of the variability in that relationship, respectively (Lavoie-Lamoureux et al. 2017). Presumably, a large proportion of the remaining variation in response would be accounted for by plant-environment interactions (Hochberg et al. 2018). As mentioned above, soil moisture, VPD and plant hydraulic properties have all been shown to influence the relationship between gs and Ψl across grapevine cultivars. However, since the initial classification by Schultz (2003), and numerous subsequent attempts by others – as reviewed by Chaves et al. (2010) – there have been no studies that have directly compared a large set of cultivars grown under the same environment conditions (soil and atmosphere) grafted onto the same rootstock in a replicated field trial.
The goal of this study was 2-fold: to (1) determine if numerous V. vinifera cultivars separate into distinct iso- or anisohydric classes in response to soil water deficits from measurements of predawn (ΨPD) and midday leaf water potential (Ψl); and (2) explore the genetic variation in the relationship of gs to Ψl among these cultivars over several seasons. To accomplish this, the study was designed to maximise the effect of cultivar on the relationships in question by controlling for numerous factors that have all been shown to be influential such as rootstock, environment, instrument operator, and sampling methodology. Several grapevine cultivars evaluated in this study have been previously classified as being isohydric (or near-isohydric) – e.g. Cabernet Sauvignon, Grenache, Tempranillo and Touriga Nacional – while others have been classified as anisohydric – e.g. Cabernet Sauvignon, Grenache, Montepulciano, Syrah (syn. Shiraz) and Touriga Nacional (Schultz 2003; Chouzouri and Schultz 2005; Soar et al. 2006b; Williams and Baeza 2007; Chaves et al. 2010; Lovisolo et al. 2010; Rodrigues et al. 2012). Finally, an additional 11, red wine grape cultivars were evaluated representing a broad range of plant material grown in warmer viticultural regions around the world.
Materials and methods
Vineyard site, design, and plant material
The experimental site used in this study was an existing variety trial that was planted with 1103 Paulsen (1103P) (Vitis berlandieri Planch. × Vitis rupestris Scheele) rootstock in June 2003 at the University of California Kearney Agricultural Research and Extension (KARE) Center near Parlier, California (36°48′N, 119°30′W). The soil was a Hanford fine sandy loam. The total vineyard area was ~0.53 ha. Vine and row spacing was 1.83 and 3.05 m respectively (5.58 m2 per vine).
The trellis system consisted of a 0.3 m cross-arm atop a stake, 1.7 m above the vineyard floor. Fixed catch wires were positioned at each end of the cross-arms. Vines were trained to bilateral cordons on a single fruiting wire at a height of 1.2 m, and were pruned to spurs. In early spring, shoots were thinned to a density of ~15 shoots per meter of row. The vineyard was drip-irrigated with two, 2 L h–1 emitters per vine placed 0.5 m on either side of the trunk. The dripline was attached to a wire installed 0.5 m above the vineyard floor.
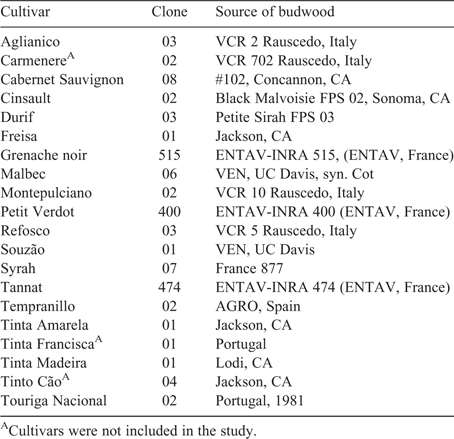
The vineyard layout consisted of 20 rows with 48 vines per row. In May 2004, 20 different Vitis vinifera L. cultivars were field-budded onto the rootstock. Foundation Plant Services (University of California, Davis, CA, USA) supplied budwood for the scion varieties. The cultivars, clonal selection, and sources of budwood are given in Table 1. Carmenere, Tinta Francisca and Tinta Cão were not included in this study due to foliar symptoms of viral infection.
|
Irrigation treatments
Three irrigation treatments were imposed beginning in the 2012 growing season as detailed below.
-
Late deficit (LD) – irrigated at 100% of estimated ETc from berry set until the onset of ripening (veraison); no applied water from veraison until harvest.
-
Sustained deficit (SD) – irrigated at 50% of estimated ETc throughout the entire growing season.
-
Early deficit (ED) – no applied water from berry set until veraison; irrigated at 50% of estimated ETc from veraison until harvest.
Prior to berry set, all vines were irrigated at 100% of estimated ETc to maintain midday leaf water potential (Ψl) at or above –1.0 MPa. After berry set, irrigation treatments were imposed once Ψl reached the threshold value of –1.0 MPa (averaged across cultivars). Drip emitters were plugged or unplugged to impose the irrigation treatments, and inline water meters were used to quantify applied water amounts. Veraison timing was averaged across replicates of a cultivar-treatment combination (e.g. separately quantified for Syrah-ED vs Syrah-LD). Therefore, treatment transition (e.g. in the ED and LD treatments) occurred over a period of several weeks across cultivars. After harvest, all vines were irrigated at 50% of estimated ETc until leaf fall.
Vineyard ETc was estimated using the following equation: ETc = ETo × Kc, where ETo is reference ET and Kc is the crop coefficient. Prior to the initiation of the experiment, seasonal crop coefficients were developed in this vineyard across seven growing seasons during which vines were kept well watered, so as to not inhibit canopy development (LE Williams, unpubl. data). They were determined by measuring the shade cast on the ground beneath the canopy, and then using the relationship between the percentage of canopy shaded area and Kc according to Williams and Ayars (2005). Finally, a relationship between Kc and accumulated growing degree-days (GDD) was established. During the experiment, evolution of the Kc over the course of the growing season was calculated using accumulated GDD from average budbreak. GDD data were downloaded from the University of California Statewide Integrated Pest Management Project’s website (www.ipm.ucdavis.edu, accessed 12 September 2019). ETo and GDD data were obtained from the CIMIS weather station at the KARE Center (Parlier #39, California Irrigation Management Information System). GDD were calculated using the single sine method with a lower threshold of 10°C.
Vine water status measurements
Vine water status (water potential; Ψ) was measured according to (Williams and Araujo 2002) using a pressure chamber (Model 1000, PMS Instrument Co.). Specifically, predawn water potential (ΨPD) measurements began at ~0400 h Pacific daylight time (PDT) and were completed before sunrise, whereas midday leaf water potential (Ψl) measurements were generally taken between 12:30 and 13:30 hours PDT. ΨPD was only measured in 2012. Leaves chosen at predawn and at midday were fully expanded, mature leaves – and at midday exposed to direct solar radiation. Leaf blades chosen across all times of day were covered with a plastic bag, quickly sealed, and petioles then cut within 1 to 2 s (s). Time between leaf excision and pressurisation was generally between 10 to 15 s.
Stomatal conductance (gs) was measured with a steady-state diffusion porometer (LI-1600, LI-COR Biosciences). The porometer was allowed to warm-up to ambient vineyard conditions (placed in the shade) at least 30 min (min) before the first measurement. The null point humidity was set to ambient after warm-up and before first measurements, and subsequently reset every 20 to 30 min. Measurements were recorded after steady-state conditions were reached (10–20 s). Care was taken to reduce sensor head exposure to direct sunlight in between measurements so as not to increase cuvette temperature. Sampled leaves were of same age and position in canopy as for Ψl measurements described above, thus ensuring light saturation.
During the 2012 growing season, the measurements of Ψl and gs were made by LE Williams for all cultivar/irrigation treatment replicates and required two days. In 2013, measurement techniques were refined by AD Levin (data not reported). During the last 2 years of the study, gs was measured on the same leaves used for the measurement of Ψl, just before leaf excision. Sample leaves were selected for gs, which was measured by LE Williams, and Ψl was measured by AD Levin both years, thus minimising potential operator error (Goldhamer and Fereres 2001). Measurements began at 12:00 hours and were completed by 14:00 hours those 2 years. A single leaf from each three-vine irrigation treatment replicate was measured and used for data analysis (n = 3–4). There were two, 16, and four measurement dates for the 2012, 2014 and 2015 growing seasons respectively.
