

Remote sensing shows south-east Queensland koalas (Phascolarctos cinereus) prefer areas of higher tree canopy height within their home ranges
Dave L. Mitchell A B * , Mariela Soto-Berelov B and Simon D. Jones B
A B * , Mariela Soto-Berelov B and Simon D. Jones B
A Australian Koala Foundation, GPO Box 2659, Brisbane, Qld 4001, Australia.
B School of Science, RMIT University, Melbourne, Vic. 3000, Australia.
Wildlife Research 50(11) 939-953 https://doi.org/10.1071/WR22093
Submitted: 1 June 2022 Accepted: 22 December 2022 Published: 27 January 2023
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Home range studies allow investigation of faunal habitat use within a well-defined area, and for some species, the concept of ‘core’ and ‘non-core’ home ranges provides the means to examine how resource use varies within home ranges. Taking this approach, we investigated whether koalas preferentially used areas of taller forest canopy within home ranges. After an extensive examination of data quality and home range estimation methods, we used remote sensing techniques to provide canopy height information at high resolution.
Aims: In many areas, koalas prefer taller individual trees at the plot scale; our aim was to investigate whether koalas prefer forest areas with higher canopy height within their home ranges.
Methods: In our southeast Queensland study area, we developed a canopy height model (CHM) from airborne LiDAR (Light Detection and Ranging) data. Existing radio telemetry and GPS data from 135 koalas were used to generate home ranges using 95% kernel density estimators, and 50% kernels represented core home ranges. Some home ranges occupied more than one forest type (Regional Ecosystem – RE); we treated each RE as an individual patch, and used 225 patches in our analysis. We intersected the 95% kernels with the CHM, and used hierarchical spatial clustering to derive four categorical canopy height classes within each patch. We then compared differences in height class area proportions between core and non-core areas for each patch.
Key results: The highest of the four canopy height classes comprised a significantly higher proportion of core areas (42.3%) than non-core areas (30.7%). Classes 2 and 3 were evenly distributed, and the proportion of Class 4 (lowest canopy height) was 20.3% of non-core areas and 11.0% in core areas. Results were similar for REs grouped by Land Zone and individual REs.
Conclusions and implications: We conclude that areas of higher canopy are an important habitat resource for koalas. We have, for the first time, examined resource variability within entire koala home ranges using remote sensing, and our methods demonstrate an avenue for further research using other forms of remote sensing. Classified canopy height models could also be used for strategic conservation planning, and at population-level koala habitat management when combined with other relevant habitat factors.
Keywords: ecology, habitat preference, habitat use, koala, LiDAR, radio telemetry, spatial clustering, spatial ecology, wildlife management.
Introduction
An animal’s home range is broadly defined as ‘that area traversed by the individual in its normal activities of food gathering, mating, and caring for young’ (Burt 1943). Powell and Mitchell (2012) proposed that a better concept of the home range is ‘that part of an animal’s cognitive map of its environment that it chooses to keep updated’. For animal ecology researchers, understanding the cognitive map requires integration of spatial and temporal animal location data along with the spatial distribution of resource availability and complexity (Heit et al. 2021).
VHF radio telemetry is routinely used to track fauna species, with location recorded at daily to weekly intervals using a hand-held GPS. Location data are then used to define home ranges using one of several methods, e.g. grid cell counts, minimum convex polygon, kernel density estimators, or low convex hull (Gregory 2017). More recently, GPS collars have been used to automatically record location data at much higher temporal frequency, e.g. hourly, but no habitat information is captured. High-frequency locations introduce spatial and temporal autocorrelation (Noonan et al. 2019), and location errors are also more likely with GPS collar data (Fleming et al. 2020). Approaches to reduce the influence of GPS errors on home range estimation include: (1) removing data points with lower precision, defined by the estimated horizontal dilution of precision (HDOP), (Adams et al. 2013); (2) calibration of collars at known locations so that an error term can be used in home range estimates (Ellis et al. 2011; Ironside et al. 2017); and (3) removing fixes with unlikely movement speeds (Wilson-Aggarwal et al. 2021). Many studies do not address GPS error (Meyer et al. 2021), and Ironside et al. (2017) concluded that screening data by HDOP resulted in a major data reduction and had few benefits, whereas Frair et al. (2010) found that home range estimates were robust to GPS error. Some home range packages specifically incorporate GPS error (Fleming et al. 2020). Home range studies may have other shortcomings, including: (1) the small number of individuals tracked; (2) the number of location fixes used to define home ranges; (3) insufficient study length to capture seasonal shifts in home ranges; and (4) indiscriminate use of the term ‘home range’, e.g. 86% of studies lasting less than 3 months referred to estimated areas as home ranges (Goldingay 2015).
Within home ranges ‘core areas’ have been defined as areas of more intensive use (Samuel et al. 1985), but for some species the concept of core home ranges is contentious because it may have little relevance to an animal’s home range use, e.g. northern bettongs (Bettongia tropica) utilise their home ranges evenly (Vernes and Pope 2001), and Bengsen et al. (2012) found that feral cats had no distinct core areas. Other studies have shown that core areas contain critical resources, e.g., an Indonesian study found that core areas contained all Bornean southern gibbon (Hylobates albibarbis) sleeping trees (Cheyne et al. 2019); in the pacific northwest (Washington, USA), female fishers (Pekania pennanti) select core areas dominated by larger trees (Lewis et al. 2016), and a study on spider monkeys (Ateles geoffroyi) found that, because core areas had better habitat quality than non-core areas, they were key to a wider understanding of habitat preferences (Asensio et al. 2012).
Most international papers using kernel density estimators have employed a 50% isopleth to define home range cores (Goldingay 2015), but other studies have used slightly different isopleths; e.g. Stirrat (2003) in their study of agile wallabies (Macropus agilis) used utilisation distribution to define the core area as the 55% isopleth. Using the same methods, Telfer and Griffiths (2006) adopted the 50% isopleth in their short-eared rock wallaby (Petrogale brachyotis) study; and Moseby et al. (2009) adopted a 60% isopleth for their study on feral cats and foxes. For Koala (Phascolarctos cinereus, henceforth koala) studies, Whisson et al. (2020) employed an animal-determined method (Powell 2000) to use 60% and 70% isopleths to define core areas within urban areas; de Oliveira et al. (2014), Goldingay and Dobner (2014), Matthews et al. (2016), Phillips (2016), and Rus et al. (2021) all used 50% isopleths.
The koala was listed as Endangered in New South Wales, Queensland and the Australian Capital Territory in February 2022 under the Environment Protection and Biodiversity Conservation Act 1999 (EPBC Act), with the major reason given as loss of habitat (DAWE 2022). Further research into how koalas select and use habitat is required, which can guide strategic planning and habitat management programs.
Knowledge of habitat use by koalas has so far relied on data collected at high resolution – with specific study aims guiding survey design, sampling intensity and scale. Data is typically collected from: (1) recorded koala locations (Gutteridge Haskins & Davey (GHD) 2009; Hanger et al. 2017); (2) plots (Phillips and Callaghan 2000); (3) transects (Hindell and Lee 1987; Dique et al. 2003); and (4) faecal pellet examination to determine food resources (Ellis et al. 1999). Current habitat management primarily relies on information derived from these data. For example, preferred tree species information obtained at plot scale is used to classify low-resolution, regional-scale mapped vegetation communities according to the proportion of preferred tree species within those communities (Lunney et al. 2000; Callaghan et al. 2011; Department of Environment and Science (DES) 2021a). Occasionally, other data, including landscape configuration and distance to roads, is incorporated into landscape or regional-scale studies (McAlpine et al. 2006; Januchowski et al. 2008; Crowther et al. 2014). Although these approaches are vital, regional-scale habitat maps may not capture habitat variability at more local scales, potentially leading to poor management decisions (Mitchell et al. 2021a).
Studying habitat preferences of individual koalas within their home ranges provides an additional approach to plot or landscape-scale studies. Koalas have high site fidelity (Thompson 2006), and home range sizes are often reported and used, e.g. for comparison within and between regions (Davies et al. 2013). Other data (e.g. tree species, tree height) are routinely collected with koala telemetry (Ellis et al. 2002; Janssen 2012; Davies et al. 2013; Goldingay and Dobner 2014; Matthews et al. 2016; Hanger et al. 2017). These data can also be used to assist with population-level inferences (Aarts et al. 2008) required for broader habitat management. For instance, Rhodes et al. (2005) showed that vegetation associations with a higher proportion of preferred tree species were used more often than other areas. Ellis et al. (2009) suggested that, although tree girth might guide koala foraging patterns, other factors including shelter tree availability and social networks also guide ranging behaviour. Foliar nitrogen availability is a strong driver for tree revisitation, including for isolated trees (Crowther et al. 2022). Crowther et al. (2014) also highlighted the importance of shelter trees for thermoregulation. Barth et al. (2020) found that isolated paddock trees provide ‘stepping stones’ between habitat patches and can be used for forage, particularly during the breeding season. In urban environments Goldingay and Dobner (2014) found that home ranges can require large areas of non-habitat to enable koalas to access patchy food resources. Davies et al. (2013) found that koala core areas in southwest Queensland were restricted to riparian habitats, and Goldingay and Dobner (2014) noted that core areas (50% fixed kernel) varied in proportion to home ranges (95% fixed kernel). Studies examining resource variability within home ranges are particularly rare, and we are aware of only one study (Gallahar et al. 2021) that assessed preferred tree species variability and tree size within home ranges. While providing useful information, few studies suggest how this knowledge can be specifically incorporated into habitat management (Goldingay 2015), exceptions being Crowther et al. (2014), who emphasised that shelter trees should be retained as well as food trees, and Gallahar et al. (2021), who recommended protection of remnant forest on higher-nutrient soils and retention of connecting corridors.
Methods
Koala location data
We acquired existing koala tracking datasets from three studies in south east Queensland covered by airborne LiDAR (Light Detection And Ranging) data: Coomera on the Gold Coast (14 koalas), and Redlands (10), both of which used Sirtrack koala-GPS collars (Ellis et al. 2016); and the Moreton Bay Rail Link (hereafter, MBR) research project, which tracked 471 koalas using a mixture of radiotelemetry locations recorded at least once every 3–4 days, and GPS collars (Hanger et al. 2017). After data examination (see below), we excluded some datasets and used locations from 135 koalas (Fig. 1) for our study.

