

Size-selective predation by three estuarine zooplanktivorous fish species
Hayden T. Schilling A B * , James A. Smith A B , Jason D. Everett A B C , Daniel P. Harrison B D E and Iain M. Suthers A B
A B * , James A. Smith A B , Jason D. Everett A B C , Daniel P. Harrison B D E and Iain M. Suthers A B
A Centre for Marine Science and Innovation, UNSW Sydney, NSW 2052, Australia.
B Sydney Institute of Marine Science, Mosman, NSW 2088, Australia.
C School of Mathematics and Physics, The University of Queensland, Saint Lucia, Qld 4072, Australia.
D Marine Studies Institute, School of Geosciences, University of Sydney, NSW 2006, Australia.
E National Marine Science Centre, Southern Cross University, Coffs Harbour, NSW 2450, Australia.
Marine and Freshwater Research 73(6) 823-832 https://doi.org/10.1071/MF21344
Submitted: 3 December 2021 Accepted: 12 March 2022 Published: 4 May 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial 4.0 International License (CC BY-NC)
Abstract
Context: Zooplanktivorous fish are a key link between abundant zooplankton and higher trophic levels but the foraging behaviour of zooplanktivorous fish is not fully understood. Selective feeding behaviours have been observed, with many species of planktivorous fish targeting certain species and sizes of zooplankton for prey. However, why certain size classes of zooplankton are preferred remains unclear.
Aim: This study investigated prey selection by three zooplanktivorous fish species through the lens of optimal foraging theory.
Methods: We assessed the size structure of zooplankton in the environment and compared this with the size distribution of zooplankton in gut contents from three zooplanktivorous fish.
Key results: The targeted prey size of Atypichthys strigatus and Scorpis lineolata aligns with the prey size classes in the environment that contain the highest overall biomass. Trachurus novaezelandiae showed little evidence of targeting these size classes.
Conclusions: These prey sizes therefore represent the most efficient prey to target because the return on foraging effort is greatest. By contrast, T. novaezelandiae showed only an underselection of large and small prey.
Implications: By incorporating this information on this key trophic link between zooplankton and fish, ecosystem models could better resolve the size dependant predation, particularly in size-based models.
Keywords: Atypichthys strigatus, diet, estuarine ecosystem, gut contents, laser optical plankton counter, optimal foraging theory, planktivory, prey choice, Scorpis lineolata, Trachurus novaezelandiae.
Introduction
Optimal foraging theory is a broad framework that seeks to understand foraging behaviour, including predation that transfers biomass from lower to higher trophic levels. Underlying this theory is the assumption that an animal will make foraging decisions that maximise the amount of energy ingested, while minimising the energy used during feeding (Pyke et al. 1977). Although sometimes criticised for being too simple and not representative of the natural environment (Pierce and Ollason 1987), this theory continues to provide a useful framework for exploring foraging behaviour and has been used recently to make and test predictions about foraging for a range of taxa, including marine mammals (Foo et al. 2016; Tyson et al. 2016), birds (Hernández-Pliego et al. 2017), lions (Barnardo et al. 2020) and fish (Thygesen et al. 2016).
Marine ecosystems are strongly structured by size (Sheldon et al. 1972; Andersen et al. 2016; Hatton et al. 2021), resulting in small prey being highly abundant compared with larger prey. Yet, small prey contain less biomass per individual and may be harder to detect than are larger prey (Hansen et al. 2013). This presents marine predators with a choice of whether to feed on the abundant easy to catch prey or target larger prey?
Zooplanktivorous fish are a key trophic link between abundant zooplankton and larger predators. On temperate rocky reefs, zooplankton can support over 50% of the fish community biomass, with much of this flowing through small zooplanktivorous fish (Truong et al. 2017; Goddard et al. 2022). The foraging strategy of zooplanktivorous fish presents an interesting test of optimal foraging theory. Zooplankton are highly abundant (often >1000 individuals m−3) and small zooplankton are often an order of magnitude more abundant than large zooplankton (Sheldon et al. 1972; Heneghan et al. 2016), meaning that the chance of a zooplanktivorous fish encountering small prey is much greater than is their chance of encountering large prey. However, a large copepod can contain up to 15 times more biomass than a small copepod (Chisholm and Roff 1990), which may make large zooplankton a preferential prey source. This creates a situation where zooplanktivorous fish could feed randomly on the basis of encounter rates of zooplankton (commonly known as filter feeding), essentially eating mostly smaller zooplankton or they could target larger zooplankton (particulate feeding) if the trade-off in terms of biomass return for energy used in searching for and capturing the larger prey is favourable. If consuming the greatest amount of biomass for the least effort is the goal of zooplanktivorous fish, then perhaps their feeding strategy is mediated not by the abundance of different prey but by the biomass of each prey type in the environment. It is plausible that they would target the prey type with the highest biomass in the environment, rather than abundance. Some species have been observed to switch between filter-feeding and particulate-feeding behaviours in response to changes in prey density (Jansen et al. 2019). A similar but alternative theory may be the targeting of calories rather as biomass as the calories are a direct measure of energy (Cumminns and Wuycheck 1971; Balogh et al. 2022). This is most likely if the different prey types have large differences in calorific content but not biomass.
Prey selectivity of zooplankton by zooplanktivorous fish has previously been explored, with a focus on mouth size and gill raker dimensions proving mixed results. Although mouth size is important for small fish (<30 mm), fish are quickly capable of feeding on almost all zooplankton and, at larger fish sizes, the limiting factor becomes handling time and capturing prey (Wanzenbock 1995). Similarly, gill raker size has been shown to not be a limiting factor on the capture of small particles, although it may reduce the efficiency of capture, leading to increased relative capture of larger particles (Langeland and Nøst 1995; Budy et al. 2005). Overall, prey selectivity in zooplanktivorous fish is not simply driven by morphology, but there is also active selection of different size prey, particularly for larger zooplanktivorous fish, which have the ability to capture all sizes of prey and can significantly alter their own behaviour while foraging (Tanaka et al. 2006).
