

Distribution, site affinity and regional movements of the manta ray, Manta alfredi (Krefft, 1868), along the east coast of Australia
Lydie I. E. Couturier A B G , Fabrice R. A. Jaine B C E , Kathy A. Townsend D , Scarla J. Weeks C E , Anthony J. Richardson B F and Michael B. Bennett AA School of Biomedical Sciences, The University of Queensland, St Lucia, Qld 4067, Australia.
B Climate Adaptation Flagship, CSIRO Marine and Atmospheric Research, Cleveland, Qld 4163, Australia.
C Centre for Spatial Environmental Research, The University of Queensland, St Lucia, Qld 4067, Australia.
D School of Biological Sciences, The University of Queensland, St Lucia, Qld 4067, Australia.
E Global Change Institute, The University of Queensland, St Lucia, Qld 4067, Australia.
F Centre for Applications in Natural Resource Mathematics (CARM), School of Mathematics and Physics, University of Queensland, St Lucia, Qld 4067, Australia.
G Corresponding author. Email: l.couturier@uq.edu.au
Marine and Freshwater Research 62(6) 628-637 https://doi.org/10.1071/MF10148
Submitted: 18 June 2010 Accepted: 12 December 2010 Published: 24 June 2011
Journal Compilation © CSIRO Publishing 2011 Open Access CC BY-NC-ND
Abstract
Despite the increasing tourism interest worldwide for the manta ray, Manta alfredi, very little is known about its biology and ecology. Knowledge of its distribution and movement patterns is important for conservation purposes. Here we describe the distribution, site visitation and movements of M. alfredi along the east coast of Australia. Photographic identification techniques were used to identify individual manta rays at three study sites: Lady Elliot Island, North Stradbroke Island and Byron Bay. Of 388 M. alfredi individuals identified at Lady Elliot Island, 187 (48%) were subsequently re-identified at least once at the same site. In total, 31 individuals were identified at both Lady Elliot Island and North Stradbroke Island (~380 km to the south) and 4 at both Lady Elliot Island and Byron Bay (~500 km to the south). Manta alfredi was present all year around at Lady Elliot Island, although in higher numbers in winter, and was mainly observed at North Stradbroke Island and Byron Bay from mid-spring to mid-autumn. This is the first report of seasonal movements and site affinity for M. alfredi in eastern Australian waters and emphasises the value of photographic identification for monitoring the occurrence of individuals.
Additional keywords: elasmobranch, migration, photographic identification, seasonality.
Introduction
Manta rays (Manta spp.) are large filter-feeding elasmobranch fishes that have a circumglobal distribution (Last and Stevens 2009; Marshall et al. 2009). The placid disposition of these animals and the fact that they can be readily approached by SCUBA divers has resulted in the development of dive-based ecotourism at many sites around the world (Anderson 2002; MacCarthy et al. 2006). Until recently, the genus Manta was considered to be monospecific, with M. birostris having a worldwide distribution. However, a recent taxonomic revision recognised at least two distinct species, M. birostris (Walbaum, 1792) and M. alfredi (Krefft, 1868), and a putative third species, M. cf. birostris (Marshall et al. 2009). The inshore or reef manta ray, M. alfredi, is commonly sighted on the continental shelf, around tropical and subtropical coral and rocky reefs, islands and along coastlines. In contrast, M. birostris is considered to be a more oceanic species that migrates longer distances and is found in cooler waters (Marshall 2008; Marshall et al. 2009). While the large pelagic manta, M. birostris, may occur at certain localities around the coast of Australia, M. alfredi is thought to be more common along the Pacific and Indian Ocean coastal regions of Australia (Whitley 1936; Marshall et al. 2009). Although these two species are morphologically and behaviourally distinct (Marshall et al. 2009), the failure to differentiate them before 2009 has resulted in confusion concerning almost all existing biological information (e.g. White et al. 2006; Dewar et al. 2008; Marshall et al. 2008). Published information is primarily attributed to the original species, M. birostris, although some or even most may refer to M. alfredi. Consequently, many aspects of the biology of M. alfredi, including its population ecology, biology, behaviour and movements are uncertain or unknown.
Manta ray aggregations have been monitored and investigated in various locations around the world, including Western Australia (McGregor et al. 2008), Hawaii (Clark and Laros 2008; Deakos 2010), Japan (Ishihara and Homma 1995; Homma et al. 1999; Yano et al. 1999), Tahiti (De Rosemont 2008), Mozambique (Marshall 2008; Marshall et al. 2008), the Maldives (Kitchen-Wheeler 2010), Indonesia (Dewar et al. 2008), Mexico (Graham et al. 2008; Rubin and Kumli 2008) and Brazil (Luiz et al. 2009). Some of these locations experience regular peaks in manta ray abundance, while others are only visited by the species seasonally. The strong seasonal visitation pattern of manta rays in the Komodo Marine Park, Indonesia, was linked to changes in sea temperature and productivity, both factors being influenced by monsoonal shifts (Dewar et al. 2008). Annual migration of M. alfredi in the Maldives has also been associated with seasonal monsoonal conditions, including changes in zooplankton abundance (Kitchen-Wheeler 2010). Long-term site fidelity by a part of the manta ray population has been reported at both Komodo Marine Park, Indonesia (Dewar et al. 2008) and Yaeyama Islands, Japan (Ishihara and Homma 1995; Homma et al. 1999). At other locations, such as Hawaii, the Maldives and Mozambique, manta rays have been shown to revisit the same site multiple times over several years (Clark and Laros 2008; Graham et al. 2008; Marshall 2008). However, more data are required to determine whether M. alfredi displays true site fidelity or simply moves within a large home range, with individuals periodically resighted at specific monitored locations. We thus refer to these same site visitation patterns as site affinity. The longest movement reported for M. alfredi was between the Yaeyama Islands and Kerama Islands, Japan, with two individuals swimming at least 400 km (Ishihara and Homma 1995; Kashiwagi et al. 2008).