|
Coomera and Redlands koala tracking data were acquired between 2008 and 2010, contemporaneous with LiDAR acquisition in 2009. For MBR, we used the Historical Imagery function in Google Earth Pro (Google 2021) to discard data for individual koalas where any land clearing had occurred, either after LiDAR acquisition or during koala tracking data acquisition (2013–2017), i.e. we did not use any MBR data obtained with GPS collars that replaced telemetry after 2015.
For the MBR telemetry data, we rejected koalas with fewer than 80 locations (i.e. koalas that had been tracked for fewer than 6 months), and removed outliers which we defined as single locations more than 200 m from other locations. For Coomera and Redlands GPS data, to assess the influence of GPS accuracy on home range size estimation (95% and 50% isopleths), we generated several home ranges (see below) for one koala (Redlands K1009, n locations = 1359) by partitioning location data. Tracking data are commonly reported with HDOP (estimated horizontal dilution of precision, a measure of GPS satellite configuration). Firstly, we removed data with HDOP >10 (Adams et al. 2013), then removed data in two further stages (HDOP ≤ 5, HDOP ≤ 2.5). We repeated the process using only data from a minimum of four satellites, then used one daily (midday) location, and finally, split the dataset into two 4-week periods.
This exercise demonstrated the differences between home range estimators incorporating spatial and temporal autocorrelation (AC) locations, and those estimators treating locations as independent and identically distributed (IID). For AC methods, the difference between isopleths was smaller compared with IID methods and suggests that AC models handle location error better than IID models (Fig. 2). So, because the GPS data, and potentially, the telemetry data, were autocorrelated, we retained all data to minimise information loss. Fig. 2 shows that AC-generated home ranges were approximately 20% larger than IID. Splitting the data into two 4-week periods had the greatest effect on home range size – for the AC 95% isopleth, the home range was twice the size of the second 4-week period, and for the IID 50% isopleth, approximately 60% larger.

|
Home range estimation
To determine home range and core areas, we used ctmm (Continuous-Time Movement Modelling; R Core Team 2021) with R ver. Ri186 4.1.1 (R Core Team 2015), RStudio ver. 1.4.1717 (RStudio Team 2021) and ctmm ver. 0.6.1 (Fleming and Calabrese 2017). As we illustrated above (Fig. 2), ctmm specifically addresses spatial and temporal autocorrelation arising from the use of modern GPS collars which, while recording multiple locations per day, may also include gaps in data collection and short sampling periods (Meyer et al. 2021). ctmm recognises that animal movement is a stochastic process and employs geostatistical methods to: (1) improve kernel estimation; (2) identify candidate kernel models; (3) identify the best model using maximum likelihood fitting with Akaike information criterion (AIC) selection; and (4) generates kernel density estimates at any desired isopleth (Calabrese et al. 2016). The outlie function highlights unusually large animal movements, i.e. locations more likely due to GPS error than actual movement, and we removed several of these from different datasets. We also used a visual diagnostic, the variogram function, to reject approximately 20 koalas with no defined home range (Fig. 3).

|
Canopy height model
Airborne LiDAR uses laser pulses to measure the distance between the aircraft sensor and objects on the ground and, after classification, delineates ground cover, mid-canopy, and upper canopy features. We obtained LiDAR data from ELVIS (Elevation Information System) at https://elevation.fsdf.org.au/. This site collects data from many Australian government agencies, and we downloaded the 2 km × 2 km tiles coincident with the telemetry and GPS datasets. Following the methods of Mitchell et al. (2021b), we derived an upper-canopy height model (CHM) using FUSION software (McGaughey 2020). Although several trees might be within each 10-m cell, we extracted the maximum height within cells to characterise the overall canopy height.
Vegetation map
The ‘Biodiversity status of pre clearing regional ecosystems – Queensland series’ (ver. 12.1; Department of Environment and Science (DES) 2021b) maps preclearing Regional Ecosystem (RE) vegetation communities in south east Queensland at a nominal scale of 1:25 000 (Neldner et al. 2019). Intersection of this dataset with the CHM captures isolated trees, small clumps of trees, and disturbed vegetation not included in the companion Remnant Vegetation dataset (Neldner et al. 2019). Within the 135 home ranges we examined, there were 225 RE patches with 20 distinct REs. RE tree species composition is summarised in Table 1.
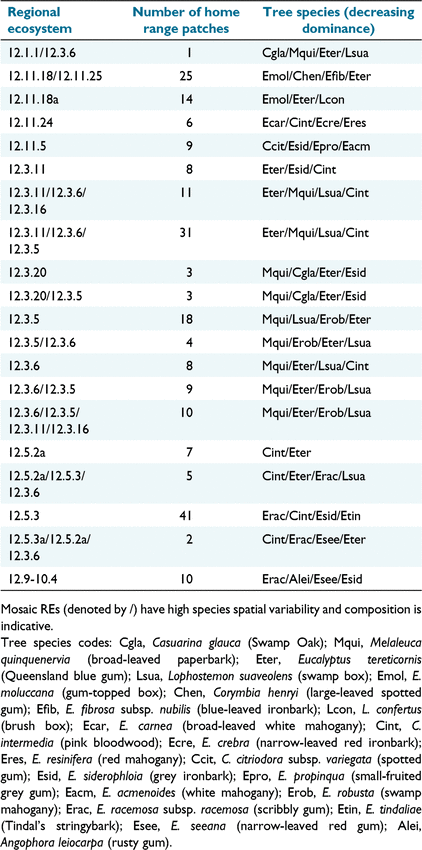
|
Within RE nomenclature, the second digit refers to the Land Zone, e.g. RE 12.3.6 is in Land Zone 3. Land Zones describe geology, landforms and geomorphic processes (Wilson and Taylor 2012), and provide a convenient means to group REs. In our study, these include Land Zone 3 (alluvial river and creek flats), Land Zone 5 (old loamy and sandy plains), Land Zone 11 (hills and lowlands on metamorphic rocks), plus an additional RE (12.9-10.4), which occurs on a combination of Land Zones (fine, and coarse-grained, sedimentary rocks).
Canopy height classification
We clipped the canopy height model to each 95% home range kernel. Canopy height varies among individual REs, so mean heights are not directly comparable among RE patches. To incorporate this variability into our analysis, we treated each patch individually, and following the methods of Mitchell et al. (2021b), we used Geoda (Anselin et al. 2006) with the Redcap agglomerative clustering algorithm (Guo 2008) to classify canopy height within each patch. We generated four height classes, i.e. highest, medium-high, medium-low, and lowest. The lowest class included areas with few or no trees. This approach allowed us to compare height classes between each RE patch rather than actual heights. Fig. 4 illustrates our approach for processing multiple REs within individual home ranges.