The goal of our study was to explore the foraging strategy of three common zooplanktivorous fish in Sydney Harbour. To investigate selective feeding, we measured the size-structured zooplankton abundance and biomass in the environment over the summer period, and compared this with the size-structured diet of planktivorous fish collected over the same period. The specific aims were to (1) examine the average size-structured zooplankton abundance and biomass in Sydney Harbour over a 3-month period, (2) determine the prey size and diet composition of three planktivorous estuarine fish, and (3) compare the prey size of the gut contents with the size structure of zooplankton available in the water to quantify size-selective predation.
Materials and methods
Zooplankton sampling
Zooplankton were sampled in the lower reach of Sydney Harbour between November 2013 and February 2014 (summer) at three sites (Site 1: −33.834°, 151.278°, Site 2: −33.839°, 151.277°, Site 3: −33.849°, 151.266°), along the southern shore close to the mouth of the estuary and the dominant tidal flow. The total distance between sites was 2.1 km. All sites were sampled in the morning on 10 days, during five ebb and five flood tides (sampled 2–3 h after the predicted high or low tide). At each site, three horizontal replicate plankton tows were made with a 40-cm diameter, 100-µm mesh net at 1-m depth. A mechanical flowmeter (Model 2030R, General Oceanics Inc., Miami, FL, USA) was attached to the net to calculate the sampling volume. A detailed description of the Sydney Harbour ecosystem is available in Johnston et al. (2015).
The zooplankton size distribution and biomass from all towed samples was determined using a laboratory-based laser optical plankton counter (LOPC; Herman et al. 2004), coupled to a pump system (Moore and Suthers 2006). When a particle passed through the beam of the LOPC, the attenuance of light was detected and recorded as the corresponding equivalent spherical diameter (ESD) of the particle. The zooplankton size was classified into a size-frequency distribution with 30-µm bins. Only particles between 300 and 3000 µm ESD were included, because particles outside this size range were unlikely to be sampled accurately by the plankton net (Moore and Suthers 2006). Zooplankton biomass was calculated from the volume of a prolate spheroid (ratio of 1:3, width:length) and the specific gravity of water (Suthers et al. 2006; Garcia et al. 2022).
Because the goal of this study was to investigate foraging behaviour of zooplanktivores rather than variation in the zooplankton community, all zooplankton samples were averaged together to provide a representative zooplankton snapshot over the whole sampling period. This aligns with the fish collection discussed below.
Fish gut content sampling
Over the same time period as the zooplankton sampling, Atypichthys strigatus (Günther, n = 17), Trachurus novaezelandiae (Richardson, n = 24) and Scorpis lineolata (Kner, n = 22) were collected either by using unbaited hook-and-line or spearfishing from the study area. These methods have been used successfully in other studies to sample these species while avoiding contamination of gut contents with bait (Gaston and Suthers 2004; Champion et al. 2015). On the basis of previous research, these species are suspected zooplanktivores and were observed to be the most abundant around our sampling sites. It was confirmed that these species are some of the most abundant small fish in Sydney Harbour by using data from fish surveys undertaken by the Reef Life Survey (Edgar and Stuart-Smith 2014). Individual fish were immediately placed on ice and later frozen, until dietary analysis took place. Fish were collected throughout the sampling period, irrespective of tide, and although fish were not collected evenly in space, all were collected within 500 m of the zooplankton sampling sites. In all, 58 of the 63 fish (92%) were collected on the same day as the zooplankton samples, with five individuals of A. strigatus being collected opportunistically on different days.
The gut contents of each fish were weighed and prey items were identified to a coarse taxonomic resolution. The fullness and percentage (by volume) of plant matter, zooplankton and unidentifiable material were recorded for each gut. Because it was not feasible to use the LOPC for partially digested gut contents, the size distribution (ESD) of zooplankton in the gut contents was manually determined from the length and width of zooplankton, to compare with the size distribution of zooplankton in the water column (the LOPC data). From each gut, a random sample of each zooplankton taxonomic group was photographed using a Leica M80 Microscope with Leica Application Suite (ver. 4.4, Leica Microsystems GmbH, Wetzlar, Germany). ImageJ (ver. 1.48, see https://imagej.nih.gov/ij/; Schneider et al. 2012) was used to measure the length (l, mm) and width (w, mm) of each prey item in the photographs. Length and width measurements were converted into an ESD (µm) by assuming the shape of an ellipsoid and using the following equation:
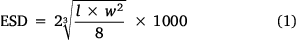
To determine the size range of zooplankton having the highest incidence of consumption by estuarine planktivorous fish, the ESD measurements of identified prey within the fish guts were compiled into frequency histograms for each fish (30-µm bins; corresponding to those of the LOPC). The prey-size distributions of all individuals from each species were then averaged to obtain an average prey-size distribution for each species.
Data analysis
All analysis was conducted using R (ver. 4.0.2, R Foundation for Statistical Computing, Vienna, Austria). A split-plot ANOVA was used to test for differences in total zooplankton biomass between tides and sites, with tide and site as fixed factors and day as a random factor. It was a split-plot design because only one level of tide (ebb or flood) was sampled per day (the ‘plot’). The zooplankton biomass was log10-transformed to meet the assumptions of normality and homogeneity of variance.
To assess selective predation in our zooplanktivorous fish, we used a simulation approach similar approach to Chappell and Smith (2016). We generated three null models of expected prey distributions under the assumption that non-selective feeding would result in prey-size distributions reflecting the zooplankton distribution in the environment. There was one model for each species. Using the abundance (%) of each 30 µm ESD zooplankton size class in the water, we created a simulated population of known size distribution. From this population, we then drew 2000 random samples of x prey items, where x represents the average number of prey items found in each species gut. From these 2000 samples, we generated a mean size-frequency distribution with a 95% confidence interval. We then assessed size-selective predation by comparing the observed predation rates for each size class with the null predictions. If a size class occurred in the fish guts more frequently than predicted by the null simulation, this suggested that this size class was disproportionately preyed on. To estimate the magnitude of the selectivity, we again followed Chappell and Smith (2016) and present the proportional effect (PE) ratio, which is the ratio of observed predation to the expected predation. A PE < 1 signifies underselection, whereas a PE > 1 signifies overselection. Compared with traditional selectivity indices such as those of Chesson (1978), the simulation approach allows for robust analyses of low sample sizes and provides a benchmark of null selection to compare against (Chappell and Smith 2016).