Photographic identification (photo-ID) is a relatively non-invasive approach that has been widely used for over 30 years in cetacean research (e.g. Perkins and Whitehead 1977; Durban et al. 2010). More recently, this technique has been applied successfully to examine aspects of the biology of several elasmobranch species, including the nurse shark Ginglymostoma cirratum (Castro and Rosa 2005), white shark Carcharodon carcharias (Domeier and Nasby-Lucas 2007), grey nurse shark Carcharias taurus (Bansemer and Bennett 2008), zebra shark Stegostoma fasciatum (Dudgeon et al. 2008) and whale shark Rhincodon typus (Speed et al. 2008). Much of the information available on manta ray movements and ecology has been obtained from photo-ID of individuals (e.g. Cerutti et al. 2008; Clark and Laros 2008; Rubin and Kumli 2008; Luiz et al. 2009; Kitchen-Wheeler 2010). Most manta rays have dark skin-pigmentation patterns on their otherwise pale ventral surfaces, which can be used to differentiate individuals. Even melanistic individuals, which are mostly dark grey or black, generally have distinctive white ventral patterning (Marshall et al. 2009; Kitchen-Wheeler 2010). Pigmentation patterns are present in near-term embryos and neonate rays, and are highly likely to be permanent and consistent through time (Homma et al. 1999; Marshall et al. 2008; Kitchen-Wheeler 2010). Many animals bear additional distinctive injuries (e.g. scars, shark bite wounds, shortened tails) that can also be used to differentiate individuals (Kitchen-Wheeler 2010; Marshall and Bennett 2010a).
While confirmed records of M. alfredi landings are limited to coastal waters near Sydney (New South Wales) and off the Whitsunday Islands (Queensland) (Whitley 1932, 1936), the species is considered to occur more broadly along the eastern Australian coast (Marshall et al. 2009). However, information on the distribution, movement and biology of M. alfredi in eastern Australia is effectively lacking.
The present study aimed to examine temporal trends, regional movements and site affinity of M. alfredi along the eastern Australian coast through the application of photo-ID of individual rays. This is the first study to investigate the population of M. alfredi in eastern Australia, and we focussed on three sites regularly visited by the species – Lady Elliot Island (southern Great Barrier Reef, Queensland), North Stradbroke Island (south-east Queensland) and Byron Bay (northern New South Wales).
Methods
Study sites
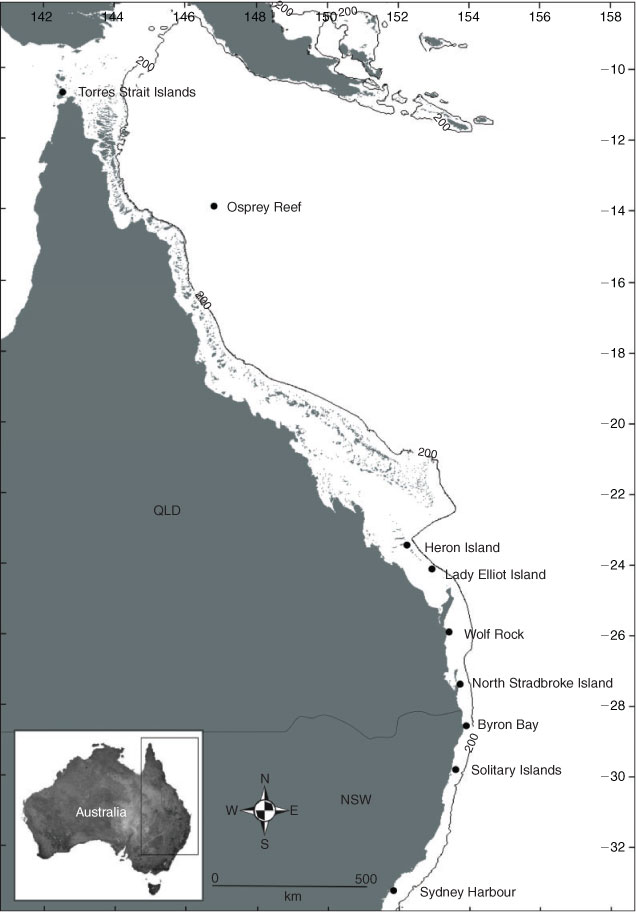
Photographs of M. alfredi were sought from local dive clubs, researchers and tourism operators spanning coastal waters from the Torres Strait Islands (North Queensland) to Sydney (New South Wales) (Fig. 1). The primary study sites for data collection were Lady Elliot Island, North Stradbroke Island and Byron Bay. Lady Elliot Island, the most southern coral cay of the Great Barrier Reef (24°06′S 152°42′E), is an ecotourism destination where manta rays are commonly seen near the surface and around cleaning stations at 8–25 m depth. At North Stradbroke Island, in south-east Queensland (27°25′S 153°32′E), ~380 km to the south of Lady Elliot Island, manta rays are mainly observed being cleaned at depths of 8–15 m around rocky reefs (known locally as The Group), 500 m off the north-eastern tip of the island. Manta rays also occur offshore from Byron Bay in New South Wales at Julian Rocks (28°37′S 153°37′E), ~126 km south of North Stradbroke Island. This rocky reef has a cleaning station at a depth of 8–17 m. Photographs of manta rays were collected over different but overlapping periods in the three locations: January 2007 to July 2010 at Lady Elliot Island, February 2005 to July 2010 at North Stradbroke Island; and February 2009 to June 2010 at Byron Bay. Specific photographic surveys were conducted by us in December 2008, March 2009, June 2009, November 2009, February 2010 and June 2010 at Lady Elliot Island.
|
Data collection
Photographs of manta rays were collected by the authors, dive instructors and recreational divers at Lady Elliot Island, North Stradbroke Island and Byron Bay. In addition, people with extensive local knowledge of manta ray visitation patterns at known sites (i.e. dive instructors and naturalists) were interviewed to identify patterns of occurrence by M. alfredi at these sites. Underwater digital still and video cameras were used while snorkelling and SCUBA diving. For each photograph, the name of the photographer, location and date of image capture were recorded. The species was identified as M. alfredi based on morphological characteristics (Marshall et al. 2009) that included dorsal colouration and patterning, the distribution of spots on the ventral surface and the absence of a caudal spine (Fig. 2).