Curve fitting stomatal response to vine water status
Stomatal response to Ψl was characterised with the following sigmoidal model:

where Ψl is midday leaf water potential, gmax is the asymptote that represents a theoretical maximum stomatal conductance, Ψl50 is the inflection point of the curve that represents the value of Ψl at which stomatal conductance is half of the maximum, and b is a scaling parameter that represents the transition width of the function. From the fitted curves, vine water status values were extracted at fixed values of gs relative to gmax (Ψl95, Ψl75, Ψl50 and Ψl25 represent the estimated Ψl at 95, 75, 50 and 25% of gmax respectively) for each cultivar, with Ψl95 assumed to be the threshold for the beginning of stomatal closure. The value of b was determined to be equal to (ΔΨl75–25 – 0.002) × 2.196–1 by linear regression, where ΔΨl75–25 is the difference in Ψl (MPa) between 75 and 25% of gmax (data not shown; R2 = 0.99, P < 0.001). The instantaneous slope of the fitted sigmoidal curve was calculated as the first derivative of the function at the inflection point and was taken as a measure of the maximum sensitivity of stomatal closure (gsensitivity; mol m–2 s–1 MPa–1).
Experimental design and data analysis
The experimental design was a randomised complete block with a strip-plot factorial treatment structure and four replications. Experimental blocks were assigned across the rows (i.e. 20 rows per block), and scion cultivars (main plot) were field-budded down the rows in a pattern of 12 vines per cultivar per block (i.e. four cultivars per row). The three irrigation treatments (strip plots) within each block were arranged down the rows in a line-source design (Williams et al. 2010a, 2010b), alternating sequentially among blocks from LD-SD-ED to ED-SD-LD such that vines in adjacent blocks received the same irrigation treatment. One non-irrigated vine was included among irrigation treatments within a block, such that an experimental unit consisted of the three remaining vines in the irrigation treatment.
Statistical analyses and graphics were done using R statistical software (R Core Team 2018). Linear regression, non-linear regression, and ANOVA were conducted using the functions lm, nls and anova, respectively, from R base package stats. For the analyses of Ψl and gs over time, a linear mixed-effects model was fit using the lmer function from the packages lme4 and lmerTest with cultivar, irrigation treatment, and sample date (and all interactions) as fixed factors, and block (and its interactions with the fixed factors) as a random factor (Bates et al. 2015; Kuznetsova et al. 2017). Calculation of least-squares means and multiple comparisons were conducted using the package lsmeans and the Tukey-Kramer adjustment method for multiplicity (Lenth 2016). Prior to non-linear regression analyses of gs in response to Ψl, gs values were pooled and means calculated across cultivars at each 0.1 MPa of Ψl. Smoothed cubic regression splines were fitted using the gam function from the package mgcv. (Wood 2017). Data were plotted using the package ggplot2 (Wickham 2016).
Results
Relationships between Ψl and ΨPD
Predawn (ΨPD) and midday leaf water potential (Ψl) were measured on vines in each irrigation treatment on numerous dates during the 2012 growing season. Although there were significant differences among irrigation treatments in both measures of vine water status, there were never significant differences among cultivars within an irrigation treatment. On 24 July 2012, just before changing the irrigation treatments that year, mean ΨPD (±s.d.) of vines in the ED treatment averaged across cultivars was –1.18 ± 0.13 MPa whereas those in the LD treatment averaged –0.13 ± 0.02 MPa. Averaged across cultivars, midday Ψl of the ED and LD treatments were –1.78 ± 0.02 and –0.98 ± 0.02 MPa, respectively, whereas that of the SD treatment averaged –1.28 MPa. Ambient temperature and VPD in the vineyard at the time of measurement (midday) was 34.9°C and 4.63 kPa respectively. Notably, mean Ψl of Cabernet Sauvignon, Grenache and Syrah on that day was –1.76 ± 0.04, –1.86 ± 0.07 and –1.80 ± 0.05 MPa, respectively. Stomatal conductance for the ED and LD treatments on that day averaged 0.089 and 0.568 mol m–2 s–1 respectively.
Midday Ψl of the LD treatment averaged across cultivars on 29 August (more than 5 weeks after irrigation had ceased for that treatment) was –1.76 ± 0.05 MPa. Midday Ψl of two vines of each cultivar (located at the end of the row on the east side of the vineyard) still being irrigated at 100% of estimated ETc averaged –0.92 ± 0.01 MPa. Midday Ψl of vines irrigated at 50% of ETc averaged –1.45 MPa. Ambient temperature and VPD at the time of measurement were 33.9°C and 4.11 kPa, respectively. On the last measurement date in 2012 (5 and 6 September) mean ΨPD and midday Ψl across cultivars were –0.73 ± 0.01 and –1.79 ± 0.02 MPa respectively. Midday Ψl of Cabernet Sauvignon, Grenache and Syrah on those dates were –1.84, –1.78 and –1.80 MPa respectively. The slope of the linear relationship between midday Ψl and ΨPD using data from Cabernet Sauvignon, Grenache and Syrah across the 2012 growing season was 0.784 with an R2 value of 0.73 (P = 0.001). Notably, the individual slopes of the three cultivars did not differ significantly from one another.
Ψl and gs seasonal responses to irrigation treatments
In general, seasonal responses of Ψl and gs to irrigation treatments were sensitive and consistent over the course of each growing season. The seasonal pattern of applied water amounts and responses of both water status variables (Ψl and gs) during 2014 is representative of those for the other growing seasons (Fig. 1). Prior to treatment imposition, all vines received 62 and 41 mm of applied water in 2014 and 2015 respectively. Following treatment imposition (12 and 11 May in 2014 and 2015 respectively), vines were irrigated at previously defined rates with applied water amounts illustrated by the curves of each treatment (Fig. 1a). Ψl and gs were significantly affected by the irrigation treatments, with three levels of water stress established before veraison, and two levels of water stress established after veraison in each year of the study (Fig. 1b, c). While Ψl and gs responded similarly to the irrigation treatments, there were small, but notable differences in each variable’s response to an irrigation treatment during each phenological period.

|
Before veraison, Ψl and gs responses were like one another in the ED and LD irrigation treatments, but they responded differently to the SD treatment. Both Ψl and gs values in LD vines remained high early in the season, with Ψl closely following the calculated non-stressed baseline Ψl value, and gs increasing slightly from berry set to a maximum near the end of June (just before veraison). Similarly, Ψl and gs values in ED vines both declined immediately following treatment imposition and were significantly lower compared with SD and LD at every measurement date from berry set until veraison (Fig. 1b, c). In contrast, while Ψl for SD vines declined after treatment imposition and was significantly different than that of LD vines from the third measurement date after treatment imposition, gs response was less sensitive compared with Ψl, and remained high in SD relative to LD for a longer period before veraison (Fig. 1b, c). Overall, preveraison gs was less sensitive to the SD irrigation treatment and remained high relative to that of LD vines despite a constantly declining Ψl until veraison.
Following veraison, Ψl and gs recovered in ED vines once irrigation water was reapplied, and both variables declined in LD vines when water was withheld (Fig. 1b, c). Ψl of LD vines was significantly lower than that of SD and ED beginning with the first measurement postveraison and remained so until harvest. Ψl and gs of ED vines recovered to values like those of SD vines and were not significantly different from SD for the remainder of the season. In general, postveraison Ψl was less stable week-to-week compared with postveraison gs, whereas preveraison Ψl was more stable week-to-week compared with preveraison gs.
The response of gs to Ψl: across and among cultivars
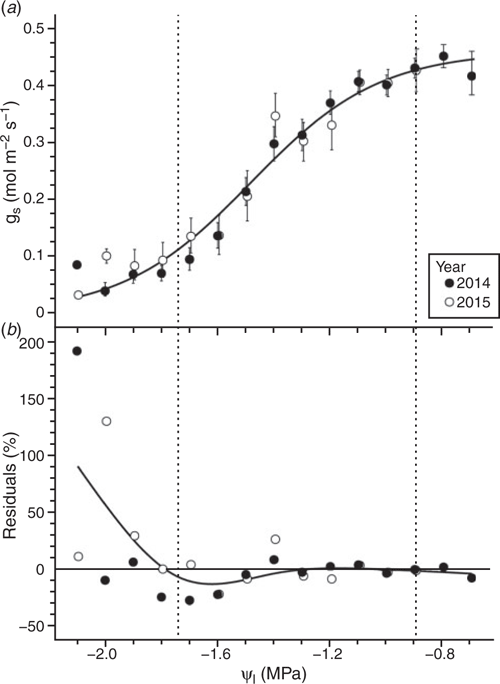
The response of gs to Ψl is summarised for all cultivars analysed together and individually in Figs 2 and 3 respectively. Because year was not a significant factor in either analysis – for all cultivars together or for all cultivars individually – data from the 2014 and 2015 growing seasons were analysed together. When all cultivars were analysed together in a composite analysis, Ψl95 and Ψl25 were estimated to be –0.89, and –1.74 MPa respectively (Fig. 2a). Across all cultivars, the response of gs to Ψl was well characterised by the sigmoidal function at Ψl values greater than Ψl25, with relative residuals generally varying within ~30% from the fitted curve, and in most cases within ~10%. Below Ψl25, the response of gs to Ψl diverged from the fitted curve with a corresponding increase in residuals to values greater than 30% of the fitted values (Fig. 2b).