Results
Zooplankton biomass
In total, 86 plankton tows were analysed using the LOPC. Four samples, each from a different site or day, contained too much gelatinous material for accurate analysis; so, they were excluded. The zooplankton biomass varied over an order of magnitude both among days and within days among sites. The smallest zooplankton biomass was recorded at Site 1 with 106.0 mg m−3 (25 November 2013), and the largest was 1722.2 mg m−3, at Site 3 (27 February 2014). The largest range in zooplankton biomass across the three sites on a single day was 1120.0 mg m−3 (27 February 2014). No significant difference in zooplankton biomass was found between ebb and flood tides (ANOVA: F1,8 = 1.17, P = 0.31), but Site 3 contained significantly more zooplankton biomass than did Site 1 across all tides (ANOVA: F2,72 = 5.48, P < 0.01, Supplementary Fig. S1). No significant interaction was found between tide and site (ANOVA: F2,72 = 0.83, P = 0.44).
Despite this variation in total biomass among samples, when the percentage biomass and abundances in each size class were investigated, consistent patterns were observed and we present an overall average zooplankton distribution, which shows consistent declines in abundance with size, and a peak in biomass between 495 and 705 µm ESD (Fig. 1). Among sites, there were only minor differences in the biomass percentage size distributions, with Site 3 having a lower percentage biomass than the other sites in the small bins (<375 µm ESD) and Site 1 having a slightly more even distribution of biomass (a lower peak between 495 and 705 µm ESD; Supplementary Fig. S1).

|
Fish diets
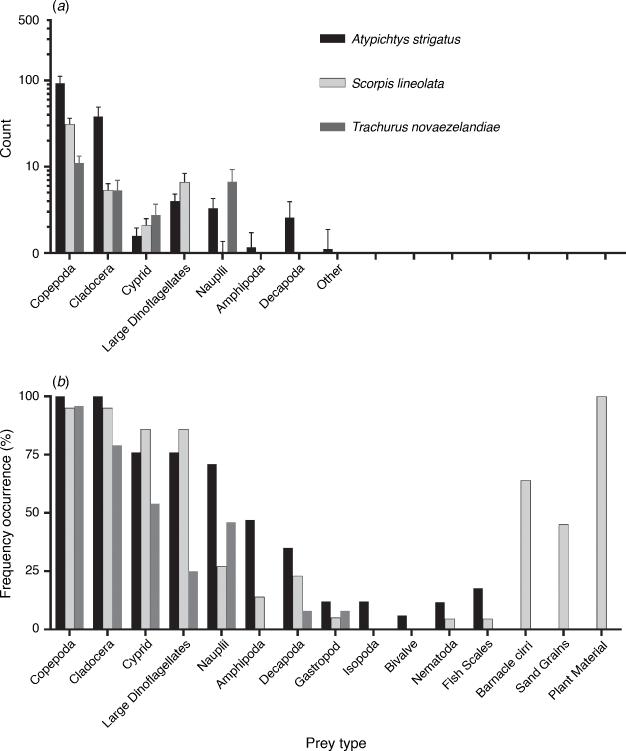
In total, 4140 prey items were identified in the guts of 17 Atypichthys strigatus, 22 Scorpis lineolata and 24 Trachurus novazelandiae individuals. No guts were empty, although seven contained fewer than ten identifiable prey items. A. strigatus, T. novaezelandiae and S. lineolata all consumed zooplankton. In A. strigatus and T. novaezelandiae, zooplankton comprised 100% of the identifiable gut contents. In S. lineolata, zooplankton comprised 40%, with plant matter making up the other 60%. Copepods were the most abundant prey items for all species (Fig. 2a). They were found in 97% of all guts (Fig. 2b) and represented 64, 66 and 41% of all prey items by count in A. strigatus, S. lineolata and T. novaezelandiae respectively. A. strigatus had the greatest average number of prey items in their guts (145.1 ± 21.4 s.e.), followed by S. lineolata (46.9 ± 6.8 s.e.), and T. novaezelandiae had the fewest (26.8 ± 4.4 s.e.; Table 1). S. lineolata was the only species to consume plant material, sand grains or barnacle cirri. T. novaezelandiae had the smallest species richness in its diet, containing eight prey types, compared with 13 and 14 prey types for A. strigatus and S. lineolata respectively.
|

|
Selective predation
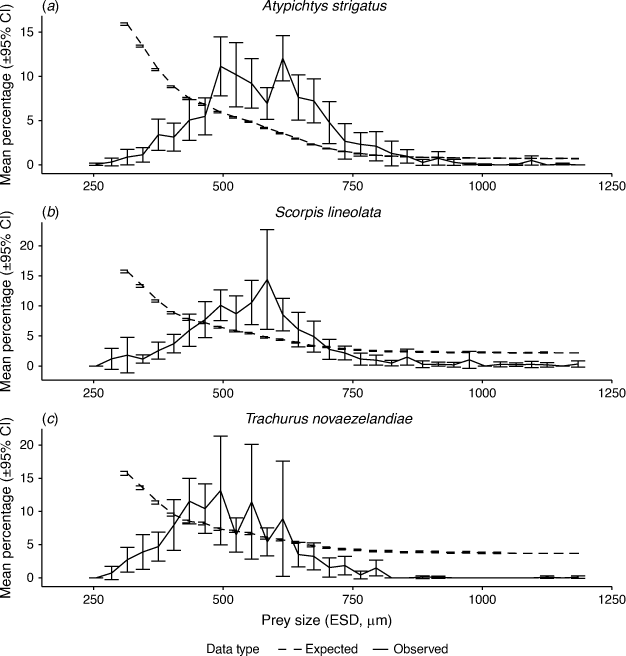
In total, 1997 prey items from gut contents were measured and converted to an equivalent spherical diameter to be comparable with the zooplankton measurements from the LOPC. The three species of fish showed a similar pattern of prey sizes in their guts, although T. novaezelandiae contained smaller prey (Figs 3, 4). The smallest size class of prey was 240–270 µm ESD, with only 0.06% (±0.02 s.d.) of prey in A. strigatus and this size class was not observed in the other two species. This smallest size class was smaller than that resolved by the LOPC, so was not included in comparisons with zooplankton in the environment. The largest size class containing prey was 2940–2970 µm ESD, containing only a single prey item, observed in the gut of a T. novaezelandiae. The median prey size classes of each species were as follows: A. strigatus, 540–570 µm ESD; S. lineolata, 540–570 µm ESD; and T. novaezelandiae, 480–510 µm ESD.