|
Distinctive spots and more diffusely pigmented patches on the ventral surfaces of animals were used to identify and differentiate individuals. Only good-quality photographs with a clear image of at least the abdominal and inter-branchial areas were used for identification. Distinctive wound marks, presumably from interactions with sharks, were also used to assist in the identification of individuals. Although most manta rays were identified using several photographs of the same sighting event, a single photograph of an individual was considered sufficient for positive identification when the entire ventral side of the body was clearly visible (e.g. Fig. 2b). The sex of individuals was determined by the presence or absence of claspers when the pelvic fins of the animal were clearly apparent on the photographs (Fig. 2d). Female : male ratio data were compared at each study site using Chi-Square Goodness of Fit with correction for continuity. Significant bias was accepted at P < 0.05.
Photo-ID procedures followed Marshall (2008). A composite sketch of the dorsal and ventral surface of each ray was made, showing all spots, wounds or other distinctive characteristics, based on all available images. These sketches, combined with our photographic database, were used to identify each individual ray. Images were matched visually, with every new identification and resighting event double-checked by two observers and approved by a single person (L. I. E. Couturier) before its inclusion in the photographic database. Re-sighting events were defined as the identification of a formerly known individual more than 24 h after it was last seen (Marshall 2008).
Sampling effort was not consistent at all sites, as it was limited by location and timing of the photographers in the water. In particular, the North Stradbroke Island study site was only accessible under relatively calm sea conditions and, consequently, the photographic sampling effort was considerably lower than at Lady Elliot Island. The Byron Bay site was only included in the last year of this study, with the provision of photographs resulting from three recreational divers in this region.
Results
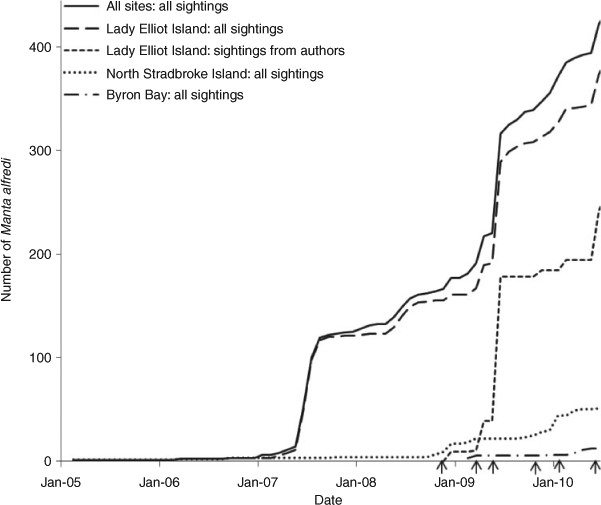
In total, 2874 photographs of M. alfredi were collected for this study, 1537 by the authors and 1337 by 52 contributors from the Torres Strait Islands (North Queensland) to South Solitary Island (New South Wales). From these, 435 individuals were identified between February 2005 and July 2010. Photographs were obtained from eight different sites: Torres Strait Islands; Osprey Reef; Heron Island; Lady Elliot Island; Wolf Rock; North Stradbroke Island; Byron Bay; and South Solitary Island (Fig. 1). The discovery curve based on all photographs collected from all sources shows a large increase in identification of new individuals since the beginning of this study (Fig. 3). It also indicates that we have not sampled the whole M. alfredi population in east Australian coastal waters as the curve did not reach an asymptote. Manta birostris was not observed in any of the photographs from these sites during this study.
|
In total, 2807 suitable photographs of M. alfredi individuals were collected from 890 sighting events at the three main study sites (Lady Elliot Island, North Stradbroke Island and Byron Bay) between 2005 and 2010. Of those, 2405 photographs were collected at Lady Elliot Island (1303 from authors and 1102 from contributors), 353 at North Stradbroke Island (217 from authors and 136 from contributors) and 49 at Byron Bay (all from contributors). From these photographs, 417 individuals were identified. Of this total, 388 (93.0%) individuals comprising 201 females, 152 males and 35 unsexed rays were identified at Lady Elliot Island. The female : male sex ratio at this site was significantly biased towards females (1.32 : 1.0, χ2 = 6.80, d.f. = 1, P < 0.01). About 67% of those individuals were identified by the research team during photographic surveys at this site (Fig. 3). Of the 51 rays identified at North Stradbroke Island (12.2%), there were 23 females, 24 males and 4 unsexed individuals. There was no sex ratio bias at this site (1.04 : 1.0, χ2 = 0.02, d.f. = 1, P > 0.05). Similarly, of the 12 individuals identified at Byron Bay (2.9%), there were 6 females, 5 males and 1 unsexed ray. A sex ratio bias was absent at this site (1.2 : 1, χ2 = 0.09, d.f. = 1, P > 0.05). The discovery curves for those three main sites show different ‘new identification’ rates due to the unequal sampling efforts at each location (Fig. 3).
Of the remaining 67 photographs, all provided by contributors, 11 individuals were identified at Heron Island, 4 at Osprey Reef and 1 each at Wolf Rock and South Solitary Island. There were no re-sightings of these identified manta rays. Photographs collected in the Torres Strait Islands region indicated that the species was M. alfredi, but were not suitable for individual identification purposes.