|

|
The three-parameter logistic function was well fit to all cultivars individually as well (Fig. 3), with adjusted R2 values ranging from 0.72 (Durif) to 0.94 (Aglianico) (Table 2). From the fit sigmoidal functions, parameter estimates and modelled values at various levels of vine water status were compared across cultivars (Tables 2, 3). Excluding Syrah, the coefficient of variation (CV) in estimates of gmax was less than 7% across cultivars, ranging from 0.398 (Grenache) to 0.489 mol m–2 s–1 (Refosco). The estimate of gmax (±s.e.) for Syrah (0.595 ± 0.130 mol m–2 s–1) was 1.2 times larger than the next greatest estimate (Refosco; gmax = 0.489 ± 0.029 mol m–2 s–1), but with a large standard error (Table 2). Interestingly, the range and standard deviation (s.d.) of measured gmax values among cultivars – measured range = 0.151 mol m–2 s–1; measured s.d. = 0.0428 mol m–2 s–1 – was smaller than those of estimated gmax values among cultivars – estimated range = 0.196 mol m–2 s–1; estimated s.d. = 0.0461 mol m–2 s–1. In either case – when comparing estimated or measured values – there were no significant differences in gmax among cultivars.

|

|
For several cultivars, the estimated asymptotic gmax value from the sigmoidal fit was greater than the highest actual measured mean gmax value (e.g. Syrah and Tinta Madeira,) or there were few data points near the estimated gmax (e.g. Tinta Amarela) (Fig. 3). One consequence was that the derived Ψl95 values fell outside of the measured values for Syrah and Tinta Madeira, or that the estimated gmax value was a poor representation of a cultivar’s actual gmax. Since subsequent analyses depended upon correct determination of gmax, the estimated gmax values were regressed on the highest mean measured gmax (taken at the highest Ψl). There was a statistically significant relationship between estimated and measured gmax (P < 0.05), though there were some notable outliers (e.g. Syrah and Durif) that weakened the relationship (R2 = 0.31). Nevertheless, the estimated slope of the relationship was not significantly different than unity (slope ± 95% confidence interval = 0.599 ± 0.493). Thus, it was assumed that the estimated gmax value from the sigmoidal fit was representative of actual gmax for all subsequent analyses.
The CV was similarly low (6%) across cultivars for estimates of Ψl50, which ranged 0.32 MPa from –1.27 (Touriga Nacional) to –1.59 MPa (Montepulciano) with no differences among cultivars. In contrast, the unitless scalar parameter b – the transition width of the function related to the slope – varied broadly among cultivars (CV = 29%). Accordingly, there was broad variation in estimates of gsensitivity (CV = 32%) among cultivars, with values ranging 3-fold from 0.428 (Petit Verdot) to 1.283 mol m–2 s–1 MPa–1 (Tannat). Further, gsensitivity was 0.562 and 0.525 mol m–2 s–1 MPa–1 for Grenache and Syrah respectively (Table 3).
When comparing modelled Ψl and gs values among cultivars at each extracted vine water status value (i.e. Ψl95, Ψl50 and Ψl25), the across-cultivar CV for modelled Ψl decreased with vine water status (i.e. Ψl95 > Ψl50 > Ψl25), whereas across-cultivar CV for modelled gs remained constant at each value (i.e. Ψl95 = Ψl50 = Ψl25) (Table 3). This was not surprising given that the model was fit to minimise variation in gs. The across-cultivar mean modelled gs and Ψl at high water status (i.e. Ψl95) was more than 0.4 mol m–2 s–1 and –0.99 MPa, respectively, whereas the across-cultivar mean modelled gs and Ψl at low water status (i.e. Ψl25) was near 0.1 mol m–2 s–1 and –1.68 MPa. Furthermore, the cultivars with high gs at high vine water status (Ψl95) tended to have similarly high gs at low vine water status. For example, Syrah had the highest gs among all cultivars at Ψl95 and at Ψl25, whereas Grenache had the lowest gs at both Ψl95and Ψl25 (Table 3). However, their gsensitivity values were similar (as stated above).
There was a significant negative relationship between gsensitivity and Ψl95 across all cultivars (Fig. 4). As the water status threshold for the onset of stomatal closure (Ψl95) decreased, sensitivity to further closure increased across the entire range of Ψl95 values. Moreover, all 17 cultivars were broadly distributed along the curve and did not cluster together. Notably, Grenache and Syrah were distributed similarly on the vertical axis (gsensitivity) and were separated only on the horizontal axis (Ψl95).

|
Relationship of cultivar mean Ψl and Ψl25
Overall cultivar mean Ψl (averaged across treatments and years) was positively related to Ψl25 both pre- and post-veraison (Fig. 5). Petit Verdot was removed from the analysis as an outlier due to its large absolute residual value in both phenological periods. There were no significant differences in response between phenological periods (slopes not significantly different), except that cultivar mean Ψl values were significantly lower post-veraison by 0.21 MPa (likely due to the lack of a fully irrigated post-veraison treatment). Nevertheless, in both phenological periods a higher Ψl25 resulted in a higher cultivar mean Ψl with nearly a 1 : 1 ratio based on the slopes of the linear regression lines. The linear regression model was highly significant, and Ψl25 explained 83% of the variation in cultivar mean Ψl when the model included both phenological periods.

|
Discussion
Since the paper published by Schultz (2003) suggesting that V. vinifera cultivars may differ in their stomatal behaviour in response to water deficits, there has been great interest among researchers in unravelling the causes and underlying mechanisms of the an/isohydric phenomenon across cultivars (Soar et al. 2006b; Collins et al. 2010; Gerzon et al. 2015; Hochberg et al. 2015; Coupel-Ledru et al. 2017; Charrier et al. 2018). However, the obvious experimental difficulty in conducting large, field-scale experiments across many cultivars has limited previous investigations to only a few cultivars at a time or to the comparison of various cultivars grown in different vineyards or locations. As such, much work has been done on potted vines whose responses may not be representative of field performance – particularly in terms of plant water relations (Passioura 2006) – and on cultivars selected based on a priori classification as either iso- or anisohydric from few data – notably Grenache and Syrah. Several reviews have been published on the topics of irrigation management and/or water relations of grapevine that have attempted to synthesise the existing literature on across-cultivar stomatal behaviour (Cifre et al. 2005; Chaves et al. 2010; Lovisolo et al. 2010; Costa et al. 2012; Domec and Johnson 2012). Unfortunately, differences in plant material, environmental conditions, and experimental methodology and techniques have limited the ability of past reviews to characterise various cultivars’ responses and have ultimately resulted in inconsistent classification. In an attempt to resolve these problems, Lavoie-Lamoureux et al. (2017) conducted a meta-analysis of 40 publications across 28 V. vinifera cultivars and hypothesised that rather than separating into two distinct groups, cultivars’ stomatal behaviours were distributed along a continuum. A similar result was found by Klein (2014) in a meta-analysis across 70 plant species.
This study explored genetic variation in vine water status and stomatal behaviour across 17 V. vinifera cultivars (clone wood and source known) grafted onto the same rootstock (1103P), grown in a replicated field trial in a high VPD area (San Joaquin Valley of California), and subjected to three season-long deficit irrigation treatments over the course of multiple seasons. Therefore, soil type, weather conditions and cultural practices were the same for all cultivars. Midday Ψl was measured by the same individual on each date in 2014 and 2015 using the technique whereby leaf blades were covered with a plastic bag just before leaf excision therefore minimising potential technique (Williams and Araujo 2002; Williams 2017) and operator errors (Goldhamer and Fereres 2001). Unfortunately, this was not one of the factors considered in the meta-analysis by Lavoie-Lamoureux et al. (2017) and even now the exact method used or a reference provided to describe how Ψl was measured is not given (Charrier et al. 2018). The timing interval between the two measurements only had to be within two hours in the meta-analysis of Lavoie-Lamoureux et al. (2017), and it is unknown to what degree this would affect their results. Stomatal conductance was measured with the same LI-COR porometer in each year on the same leaf used for the measurement of Ψl just before its excision. Thus, the design and techniques used in this study should have maximised the effect of cultivar on the parameters in question. The hypothesis that cultivars do not separate into clear iso- or anisohydric groups, but rather are distributed along a continuum was tested by (1) establishing a range of plant water deficits using variable amounts and timings of applied water, (2) measuring ΨPD and Ψl the first year and (3) measuring gs and Ψl on the same leaf samples the third and fourth years, and then comparing response curves among cultivars.