|
|
Strong evidence of size-selective predation was found for A. strigatus and S. lineolata. When compared with the null model of expected diet proportions based on the assumption of random feeding and the observed size structure in the environment, all species showed an underselection of prey in both small (<480 µm ESD for A. strigatus, <450 µm ESD for S. lineolata and <420 µm ESD for T. novaezelandiae) and large (>870 µm ESD for A. strigatus, >690 µm ESD for S. lineolata and >630 µm ESD for T. novaezelandiae) size classes (Fig. 3, Supplementary Table S1). This corresponded to strong evidence of an overselection of medium size particles in A. strigatus (480–840 µm ESD) and S. lineolata (480–690 µm ESD). By contrast, only weak evidence of selectivity was evident in T. novaezelandiae, where the small and large prey underselected, whereas the moderate-size prey showed only weak evidence of selectivity, although this could be due to the low average number of prey items within their guts creating large 95% confidence intervals (Fig. 3, Supplementary Table S1).
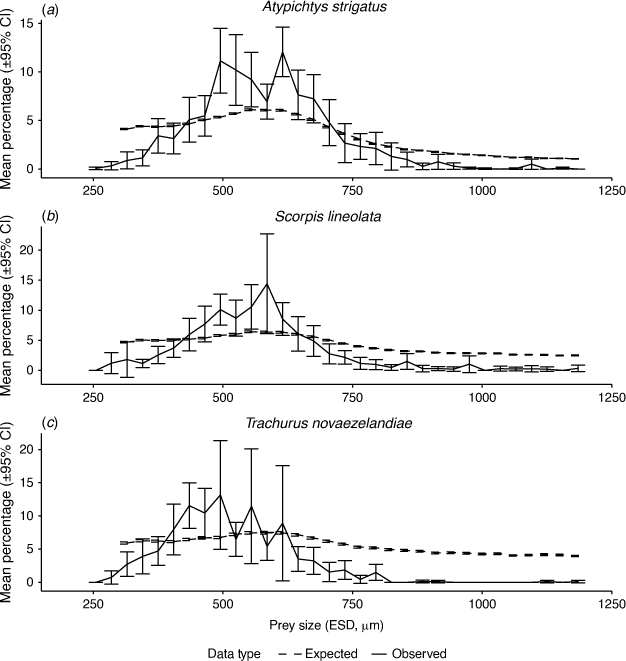
When compared with the null model of expected diet proportions calculated using the biomass of each size class in the environment rather than the abundance, the evidence for size-selective feeding was reduced and our observed prey sizes aligned more closely with expectation for A. strigatus and S. lineolata (Fig. 4). Whereas there continued to be some evidence of underselection at small and large prey sizes, there was vastly reduced evidence of positive selectivity occurring in any size class (Supplementary Table S1). The peak in observed prey size corresponded to the peak in size classes that contained the most biomass in the environment, suggesting that A. strigatus and S. lineolata were likely to be targeting available biomass. By contrast, T. novaezelandiae showed slightly more evidence of selectivity in the biomass model than did the abundance model, with the peak in size classes consumed being smaller than would be expected on the basis of available biomass, suggesting that T. novaezelandiae was likely to be feeding on the basis of abundance, not biomass.
Discussion
This study showed evidence of size-selective predation occurring in three estuarine zooplanktivorous fish, with two species (A. strigatus and S. lineolata) showing strong evidence, whereas T. novaezelandiae showed only weak evidence of size-selective predation. By comparing the observed prey-size compositions with expected prey-size compositions on the basis of feeding relative to environmental abundance and biomass in each size class, we propose that size-selective predation is driven by the increased amount of biomass available in the environment for preferred size classes relative to the other size classes. Our finding supports the theory of optimal foraging theory. Because the goal of foraging is to consume the greatest biomass for the least effort, the size classes with the greatest biomass in them represent the most ‘profitable’ food sources. Understanding the foraging decisions made by planktivorous fish is vital because they often link zooplankton as an abundant resource with the fisheries typically comprising higher trophic levels (Pikitch et al. 2014).
Fish planktivory
Previous studies have defined T. novaezelandiae as a planktivore (Kingsford 1989; Bulman et al. 2001; Dawson et al. 2020), whereas A. strigatus and S. lineolata have previously been defined as piscivores (Bulman et al. 2001) or planktivores (Kingsford 1989; Glasby and Kingsford 1994; Champion et al. 2015). However, this study found only evidence of planktivory with some benthic foraging only for A. strigatus and omnivorory for S. lineolata. It is likely that A. strigatus and S. lineolata may have a flexible diet that can vary with ontogeny and in both time and space, particularly in offshore locations such as those in Bulman et al. (2001). The classification of T. novaezelandiae is consistent with other members of the Trachurus genus (Tanaka et al. 2006).