Photo-ID indicated that the number of M. alfredi individuals that visited waters around Lady Elliot Island peaked in June and July, although they were present in every month of the year (Table 1). This observed variation in manta ray sightings was not the result of changes in ‘diver effort’, as over 60% of dives between July 2008 and April 2010 were conducted in spring and summer (Lady Elliot Island Eco Resort, Water sports statistics 2008–2010). By contrast, M. alfredi was principally sighted at North Stradbroke Island from mid-spring to mid-autumn (October–April), with few sightings during the cooler months (May–September), and from late summer to mid-autumn (February–April) off Byron Bay.

|
Site affinity
Of the 417 individuals identified at the three study sites, 211 (50.6%) were re-sighted at least once at Lady Elliot Island, North Stradbroke Island or Byron Bay. Of the 388 individuals identified at Lady Elliot Island, 188 (48.5%) were re-sighted at least once at this same site 1–1102 days later (Table 2). In addition, of the 51 individuals identified at North Stradbroke Island, 22 (43.1%) were subsequently re-sighted at the same site with an inter-sighting period of 1–741 days. Two individuals were re-sighted 9 times at Lady Elliot Island and three individuals were re-sighted three times at North Stradbroke Island.

|
Movements
Thirty-four manta rays (7.8% of total) were identified at two or three study sites, including 20 females, 13 males and one unsexed ray (Table 3). The female : male sex ratio was not significantly biased (1.54 : 1.0, χ2 = 1.48, d.f. = 1, P > 0.05). All were identified at southern sites during the austral mid-spring to early autumn and 32 out of 34 individuals were only identified at Lady Elliot Island in autumn–winter. Of these 34 manta rays, 31 were sighed at both Lady Elliot Island and North Stradbroke Island, with the time interval between re-sightings ranging from 47 to 1199 days. Return movements between these two sites were demonstrated by 10 individuals (Table 3). Four manta rays were sighted at both Byron Bay and Lady Elliot Island, and one at all three sites (Table 3).

|
Discussion
This study provides the first empirical evidence that the reef manta ray, M. alfredi, occurs in moderately large numbers along the east coast of Australia, and that individual rays are capable of movements over hundreds of kilometres. This information is of management as well as scientific interest as this species is one of the principal targets of recreational dive and ecotourism operations globally (Homma et al. 1999; Anderson 2002; Australian Government 2009; Anderson et al. 2011). In addition, although threats for M. alfredi in eastern Australia are unquantified, the species is likely to be vulnerable to unregulated diving tourism (e.g. Anderson et al. 2011) and as non-target by-catch in coastal fisheries, including shark safety net programs (Paterson 1990; Krogh and Reid 1996).
Site affinity
We show that a proportion of the manta ray population on the east coast of Australia displays site affinity to specific locations, being sighted, at Lady Elliot Island in particular, on multiple occasions. We adopt the term ‘site affinity’ for this behaviour, as it is not clear whether manta rays exhibit true site fidelity. It is unknown whether individuals remain at or in the vicinity of the same site over a long period of time, or whether resights occur due to more transient visitation from general movement within a large ‘home range’. Long-term site affinities/fidelities have been documented for M. alfredi populations in Mozambique (Marshall 2008), Hawaii (Clark and Laros 2008) and Japan (Homma et al. 1999) using intensive sight–resight data and in Indonesia using acoustic telemetry (Dewar et al. 2008). For most of these studies, the species was referred to as M. birostris, but is here identified as M. alfredi based on photographs provided by H. Dewar (Indonesia), T. Kashiwagi (Japan) and from the species distribution map in Marshall et al. (2009).
Factors that may explain site affinities that often result in aggregations of M. alfredi are likely to involve feeding areas, cleaning stations, reproductive sites and migratory landmarks, as have been reported for other elasmobranch fishes (Hueter et al. 2004; Heupel et al. 2006; Wilson et al. 2006; Dewar et al. 2008; Bansemer and Bennett 2009). Based on direct observations and video recordings of manta ray behaviour, M. alfredi individuals used all three of the main study sites for feeding and cleaning. These activities appear to be common at most, if not all sites where this species is routinely encountered (Clark and Laros 2008; Dewar et al. 2008; Marshall 2008; McGregor et al. 2008; Deakos 2010; Kitchen-Wheeler 2010), and may be regarded as key factors in relation to site affinity. The issue of whether site affinity is related to reproductive events for M. alfredi is less clear. While philopatry (i.e. site fidelity to breeding and/or natal ground) has been recorded for several elasmobranch species (e.g. Hueter et al. 2004; Heupel et al. 2006; Bansemer and Bennett 2009), there is no specific support in the literature for this behaviour in M. alfredi. Our data did show a positive female sex bias at Lady Elliot Island, but nearly equal proportions of males and females at the other two main study sites. Sex ratios biased in favour of females have also been reported for M. alfredi in Mozambique, where 74.8% of the 507 identified rays were females (Marshall and Bennett 2010b), and in the Maldives where ~63% of the 1379 M. alfredi individuals reported were females (Kitchen-Wheeler 2010). These results suggest that there are differences in the distribution of females and males within the species’ utilised habitat.
There is a possibility that rocky reefs and coral islands act as landmarks that are used by manta rays to orient themselves within their home range, and are also used during seasonal migrations. The observation of manta rays in moderately large numbers at prominent features may be analogous to the behaviour of grey nurse sharks, C. taurus, along the same coastline, in which sharks aggregate at and move between specific sites (Bansemer and Bennett 2010). It is also important to note that while the majority of our manta ray observations occurred at popular SCUBA-diving locations, as they are a major ecotourism drawcard, it does not preclude their occurrence at other non-dived sites.