|
Analysis
We extracted the Geoda-classified canopy height cells within the core area, with remaining cells constituting non-core classified cells. We tabulated data from 225 patches within 135 home ranges, and calculated the proportion of each class as a proportion of the total number of cells in core and non-core areas (Fig. 5).

|
Paired t-tests were used to examine whether differences between core and non-core proportions were statistically significant for: (1) all patches; (2) patches grouped by Land Zone; and (3) patches within individual REs. Anticipating that some differences would not meet the standard t-test significance criterion (P = 0.05), we also used Cohen’s d, a secondary statistical measure that standardises effect size across datasets with different degrees of freedom (Eqn 1), where values between 0.3 and 0.5 are regarded as having medium effect size (Cohen 1988).
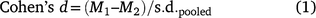
where M1 = core class mean, M2 = non-core class mean,  , s.d. = standard deviation of the mean.
, s.d. = standard deviation of the mean.
The entire data processing workflow is illustrated in Fig. 6.

|
Ethics
Collection of Coomera/Redlands koala location data (Ellis et al. 2016) was approved by the University of Queensland animal ethics committee (CMLR/937/08/ARC/RIOTINTO, ZOO/ENT/115/04/RT) and San Diego Zoo Global IACUC (no. 09-006), and conducted under permits from Queensland Department of Wildlife and Heritage (WITK05609808), WISP05609708, WISP00491303, WISP05609708). Collection of Moreton Bay Rail Link location data (Hanger et al. 2017) was approved by the Sunshine Coast (USC) Animal Ethics Committee (Animal ethics number AN/A/13/80) and by the Queensland Government (Scientific Purposes Permit, WISP11532912).
Results
Home ranges
Mean home range size for our south east Queensland koala dataset was 13.9 ha (n = 135, median = 7.9 ha, range 0.8–113.5 ha). Home ranges on Land Zone 3 (alluvial soils) and Land Zone 5 (plains) appeared to be substantially smaller than those on Land Zone 11 (hillier metamorphic substrates); home range sizes in Land Zone 11 were also highly variable (Table 2). We did not test for significant differences among Land Zones because of unequal sample sizes, unequal variances, non-normal data distribution, and outliers.

|
Mean core home range area was 3.3 ha (n = 135, median 2.0 ha, range 0.19–22.5 ha), an average 23.8% of all home ranges. There were slight differences among Land Zones (range 23.4–26.4%), but none were significant (PANOVA = 0.48).
All home range Regional Ecosystem patches
Class 1 (highest canopy) comprised 33.6% of entire home ranges, followed by Class 2 (27.1%) and Class 3 (21.0%), with Class 4 (lowest canopy height class) last at 18.5% (Fig. 7). Class 1 comprised 42.3% of core areas compared with 30.7% of non-core areas, Classes 2 and 3 were evenly distributed, and Class 4 occupied a greater proportion of non-core areas (20.3%) compared with core areas (11.0%).

|
Differences between core and non-core area proportions were significant for Class 1. Class 2 proportions were not significantly different, and Class 3 proportions were similar but still significantly different. Class 4 (which, as well as areas of lower canopy height, might include disturbed areas) differences were the most significant (Table 3). Variance (not shown) was similar in all classes, and Classes 1 and 4 had largest Cohen’s d.

|
Land Zones
The same trends were evident for RE patches grouped by Land Zones. Patches in Land Zone 11 (with lower-nutrient soils) have the highest proportion of Class 1 in core areas (50.2%) compared with the mean of other Land Zones, which have higher-nutrient soils (40.5%) (Fig. 8).

|
Differences in Class 1 distribution were statistically significant for all Land Zones except 9–10, with a small sample size (d.f. = 9). Cohen’s d indicates a small-to-moderate effect for this Land Zone (Table 4). Generally, there were no significant differences for Classes 2 and 3, and for Class 4 there were significant differences across all Land Zones (Table 4).

|
Individual REs
The same trends (Class 1 proportions higher in core areas, Classes 2 and 3 distributed equally, Class 4 more prominent in non-core areas) were graphically evident across individual REs, and these differences were significant for most REs, e.g. for RE 12.3.11/12.3.6/12.3.5 (Fig. 9, Table 5; see Supplementary material for all REs).

|

|
The only exception in these trends was for RE 12.3.5, where there were no apparent (Fig. 10) or significant (Table 6) differences in class distribution between core and non-core areas.