Isohydric, near-isohydric, anisohydric or isohydrodynamic?
In general, an isohydric plant will maintain its water potential value similar between water stressed and well-watered cohorts (Tardieu and Simonneau 1998). The results from the first year of the study clearly demonstrate that none of the cultivars in this study responded to soil water deficits in an isohydric manner but were more anisohydric-like, in agreement with that of (Soar et al. 2006b). Midday Ψl of vines in the three irrigation treatments differed significantly from one another – midday Ψl of well-watered vines across cultivars were always much greater than the other irrigation treatments and as the water in the soil profile was depleted, midday Ψl decreased for all cultivars including that of Grenache. None of the cultivars examined in this study maintained a minimal midday Ψl plateau (> –1.5 MPa) as reported previously for near-isohydric grape cultivars (Schultz 2003; Prieto et al. 2010). In addition, the slope of the relationship between midday Ψl and ΨPD reported here did not differ among cultivars, similar to that observed by Charrier et al. (2018) for Grenache and Syrah. Lastly, the cultivars did not appear to respond to soil water deficits in an isohydrodynamic manner (Franks et al. 2007; Shellie and Bowen 2014).
Exploring the curve: cultivar differences in the relation of gs to Ψl
A sigmoidal gs relation to Ψl was present in all 17 cultivars and in a composite analysis. A similar result was found by Charrier et al. (2018). Linear relationships between gs and Ψl (and Ψstem) have been commonly reported in grape (Williams and Araujo 2002; Shackel 2007; Williams 2012; Williams et al. 2012). However, nonlinearity may have been overlooked in those studies. The reported ranges of Ψl or Ψstem were narrower (~0.5 to 1.0 MPa) than that reported herein (1.4 MPa), thus may have only represented the linear portion of the entire response curve. Furthermore, there are few data in the aforementioned studies at low vine water status (Ψl or Ψstem < –1.5 MPa), and the reported data at high vine water status (Ψl or Ψstem > –1.0 MPa) show increasing vertical scatter. Indeed, when a broader range of vine water status was evaluated, non-linear relationships between gs and Ψl (and Ψstem) have been shown for V. vinifera cv. Sauvignon blanc (Naor et al. 1994, 1997), and between gs and ΨPD across five cultivars (Prieto et al. 2010). Recently, sigmoidal functions were also well fit to data relating grapevine leaf petiole percent loss of conductivity to xylem tension in potted grapevine (cv. Merlot) (Hochberg et al. 2017a). Finally, sigmoidal relations of gs to Ψl has been reported for woody and non-woody crops alike, including: Populus spp. (Silim et al. 2009), Olea europea (Ennajeh et al. 2008), Oryza sativa (Dingkuhn et al. 1989), Sorghum bicolor (Henzell et al. 1976); and across a broad range of tree species (Klein 2014). Nevertheless, in this study the nature of the sigmoidal relationship between gs and Ψl depended greatly upon cultivar.
No cultivar differences at high water status
Under well-watered conditions, there were no cultivar differences in gmax. While both high gmax (Henzell et al. 1976) and low gmax (Sinclair et al. 2005) have been proposed means to improve productivity under dry conditions, there was surprisingly little variation in gmax under well-watered conditions among cultivars over two years. Measured and estimated gmax values in this study (~0.4 to 0.6 mol m–2 s–1 across cultivars) were similar to other reported values for well-watered grapevines (Williams and Baeza 2007; Bota et al. 2016), and there were no significant differences among cultivars. Interestingly, measured gmax values among cultivars were more similar than were estimated gmax values, further supporting the point that cultivars behave similarly under well-watered conditions. Thus, no prospects were revealed for improved efficiency at either high water status (Price et al. 2002) or at low water status (Silim et al. 2009).
The lone gmax outlier – Syrah, who’s estimated gmax was more than 0.1 mol m–2 s–1 greater than measured gmax – was likely due a lack of data density in the well-watered range (> –1.0 MPa) that ultimately resulted in a poor estimate of the asymptote by the sigmoidal fit. This was also true for Tinta Madeira, though the difference between measured and estimated gmax was smaller. The question remains whether or not higher gs values would be measured for those cultivars – and a more accurate estimate for gmax would be obtained – at higher Ψl values. Although it is not biologically impossible – Soar et al. (2009) reported gs values for Shiraz (syn = Syrah) near 0.8 mol m–2 s–1 – it is unlikely under field conditions, primarily because those values were obtained through artificial heating and under high relative humidity. Also, significantly higher mid-season Ψl values (> –0.7 MPa) would also be difficult to reach under field conditions (Williams and Baeza 2007).
Generally speaking, the role of gmax in terms of drought tolerance has been evaluated almost exclusively in the context of crop species that are grown for agronomic purposes (yield/biomass production) – e.g. Populus spp., O. sativa, and S. bicolor. Considering that grapevine water use is highly correlated with gs (Williams et al. 2012) and canopy percent shaded area/canopy coverage (Williams and Ayars 2005), similar gmax across cultivars would suggest that canopy percent shaded area alone – ultimately determined by relative cultivar vigour and canopy training/trellising system – determines total consumptive water use in grapevine. However, understanding the importance of high or low gmax in wine grape production is further complicated by the common horticultural purposes (fruit quality) in which water deficits are often beneficial (Roby et al. 2004).
Cultivar differences become apparent at moderate water status
As water deficits increased, differences among cultivars in the onset of stomatal closure (Ψl95) became apparent, ranging from Syrah at –0.76 MPa to Tannat at –1.26 MPa. There are few reported data regarding the Ψl at the onset of closure for grapevines, primarily due to the predominant practice of fitting linear curves through the data (as discussed above), or simply not conducting a regression analysis. Williams and Baeza (2007) suggest a Ψl value of –1.2 MPa as signifying when grapevines are no longer responsive to changes in VPD, but rather to changes in soil moisture. In their recent meta-analysis of the literature, Lavoie-Lamoureux et al. (2017) suggest using –1.214 MPa as the water status level at which to compare cultivars. It is likely that this value – Ψl = –1.2 MPa – varies by cultivar, and indeed falls in the lower end of the across-cultivar Ψl95 range obtained herein. The data of Hochberg et al. (2017b) show that Ψl95 for Chardonnay was closer to Ψl ≈ –0.8 MPa, and Hochberg et al. (2017a) show a 12% loss of conductivity (i.e. Ψl88) in Merlot occurring at Ψstem ≈ –0.66 to –0.91 MPa – which would correspond to a Ψl ≈ –0.97 to –1.20 MPa (Williams and Araujo 2002). Finally, a similar range in the onset of stomatal closure has been reported among Poplar clones (Silim et al. 2009) and across 70 other tree species (Klein 2014). Therefore, the cultivar-specific water potential values for the onset of stomatal closure in this study are within the expected range for V. vinifera, and moreover seem to be highly conserved across other woody perennial plant species as well.
Like the onset of stomatal closure, gsensitivity was cultivar-specific, revealing a 3-fold difference in slopes (Fig. 4). Prevailing definitions of iso- and anisohydry state that isohydric plants maintain Ψl almost constant as soil dries due to more sensitive stomata, whereas anisohydric plants maintain gs and allow Ψl to decline with soil drying (Tardieu and Simonneau 1998; Jones 2014). Using this definition, cultivars in this study with high gsensitivity (e.g. Tannat, Durif and Montepulciano) could be categorised as isohydric/near-isohydric, whereas those with low gsensitivity (e.g. Petit Verdot, Tinta Madeira, Freisa) could be categorised as anisohydric/near-anisohydric.
In the present study, Grenache (often considered the archetypal isohydric cultivar) and Syrah (often considered the archetypal anisohydric cultivar) had almost the same gsensitivity (maximum slope of the linear part of the curves), and did not segregate based on this criterion. However, simple observation of the sigmoidal functions fit to Grenache and Syrah clearly shows that they behave differently at moderate water status – Grenache is more sigmoidal with a clear asymptotic levelling off of gs at high Ψl values, whereas Syrah is less sigmoidal without a clear asymptotic levelling off of gs at high Ψl. If gsensitivity were calculated at moderate water status values (e.g. Ψl95 > Ψl > Ψl50) instead of in the linear portion (e.g. at Ψl50), it is possible that Grenache and Syrah would segregate clearly based on this criterion. Simply looking at this constrained dataset supports some previous classification of these two cultivars. Yet the fact that their behaviour – and ultimate classification – depends on how the stomatal sensitivity parameter is defined calls into question its usefulness for classification.