Movements
This is the first report of regional movements undertaken by M. alfredi using photo-ID techniques in Australian waters. Of the 31 individual manta rays identified at both Lady Elliot Island and North Stradbroke Island, 380 km apart, 32% displayed repeated movements between the two sites within 3 years. In addition, the four individuals sighted at both Lady Elliot Island and at Byron Bay travelled at least 506 km between these sites. The only previous report of movement for this species over similar distances was a 400-km movement made by two individual manta rays in the Yeayama Islands group (Japan), based on a 30-years survey using photo-ID (Kashiwagi et al. 2008). Our results indicate that M. alfredi undertakes regional movements, which probably represent seasonal migrations given the seasonality of sightings at the three sites.
Broad-scale movements are typical amongst large planktivorous elasmobranchs. The whale shark, Rhincodon typus, and the basking shark, Cetorhinus maximus, have similar life histories to the manta rays (i.e. filter-feeders, long-lived and late maturity) and exhibit long-distance migrations. Sims et al. (2003) reported a tagged basking shark that travelled at least 1878 km in 77 days, and a whale shark was shown to travel at least 1501 km from Ningaloo reef to the north-east Indian Ocean in 57 days (Wilson et al. 2006). Migrations and seasonal aggregations of whale and basking sharks have been associated with physical and biological oceanographic conditions, with zooplankton abundance and composition appearing to play a major role in the movement, distribution and aggregations of these large filter-feeders (Heyman et al. 2001; Duffy 2002; Sims et al. 2006; Humphries et al. 2010). Their movements were related to ocean currents and associated thermal fronts that stimulate, aggregate and transport food resources (Taylor and Pearce 1999; Wilson et al. 2001; Cotton et al. 2005; Sleeman 2007).
We show that M. alfredi seasonally occurs at different sites, with animals mainly observed at the two southerly sites during mid-spring to early autumn and at Lady Elliot Island during autumn and winter. Population overlap between the northern and southern aggregation sites was shown by 34 individual rays sighted at two or more of the study sites. These particular individuals were sighted during warmer months at the southern sites and nearly all individuals were sighted only during cooler months at Lady Elliot Island. This distinct pattern of seasonal north–south movements corroborates the anecdotal information obtained from interviews with local collaborators, and is consistent with the relative sighting abundances. These seasonal patterns may be timed to benefit from the temporal and spatial movement of warm waters of the East Australian Current. This current increases in strength during spring and summer, bringing warm waters further south, and decreases in winter (Boland and Church 1981; Ridgway and Dunn 2003). The timing of manta ray movements would minimise the energy required for migration, as manta rays could use the relatively strong East Australian Current flow to migrate southwards during summer and return northward during winter, when the East Australian Current is weakest. The knowledge of manta ray feeding ecology is limited and mainly anecdotal (e.g. Whitley 1936). Although manta rays have been observed feeding at both Lady Elliot Island and North Stradbroke Island, data are currently insufficient to relate their regional movements to their foraging ecology. In addition, seasonal trends in zooplankton productivity at these study areas are still poorly understood. The use of acoustic and satellite tracking technologies in the future will bring new understanding of the movements and habitat use of manta rays. We suggest that temporary and repeated affinity for specific sites appears to be a common feature of M. alfredi behaviour, and that same-site visitation probably relates to seasonal movement patterns linked to changes in sea water temperature, current flow and food abundance. This seasonal migration requires further investigation.
Acknowledgements
We would like to thank our colleagues and dive associates who contributed photographs and without whom this study would not have been possible: M. Atkinson, D. Biddulph, C. Garraway, K. Fiora, M. Gray, C. Roelfsema, M. Ryan, R. Cheseldene-Culley, A. Myers, T. Ito, K. Ito, T. Okohira, K. Sprogis, C. Rohner, J. Natoli, J. Watterson, L. Botterill, K. Coufal, R. Davis, A. Donnelly, J. Chinnick, K. Spence, L. Newey, J. Loder, R. Flynn, J. Flynn, Dobbi, P. Rokic, G. Mac Donald, S. Kucera, J. Sheils, C. Wong, C. Proske, P. Gartrell, S. Marshall, P. Good, M. Chafin, G. Giess, M. Myer, G.Haas, P. Smedley, S. Kirby, M. Garrido, R. Manson, X. Rivett, S. Carter, L. and L. Stewart, A. M. Winkelhagen, M. Canto, A. Gutteridge, C. Josey, Y. Adjei, A. Cox and G. Heritage. We are grateful to the Guest Editors and two anonymous reviewers for their valuable feedback on this paper. We especially thank Earthwatch Institute Australia, Brother Pty Ltd and Lady Elliot Island Eco Resort for their support with field trips to Lady Elliot Island. We thank James Griffith and Manta Lodge & Scuba Centre for their ongoing assistance and field support at North Stradbroke Island. This work is part of the Project Manta supported by the University of Queensland, EarthWatch Australia, Australian Marine Conservation Society, Brother Pty Ltd, Consolidated Rutile Ltd, David Hannan Prod, Eye on the Reef Program, Lady Elliot Island Eco-Resort, Manta Lodge and Scuba Centre, Redland City Council, Project AWARE, PADI foundation, NASA and SEQ Catchment. This study was conducted under Great Barrier Reef Marine Park permit (G09/29853.1), Moreton Bay Marine Park permit (QS2008/CVL1440a) and Ethics approval (SBMS/071/08/SEAWORLD).
References
Anderson, R. C. (2002). Elasmobranchs as a recreational resources. In ‘Elasmobranch Biodiversity, Conservation and Managment. Proceedings of the International Seminar and Workshop, Sabah, Malaysia, July 1997’. (Eds S. L. Fowler, T. M. Reed and F. A. Dipper.) pp. 46–51. (IUCN Species Commission: Sabah.)Anderson, R. C., Shiham, A. M., Kitchen-Wheeler, A. M., and Stevens, G. (2011). Extent and economic value of manta ray watching in Maldives. Tourism in Marine Environments 7, 15–27.
| Extent and economic value of manta ray watching in Maldives.Crossref | GoogleScholarGoogle Scholar |
Australian Government (2009). The direct value of sharks to the marine tourism industry. Available at http://www.rrrc.org.au/publications/downloads/value_of_sharks.pdf [Accessed 15 March 2009].