|

|
Discussion
Our aim was to investigate whether koalas prefer forest areas with higher canopy height within their home ranges. We took a straightforward approach by firstly using spatial clustering to generate contiguous height classes from LiDAR within individual home ranges, and then examined differences in height class allocation between core and non-core home range areas.
Results
Within our dataset of 225 Regional Ecosystem (REs – mapped forest communities) patches within 135 home ranges, we found that areas of highest canopy height (Class 1) comprised a significantly higher proportion of core areas (42.3%) compared with non-core areas (30.7%). For Classes 2 and 3 there was no significant difference between the two areas, but the lowest height class (Class 4) occupied 11.0% of core areas and 20.3% of non-core areas. Our results were consistent across both REs grouped by Land Zones (Wilson and Taylor 2012) and individual REs.
Interpretation of results
Our study examines use versus availability (Johnson 1980) of a potential resource within estimated koala home ranges. We achieved this by comparing forest canopy height distribution within non-core and core home ranges, where koalas can be expected to be found 50% of the time. Core areas, occupying 25% of the total home range, have a higher proportion of the highest-canopy class compared with non-core areas. We conclude that areas of higher canopy height are therefore likely to be an important habitat resource for koalas. Our results concur with previous findings that show koalas generally prefer larger or taller trees (Hindell and Lee 1987; Phillips and Callaghan 2000; Moore and Foley 2005; Matthews et al. 2007; Ellis et al. 2009; Smith et al. 2013; Office of Environment and Heritage (OEH) 2018).
Our results were similarly consistent across Land Zones, but we note that for Land Zone 11, the proportion of Class 1 (highest canopy) in core areas was 24% higher than the average of remaining Land Zones. This Land Zone has lower-fertility soils, therefore we suggest that these localised areas of higher canopy are more likely to be on soils with slightly higher nutrient availability, an important factor in habitat use (Januchowski et al. 2008; Law et al. 2017). Another explanation might be that core areas have a higher density of preferred food trees, e.g. E. tereticornis on alluvial soils (White 1999; Phillips et al. 2000; Office of Environment and Heritage (OEH) 2018). This species is generally taller than co-occurring tree species such as Corymbia spp. (Mitchell et al. 2021b) and in undisturbed areas, any patches with higher density of E. tereticornis might be captured by the highest canopy height class. We also note that Class 4 (lowest canopy height class) showed the greatest difference in use between core and non-core areas. Why koalas use non-core areas less is a complex issue – in some home ranges it might be because of habitat disturbance, in others because of lower nutrient status, local variations in tree species composition, or social factors. Nevertheless, these areas form a necessary component of home ranges. For individual REs, core area Class 1 proportions were also greater than non-core proportions. The only exception was RE 12.3.5, where we did not distinguish any significant difference between core and non-core areas for any height class. RE 12.3.5 is composed of Melaleuca quinquenervia open forest with Eucalyptus robusta and E. tereticornis as scattered individuals or clumps (Department of Environment and Science (DES) 2021c). These species are generally higher than the M. quinquenervia canopy (D.M., pers. obs), but do not have the density required to generate a distinct height class within home ranges.
By using spatial clustering instead of a continuous canopy height model, we have avoided two potential complications. Adjoining cells are more likely to belong to the same class, thereby reducing the influence of under-canopy GPS errors, which can be in the order of 30 m (Frair et al. 2010). Secondly, all spatial layers we used were classified (core, non-core, and four height classes), enabling us to use simple paired t-tests for our analysis.
Study limitations
Our research relied on existing koala tracking data, so we could not design a study capturing a more representative sample of koala habitats in southeast Queensland, or other regions with koalas, nor could we observe fine-scale aspects of forest structure and tree use by tracked koalas. We only examined the potential influence of canopy height class stratified by low-resolution pre-clearing Regional Ecosystems (REs) maps with a nominal 1:25 000 accuracy (Neldner et al. 2019); for smaller home ranges encompassing more than one RE, this limitation could introduce error into our analysis. We recognise that many other variables might influence core home range establishment, e.g. the proportion of preferred tree species within REs (McAlpine et al. 2006) or the availability of shelter trees (Crowther et al. 2014). Other evidence suggests that social interaction influences koala movement (Ellis et al. 2009; Jiang et al. 2022) and therefore core area extent, but we did not attempt to account for this variable because so many core home ranges overlapped. Core extents are likely influenced by both seasonal use of shelter trees (Ellis et al. 2009) and use of different tree species during breeding and non-breeding seasons (Martin 1985; Goldingay 2015), but few koalas were tracked for 12 months or longer so we could not capture seasonal home range shifts. For Coomera and Redlands, telemetry data was contemporaneous with LiDAR, but for the Moreton Bay Rail data this was not possible. We addressed this issue by using Google Earth historical imagery to reject data with habitat disturbance between LiDAR and telemetry acquisition.
Relation to other studies
Where koalas are present, the major factor in habitat occupancy is the density of preferred food and shelter tree species (McAlpine et al. 2006; Ellis et al. 2009; Davies et al. 2013), but low-resolution maps depicting these resources do not capture spatial variability at finer scales where koala habitat management decisions are made (Mitchell et al. 2021a). To our knowledge only one study has mapped all trees, and tree species, within an area occupied by koalas (Moore et al. 2010), which would be the ideal basis for habitat management, but across large areas this approach is impractical. Our study exploits another known koala habitat preference, the use of taller trees, and this information is available over large areas within the koala’s geographic range using LiDAR.
Higher soil fertility (Phillips et al. 2000; Callaghan et al. 2011), gentler slopes and lower elevation (Gutteridge Haskins & Davey (GHD) 2009), and landscape position (Callaghan et al. 2011) are other abiotic factors influencing koala habitat quality; one study in Redlands links all these (Biolink 2019). To a large degree these factors also determine the growth rate and size of trees (Bell and Williams 1997), as well as local variation in tree species diversity and forest structure (Neldner et al. 2019), which can confound the utility of current broad-scale habitat maps at the higher spatial resolution required for habitat management (Mitchell et al. 2021a). We surmise that areas of higher canopy, which can be mapped at high resolution, encapsulate some of these unmapped and highly variable abiotic factors that contribute to habitat quality, and hence habitat use by koalas.
Our calculated home ranges may be larger than comparable home ranges shown in the Moreton Bay Rail Link (MBR) study (Hanger et al. 2017) or other studies, for three reasons. Firstly, we used telemetry datasets with a minimum of 80 locations (i.e. a minimum of 6 months) compared with a minimum of 20 locations used in the MBR study (Hanger et al. 2017). Short-term studies may only capture part of a larger home range (Goldingay 2015); in our exploratory analyses, we detected temporary shifts (not shown) within larger home ranges for some koalas over a period of a few weeks using variogram plots within ctmm. Secondly, using GPS locations for one koala and all suitable kernel density estimators, we confirmed that estimators incorporating autocorrelation (AC) increase calculated home range size by approximately 20% compared with estimators assuming location independence (IID), similar to results obtained by Fleming et al. (2015). Lastly, many telemetry studies assume that, when data is collected daily or weekly, or by thinning GPS data (Matthews et al. 2016), then temporal autocorrelation need not be considered. This may be the case, but spatial autocorrelation may still occur. For example, we used daily-to-weekly telemetry data from 114 koalas tracked during the MBR study, and AC estimators for 45 koalas (39%) were ranked higher than IID estimators. Therefore, reported home range estimates relying on IID likely underestimate home range size.