In their comparison of five cultivars, Prieto et al. (2010) reported that a stomatal sensitivity factor, k – calculated using the Ball, Woodrow, and Berry model – significantly differed between Grenache and Syrah, but their level of significance for this parameter was α = 0.10. Furthermore, they show regressions of k to predawn water potential for Syrah and Ekigaina (as examples of extreme cases), but do not show the comparison data for Grenache. Recent work has also shown that there are no statistically significant differences between Grenache and Syrah with respect to sensitivity of transpiration rate to declining soil water availability under both field and greenhouse conditions (Charrier et al. 2018). Therefore, other factors affecting stomatal behaviour such as leaf hydraulics or environment may be more important to consider for cultivar classification going forward.
It is notable that Grenache and Syrah – the two cultivars first compared by Schultz (2003) – differed in this study only in their Ψl95 value, but had almost the same gsensitivity (Fig. 4). Although widely cited as the foundational evidence for the an/isohydric phenomenon within V. vinifera cultivars, it is seldom discussed that Schultz (2003) actually used Grenache and Syrah scions grafted onto different rootstocks, 140 Ruggeri and 110 Richter respectively (Schultz 1996). Vitis spp. rootstocks differ in their effects on vine water relations and gas exchange (Padgett-Johnson et al. 2003), and have also been shown to significantly affect scion gas exchange (Soar et al. 2006a) as well as scion growth and productivity (Williams 2010). Lavoie-Lamoureux et al. (2017) reported that rootstock generally accounts for 19% of the variability in scion gs response to water deficits. When grafted onto the same rootstock, similar rates of stomatal closure between Grenache and Syrah in response to Ψstem (ranging from –0.5 to –1.5 MPa) have been reported in a recent three-year field study (Bota et al. 2016). It did appear that Grenache and Syrah behaved differently at moderate water status in this study, in contrast to Schultz (2003) who showed a greater divergence between the two cultivars at lower water status. Well-watered conditions have been shown to increase variation in stomatal response across Grenache-Syrah progeny (Coupel-Ledru et al. 2017). Indeed, it should be noted that Syrah maintained a higher gs compared with Grenache at all water status levels in this study. Nevertheless, whether considering Grenache and Syrah separately, or all the cultivars together in this study, there were more differences among them in stomatal behaviour at moderate water status relative to low water status.
Few cultivar differences at low water status
Analysis of gs at Ψl25 and the similarity of Ψl among cultivars at Ψl25 suggests a common water status limit to low gs among grapevine cultivars. At Ψl25, gs was already down to 0.08 to 0.1 mol m–2 s–1 and declined gradually thereafter. Grapevines have been previously classified as severely stressed when gs <0.05 to 0.1 mol m–2 s–1 (Cifre et al. 2005; Lovisolo et al. 2010). The lack of apparent cultivar differences at low water status may have been due to the relatively poor fit of the sigmoidal functions at these levels (shown by increasing homoscedasticity in residuals in Fig. 2b). However, all extracted Ψl25 values were above these poorly fit regions of the dataset. Brodribb and Holbrook (2003) regarded Ψl20 as the point of stomatal closure in their analysis of stomatal behaviour across four tropical tree species, and Klein (2014) used Ψl25 as a functional parameter by which to compare stomatal behaviour of 70 woody perennial plant species. Hence, it is reasonable to regard the value of Ψl25 as the functional lower limit of stomatal contribution to leaf gas exchange and overall vine performance.
Stomata function in part to protect the hydraulic integrity of the xylem (Tyree and Sperry 1988; Brodribb and Holbrook 2003), but also close in response to reduction in leaf turgor (Shackel et al. 1987). Thus, a reduction in gs is both an impact of and a protective response to declining plant water status. This inherently complicates behavioural interpretation. Choat et al. (2010) showed that xylem cavitation commences at much lower water status than previously accepted, and Hochberg et al. (2017b) recently showed that stomata close well in advance of xylem cavitation in grapevine. In the latter work, gs was less than 10% of gmax before the first embolisms were detected in the xylem (–1.46 > Ψl > –1.75 MPa). Those Ψl values at the appearance of the first embolisms closely correspond to the Ψl25 values found among cultivars in this study (-1.5 > Ψl > –1.8 MPa; Fig. 5). Therefore, our data indicate most stomata were closed for all cultivars at or below their Ψl25 value, and that the cultivar-specific Ψl25 value may be related to the appearance of first embolisms in the xylem. Thus, each cultivar-specific Ψl25 value may indicate a hydraulically determined water status limit, which may be governed by cultivar differences in vascular anatomy (Gerzon et al. 2015; Hochberg et al. 2015).
Potential physiological mechanisms to explain cultivar differences
It is likely that the different stomatal behaviours observed among cultivars were mediated by hydraulic or turgor-dependent signals, which are closely associated with incipient stomatal closure (Brodribb and Holbrook 2003). In their modelling work on partitioning stomatal responses to drought, Rodriguez-Dominguez et al. (2016) found that leaf turgor-dependent mechanisms accounted for a large majority of the decline in gs to soil drought across three woody perennial crop species, including grapevine. Reduced hydraulic conductivity and increased osmotic adjustment made up a large proportion of the overall stomatal response in that study.
In general, differences in stomatal closure among plant species may arise in part from differences in osmotic adjustment (Hsiao 1973), leading to acclimation of stomatal response and leaf hydraulic conductivity to low water status (Matthews and Boyer 1984; Martorell et al. 2015). Differences in osmotic adjustment have also been closely associated with differences in stomatal behaviours between tree species (Nolan et al. 2017). Grape leaves also go through significant osmotic adjustment (Schultz and Matthews 1993), and there is recent evidence for differences in osmotic adjustment among V. vinifera L. cultivars (Martorell et al. 2015). Differences in osmotic adjustment have also been previously reported among cultivars in wheat, sorghum, and cotton (Morgan 1984), and in O. europea (Ennajeh et al. 2008). Thus, genetic variation in the osmotic adjustment response may play an important role in the wide range of cultivar responses found in this study.
Conclusions
The results of this study provide clear evidence in support of an across-cultivar continuum of stomatal behaviour, and call into question previous classifications of V. vinifera cultivars as having either exclusively iso- or anisohydric stomatal responses. First, cultivars did not separate into distinct iso- or anisohydric groups based on their Ψl v. ΨPD relationships. Moreover, a deeper exploration into genetic variation among cultivars revealed that instead of separating into distinct groups, cultivars were distributed along a continuum based upon the negative relationship between the sensitivity (gsensitivity) and the onset (Ψl95) of stomatal closure in the face of declining vine water status. Notably, this continuum was well defined at moderate status values, but cultivars behaved similarly at high and low water status.
At high vine water status, cultivars had similar gmax values, suggesting that overall vine water use would simply be a function of canopy size under well watered and saturating light conditions. As Ψl decreased from well-watered conditions, all cultivars responded similarly to one another by maintaining a high gs – an anisohydric characteristic. This was despite several of the cultivars having been assigned to the isohydric/near-isohydric category in other studies. However, closer examination of the gs vs Ψl relationship showed that as vine water status declined from well-watered conditions, stomata of various cultivars responded differently to minimise water loss – an isohydric characteristic. Finally, at low water status, gs was the similar for all cultivars (<0.1 mol m–2 s–1), but the water status value at the end of stomatal closure (Ψl25) differed among them, indicating that there is a cultivar-dependent water status limit to stomatal functioning.
The large and significant differences in midday Ψl values measured as a function of irrigation treatment across all cultivars in this study would indicate that midday Ψl provides an accurate assessment of vine water status, and would therefore be useful to validate other measures of monitoring vine water status, or as a tool in an irrigation management program. In terms of cultivar-specific stomatal behaviour, all cultivars examined herein responded in an anisohydric manner at high water status by keeping stomata open as Ψl decreased. This was despite several of the cultivars having been assigned to the isohydric/near isohydric category in other studies. However, closer examination of the gs vs Ψl relationship indicates there may be times – at moderate water deficits – in which stomata of various cultivars may respond differently to minimise water loss, an isohydric response. This would indicate that V. vinifera cultivars have both iso- and anisohydric stomatal responses to water deficits, and that any future classification should incorporate a holistic evaluation of the entire response curve. Finally, differences in osmotic adjustment among cultivars may play the primary role in determining the observed differences in stomatal behaviour.