There is strong evidence for prey-size selection occurring in all three species. Prey smaller than 465 µm ESD and larger than 900 µm ESD were found in the gut contents significantly less often than would be expected on the basis of their environmental abundance. This was matched with significantly more prey of a moderate size (480–~780 µm ESD) being observed in the gut contents of both A. strigatus and S. lineolata than expected on the basis of abundance. Owing to low numbers of prey in T. novaezelandiae gut contents, there was insufficient power to detect any positive prey selection and, instead, the contents matched the expected consumption of moderate-sized prey. There was variation among species, with A. strigatus showing the strongest evidence of prey selectivity while also consuming a higher proportion of larger prey (>705 µm ESD) than do the other species. Whereas there are no comparable studies of prey-size selection for T. novaezelandiae and S. lineolata, it has previously been observed that, in coastal environments, A. strigatus had a preference for slightly larger zooplankton (Champion et al. 2015). This may be because in the offshore location, the available zooplankton were also larger than they were in the estuarine samples in the current study (Champion et al. 2015).
When compared with biomass available in each size class of zooplankton prey (rather than raw abundance), the observed prey-size distributions were a much closer match to the expected distributions. There continued to be an underselection of small and large particles, but the peak in prey size consumed closely matched the size classes of zooplankton in the environment containing the most zooplankton. Avoidance of small prey as an active choice was demonstrated previously in a mesocosm experiment, which demonstrated that turbidity was not an influence on selection by planktivorous bluegill sunfish (Gardner 1981). The present study suggests that the reason for this avoidance may be that the energetic costs of capturing the small prey are not as efficient as when targeting the size classes with the most biomass. In the future, it would be useful to investigate whether prey selectivity changes with the ontogeny of the predator because this has been observed in larval and juvenile yellow perch that switch from a high capture-efficiency technique to a lower capture but higher biomass-return strategy as they mature (Graeb et al. 2004).
As our prey-size measurements for the environmental zooplankton and gut contents were obtained using different methodologies (LOPC vs manual sizing), it is possible that there could be a methodological bias between the measurements. The LOPC has been rigorously validated and used in many studies and has been shown to provide accurate measurements across the size range observed in our study (Herman et al. 2004; Herman and Harvey 2006). By contrast, manual measurements are variable in method and may be subject to bias, particularly because zooplankton in gut contents are not in pristine condition. Previous manual measurements of copepods have shown to be highly accurate (within 1% accuracy), giving us confidence in our method (Araoz 1991). Our measurement method followed that of Skjoldal et al. (2013) and because the majority of zooplankton prey observed in this study were typical copepods with an elliptical shape (Araoz 1991), our method of calculating their volume is valid for most prey items, although it does ignore appendages, which had often fallen off. This may result in a small underestimation of size; however, because the appendages contain very small amounts of total volume and, therefore, biomass (<5%), it is reasonable to assume that our measurements and prey size estimate would be within 5% accuracy and any variation around this would not change the interpretations of the patterns observed in this study.
Copepods were the most abundant prey item in the gut contents of all three species, followed by cladocerans for A. strigatus and T. novaezelandiae and large dinoflagellates for S. lineolata (along with plant material), with other prey types being almost an order of magnitude less abundant. Copepods and cladocerans have very similar calorific contents (Cumminns and Wuycheck 1971), suggesting that the selection in this case was most likely driven by the available biomass and search time trade-off (which will correlate with available calories).
The overlapping prey-size range also provides insight into the niche partitioning occurring in Sydney Harbour; although all three of our studied species consume zooplankton of the same size, they occur in different habitats and are consuming the zooplankton in different parts of the estuary. A. strigatus is an extremely reef-associated zooplanktivore, S. lineolata is also reef associated but also eats plants, and T. novaezelandiae is a zooplanktivore but is less associated with reefs, particularly within estuarine environments.
Zooplankton variability
Although not consistent, there was, on average, higher zooplankton biomass at Site 3 (inner site) than at Site 1 (outer site). This suggests that zooplankton may accumulate inside the estuary and not all is discharged on the ebb tide. This accumulation may be due to either estuarine production or retention within the estuary (Avila et al. 2012) or active use of tidal currents by zooplankton (Simons et al. 2006), and further research looking at estuarine gradients in zooplankton biomass and productivity would provide valuable insight in how estuarine and coastal zooplankton support higher trophic levels. The idea of retention is supported by previous research showing that 50% of the water in this lower-estuary region of Sydney Harbour is retained and not exchanged with the ocean for up to 80 days, increasing up to 90% in the inner estuary (Das et al. 2000).
Regardless of the large variation observed in zooplankton biomass and abundance in the environment, once this was standardised to percentage composition of size classes, there was a consistent trend in the proportions of each size class present. This showed that although there are fluctuations in the overall zooplankton abundance, the size structure of the zooplankton community is stable. This stable size structure of the zooplankton community potentially enables the zooplanktivorous fish to match their prey-size preference to the greatest available biomass that occurs in specific size classes.
Conclusions
Increased understanding of lower trophic level predation dynamics will enable ecosystem modellers to better capture predator–prey dynamics within their models. It is now well recognised that modelling zooplankton specifically in ecosystem models is important (Heneghan et al. 2016), and that size-based modelling approaches may offer significant advantages over traditional food-web models (Blanchard et al. 2017), particularly when predators have diverse prey. By empirically matching predator–prey dynamics to specific size classes of zooplankton with a mechanism (biomass availability), our findings should enable more confidence in the modelling of zooplanktivorous fish predation.
This study has both demonstrated size-selective zooplankton predation by estuarine fish and shown that the targeted size range is likely to be preferred because it contains the largest amount of total biomass, although the extent of the selectivity varies by species. This results in the targeted size classes being the most efficient prey source in terms of return and effort during foraging. This has important considerations for zooplanktivorous fish trophic ecology because it highlights how these species do not feed randomly on zooplankton in the water. The information highlighted in this paper will enable the creation of more accurate lower trophic level and size-based ecosystem models.