A study in Lismore (New South Wales) by Goldingay and Dobner (2014) found that core areas were proportionate to home range areas, occupying an average 4.7% of home ranges. These home ranges included cleared and developed land, i.e. areas with no food or shelter resources. In our study, home ranges occupied relatively undisturbed habitats, albeit within peri-urban landscapes, and core areas occupied an average of 23.8% of all home ranges. This contrast highlights that consideration of core areas may be a useful addition to future koala ecology studies.
Future work
Our study has shown that higher canopy height is a koala habitat resource, but how important is it? We looked at canopy height in isolation, in a future study we will insert this resource into landscape-scale multivariate mixed-effect models (McAlpine et al. 2006; Januchowski et al. 2008; Crowther et al. 2014) to establish the relative importance of canopy height compared with known predictor variables.
Previous remote sensing research at home range scales has concentrated on image classification (for koalas, see Ream 2013; Wu et al. 2019), which does not produce contiguous map classes to present readily interpretable research findings, nor encourage subsequent use in habitat management maps. For the first time, we have used spatial clustering techniques with remotely sensed data to identify and quantify the variability of a resource within home ranges. Our methods demonstrate the utility of this approach and provide a template for future research. Ellis et al. (2009) established the importance of lower-height shelter trees to koalas, and Eyre (2006) identified forest structure as a resource for greater gliders (Petauroides volans); both resources could be mapped using the classified standard deviation of LiDAR-derived canopy heights. Crowther et al. (2014) identified sheltered gullies, taller trees, lower elevations, and canopy shelter as important koala habitat variables – these can all be mapped at any desired resolution using LiDAR. The clustering algorithm detected no differences for RE 12.3.5 (Melaleuca quinquenervia open forest with Eucalyptus robusta and E. tereticornis), a vegetation community important to koalas along the New South Wales north coast (Phillips et al. 2000), as well as in south east Queensland. These species are generally higher than the M. quinquenervia canopy (D.M., pers. obs), and can be considered as emergent trees requiring a different approach, e.g. using LiDAR and non-spatial clustering to identify areas with higher trees. Ultimately, combined remotely sensed data types could be integrated with existing habitat maps to form a resource model (Di Stefano et al. 2011) and, when extended to population level, provide the quantitative framework required to better inform fauna habitat management.
Conclusion
Core and non-core home ranges provide a convenient means to examine resource availability and resource use within home ranges. We used this approach to confirm that areas of higher canopy are indeed important to koalas. Relatively, core areas have 30% more of the highest canopy class compared with non-core areas. This is a useful addition to our knowledge about how koalas use their home ranges, and has wider implications for habitat management, e.g. Callaghan et al. (2011) suggested a height overlay map added to existing habitat maps would be beneficial for koala management. Our approach, using core and non-core areas, might be useful to examine other factors determining habitat quality.
Supplementary material
Supplementary material is available online.
Data availability
We are not authorised to redistribute koala location data, but our derived data (core and non-core home ranges, canopy height model etc.) can be shared upon reasonable request to the corresponding author. On request, we can provide the R scripts required to facilitate the use of ctmm for home range estimation.
Conflicts of interest
The authors declare no conflicts of interest. Dave L. Mitchell is employed by the Australian Koala Foundation as a Landscape Ecologist.
Declaration of funding
We are grateful to the Australian Koala Foundation (AKF), who funded our research. The funder had no role in study design, data analysis, decision to publish, or preparation of the manuscript.
Personal communication
W. (Bill) Ellis, Research Fellow, School of Agriculture and Food Sciences, Faculty of Science, University of Queensland.
Acknowledgements
We thank Deborah Tabart OAM (AKF Chief Executive Officer) and the AKF Board for their ongoing dedication and support. At the School of Agriculture and Food Sciences, University of Queensland, we particularly thank Bill Ellis and Sean Fitzgibbon for graciously providing their GPS tracking data, and Cassandra Arkinstall for assistance with ctmm. Deirdre de Villiers (Endeavour Veterinary Ecology) kindly facilitated access to the Moreton Bay Rail koala telemetry data, which was provided by the Department of Transport and Main Roads, Queensland Government. We also thank Regan Kelly (RMIT) for processing some of the canopy height data.
References
Aarts, G, MacKenzie, M, McConnell, B, Fedak, M, and Matthiopoulos, J (2008). Estimating space-use and habitat preference from wildlife telemetry data. Ecography 31, 140–160.| Estimating space-use and habitat preference from wildlife telemetry data.Crossref | GoogleScholarGoogle Scholar |
Adams, AL, Dickinson, KJM, Robertson, BC, and van Heezik, Y (2013). An evaluation of the accuracy and performance of lightweight GPS collars in a suburban environment. PLoS ONE 8, e68496.
| An evaluation of the accuracy and performance of lightweight GPS collars in a suburban environment.Crossref | GoogleScholarGoogle Scholar |
Anselin, L, Syabri, I, and Kho, Y (2006). GeoDa: an introduction to spatial data analysis. Geographical Analysis 38, 5–22.
| GeoDa: an introduction to spatial data analysis.Crossref | GoogleScholarGoogle Scholar |
Asensio, N, Lusseau, D, Schaffner, CM, and Aureli, F (2012). Spider monkeys use high-quality core areas in a tropical dry forest. Journal of Zoology 287, 250–258.
| Spider monkeys use high-quality core areas in a tropical dry forest.Crossref | GoogleScholarGoogle Scholar |
Barth, BJ, FitzGibbon, SI, Gillett, A, Wilson, RS, Moffitt, B, Pye, GW, Adam, D, Preece, H, and Ellis, WA (2020). Scattered paddock trees and roadside vegetation can provide important habitat for koalas (Phascolarctos cinereus) in an agricultural landscape. Australian Mammalogy 42, 194–203.
| Scattered paddock trees and roadside vegetation can provide important habitat for koalas (Phascolarctos cinereus) in an agricultural landscape.Crossref | GoogleScholarGoogle Scholar |
Bell DT, Williams JE (1997) Eucalypt ecophysiology. In ‘Eucalypt Ecology’. (Eds J Williams, J Woinarski) pp. 168–196. (Cambridge University Press: Cambridge, UK)
Bengsen, AJ, Butler, JA, and Masters, P (2012). Applying home-range and landscape-use data to design effective feral-cat control programs. Wildlife Research 39, 258–265.
| Applying home-range and landscape-use data to design effective feral-cat control programs.Crossref | GoogleScholarGoogle Scholar |
Biolink (2019) Redlands Coast koala population and habitat assessment. Final report to Redlands City Council April 2019. Biolink Ecological Consultants, Uki, NSW, Australia. Available at https://www.redland.qld.gov.au/download/downloads/id/3297/redlands_coast_koala_population_and_habitat_assessment.pdf. [Accessed 27 March 2022]
Burt, WH (1943). Territoriality and home range concepts as applied to mammals. Journal of Mammalogy 24, 346–352.
| Territoriality and home range concepts as applied to mammals.Crossref | GoogleScholarGoogle Scholar |
Calabrese, JM, Fleming, CH, Gurarie, E, and Freckleton, R (2016). ctmm: an R package for analyzing animal relocation data as a continuous-time stochastic process. Methods in Ecology and Evolution 7, 1124–1132.
| ctmm: an R package for analyzing animal relocation data as a continuous-time stochastic process.Crossref | GoogleScholarGoogle Scholar |