Conflicts of interest
Larry E Williams, Mark A Matthews and Alexander D Levin conceived of and designed the research. ADL conducted experiments, analysed data, and wrote the manuscript. All authors read and approved the manuscript. The authors declare no conflicts of interest.
Acknowledgements
The authors would like to thank GA Gambetta, KA Shackel and AJ McElrone for helpful discussions related to earlier drafts of this manuscript; and Judith Chiginsky and Laura Fontaine for their technical assistance in data collection. The authors would also like to thank the staff at the KARE Center for vineyard management services. This work was supported in part by the American Vineyard Foundation (award number 2014–1533) and the California Institute of Water Resources Joseph G Prosser Trust Grant (award number 2015CA345B). This work also served as partial satisfaction of the requirements for the completion of A Levin’s doctoral thesis, which was generously supported by the Henry A Jastro Research Award, the Wine Spectator Scholarship, the Horace O Lanza Scholarship, the Adolf C and Richie C Heck Research Fellowship, the Louis R Gomberg Scholarship, CO Foerster Jr Scholarship, the Nathan Fay Scholarship, the Pearl and Albert J Winkler Scholarship in Viticulture, the Harold P Olmo Scholarship, the Richard and Saralee Kunde Scholarship, the David E Gallo Award, the John Ferrington Award, the Robert Lawrence Balzer Scholarship, the Curtis J Alley Memorial Research Scholarship, the Leon D Adams Research Scholarship, and the Andre Tchelistcheff and Dr Richard Peterson Scholarship.
References
Bates LM, Hall AE (1981) Stomatal closure with soil water depletion not associated with changes in bulk leaf water status. Oecologia 50, 62–65.| Stomatal closure with soil water depletion not associated with changes in bulk leaf water status.Crossref | GoogleScholarGoogle Scholar | 28310062PubMed |
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67, 1–48.
| Fitting linear mixed-effects models using lme4.Crossref | GoogleScholarGoogle Scholar |
Bota J, Tomas M, Flexas J, Medrano H, Escalona JM (2016) Differences among grapevine cultivars in their stomatal behavior and water use efficiency under progressive water stress. Agricultural Water Management 164, 91–99.
| Differences among grapevine cultivars in their stomatal behavior and water use efficiency under progressive water stress.Crossref | GoogleScholarGoogle Scholar |
Brodribb TJ, Holbrook NM (2003) Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiology 132, 2166–2173.
| Stomatal closure during leaf dehydration, correlation with other leaf physiological traits.Crossref | GoogleScholarGoogle Scholar | 12913171PubMed |
Charrier G, Delzon S, Domec JC, Zhang L, Delmas CEL, Merlin I, Corso D, King A, Ojeda H, Ollat N, Prieto JA, Scholach T, Skinner P, van Leeuwen C, Gambetta G (2018) Drought will not leave your glass empty: low risk of hydraulic failure by long-term drought observations in world’s top wine regions. Science Advances 4, eaao6969
| Drought will not leave your glass empty: low risk of hydraulic failure by long-term drought observations in world’s top wine regions.Crossref | GoogleScholarGoogle Scholar | 29404405PubMed |
Chaves MM, Zarrouk O, Francisco R, Costa JM, Santos T, Regalado AP, Rodrigues ML, Lopes CM (2010) Grapevine under deficit irrigation: hints from physiological and molecular data. Annals of Botany 105, 661–676.
| Grapevine under deficit irrigation: hints from physiological and molecular data.Crossref | GoogleScholarGoogle Scholar | 20299345PubMed |
Choat B, Drayton WM, Brodersen C, Matthews MA, Shackel KA, Wada H, McElrone AJ (2010) Measurement of vulnerability to water stress-induced cavitation in grapevine: a comparison of four techniques applied to a long-vesseled species. Plant, Cell & Environment 33, 1502–1512.
Chouzouri A, Schultz HR (2005) Hydraulic anatomy, cavitiation susceptibility and gas-exchange of several grapevine cultivars of different geographic origin. Acta Horticulturae 689, 325–332.
| Hydraulic anatomy, cavitiation susceptibility and gas-exchange of several grapevine cultivars of different geographic origin.Crossref | GoogleScholarGoogle Scholar |
Cifre J, Bota J, Escalona JM, Medrano H, Flexas J (2005) Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.). Agriculture, Ecosystems & Environment 106, 159–170.
| Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.).Crossref | GoogleScholarGoogle Scholar |
Collins MJ, Fuentes S, Barlow EWR (2010) Partial rootzone drying and deficit irrigation increase stomatal sensitivity to vapour pressure deficit in anisohydric grapevines. Functional Plant Biology 37, 128
| Partial rootzone drying and deficit irrigation increase stomatal sensitivity to vapour pressure deficit in anisohydric grapevines.Crossref | GoogleScholarGoogle Scholar |
Costa JM, Ortuño MF, Lopes CM, Chaves MM (2012) Grapevine varieties exhibiting differences in stomatal response to water deficit. Functional Plant Biology 39, 179–189.
| Grapevine varieties exhibiting differences in stomatal response to water deficit.Crossref | GoogleScholarGoogle Scholar |
Coupel-Ledru A, Tyerman SD, Masclef D, Lebon E, Christophe A, Edwards EJ, Simonneau T (2017) Abscisic acid down-regulates hydraulic conductance of grapevine leaves in isohydric genotypes only. Plant Physiology 175, 1121–1134.
| Abscisic acid down-regulates hydraulic conductance of grapevine leaves in isohydric genotypes only.Crossref | GoogleScholarGoogle Scholar | 28899961PubMed |
Dingkuhn M, Cruz RT, O’Toole JC, Dorffling K (1989) Net photosynthesis, water use efficiency, leaf water potential and leaf rolling as affected by water deficit in tropical upland rice. Australian Journal of Agricultural Research 40, 1171–1181.
| Net photosynthesis, water use efficiency, leaf water potential and leaf rolling as affected by water deficit in tropical upland rice.Crossref | GoogleScholarGoogle Scholar |
Domec JC, Johnson DM (2012) Does homeostasis or disturbance of homeostasis in minimum leaf water potential explain the isohydric versus anisohydric behavior of Vitis vinifera L. cultivars? Tree Physiology 32, 245–248.
| Does homeostasis or disturbance of homeostasis in minimum leaf water potential explain the isohydric versus anisohydric behavior of Vitis vinifera L. cultivars?Crossref | GoogleScholarGoogle Scholar | 22427373PubMed |
Ennajeh M, Tounekti T, Vadel AM, Khemira H, Cochard H (2008) Water relations and drought-induced embolism in olive (Olea europaea) varieties ‘Meski’ and ‘Chemlali’ during severe drought. Tree Physiology 28, 971–976.
| Water relations and drought-induced embolism in olive (Olea europaea) varieties ‘Meski’ and ‘Chemlali’ during severe drought.Crossref | GoogleScholarGoogle Scholar | 18381277PubMed |
Franks PJ, Drake PL, Froend RH (2007) Anisohydric but isohydrodynamic: seasonally constant plant water potential gradient explained by a stomatal control mechanism incorporating variable plant hydraulic conductance. Plant, Cell & Environment 30, 19–30.
| Anisohydric but isohydrodynamic: seasonally constant plant water potential gradient explained by a stomatal control mechanism incorporating variable plant hydraulic conductance.Crossref | GoogleScholarGoogle Scholar |
Gerzon E, Biton I, Yaniv Y, Zemach H, Netzer Y, Schwartz A, Fait A, Ben-Ari G (2015) Grapevine anatomy as a possible determinant of isohydric or anisohydric behavior. American Journal of Enology and Viticulture 66, 340–347.
| Grapevine anatomy as a possible determinant of isohydric or anisohydric behavior.Crossref | GoogleScholarGoogle Scholar |
Goldhamer DA, Fereres E (2001) Simplified tree water status measurements can aid almond irrigation. California Agriculture 55, 32–37.
| Simplified tree water status measurements can aid almond irrigation.Crossref | GoogleScholarGoogle Scholar |
Henzell RG, McCree KJ, Van Bavel CHM, Schertz KF (1976) Sorghum genotype variation in stomatal sensitivity to leaf water deficit. Crop Science 16, 660–662.