Data availability
All code and data are available at https://github.com/HaydenSchilling/Estuarine-Size-Selective-Predation.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Declaration of funding
This study was conducted as part of the Sydney Institute of Marine Science Sydney Harbour Research Project, which provided financial assistance through their Seed Funding Program. J. A. Smith and J. D. Everett were funded by the Australian Research Council (LP120100592 and DPI120100728 respectively). D. P. Harrison was funded by an Australian Postgraduate Award.
Supplementary material
Supplementary material is available online.
Acknowledgements
Thanks go to D. Cruz, C. Champion, J. Halstead, A. Ryan, J. Fenton and G. Brook for assistance in the field. Samples were collected under NSW Department of Primary Industries Scientific Collection Permit No P03/0086(F)-8.0, with approval from the UNSW Animal Care and Ethics Committee (ACEC; 12/111A). This is paper number 289 from the Sydney Institute of Marine Science.
References
Andersen, KH, Berge, T, Gonçalves, RJ, Hartvig, M, Heuschele, J, Hylander, S, Jacobsen, NS, Lindemann, C, Martens, EA, Neuheimer, AB, Olsson, K, Palacz, A, Prowe, AEF, Sainmont, J, Traving, SJ, Visser, AW, Wadhwa, N, and Kiørboe, T (2016). Characteristic sizes of life in the oceans, from bacteria to whales. Annual Review of Marine Science 8, 217–241.| Characteristic sizes of life in the oceans, from bacteria to whales.Crossref | GoogleScholarGoogle Scholar | 26163011PubMed |
Araoz, N (1991). Individual biomass, based on body measures, of copepod species considered as main forage items for fishes of the Argentine shelf. Oceanologica Acta 14, 575–580.
Avila, TR, Machado, AA S, and Bianchini, A (2012). Estimation of zooplankton secondary production in estuarine waters: comparison between the enzymatic (chitobiase) method and mathematical models using crustaceans. Journal of Experimental Marine Biology and Ecology 416–417, 144–152.
| Estimation of zooplankton secondary production in estuarine waters: comparison between the enzymatic (chitobiase) method and mathematical models using crustaceans.Crossref | GoogleScholarGoogle Scholar |
Balogh, C, Serfőző, Z, and Kobak, J (2022). Factors determining selective predation of the common carp on quagga versus zebra mussels. Freshwater Biology 67, 619–629.
| Factors determining selective predation of the common carp on quagga versus zebra mussels.Crossref | GoogleScholarGoogle Scholar |
Barnardo, T, Tambling, CJ, Davies, AB, Klein-Snakenborg, S, Asner, GP, le Roux, E, Cromsigt, JPGM, Druce, DJ, and Kerley, GIH (2020). Opportunistic feeding by lions: non-preferred prey comprise an important part of lion diets in a habitat where preferred prey are abundant. Mammal Research 65, 235–243.
| Opportunistic feeding by lions: non-preferred prey comprise an important part of lion diets in a habitat where preferred prey are abundant.Crossref | GoogleScholarGoogle Scholar |
Blanchard, JL, Heneghan, RF, Everett, JD, Trebilco, R, and Richardson, AJ (2017). From bacteria to whales: using functional size spectra to model marine ecosystems. Trends in Ecology & Evolution 32, 174–186.
| From bacteria to whales: using functional size spectra to model marine ecosystems.Crossref | GoogleScholarGoogle Scholar |
Budy, P, Haddix, T, and Schneidervin, R (2005). Zooplankton size selection relative to gill raker spacing in rainbow trout. Transactions of the American Fisheries Society 134, 1228–1235.
| Zooplankton size selection relative to gill raker spacing in rainbow trout.Crossref | GoogleScholarGoogle Scholar |
Bulman, C, Althaus, F, He, X, Bax, NJ, and Williams, A (2001). Diets and trophic guilds of demersal fishes of the south-eastern Australian shelf. Marine and Freshwater Research 52, 537–548.
| Diets and trophic guilds of demersal fishes of the south-eastern Australian shelf.Crossref | GoogleScholarGoogle Scholar |
Champion, C, Suthers, IM, and Smith, JA (2015). Zooplanktivory is a key process for fish production on a coastal artificial reef. Marine Ecology Progress Series 541, 1–14.
| Zooplanktivory is a key process for fish production on a coastal artificial reef.Crossref | GoogleScholarGoogle Scholar |
Chappell, BF, and Smith, KG (2016). Patterns of predation of native reef fish by invasive Indo-Pacific lionfish in the western Atlantic: evidence of selectivity by a generalist predator. Global Ecology and Conservation 8, 18–23.
| Patterns of predation of native reef fish by invasive Indo-Pacific lionfish in the western Atlantic: evidence of selectivity by a generalist predator.Crossref | GoogleScholarGoogle Scholar |
Chesson, J (1978). Measuring preference in selective predation. Ecology 59, 211–215.
| Measuring preference in selective predation.Crossref | GoogleScholarGoogle Scholar |
Chisholm, LA, and Roff, JC (1990). Size–weight relationships and biomass of tropical neritic copepods off Kingston, Jamaica. Marine Biology 106, 71–77.
| Size–weight relationships and biomass of tropical neritic copepods off Kingston, Jamaica.Crossref | GoogleScholarGoogle Scholar |
Cumminns, KW, and Wuycheck, JC (1971). Caloric equivalents for investigations in ecological energetics. SIL Communications, 1953-1996 18, 1–158.
| Caloric equivalents for investigations in ecological energetics.Crossref | GoogleScholarGoogle Scholar |
Das, P, Marchesiello, P, and Middleton, JH (2000). Numerical modelling of tide-induced residual circulation in Sydney Harbour. Marine and Freshwater Research 51, 97–112.
| Numerical modelling of tide-induced residual circulation in Sydney Harbour.Crossref | GoogleScholarGoogle Scholar |
Dawson, G, Suthers, IM, Brodie, S, and Smith, JA (2020). The bioenergetics of a coastal forage fish: importance of empirical values for ecosystem models. Deep-sea Research – II. Topical Studies in Oceanography 175, 104700.