Bansemer, C. S., and Bennett, M. B. (2008). Multi-year validation of photographic identification of grey nurse sharks, Carcharias taurus, and applications for non-invasive conservation research. Marine and Freshwater Research 59, 322–331.
| Multi-year validation of photographic identification of grey nurse sharks, Carcharias taurus, and applications for non-invasive conservation research.Crossref | GoogleScholarGoogle Scholar |
Bansemer, C. S., and Bennett, M. B. (2009). Reproductive periodicity, localised movements and behavioural segregation of pregnant Carcharias taurus at Wolf Rock, southeast Queensland, Australia. Marine Ecology Progress Series 374, 215–227.
| Reproductive periodicity, localised movements and behavioural segregation of pregnant Carcharias taurus at Wolf Rock, southeast Queensland, Australia.Crossref | GoogleScholarGoogle Scholar |
Bansemer, C. S., and Bennett, M. B. (2010). Retained fishing gear and associated injuries in the east Australian grey nurse sharks (Carcharias taurus): implications for population recovery. Marine and Freshwater Research 61, 97–103.
| Retained fishing gear and associated injuries in the east Australian grey nurse sharks (Carcharias taurus): implications for population recovery.Crossref | GoogleScholarGoogle Scholar |
Boland, F. M., and Church, J. A. (1981). The East Australian Current 1978. Deep-Sea Research 28, 937–957.
| The East Australian Current 1978.Crossref | GoogleScholarGoogle Scholar |
Castro, A. L. F., and Rosa, R. S. (2005). Use of natural marks on population estimates of the nurse shark, Ginglymostoma cirratum, at Atol das Rocas Biological Reserve, Brazil. Environmental Biology of Fishes 72, 213–221.
| Use of natural marks on population estimates of the nurse shark, Ginglymostoma cirratum, at Atol das Rocas Biological Reserve, Brazil.Crossref | GoogleScholarGoogle Scholar |
Cerutti, F., McGregor, F., Stevens, G., Bradshaw, C., and Meekan, M. (2008). Photo identification of manta rays in the Indo-Pacific Ocean. In ‘Joint Meeting of Ichthyologists and Herpetologists. American Elasmobranch Society Devil Ray Symposium, Montreal, 23–28 July 2008’. Abstract 0747. Available at http://www.asih.org/meetingabstracts2008 [Accessed 19 April 2009].
Clark, T., and Laros, K. (2008). Sixteen years of photo-identification of Hawaiian manta rays (Manta birostris). In ‘Joint Meeting of Ichthyologists and Herpetologists. American Elasmobranch Society Devil Ray Symposium, Montreal, 23–28 July 2008’. Abstract 0116. Available at http://www.asih.org/meetingabstracts2008 [Accessed 19 April 2009].
Cotton, P. A., Sims, D. W., Fanshawe, S., and Chadwick, M. (2005). The effect of climate variability on zooplankton and basking shark (Cetorhinus maximus) relative abundance off southwest Britain. Fisheries Oceanography 14, 151–155.
| The effect of climate variability on zooplankton and basking shark (Cetorhinus maximus) relative abundance off southwest Britain.Crossref | GoogleScholarGoogle Scholar |
De Rosemont, M. (2008). Observation and sighting description of the Manta birostris, in BoraBora Island (French Polynesia – South Pacific). In ‘Joint Meeting of Ichthyologists and Herpetologists. American Elasmobranch Society Devil Ray Symposium, Montreal, 23–28 July 2008’. Abstract 0302. Available at http://www.asih.org/meetingabstracts2008 [Accessed 19 April 2009].
Deakos, M. H. (2010). Paired-laser photogrammetry as a simple and accurate system for measuring the body size of free-ranging manta rays Manta alfredi. Aquatic Biology 10, 1–10.
| Paired-laser photogrammetry as a simple and accurate system for measuring the body size of free-ranging manta rays Manta alfredi.Crossref | GoogleScholarGoogle Scholar |
Dewar, H., Mous, P., Domeier, M., Muljadi, A., Pet, J., et al. (2008). Movements and site fidelity of the giant manta ray, Manta birostris, in the Komodo Marine Park, Indonesia. Marine Biology 155, 121–133.
| Movements and site fidelity of the giant manta ray, Manta birostris, in the Komodo Marine Park, Indonesia.Crossref | GoogleScholarGoogle Scholar |
Domeier, M. L., and Nasby-Lucas, N. (2007). Annual re-sightings of photographically identified white sharks (Carcharodon carcharias) at an eastern Pacific aggregation site (Guadalupe Island, Mexico). Marine Biology 150, 977–984.
| Annual re-sightings of photographically identified white sharks (Carcharodon carcharias) at an eastern Pacific aggregation site (Guadalupe Island, Mexico).Crossref | GoogleScholarGoogle Scholar |
Dudgeon, C. L., Noad, M. J., and Lanyon, J. M. (2008). Abundance and demography of a seasonal aggregation of zebra sharks Stegostoma fasciatum. Marine Ecology Progress Series 368, 269–281.
| Abundance and demography of a seasonal aggregation of zebra sharks Stegostoma fasciatum.Crossref | GoogleScholarGoogle Scholar |
Duffy, C. A. J. (2002). Distribution, seasonality, lengths, and feeding behaviour of whale sharks (Rhincodon typus) observed in New Zealand waters. New Zealand Journal of Marine and Freshwater Research 36, 565–570.