Callaghan, J, McAlpine, C, Mitchell, D, Thompson, J, Bowen, M, Rhodes, J, de Jong, C, Domalewski, R, and Scott, A (2011). Ranking and mapping koala habitat quality for conservation planning on the basis of indirect evidence of tree-species use: a case study of Noosa Shire, south-eastern Queensland. Wildlife Research 38, 89–102.
| Ranking and mapping koala habitat quality for conservation planning on the basis of indirect evidence of tree-species use: a case study of Noosa Shire, south-eastern Queensland.Crossref | GoogleScholarGoogle Scholar |
Cheyne, SM, Capilla, BR, Abdulaziz, K, Supiansyah, , Adul Cahyaningrum, E, and Smith, DE (2019). Home range variation and site fidelity of Bornean southern gibbons [Hylobates albibarbis] from 2010-2018. PLoS ONE 14, e0217784.
| Home range variation and site fidelity of Bornean southern gibbons [Hylobates albibarbis] from 2010-2018.Crossref | GoogleScholarGoogle Scholar |
Cohen J (1988) ‘Statistical Power Analysis for the Behavioral Sciences.’ (Routledge Academic: New York, NY, USA)
Crowther, MS, Lunney, D, Lemon, J, Stalenberg, E, Wheeler, R, Madani, G, Ross, KA, and Ellis, M (2014). Climate-mediated habitat selection in an arboreal folivore. Ecography 37, 336–343.
| Climate-mediated habitat selection in an arboreal folivore.Crossref | GoogleScholarGoogle Scholar |
Crowther, MS, Rus, AI, Mella, VSA, Krockenberger, MB, Lindsay, J, Moore, BD, and McArthur, C (2022). Patch quality and habitat fragmentation shape the foraging patterns of a specialist folivore. Behavioral Ecology 33, 1007–1017.
| Patch quality and habitat fragmentation shape the foraging patterns of a specialist folivore.Crossref | GoogleScholarGoogle Scholar |
Davies, N, Gramotnev, G, Seabrook, L, Bradley, A, Baxter, G, Rhodes, J, Lunney, D, and McAlpine, C (2013). Movement patterns of an arboreal marsupial at the edge of its range: a case study of the koala. Movement Ecology 1, 8.
| Movement patterns of an arboreal marsupial at the edge of its range: a case study of the koala.Crossref | GoogleScholarGoogle Scholar |
DAWE (2022) Koala conservation. Department of Agriculture, Water and the Environment, Canberra, ACT, Australia. Available at https://www.awe.gov.au/environment/biodiversity/bushfire-recovery/funding-support/koala-conservation [Accessed 7 March 2022]
de Oliveira, SM, Murray, PJ, de Villiers, DL, and Baxter, GS (2014). Ecology and movement of urban koalas adjacent to linear infrastructure in coastal south-east Queensland. Australian Mammalogy 36, 45–54.
| Ecology and movement of urban koalas adjacent to linear infrastructure in coastal south-east Queensland.Crossref | GoogleScholarGoogle Scholar |
Department of Environment and Science (DES) (2021a) Spatial modelling for koalas in South East Queensland: report version 2.0. Koala Habitat Areas (KHA) v2.0, Locally Refined Koala Habitat Areas (LRKHA) v2.0, Koala Priority Areas (KPA) v1.0, Koala Habitat Restoration Areas (KHRA) v1.0. Queensland Department of Environment and Science, Brisbane, Qld, Australia. Available at https://environment.des.qld.gov.au/wildlife/koalas/mapping/spp-values.html [Accessed 25 February 2022]
Department of Environment and Science (DES) (2021b) Biodiversity status of pre-clearing regional ecosystems – Queensland series, version 12. Queensland Department of Environment and Science, Brisbane, Qld, Australia. Available at http://qldspatial.information.qld.gov.au/catalogue/custom/detail.page?fid=%7B8FDF54D2-654C-4822-8295-1D8E8E772373%7D [Accessed 5 July 2021]
Department of Environment and Science (DES) (2021c) Regional Ecosystem Description Database (REDD). Version 12. Queensland Department of Environment and Science, Brisbane, Qld, Australia. Available at https://www.qld.gov.au/environment/plants-animals/plants/ecosystems/descriptions/download [Accessed 5 July 2021]
Di Stefano, J, Coulson, G, Greenfield, A, and Swan, M (2011). Resource heterogeneity influences home range area in the swamp wallaby Wallabia bicolor. Ecography 34, 469–479.
| Resource heterogeneity influences home range area in the swamp wallaby Wallabia bicolor.Crossref | GoogleScholarGoogle Scholar |
Dique, DS, de Villiers, DL, and Preece, HJ (2003). Evaluation of line-transect sampling for estimating koala abundance in the Pine Rivers Shire, south-east Queensland. Wildlife Research 30, 127–133.
| Evaluation of line-transect sampling for estimating koala abundance in the Pine Rivers Shire, south-east Queensland.Crossref | GoogleScholarGoogle Scholar |
Ellis, W, Carrick, F, Lundgren, P, Veary, A, and Cohen, B (1999). The use of faecal cuticle examination to determine the dietary composition of koalas. Australian Zoologist 31, 127–133.
| The use of faecal cuticle examination to determine the dietary composition of koalas.Crossref | GoogleScholarGoogle Scholar |
Ellis, WAH, Melzer, A, Carrick, FN, and Hasegawa, M (2002). Tree use, diet and home range of the koala (Phascolarctos cinereus) at Blair Athol, central Queensland. Wildlife Research 29, 303–311.
| Tree use, diet and home range of the koala (Phascolarctos cinereus) at Blair Athol, central Queensland.Crossref | GoogleScholarGoogle Scholar |
Ellis, WAH, Melzer, A, and Bercovitch, FB (2009). Spatiotemporal dynamics of habitat use by koalas: the checkerboard model. Behavioral Ecology and Sociobiology 63, 1181–1188.
| Spatiotemporal dynamics of habitat use by koalas: the checkerboard model.Crossref | GoogleScholarGoogle Scholar |
Ellis, W, Bercovitch, F, FitzGibbon, S, Roe, P, Wimmer, J, Melzer, A, and Wilson, R (2011). Koala bellows and their association with the spatial dynamics of free-ranging koalas. Behavioral Ecology 22, 372–377.
| Koala bellows and their association with the spatial dynamics of free-ranging koalas.Crossref | GoogleScholarGoogle Scholar |
Ellis, WA, FitzGibbon, SI, Barth, BJ, Niehaus, AC, David, GK, Taylor, BD, Matsushige, H, Melzer, A, Bercovitch, FB, Carrick, F, Jones, DN, Dexter, C, Gillett, A, Predavec, M, Lunney, D, and Wilson, RS (2016). Daylight saving time can decrease the frequency of wildlife–vehicle collisions. Biology Letters 12, 20160632.
| Daylight saving time can decrease the frequency of wildlife–vehicle collisions.Crossref | GoogleScholarGoogle Scholar |
Eyre, TJ (2006). Regional habitat selection of large gliding possums at forest stand and landscape scales in southern Queensland, Australia I. Greater glider (Petauroides Volans). Forest Ecology and Management 235, 270–282.
| Regional habitat selection of large gliding possums at forest stand and landscape scales in southern Queensland, Australia I. Greater glider (Petauroides Volans).Crossref | GoogleScholarGoogle Scholar |
Fleming, CH, and Calabrese, JM (2017). A new kernel density estimator for accurate home-range and species-range area estimation. Methods in Ecology and Evolution 8, 571–579.
| A new kernel density estimator for accurate home-range and species-range area estimation.Crossref | GoogleScholarGoogle Scholar |
Fleming, CH, Fagan, WF, Mueller, T, Olson, KA, Leimgruber, P, and Calabrese, JM (2015). Rigorous home range estimation with movement data: a new autocorrelated kernel density estimator. Ecology 96, 1182–1188.
| Rigorous home range estimation with movement data: a new autocorrelated kernel density estimator.Crossref | GoogleScholarGoogle Scholar |
Fleming, CH, Drescher-Lehman, J, Noonan, MJ, Akre, TSB, Brown, DJ, Cochrane, MM, Dejid, N, DeNicola, V, DePerno, CS, Dunlop, JN, Gould, NP, Harrison, A-L, Hollins, J, Ishii, H, Kaneko, Y, Kays, R, Killen, SS, Koeck, B, Lambertucci, SA, LaPoint, SD, Medici, EP, Meyburg, B-U, Miller, TA, Moen, RA, Mueller, T, Pfeiffer, T, Pike, KN, Roulin, A, Safi, K, Séchaud, R, Scharf, AK, Shephard, JM, Stabach, JA, Stein, K, Tonra, CM, Yamazaki, K, Fagan, WF, and Calabrese, JM (2020). A comprehensive framework for handling location error in animal tracking data. bioRxiv , 2020.06.12.130195.
| A comprehensive framework for handling location error in animal tracking data.Crossref | GoogleScholarGoogle Scholar |
Frair, JL, Fieberg, J, Hebblewhite, M, Cagnacci, F, DeCesare, NJ, and Pedrotti, L (2010). Resolving issues of imprecise and habitat-biased locations in ecological analyses using GPS telemetry data. Philosophical Transactions of the Royal Society B: Biological Sciences 365, 2187–2200.
| Resolving issues of imprecise and habitat-biased locations in ecological analyses using GPS telemetry data.Crossref | GoogleScholarGoogle Scholar |
Gallahar, N, Leigh, K, and Phalen, D (2021). Koala tree selection in a mixed-tenure landscape and post-fire implications. Wildlife Research 48, 737–755.