| The bioenergetics of a coastal forage fish: importance of empirical values for ecosystem models.Crossref | GoogleScholarGoogle Scholar |
Edgar, GJ, and Stuart-Smith, RD (2014). Systematic global assessment of reef fish communities by the Reef Life Survey program. Scientific Data 1, 140007.
| Systematic global assessment of reef fish communities by the Reef Life Survey program.Crossref | GoogleScholarGoogle Scholar | 25977765PubMed |
Foo, D, Semmens, JM, Arnould, JPY, Dorville, N, Hoskins, AJ, Abernathy, K, Marshall, GJ, and Hindell, MA (2016). Testing optimal foraging theory models on benthic divers. Animal Behaviour 112, 127–138.
| Testing optimal foraging theory models on benthic divers.Crossref | GoogleScholarGoogle Scholar |
Garcia, V, Schilling, HT, Cruz, D, Hawes, SM, Everett, JD, Roughan, M, Miskiewicz, AG, Pakhomov, EA, Jeffs, A, and Suthers, IM (2022). Entrainment and development of larval fish assemblages in two contrasting cold core eddies of the East Australian Current system. Marine Ecology Progress Series 685, 1–18.
| Entrainment and development of larval fish assemblages in two contrasting cold core eddies of the East Australian Current system.Crossref | GoogleScholarGoogle Scholar |
Gardner, MB (1981). Mechanisms of size selectivity by planktivorous fish: a test of hypotheses. Ecology 62, 571–578.
| Mechanisms of size selectivity by planktivorous fish: a test of hypotheses.Crossref | GoogleScholarGoogle Scholar |
Gaston, TF, and Suthers, IM (2004). Spatial variation in δ13C and δ15N of liver, muscle and bone in a rocky reef planktivorous fish: the relative contribution of sewage. Journal of Experimental Marine Biology and Ecology 304, 17–33.
| Spatial variation in δ13C and δ15N of liver, muscle and bone in a rocky reef planktivorous fish: the relative contribution of sewage.Crossref | GoogleScholarGoogle Scholar |
Glasby, TM, and Kingsford, MJ (1994). Atypichthys strigatus (Pisces: Scorpididae): an opportunistic planktivore that responds to benthic disturbances and cleans other fishes. Australian Journal of Ecology 19, 385–394.
| Atypichthys strigatus (Pisces: Scorpididae): an opportunistic planktivore that responds to benthic disturbances and cleans other fishes.Crossref | GoogleScholarGoogle Scholar |
Goddard, BK, Becker, A, Harasti, D, Smith, JA, Subramaniam, RC, and Suthers, IM (2022). The trophic basis of fish assemblages in temperate estuarine and coastal ecosystems. Marine Biology 169, 19.
| The trophic basis of fish assemblages in temperate estuarine and coastal ecosystems.Crossref | GoogleScholarGoogle Scholar |
Graeb, BDS, Dettmers, JM, Wahl, DH, and Cáceres, CE (2004). Fish size and prey availability affect growth, survival, prey selection, and foraging behavior of larval yellow perch. Transactions of the American Fisheries Society 133, 504–514.
| Fish size and prey availability affect growth, survival, prey selection, and foraging behavior of larval yellow perch.Crossref | GoogleScholarGoogle Scholar |
Hansen, AG, Beauchamp, DA, and Schoen, ER (2013). Visual prey detection responses of piscivorous trout and salmon: effects of light, turbidity, and prey size. Transactions of the American Fisheries Society 142, 854–867.
| Visual prey detection responses of piscivorous trout and salmon: effects of light, turbidity, and prey size.Crossref | GoogleScholarGoogle Scholar |
Hatton, IA, Heneghan, RF, Bar-On, YM, and Galbraith, ED (2021). The global ocean size spectrum from bacteria to whales. Science Advances 7, eabh3732.
| The global ocean size spectrum from bacteria to whales.Crossref | GoogleScholarGoogle Scholar | 34757796PubMed |
Heneghan, RF, Everett, JD, Blanchard, JL, and Richardson, AJ (2016). Zooplankton are not fish: improving zooplankton realism in size-spectrum models mediates energy transfer in food webs. Frontiers in Marine Science 3, 201.
| Zooplankton are not fish: improving zooplankton realism in size-spectrum models mediates energy transfer in food webs.Crossref | GoogleScholarGoogle Scholar |
Herman, AW, and Harvey, M (2006). Application of normalized biomass size spectra to laser optical plankton counter net intercomparisons of zooplankton distributions. Journal of Geophysical Research: Oceans 111, C05S05.
| Application of normalized biomass size spectra to laser optical plankton counter net intercomparisons of zooplankton distributions.Crossref | GoogleScholarGoogle Scholar |
Herman, AW, Beanlands, B, and Phillips, EF (2004). The next generation of optical plankton counter: the laser-OPC. Journal of Plankton Research 26, 1135–1145.
| The next generation of optical plankton counter: the laser-OPC.Crossref | GoogleScholarGoogle Scholar |
Hernández-Pliego, J, Rodríguez, C, Dell’Omo, G, and Bustamante, J (2017). Combined use of tri-axial accelerometers and GPS reveals the flexible foraging strategy of a bird in relation to weather conditions. PLoS One 12, e0177892.
| Combined use of tri-axial accelerometers and GPS reveals the flexible foraging strategy of a bird in relation to weather conditions.Crossref | GoogleScholarGoogle Scholar | 28591181PubMed |
Jansen, T, Post, S, Olafsdottir, AH, Reynisson, P, Óskarsson, GJ, and Arendt, KE (2019). Diel vertical feeding behaviour of Atlantic mackerel (Scomber scombrus) in the Irminger current. Fisheries Research 214, 25–34.
| Diel vertical feeding behaviour of Atlantic mackerel (Scomber scombrus) in the Irminger current.Crossref | GoogleScholarGoogle Scholar |
Johnston, EL, Mayer-Pinto, M, Hutchings, PA, Marzinelli, EM, Ahyong, ST, Birch, G, Booth, DJ, Creese, RG, Doblin, MA, Figueira, W, Gribben, PE, Pritchard, T, Roughan, M, Steinberg, PD, and Hedge, LH (2015). Sydney Harbour: what we do and do not know about a highly diverse estuary. Marine and Freshwater Research 66, 1073–1087.