| Distribution, seasonality, lengths, and feeding behaviour of whale sharks (Rhincodon typus) observed in New Zealand waters.Crossref | GoogleScholarGoogle Scholar |
Durban, J., Ellifrit, D., Dahlheim, M., Waite, J., Barrett-Lennard, L., Ellis, G., et al. (2010). Photographic mark-recapture analysis of clustered mammal eating killer-whales around Aleutian Islands and Gulf of Alaska. Marine Biology (Berlin) 157, 1591–1604.
| Photographic mark-recapture analysis of clustered mammal eating killer-whales around Aleutian Islands and Gulf of Alaska.Crossref | GoogleScholarGoogle Scholar |
Graham, R., Hickerson, E., Castellanos, D., and Nuttall, M. (2008). Site fidelity and movements of juvenile manta rays in the Gulf of Mexico. In ‘Joint Meeting of Ichthyologists and Herpetologists. American Elasmobranch Society Devil Ray Symposium, Montreal, 23–28 July 2008’. Abstract 0312. Available at http://www.asih.org/meetingabstracts2008 [Accessed 19 April 2009].
Heupel, M. R., Simpfendorfer, C. A., Collins, A. B., and Tyminsky, J. P. (2006). Residency and movement patterns of bonnethead sharks, Sphyrna tiburo, in a large Florida estuary. Environmental Biology of Fishes 76, 47–67.
| Residency and movement patterns of bonnethead sharks, Sphyrna tiburo, in a large Florida estuary.Crossref | GoogleScholarGoogle Scholar |
Heyman, W. D., Graham, R. T., Kjerfve, B., and Johannes, R. E. (2001). Whale sharks Rhincodon typus aggregate to feed on fish spawn in Belize. Marine Ecology Progress Series 215, 275–282.
| Whale sharks Rhincodon typus aggregate to feed on fish spawn in Belize.Crossref | GoogleScholarGoogle Scholar |
Homma, K., Maruyama, T., Itoh, T., Ishihara, H., and Uchida, S. (1999). Biology of the manta ray, Manta birostris Walbaum, in the Indo-Pacific. In ‘Proceedings of the 5th Indo-Pacific Fish Conference, Noumea, 3–8 November 1997’. (Eds B. Seret and J. Sire.) pp. 209–216. (Société Française d’Ichtyologie: Paris.)
Hueter, R. E., Heupel, M. R., Heist, E. J., and Keeney, D. B. (2004). Evidence of philopatry in sharks and implications for the management of shark fisheries. Journal of Northwest Atlantic Fishery Science 35, 239–247.
| Evidence of philopatry in sharks and implications for the management of shark fisheries.Crossref | GoogleScholarGoogle Scholar |
Humphries, N. E., Queiroz, N., Dyer, J. R. M., Pade, N. G., Musyl, M. K., et al. (2010). Environmental context explains Lévy and Brownian movement patterns of marine predators. Nature , .
| Environmental context explains Lévy and Brownian movement patterns of marine predators.Crossref | GoogleScholarGoogle Scholar |
Ishihara, H., and Homma, K. (1995). Manta rays in the Yaeyama Islands. Shark News 5, 3..
Kashiwagi, T., Ito, T., Ovenden, J., and Bennett, M. B. (2008). Population characteristics of Manta birostris observed in Yaeyama, Okinawa, Japan, 1987–2006. In ‘Joint Meeting of Ichthyologists and Herpetologists. American Elasmobranch Society Devil Ray Symposium, Montreal, 23–28 July 2008’. Abstract 0316. Available at http://www.asih.org/meetingabstracts2008 [Accessed 19 April 2009].
Kitchen-Wheeler, A. M. (2010). Visual identification of individual manta ray (Manta alfredi) in the Maldives Islands, Western Indian Ocean. Marine Biology Research 6, 351–363.
| Visual identification of individual manta ray (Manta alfredi) in the Maldives Islands, Western Indian Ocean.Crossref | GoogleScholarGoogle Scholar |
Krogh, M., and Reid, D. (1996). Bycatch in the protective shark meshing programme off south-eastern New South Wales, Australia. Biological Conservation 77, 219–226.
| Bycatch in the protective shark meshing programme off south-eastern New South Wales, Australia.Crossref | GoogleScholarGoogle Scholar |
Last, P. R., and Stevens, J. D. (2009). ‘Sharks and Rays of Australia.’ 2nd edn. (CSIRO Publishing: Melbourne.)
Luiz, O. J., Balboni, A. P., Kodja, G., Andrade, M., and Marum, H. (2009). Seasonal occurrences of Manta birostris (Chondrichthyes: Mobulidae) in southeastern Brazil. Ichthylogcal Research 56, 96–99.
| Seasonal occurrences of Manta birostris (Chondrichthyes: Mobulidae) in southeastern Brazil.Crossref | GoogleScholarGoogle Scholar |
MacCarthy, M., O’Neill, M., and Williams, P. (2006). Customer satisfaction and scuba-diving: some insights from the deep. The Services Industries Journal 26, 537–555.
| Customer satisfaction and scuba-diving: some insights from the deep.Crossref | GoogleScholarGoogle Scholar |
Marshall, A. D. (2008). Biology and population ecology of Manta birostris in southern Mozambique. Ph.D. Thesis, The University of Queensland, Brisbane.
Marshall, A. D., and Bennett, M. B. (2010a). The frequency and effect of shark-inflicted bite injuries to the reef manta ray (Manta alfredi). African Journal of Marine Science 32, 573–580
Marshall, A. D., and Bennett, M. B. (2010). Reproductive ecology of the reef manta ray Manta alfredi in southern Mozambique. Journal of Fish Biology 77, 169–190.
| Reproductive ecology of the reef manta ray Manta alfredi in southern Mozambique.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3cnoslyhsQ%3D%3D&md5=44cd88177c1dc6a22fc236b4f9352964CAS | 20646146PubMed |
Marshall, A. D., Pierce, S. J., and Bennett, M. B. (2008). Morphological measurements of manta rays (Manta birostris) with a description of a foetus from the east coast of Southern Africa. Zootaxa 117, 24–30.