| Koala tree selection in a mixed-tenure landscape and post-fire implications.Crossref | GoogleScholarGoogle Scholar |
Goldingay, RL (2015). A review of home-range studies on Australian terrestrial vertebrates: adequacy of studies, testing of hypotheses, and relevance to conservation and international studies. Australian Journal of Zoology 63, 136–146.
| A review of home-range studies on Australian terrestrial vertebrates: adequacy of studies, testing of hypotheses, and relevance to conservation and international studies.Crossref | GoogleScholarGoogle Scholar |
Goldingay, RL, and Dobner, B (2014). Home range areas of koalas in an urban area of north-east New South Wales. Australian Mammalogy 36, 74–80.
| Home range areas of koalas in an urban area of north-east New South Wales.Crossref | GoogleScholarGoogle Scholar |
Google (2021) Google Earth Pro 7.3.4.8248. Available at https://www.google.com/earth/download/gep/agree.html?hl=en-GB [Accessed 26 February 2022]
Gregory T (2017) Chapter 7: Home range estimation. In ‘The International Encyclopedia of Primatology’. (Eds M Bezanson, KC MacKinnon, E Riley, CJ Campbell, K Nekaris, A Estrada, AF Di Fiore, S Ross, LE Jones-Engel, B Thierry, RW Sussman, C Sanz, J Loudon, S Elton, A Fuentes) pp. 1–4. (John Wiley & Sons, Inc: Hoboken, NJ, USA)
Guo, D (2008). Regionalization with dynamically constrained agglomerative clustering and partitioning (REDCAP). International Journal of Geographical Information Science 22, 801–823.
| Regionalization with dynamically constrained agglomerative clustering and partitioning (REDCAP).Crossref | GoogleScholarGoogle Scholar |
Gutteridge Haskins & Davey (GHD) (2009) South east Queensland koala habitat assessment and mapping project. Prepared by GHD for the Queensland Department of Environment and Resource Management, Brisbane, Qld, Australia.
Hanger J, de Villiers D, Forbes N, Nottidge B, Beyer, H, Loader J, Timms P. (2017) Final technical report, Moreton Bay Rail Koala Management Program. Queensland Department of Transport Main Roads, Brisbane, Qld, Australia. Available at https://www.scribd.com/document/408104224/Moreton-Bay-Koala-report-1-pdf [Accessed 23 August 2022]
Heit, DR, Ortiz-Calo, W, and Montgomery, RA (2021). Landscape complexity persists as a critical source of bias in terrestrial animal home range estimation. Ecology 102, e03427.
| Landscape complexity persists as a critical source of bias in terrestrial animal home range estimation.Crossref | GoogleScholarGoogle Scholar |
Hindell, MA, and Lee, AK (1987). Habitat use and tree preferences of koalas in a mixed eucalypt forest. Australian Wildlife Research 14, 349–360.
| Habitat use and tree preferences of koalas in a mixed eucalypt forest.Crossref | GoogleScholarGoogle Scholar |
Ironside, KE, Mattson, DJ, Arundel, TR, and Hansen, JR (2017). Is GPS telemetry location error screening beneficial? Wildlife Biology 2017, wlb.00229.
| Is GPS telemetry location error screening beneficial?Crossref | GoogleScholarGoogle Scholar |
Janssen, V (2012). Indirect tracking of drop bears using GNSS technology. Australian Geographer 43, 445–452.
| Indirect tracking of drop bears using GNSS technology.Crossref | GoogleScholarGoogle Scholar |
Januchowski, SR, McAlpine, CA, Callaghan, JG, Griffin, CB, Bowen, M, Mitchell, D, and Lunney, D (2008). Identifying multiscale habitat factors influencing koala (Phascolarctos cinereus) occurrence and management in Ballarat, Victoria, Australia. Ecological Management & Restoration 9, 134–142.
| Identifying multiscale habitat factors influencing koala (Phascolarctos cinereus) occurrence and management in Ballarat, Victoria, Australia.Crossref | GoogleScholarGoogle Scholar |
Jiang, AZ, Murray, P, Phillips, C, Tribe, A, and Ellis, W (2022). Movement of free-ranging koalas in response to male vocalisation playbacks. Animals 12, 287.
| Movement of free-ranging koalas in response to male vocalisation playbacks.Crossref | GoogleScholarGoogle Scholar |
Johnson, DH (1980). The comparison of usage and availability measurements for evaluating resource preference. Ecology 61, 65–71.
| The comparison of usage and availability measurements for evaluating resource preference.Crossref | GoogleScholarGoogle Scholar |
Law, B, Caccamo, G, Roe, P, Truskinger, A, Brassil, T, Gonsalves, L, McConville, A, and Stanton, M (2017). Development and field validation of a regional, management-scale habitat model: a koala Phascolarctos cinereus case study. Ecology and Evolution 7, 7475–7489.
| Development and field validation of a regional, management-scale habitat model: a koala Phascolarctos cinereus case study.Crossref | GoogleScholarGoogle Scholar |
Lewis, JC, Jenkins, KJ, Happe, PJ, Manson, DJ, and McCalmon, M (2016). Landscape-scale habitat selection by fishers translocated to the Olympic Peninsula of Washington. Forest Ecology and Management 369, 170–183.
| Landscape-scale habitat selection by fishers translocated to the Olympic Peninsula of Washington.Crossref | GoogleScholarGoogle Scholar |
Lunney, D, Matthews, A, Moon, C, and Ferrier, S (2000). Incorporating habitat mapping into practical koala conservation on private lands. Conservation Biology 14, 669–680.
| Incorporating habitat mapping into practical koala conservation on private lands.Crossref | GoogleScholarGoogle Scholar |
Martin, RW (1985). Overbrowsing, and decline of a population of the koala, Phascolarctos cinereus, in Victoria. I. Food preference and food tree defoliation. Wildlife Research 12, 355–365.
| Overbrowsing, and decline of a population of the koala, Phascolarctos cinereus, in Victoria. I. Food preference and food tree defoliation.Crossref | GoogleScholarGoogle Scholar |
Matthews, A, Lunney, D, Gresser, S, and Maitz, W (2007). Tree use by koalas (Phascolarctos cinereus) after fire in remnant coastal forest. Wildlife Research 34, 84–93.
| Tree use by koalas (Phascolarctos cinereus) after fire in remnant coastal forest.Crossref | GoogleScholarGoogle Scholar |
Matthews, A, Lunney, D, Gresser, S, and Maitz, W (2016). Movement patterns of koalas in remnant forest after fire. Australian Mammalogy 38, 91–104.
| Movement patterns of koalas in remnant forest after fire.Crossref | GoogleScholarGoogle Scholar |
McAlpine, CA, Rhodes, JR, Callaghan, JG, Bowen, ME, Lunney, D, Mitchell, DL, Pullar, DV, and Possingham, HP (2006). The importance of forest area and configuration relative to local habitat factors for conserving forest mammals: a case study of koalas in Queensland, Australia. Biological Conservation 132, 153–165.
| The importance of forest area and configuration relative to local habitat factors for conserving forest mammals: a case study of koalas in Queensland, Australia.Crossref | GoogleScholarGoogle Scholar |
McGaughey RJ (2020) FUSION/LDV: software for LIDAR data analysis and visualization, FUSION version 4.0. United States Department of Agriculture Forest Service, Pacific Northwest Research Station, Portland, OR, USA. Available at http://forsys.cfr.washington.edu/FUSION/fusion_overview.html [Accessed 30 March 2022]
Meyer, NFV, King, J-P, Mahony, M, Clulow, J, Beranek, C, Reedman, C, Balkenhol, N, and Hayward, MW (2021). Large area used by squirrel gliders in an urban area, uncovered using GPS telemetry. Ecology and Evolution 11, 7147–7153.
| Large area used by squirrel gliders in an urban area, uncovered using GPS telemetry.Crossref | GoogleScholarGoogle Scholar |
Mitchell, DL, Soto-Berelov, M, Langford, WT, and Jones, SD (2021a). Factors confounding koala habitat mapping at multiple decision-making scales. Ecological Management & Restoration 22, 171–182.
| Factors confounding koala habitat mapping at multiple decision-making scales.Crossref | GoogleScholarGoogle Scholar |
Mitchell, DL, Soto-Berelov, M, and Jones, SD (2021b). Regional variation in forest canopy height and implications for koala (Phascolarctos cinereus) habitat mapping and forest management. Forests 12, 1494.
| Regional variation in forest canopy height and implications for koala (Phascolarctos cinereus) habitat mapping and forest management.Crossref | GoogleScholarGoogle Scholar |
Moore, BD, and Foley, WJ (2005). Tree use by koalas in a chemically complex landscape. Nature 435, 488–490.
| Tree use by koalas in a chemically complex landscape.Crossref | GoogleScholarGoogle Scholar |
Moore, BD, Lawler, IR, Wallis, IR, Beale, CM, and Foley, WJ (2010). Palatability mapping: a koala’s eye view of spatial variation in habitat quality. Ecology 91, 3165–3176.
| Palatability mapping: a koala’s eye view of spatial variation in habitat quality.Crossref | GoogleScholarGoogle Scholar |