| Sorghum genotype variation in stomatal sensitivity to leaf water deficit.Crossref | GoogleScholarGoogle Scholar |
Hochberg U, Degu A, Gendler T, Fait A, Rachmilevitch S (2015) The variability in the xylem architecture of grapevine petiole and its contribution to hydraulic differences. Functional Plant Biology 42, 357
| The variability in the xylem architecture of grapevine petiole and its contribution to hydraulic differences.Crossref | GoogleScholarGoogle Scholar |
Hochberg U, Bonel AG, David-Schwartz R, Degu A, Fait A, Cochard H, Peterlunger E, Herrera JC (2017a) Grapevine acclimation to water deficit: the adjustment of stomatal and hydraulic conductance differs from petiole embolism vulnerability. Planta 245, 1091–1104.
| Grapevine acclimation to water deficit: the adjustment of stomatal and hydraulic conductance differs from petiole embolism vulnerability.Crossref | GoogleScholarGoogle Scholar | 28214919PubMed |
Hochberg U, Windt CW, Ponomarenko A, Zhang YJ, Gersony J, Rockwell FE, Holbrook NM (2017b) Stomatal closure, basal leaf embolism, and shedding protect the hydraulic integrity of grape stems. Plant Physiology 174, 764–775.
| Stomatal closure, basal leaf embolism, and shedding protect the hydraulic integrity of grape stems.Crossref | GoogleScholarGoogle Scholar | 28351909PubMed |
Hochberg U, Rockwell FE, Holbrook NM, Cochard H (2018) Iso/anisohydry: a plant–environment interaction rather than a simple hydraulic trait. Trends in Plant Science 23, 112–120.
| Iso/anisohydry: a plant–environment interaction rather than a simple hydraulic trait.Crossref | GoogleScholarGoogle Scholar | 29223922PubMed |
Hsiao TC (1973) Plant responses to water stress. Annual Review of Plant Physiology 24, 519–570.
| Plant responses to water stress.Crossref | GoogleScholarGoogle Scholar |
Jones HG (1974) Assessment of stomatal control of plant water status. New Phytologist 73, 851–859.
| Assessment of stomatal control of plant water status.Crossref | GoogleScholarGoogle Scholar |
Jones HG (2014) ‘Plants and microclimate: a quantitative approach to environmental plant physiology.’ (Cambridge University Press: Cambridge, UK)
Klein T (2014) The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours. Functional Ecology 28, 1313–1320.
| The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours.Crossref | GoogleScholarGoogle Scholar |
Kuznetsova A, Brockoff PB, Rune HB (2017) lmerTest Package: tests in linear mixed effects models. Journal of Statistical Software 82, 1–26.
| lmerTest Package: tests in linear mixed effects models.Crossref | GoogleScholarGoogle Scholar |
Lavoie-Lamoureux A, Sacco D, Risse PA, Lovisolo C (2017) Factors influencing stomatal conductance in response to water availability in grapevine: a meta-analysis. Physiologia Plantarum 159, 468–482.
| Factors influencing stomatal conductance in response to water availability in grapevine: a meta-analysis.Crossref | GoogleScholarGoogle Scholar | 27859326PubMed |
Lenth R (2016) Least-squares means: the R package lsmeans. Journal of Statistical Software 69, 1–33.
| Least-squares means: the R package lsmeans.Crossref | GoogleScholarGoogle Scholar |
Lovisolo C, Perrone I, Carra A, Ferrandino A, Flexas J, Medrano H, Schubert A (2010) Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non-hydraulic interactions at the whole-plant level: a physiological and molecular update. Functional Plant Biology 37, 98–116.
| Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non-hydraulic interactions at the whole-plant level: a physiological and molecular update.Crossref | GoogleScholarGoogle Scholar |
Martin-St. Paul N, Delzon S, Cochard H (2017) Plant resistance to drought relies on early stomatal closure. Available at bioRxiv.org [Verified 12 September 2019].
Martinez-Vilalta J, Garcia-Forner N (2017) Water potential regulation, stomatal behaviour and hydraulic transport under drought: deconstructing the iso/anisohydric concept. Plant, Cell & Environment 40, 962–976.
| Water potential regulation, stomatal behaviour and hydraulic transport under drought: deconstructing the iso/anisohydric concept.Crossref | GoogleScholarGoogle Scholar |
Martorell S, Medrano H, Tomas M, Escalona JM, Flexas J, Diaz-Espejo A (2015) Plasticity of vulnerability to leaf hydraulic dysfunction during acclimation to drought in grapevines: an osmotic-mediated process. Physiologia Plantarum 153, 381–391.
| Plasticity of vulnerability to leaf hydraulic dysfunction during acclimation to drought in grapevines: an osmotic-mediated process.Crossref | GoogleScholarGoogle Scholar | 25132228PubMed |
Matthews MA, Boyer JS (1984) Acclimation of photosynthesis to low leaf water potentials. Plant Physiology 74, 161–166.
| Acclimation of photosynthesis to low leaf water potentials.Crossref | GoogleScholarGoogle Scholar | 16663372PubMed |
Morgan JM (1984) Osmoregulation and water stress in higher plants. Annual Review of Plant Physiology 35, 299–319.
| Osmoregulation and water stress in higher plants.Crossref | GoogleScholarGoogle Scholar |
Naor A, Bravdo B, Gelobter J (1994) Gas exchange and water relations in field-grown Sauvignon blanc grapevines. American Journal of Enology and Viticulture 45, 423–428.
Naor A, Gal Y, Bravdo B (1997) Crop lad affects assimilation rate, stomatal conductance, stem water potential and water relations of field-grown Sauvignon blanc grapevines. Journal of Experimental Botany 48, 1675–1680.
| Crop lad affects assimilation rate, stomatal conductance, stem water potential and water relations of field-grown Sauvignon blanc grapevines.Crossref | GoogleScholarGoogle Scholar |
Nolan RH, Tarin T, Santini NS, McAdam SAM, Ruman R, Eamus D (2017) Differences in osmotic adjustment, foliar abscisic acid dynamics, and stomatal regulation between an isohydric and anisohydric woody angiosperm during drought. Plant, Cell & Environment 40, 3122–3134.
| Differences in osmotic adjustment, foliar abscisic acid dynamics, and stomatal regulation between an isohydric and anisohydric woody angiosperm during drought.Crossref | GoogleScholarGoogle Scholar |
Padgett-Johnson M, Williams LE, Walker MA (2003) Vine water relations, gas exchange, and vegetative growth of seventeen Vitis species grown under irrigated and nonirrigated conditions in California. Journal of the American Society for Horticultural Science 128, 269–276.
| Vine water relations, gas exchange, and vegetative growth of seventeen Vitis species grown under irrigated and nonirrigated conditions in California.Crossref | GoogleScholarGoogle Scholar |
Passioura JB (2006) Viewpoint: the perils of pot experiments. Functional Plant Biology 33, 1075
| Viewpoint: the perils of pot experiments.Crossref | GoogleScholarGoogle Scholar |
Price AH, Cairns JE, Horton P, Jones HG, Griffiths H (2002) Linking drought-resistance mechanisms to drought avoidance in upland rice using a QTL approach: progress and new opportunities to integrate stomatal and mesophyll responses. Journal of Experimental Botany 53, 989–1004.
| Linking drought-resistance mechanisms to drought avoidance in upland rice using a QTL approach: progress and new opportunities to integrate stomatal and mesophyll responses.Crossref | GoogleScholarGoogle Scholar | 11971911PubMed |
Prieto JA, Lebon E, Ojeda H (2010) Stomatal behavior of different grapevine cultivars in responses to soil water status and air water vapor pressure deficit. Journal International des Sciences de la Vigne et du Vin 44, 9–20.