Marshall, A. D., Compagno, L. J. V., and Bennett, M. B. (2009). Redescription of the genus Manta with resurrection of Manta alfredi (Krefft, 1868) (Chondrichthyes; Myliobatoidei; Mobulidae). Zootaxa 2301, 1–28.
McGregor, F., Keulen, M. V., Waite, A., and Meekan, M. (2008). Foraging ecology and population dynamics of the manta ray, Manta birostris in lagoonal waters of Ningaloo Reef, Western Australia. In ‘Joint Meeting of Ichthyologists and Herpetologists. American Elasmobranch Society Devil Ray Symposium, Montreal, 23–28 July 2008’. Abstract 0338. Available at http://www.asih.org/meetingabstracts2008 [Accessed 19 April 2009].
Paterson, R. A. (1990). Effects of long-term anti-shark measures on target and non-target species in Queensland, Australia. Biological Conservation 52, 147–159.
| Effects of long-term anti-shark measures on target and non-target species in Queensland, Australia.Crossref | GoogleScholarGoogle Scholar |
Perkins, J. S., and Whitehead, H. P. (1977). Observation on three species of baleen whales off northern Newfoundland and adjacent waters. Journal of the Fisheries Research Board of Canada 35, 1436–1440.
Ridgway, K. R., and Dunn, J. R. (2003). Mesoscale structure of the mean East Australian Current system and its relationship with topography. Progress in Oceanography 56, 189–222.
| Mesoscale structure of the mean East Australian Current system and its relationship with topography.Crossref | GoogleScholarGoogle Scholar |
Rubin, R. D., and Kumli, K. R. (2008). Photo-identification of the manta ray, Manta birostris, in the Revillagigedos Islands, Mexico. In ‘Joint Meeting of Ichthyologists and Herpetologists. American Elasmobranch Society Devil Ray Symposium, Montreal, 23–28 July 2008’. Abstract 0714. Available at http://www.asih.org/meetingabstracts2008 [Accessed 19 April 2009].
Sims, D. W., Southall, E. J., Richardson, A. J., Reid, P. C., and Metcalfe, J. D. (2003). Seasonal movements and behaviour of basking sharks from archival tagging: no evidence of winter hibernation. Marine Ecology Progress Series 248, 187–196.
| Seasonal movements and behaviour of basking sharks from archival tagging: no evidence of winter hibernation.Crossref | GoogleScholarGoogle Scholar |
Sims, D. W., Witt, M. J., Richardson, A. J., Southall, E. J., and Metcalfe, J. D. (2006). Encounter success of free-ranging marine predator movements across a dynamic prey landscape. Proceedings. Biological Sciences 273, 1195–1201.
| Encounter success of free-ranging marine predator movements across a dynamic prey landscape.Crossref | GoogleScholarGoogle Scholar |
Sleeman, J. C. (2007). Biophysical correlates of relative abundances of marine megafauna at Ningaloo Reef, Western Australia. Marine and Freshwater Research 58, 608–623.
| Biophysical correlates of relative abundances of marine megafauna at Ningaloo Reef, Western Australia.Crossref | GoogleScholarGoogle Scholar |
Speed, C. W., Meekan, M. G., Rowat, D., Pierce, S. J., Marshall, A. D., et al. (2008). Scarring patterns and relative mortality rates of Indian Ocean whale sharks. Journal of Fish Biology 72, 1488–1503.
| Scarring patterns and relative mortality rates of Indian Ocean whale sharks.Crossref | GoogleScholarGoogle Scholar |
Taylor, J. G., and Pearce, A. F. (1999). Ningaloo Reef currents: Implications for coral spawn dispersal, zooplankton and whale shark abundance. Journal of the Royal Society of Western Australia 82, 57–65.
White, W. T., Giles, J., Dharmadi, , and Potter, I. C. (2006). Data on the bycatch fishery and reproductive biology of mobulid rays (Myliobatiformes) in Indonesia. Fisheries Research 82, 65–73.
| Data on the bycatch fishery and reproductive biology of mobulid rays (Myliobatiformes) in Indonesia.Crossref | GoogleScholarGoogle Scholar |
Whitley, G. P. (1932). Studies in ichthyology No. 6. Records of the Australian Museum 18, 321–348 [pls XXXVI–XXXIX]
Whitley, G. P. (1936). The Australian devil ray, Daemomanta alfredi (Krefft), with remarks on the superfamily Mobuloidae (order Batoidei). Australian Zoologist 8, 164–188.
Wilson, S. G., Taylor, J. G., and Pearce, A. F. (2001). The seasonal aggregation of whale sharks at Ningaloo reef, Western Australia: currents, migrations and the El Niño/Southern oscillation. Environmental Biology of Fishes 61, 1–11.
| The seasonal aggregation of whale sharks at Ningaloo reef, Western Australia: currents, migrations and the El Niño/Southern oscillation.Crossref | GoogleScholarGoogle Scholar |
Wilson, S. G., Polovina, J. J., Stewart, B. S., and Meekan, M. G. (2006). Movements of whale sharks (Rhincodon typus) tagged at Ningaloo Reef, Western Australia. Marine Biology 148, 1157–1166.
| Movements of whale sharks (Rhincodon typus) tagged at Ningaloo Reef, Western Australia.Crossref | GoogleScholarGoogle Scholar |
Yano, K., Sato, F., and Takahashi, T. (1999). Observations of mating behavior of the manta ray, Manta birostris, at the Ogasawara Islands, Japan. Ichthylogcal Research 46, 289–296.
| Observations of mating behavior of the manta ray, Manta birostris, at the Ogasawara Islands, Japan.Crossref | GoogleScholarGoogle Scholar |