Moseby, KE, Stott, J, and Crisp, H (2009). Movement patterns of feral predators in an arid environment – implications for control through poison baiting. Wildlife Research 36, 422–435.
| Movement patterns of feral predators in an arid environment – implications for control through poison baiting.Crossref | GoogleScholarGoogle Scholar |
Neldner VJ, Wilson BA, Dillewaard HA, Ryan TS, Butler DW, McDonald WJF, Addicott EP, Appelman CN (2019) Methodology for survey and mapping of regional ecosystems and vegetation communities in Queensland. Version 5.0. Updated March 2019. Queensland Herbarium, Queensland Department of Environment and Science, Brisbane. Available at https://www.publications.qld.gov.au/dataset/d8244c14-d879-4a11-878c-2b6d4f01a932/resource/6dee78ab-c12c-4692-9842-b7257c2511e4/download/methodology-mapping-surveying-v6.pdf
Noonan, MJ, Tucker, MA, Fleming, CH, Akre, TS, Alberts, SC, Ali, AH, Altmann, J, Antunes, PC, Belant, JL, Beyer, D, Blaum, N, Böhning-Gaese, K, Cullen, L, Cunha, R, de Paula, RC, Dekker, J, Drescher-Lehman, J, Farwig, N, Fichtel, C, Fischer, C, Ford, AT, Goheen, JR, Janssen, R, Jeltsch, F, Kauffman, M, Kappeler, PM, Koch, F, LaPoint, S, Markham, AC, Medici, EP, Morato, RG, Nathan, R, Luiz, GR, Oliveira-Santos, LGR, Olson, KA, Patterson, BD, Paviolo, A, Ramalho, EE, Rösner, S, Schabo, DG, Selva, N, Sergiel, A, Xavier, M, da Silva, MX, Spiegel, O, Thompson, P, Ullmann, W, Zięba, F, Zwijacz-Kozica, T, Fagan, WF, Mueller, T, and Calabrese, JM (2019). A comprehensive analysis of autocorrelation and bias in home range estimation. Ecological Monographs 89, e01344.
| A comprehensive analysis of autocorrelation and bias in home range estimation.Crossref | GoogleScholarGoogle Scholar |
Office of Environment and Heritage (OEH) (2018) A review of koala tree use across New South Wales. Office of Environment and Heritage, Sydney, NSW. Available at https://www.environment.nsw.gov.au/research-and-publications/publications-search/a-review-of-koala-tree-use-across-new-south-wales. [Accessed 25 March 2022]
Phillips, S (2016). Aversive behaviour by koalas (Phascolarctos cinereus) during the course of a music festival in northern New South Wales, Australia. Australian Mammalogy 38, 158–163.
| Aversive behaviour by koalas (Phascolarctos cinereus) during the course of a music festival in northern New South Wales, Australia.Crossref | GoogleScholarGoogle Scholar |
Phillips, S, and Callaghan, J (2000). Tree species preferences of koalas (Phascolarctos cinereus) in the Campbelltown area south-west of Sydney, New South Wales. Wildlife Research 27, 509–516.
| Tree species preferences of koalas (Phascolarctos cinereus) in the Campbelltown area south-west of Sydney, New South Wales.Crossref | GoogleScholarGoogle Scholar |
Phillips, S, Callaghan, J, and Thompson, V (2000). The tree species preferences of koalas (Phascolarctos cinereus) inhabiting forest and woodland communities on Quaternary deposits in the Port Stephens area, New South Wales. Wildlife Research 27, 1–10.
| The tree species preferences of koalas (Phascolarctos cinereus) inhabiting forest and woodland communities on Quaternary deposits in the Port Stephens area, New South Wales.Crossref | GoogleScholarGoogle Scholar |
Powell, RA, and Mitchell, MS (2012). What is a home range? Journal of Mammalogy 93, 948–958.
| What is a home range?Crossref | GoogleScholarGoogle Scholar |
Powell R (2000) Animal home ranges and territories and home range estimators. In ‘Research techniques in animal ecology: controversies and consequences’. (Eds L Boitani, TK Fuller) pp. 65–110. (Columbia University Press: New York, NY, USA)
R Core Team (2015) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at https://www.R-project.org/
R Core Team (2021) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at https://www.R-project.org/
Ream B (2013) Mapping Eucalypts in South-West Queensland: answering the question can fine resolution satellite remote sensing be used to map Eucalypt composition. MPhil thesis, School of Geography, Planning & Environmental Management, The University of Queensland, Brisbane, Qld, Australia.
Rhodes, JR, McAlpine, CA, Lunney, D, and Possingham, HP (2005). A spatially explicit habitat selection model incorporating home range behavior. Ecology 86, 1199–1205.
| A spatially explicit habitat selection model incorporating home range behavior.Crossref | GoogleScholarGoogle Scholar |
RStudio Team (2021) RStudio: Integrated Development for R. RStudio, PBC, Boston, MA. Available at http://www.rstudio.com/
Rus, AI, McArthur, C, Mella, VSA, and Crowther, MS (2021). Habitat fragmentation affects movement and space use of a specialist folivore, the koala. Animal Conservation 24, 26–37.
| Habitat fragmentation affects movement and space use of a specialist folivore, the koala.Crossref | GoogleScholarGoogle Scholar |
Samuel, MD, Pierce, DJ, and Garton, EO (1985). Identifying areas of concentrated use within the home range. The Journal of Animal Ecology 54, 711–719.
| Identifying areas of concentrated use within the home range.Crossref | GoogleScholarGoogle Scholar |
Smith, AG, McAlpine, C, Rhodes, JR, Seabrook, L, Baxter, G, Lunney, D, and Bradley, A (2013). At what spatial scales does resource selection vary? A case study of koalas in a semi-arid region. Austral Ecology 38, 230–240.
| At what spatial scales does resource selection vary? A case study of koalas in a semi-arid region.Crossref | GoogleScholarGoogle Scholar |
Stirrat, SC (2003). Seasonal changes in home-range area and habitat use by the agile wallaby (Macropus agilis). Wildlife Research 30, 593–600.
| Seasonal changes in home-range area and habitat use by the agile wallaby (Macropus agilis).Crossref | GoogleScholarGoogle Scholar |
Telfer, WR, and Griffiths, AD (2006). Dry-season use of space, habitats and shelters by the short-eared rock-wallaby (Petrogale brachyotis) in the monsoon tropics. Wildlife Research 33, 207–214.
| Dry-season use of space, habitats and shelters by the short-eared rock-wallaby (Petrogale brachyotis) in the monsoon tropics.Crossref | GoogleScholarGoogle Scholar |
Thompson J (2006) The comparative ecology and population dynamics of koalas in the Koala Coast region of south-east Queensland. PhD thesis, School of Integrative Biology, University of Queensland, Brisbane, Qld, Australia.
Vernes, K, and Pope, LC (2001). Stability of nest range, home range and movement of the northern bettong (Bettongia tropica) following moderate-intensity fire in a tropical woodland, north-eastern Queensland. Wildlife Research 28, 141–150.
| Stability of nest range, home range and movement of the northern bettong (Bettongia tropica) following moderate-intensity fire in a tropical woodland, north-eastern Queensland.Crossref | GoogleScholarGoogle Scholar |
Whisson, DA, Zylinski, S, Ferrari, A, Yokochi, K, and Ashman, KR (2020). Patchy resources and multiple threats: how do koalas navigate an urban landscape? Landscape and Urban Planning 201, 103854.
| Patchy resources and multiple threats: how do koalas navigate an urban landscape?Crossref | GoogleScholarGoogle Scholar |
White, NA (1999). Ecology of the koala (Phascolarctos cinereus) in rural south-east Queensland, Australia. Wildlife Research 26, 731–744.
| Ecology of the koala (Phascolarctos cinereus) in rural south-east Queensland, Australia.Crossref | GoogleScholarGoogle Scholar |
Wilson-Aggarwal, JK, Goodwin, CED, Moundai, T, Sidouin, MK, Swan, GJF, Léchenne, M, and McDonald, RA (2021). Spatial and temporal dynamics of space use by free-ranging domestic dogs Canis familiaris in rural Africa. Ecological Applications 31, e02328.
| Spatial and temporal dynamics of space use by free-ranging domestic dogs Canis familiaris in rural Africa.Crossref | GoogleScholarGoogle Scholar |
Wilson PR, Taylor PM (2012) Land Zones of Queensland. Queensland Herbarium, Queensland Department of Science, Information Technology, Innovation and the Arts, Brisbane, Qld, Australia. Available at https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwi6oJa4o7P2AhWQEYgKHdeXCqcQFnoECAIQAQ&url=https%3A%2F%2Fenvironment.des.qld.gov.au%2F__data%2Fassets%2Fpdf_file%2F0038%2F89669%2Fland-zones-queensland.pdf&usg=AOvVaw2nztWej1nZqfK8jwos8YJE [Accessed 26 February 2022]
Wu, H, Levin, N, Seabrook, L, Moore, BD, and McAlpine, C (2019). Mapping foliar nutrition using WorldView-3 and WorldView-2 to assess koala habitat suitability. Remote Sensing 11, 215.
| Mapping foliar nutrition using WorldView-3 and WorldView-2 to assess koala habitat suitability.Crossref | GoogleScholarGoogle Scholar |