| Sydney Harbour: what we do and do not know about a highly diverse estuary.Crossref | GoogleScholarGoogle Scholar |
Kingsford, MJ (1989). Distribution patters of planktivorous reef fish along the coast of northeastern New Zealand. Marine Ecology Progress Series 54, 13–24.
| Distribution patters of planktivorous reef fish along the coast of northeastern New Zealand.Crossref | GoogleScholarGoogle Scholar |
Langeland, A, and Nøst, T (1995). Gill raker structure and selective predation on zooplankton by particulate feeding fish. Journal of Fish Biology 47, 719–732.
| Gill raker structure and selective predation on zooplankton by particulate feeding fish.Crossref | GoogleScholarGoogle Scholar |
Moore, SK, and Suthers, IM (2006). Evaluation and correction of subresolved particles by the optical plankton counter in three Australian estuaries with pristine to highly modified catchments. Journal of Geophysical Research: Oceans 111, 1–14.
| Evaluation and correction of subresolved particles by the optical plankton counter in three Australian estuaries with pristine to highly modified catchments.Crossref | GoogleScholarGoogle Scholar |
Pierce, GJ, and Ollason, JG (1987). Eight reasons why optimal foraging theory is a complete waste of time. Oikos 49, 111–118.
| Eight reasons why optimal foraging theory is a complete waste of time.Crossref | GoogleScholarGoogle Scholar |
Pikitch, EK, Rountos, KJ, Essington, TE, Santora, C, Pauly, D, Watson, R, Sumaila, UR, Boersma, PD, Boyd, IL, Conover, DO, Cury, P, Heppell, SS, Houde, ED, Mangel, M, Plagányi, É, Sainsbury, K, Steneck, RS, Geers, TM, Gownaris, N, and Munch, SB (2014). The global contribution of forage fish to marine fisheries and ecosystems. Fish and Fisheries 15, 43–64.
| The global contribution of forage fish to marine fisheries and ecosystems.Crossref | GoogleScholarGoogle Scholar |
Pyke, GH, Pulliam, HR, and Charnov, EL (1977). Optimal foraging: a selective review of theory and tests. The Quarterly Review of Biology 52, 137–154.
| Optimal foraging: a selective review of theory and tests.Crossref | GoogleScholarGoogle Scholar |
Schneider, CA, Rasband, WS, and Eliceiri, KW (2012). NIH Image to ImageJ: 25 years of image analysis. Nature Methods 9, 671–675.
| NIH Image to ImageJ: 25 years of image analysis.Crossref | GoogleScholarGoogle Scholar | 22930834PubMed |
Sheldon, RW, Prakash, A, and Sutcliffe, WH (1972). The size distribution of particles in the ocean. Limnology and Oceanography 17, 327–340.
| The size distribution of particles in the ocean.Crossref | GoogleScholarGoogle Scholar |
Simons, RD, Monismith, SG, Johnson, LE, Winkler, G, and Saucier, FJ (2006). Zooplankton retention in the estuarine transition zone of the St Lawrence Estuary. Limnology and Oceanography 51, 2621–2631.
| Zooplankton retention in the estuarine transition zone of the St Lawrence Estuary.Crossref | GoogleScholarGoogle Scholar |
Skjoldal, HR, Wiebe, PH, Postel, L, Knutsen, T, Kaartvedt, S, and Sameoto, DD (2013). Intercomparison of zooplankton (net) sampling systems: results from the ICES/GLOBEC sea-going workshop. Progress in Oceanography 108, 1–42.
| Intercomparison of zooplankton (net) sampling systems: results from the ICES/GLOBEC sea-going workshop.Crossref | GoogleScholarGoogle Scholar |
Suthers, IM, Taggart, CT, Rissik, D, and Baird, ME (2006). Day and night ichthyoplankton assemblages and zooplankton biomass size spectrum in a deep ocean island wake. Marine Ecology Progress Series 322, 225–238.
| Day and night ichthyoplankton assemblages and zooplankton biomass size spectrum in a deep ocean island wake.Crossref | GoogleScholarGoogle Scholar |
Tanaka, H, Aoki, I, and Ohshimo, S (2006). Feeding habits and gill raker morphology of three planktivorous pelagic fish species off the coast of northern and western Kyushu in summer. Journal of Fish Biology 68, 1041–1061.
| Feeding habits and gill raker morphology of three planktivorous pelagic fish species off the coast of northern and western Kyushu in summer.Crossref | GoogleScholarGoogle Scholar |
Thygesen, UH, Sommer, L, Evans, K, and Patterson, TA (2016). Dynamic optimal foraging theory explains vertical migrations of Bigeye tuna. Ecology 97, 1852–1861.
| Dynamic optimal foraging theory explains vertical migrations of Bigeye tuna.Crossref | GoogleScholarGoogle Scholar | 27859170PubMed |
Truong, L, Suthers, IM, Cruz, DO, and Smith, JA (2017). Plankton supports the majority of fish biomass on temperate rocky reefs. Marine Biology 164, 73.
| Plankton supports the majority of fish biomass on temperate rocky reefs.Crossref | GoogleScholarGoogle Scholar |
Tyson, RB, Friedlaender, AS, and Nowacek, DP (2016). Does optimal foraging theory predict the foraging performance of a large air-breathing marine predator? Animal Behaviour 116, 223–235.
| Does optimal foraging theory predict the foraging performance of a large air-breathing marine predator?Crossref | GoogleScholarGoogle Scholar |
Wanzenbock, J (1995). Changing handling times during feeding and consequences for prey size selection of O+ zooplanktivorous fish. Oecologia 104, 372–378.
| Changing handling times during feeding and consequences for prey size selection of O+ zooplanktivorous fish.Crossref | GoogleScholarGoogle Scholar | 28307594PubMed |