Roby G, Harbertson JF, Adams DO, Matthews MA (2004) Berry size and vine water deficits as factors in winegrape composition: anthocyanins and tannins. Australian Journal of Grape and Wine Research 10, 100–107.
| Berry size and vine water deficits as factors in winegrape composition: anthocyanins and tannins.Crossref | GoogleScholarGoogle Scholar |
Rodrigues P, Pedroso V, Gouveia JP, Martins S, Lopes C, Alves I (2012) Influence of soil water content and atmospheric conditions on leaf water potential in cv. ‘Touriga Nacional’ deep-rooted vineyards. Irrigation Science 30, 407–417.
| Influence of soil water content and atmospheric conditions on leaf water potential in cv. ‘Touriga Nacional’ deep-rooted vineyards.Crossref | GoogleScholarGoogle Scholar |
Rodriguez-Dominguez CM, Buckley TN, Egea G, de Cires A, Hernandez-Santana V, Martorell S, Diaz-Espejo A (2016) Most stomatal closure in woody species under moderate drought can be explained by stomatal responses to leaf turgor. Plant, Cell & Environment 39, 2014–2026.
| Most stomatal closure in woody species under moderate drought can be explained by stomatal responses to leaf turgor.Crossref | GoogleScholarGoogle Scholar |
Schultz HR (1996) Water relations and photosynthetic responses of two grapevine cultivars of different geographical origin during water stress. Acta Horticulturae 427, 251–266.
| Water relations and photosynthetic responses of two grapevine cultivars of different geographical origin during water stress.Crossref | GoogleScholarGoogle Scholar |
Schultz HR (2003) Differences in hydraulic architecture account for near-isohydric and anisohydric behaviour of two field-grown Vitis vinifera L. cultivars during drought. Plant, Cell & Environment 26, 1393–1405.
| Differences in hydraulic architecture account for near-isohydric and anisohydric behaviour of two field-grown Vitis vinifera L. cultivars during drought.Crossref | GoogleScholarGoogle Scholar |
Schultz HR, Matthews MA (1993) Growth, osmotic adjustment, and cell-wall mechanics of expanding grape leaves during water deficits. Crop Science 33, 287–294.
| Growth, osmotic adjustment, and cell-wall mechanics of expanding grape leaves during water deficits.Crossref | GoogleScholarGoogle Scholar |
Shackel KA (2007) Water relations of woody perennial plant species. Journal International des Sciences de la Vigne et du Vin 41, 121–129.
Shackel KA, Matthews MA, Morrison JC (1987) Dynamic relation between expansion and cellular turgor in growing grape (Vitis vinifera L.) leaves. Plant Physiology 84, 1166–1171.
| Dynamic relation between expansion and cellular turgor in growing grape (Vitis vinifera L.) leaves.Crossref | GoogleScholarGoogle Scholar | 16665579PubMed |
Shellie KC, Bowen P (2014) Isohydrodynamic behavior in deficit-irrigated Cabernet Sauvignon and Malbec and its relationship between yield and berry composition. Irrigation Science 32, 87–97.
| Isohydrodynamic behavior in deficit-irrigated Cabernet Sauvignon and Malbec and its relationship between yield and berry composition.Crossref | GoogleScholarGoogle Scholar |
Silim S, Nash R, Reynard D, White B, Schroeder W (2009) Leaf gas exchange and water potential responses to drought in nine poplar (Populus spp.) clones with contrasting drought tolerance. Trees 23, 959–969.
| Leaf gas exchange and water potential responses to drought in nine poplar (Populus spp.) clones with contrasting drought tolerance.Crossref | GoogleScholarGoogle Scholar |
Sinclair TR, Hammer GL, van Oosterom EJ (2005) Potential yield and water-use efficiency benefits in sorghum from limited maximum transpiration rate. Functional Plant Biology 32, 945–952.
| Potential yield and water-use efficiency benefits in sorghum from limited maximum transpiration rate.Crossref | GoogleScholarGoogle Scholar |
Soar CJ, Dry PR, Loveys BR (2006a) Scion photosynthesis and leaf gas exchange in Vitis vinifera L. cv. Shiraz: mediation of rootstock effects via xylem sap ABA. Australian Journal of Grape and Wine Research 12, 82–96.
| Scion photosynthesis and leaf gas exchange in Vitis vinifera L. cv. Shiraz: mediation of rootstock effects via xylem sap ABA.Crossref | GoogleScholarGoogle Scholar |
Soar CJ, Speirs J, Maffei SM, Penrose AB, McCarthy MG, Loveys BR (2006b) Grape vine varieties Shiraz and Grenache differ in their stomatal response to VPD: apparent links with ABA physiology and gene expression in leaf tissue. Australian Journal of Grape and Wine Research 12, 2–12.
| Grape vine varieties Shiraz and Grenache differ in their stomatal response to VPD: apparent links with ABA physiology and gene expression in leaf tissue.Crossref | GoogleScholarGoogle Scholar |
Soar CJ, Collins MJ, Sadras VO (2009) Irrigated Shiraz vines (Vitis vinifera) upregulate gas exchange and maintain berry growth in response to short spells of high maximum temperature in the field. Functional Plant Biology 36, 801–814.
| Irrigated Shiraz vines (Vitis vinifera) upregulate gas exchange and maintain berry growth in response to short spells of high maximum temperature in the field.Crossref | GoogleScholarGoogle Scholar |
Tardieu F, Simonneau T (1998) Variability among species of stomatal control under fluctuating soil water status and evaporative demand: modelling isohydric and anisohydric behaviours. Journal of Experimental Botany 49, 419–432.
| Variability among species of stomatal control under fluctuating soil water status and evaporative demand: modelling isohydric and anisohydric behaviours.Crossref | GoogleScholarGoogle Scholar |
R Core Team (2018) ‘R: A language and environment for statistical computing.’ (R Foundation for Statistical Computing: Vienna, Austria) Available at http://www.R-project.org/ [Verified 12 May 2019].
Tyree MT, Sperry JS (1988) Do woody plants operate near the point of catastrophic xylem dysfunction caused by dynamic water stress? Plant Physiology 88, 574–580.
| Do woody plants operate near the point of catastrophic xylem dysfunction caused by dynamic water stress?Crossref | GoogleScholarGoogle Scholar | 16666351PubMed |
Wickham H (2016) ‘ggplot2: Elegant graphics for data analysis.’ (Springer-Verlag: New York)
Williams LE (2010) Interaction of rootstock and applied water amounts at various fractions of estimated evapotranspiration (ETc) on productivity of Cabernet Sauvignon. Australian Journal of Grape and Wine Research 16, 434–444.
| Interaction of rootstock and applied water amounts at various fractions of estimated evapotranspiration (ETc) on productivity of Cabernet Sauvignon.Crossref | GoogleScholarGoogle Scholar |
Williams LE (2012) Effects of applied water amounts at various fractions of evapotranspiration (ETc) on leaf gas exchange of Thompson Seedless grapevines. Australian Journal of Grape and Wine Research 18, 100–108.
| Effects of applied water amounts at various fractions of evapotranspiration (ETc) on leaf gas exchange of Thompson Seedless grapevines.Crossref | GoogleScholarGoogle Scholar |
Williams LE (2017) Physiological tools to assess vine water status for use in vineyard irrigation management: review and update. Acta Horticulturae 1157, 151–166.
| Physiological tools to assess vine water status for use in vineyard irrigation management: review and update.Crossref | GoogleScholarGoogle Scholar |
Williams LE, Araujo FJ (2002) Correlations among predawn leaf, midday leaf, and midday stem water potential and their correlations with other measures of soil and plant water status in Vitis vinifera. Journal of the American Society for Horticultural Science 127, 448–454.
| Correlations among predawn leaf, midday leaf, and midday stem water potential and their correlations with other measures of soil and plant water status in Vitis vinifera.Crossref | GoogleScholarGoogle Scholar |
Williams LE, Ayars JE (2005) Grapevine water use and the crop coefficient are linear functions of the shaded area measured beneath the canopy. Agricultural and Forest Meteorology 132, 201–211.
| Grapevine water use and the crop coefficient are linear functions of the shaded area measured beneath the canopy.Crossref | GoogleScholarGoogle Scholar |
Williams LE, Baeza P (2007) Relationships among ambient temperature and vapor pressure deficit and leaf and stem water potentials of fully irrigated, field-grown grapevines. American Journal of Enology and Viticulture 58, 173–181.
Williams LE, Grimes DW, Phene CJ (2010a) The effects of applied water at various fractions of measured evapotranspiration on reproductive growth and water productivity of Thompson Seedless grapevines. Irrigation Science 28, 233–243.
| The effects of applied water at various fractions of measured evapotranspiration on reproductive growth and water productivity of Thompson Seedless grapevines.Crossref | GoogleScholarGoogle Scholar |
Williams LE, Grimes DW, Phene CJ (2010b) The effects of applied water at various fractions of measured evapotranspiration on water relations and vegetative growth of Thompson Seedless grapevines. Irrigation Science 28, 221–232.
| The effects of applied water at various fractions of measured evapotranspiration on water relations and vegetative growth of Thompson Seedless grapevines.Crossref | GoogleScholarGoogle Scholar |
Williams LE, Baeza P, Vaughn P (2012) Midday measurements of leaf water potential and stomatal conductance are highly correlated with daily water use of Thompson Seedless grapevines. Irrigation Science 30, 201–212.
| Midday measurements of leaf water potential and stomatal conductance are highly correlated with daily water use of Thompson Seedless grapevines.Crossref | GoogleScholarGoogle Scholar |
Wood SN (2017) ‘Generalized additive models: an introduction with R.’ (Chapman and Hall: London)