

Changes in botanical composition on three farmlets subjected to different pasture and grazing management strategies
L. M. Shakhane A B , J. M. Scott B F , R. Murison C , C. Mulcahy D , G. N. Hinch B , A. Morrow B and D. F. Mackay B EA Faculty of Agriculture, National University of Lesotho, PO Box 180, Roma, Lesotho.
B School of Environmental and Rural Science, University of New England, Armidale, NSW 2351, Australia.
C School of Science and Technology, University of New England, Armidale, NSW 2351, Australia.
D 79 Beardy Street, Armidale, NSW 2350, Australia.
E 3 Jayne Close, Armidale, NSW 2350, Australia.
F Corresponding author. Email: dr.jimscott@gmail.com
Animal Production Science 53(8) 670-684 https://doi.org/10.1071/AN11177
Submitted: 16 August 2011 Accepted: 20 March 2012 Published: 10 July 2013
Journal Compilation © CSIRO Publishing 2013 Open Access CC BY-NC-ND
Abstract
As part of the Cicerone Project’s farmlet experiment, conducted on the Northern Tablelands of New South Wales, Australia, between July 2000 and December 2006, this study assessed the effects of varying soil fertility, pasture species and grazing management on the botanical composition of three 53-ha farmlets subjected to different management strategies. Starting with the same initial conditions, the farmlets were managed to reach different target levels of soil phosphorus (P) and sulfur (S); Farmlet A aimed at 60 mg/kg of Colwell P and 10 mg/kg S (KCl40) whereas Farmlets B and C both aimed at 20 and 6.5 mg/kg of P and S, respectively. Pastures were renovated on six out of eight paddocks on Farmlet A, but only one paddock of each of Farmlets B (typical management) and C (intensive rotational grazing) was renovated. Flexible rotational grazing was employed on Farmlets A and B (each of eight paddocks) while Farmlet C used intensive rotational grazing over its 17 major paddocks, which were further subdivided into 37 subpaddocks. This paper focuses on the botanical composition dynamics observed across all three farmlets and the explanatory variables associated with those changes. Eight assessments of botanical composition were carried out at approximately annual intervals across each of the 37 major paddocks distributed across the farmlets and the results for each of 49 species were aggregated into seven functional groups for analysis. The strongest correlation found was a negative curvilinear relationship between sown perennial grasses (SPG) and warm-season grasses (WSG). The most significant factors affecting the functional group changes were soil P, sowing phase, paddock and date. These factors led to significant increases in SPG and correspondingly lower levels of WSG on Farmlet A compared with Farmlet B. Farmlets B and C experienced similar, declining levels of SPG, and increasing levels of WSG suggesting that intensive rotational grazing did not lead to substantial changes in botanical composition, compared with flexible rotational grazing, in spite of the fact that intensive rotational grazing had much longer grazing rests and shorter graze periods than the other two farmlets. Soil P levels were also significantly associated with levels of cool-season annual grasses, legumes and herbs, especially on Farmlet A. In general, the largest differences in botanical composition were between Farmlet A and the other two farmlets; these differences were most closely associated with those plants categorised as sown, introduced, C3 pasture species. The levels of legume were generally low on all farmlets, due largely to the dry seasons experienced over most of the trial. Efforts to increase the legume composition on all farmlets were more successful on Farmlet A than on the other two farmlets due, presumably, to higher soil fertility on Farmlet A. Farmlet C, with its long rest periods and short graze periods, had a small proportion of legumes, due to the competitive effects of the accumulated tall grass herbage between grazings. The ‘typical’ management of Farmlet B also resulted in low levels of legume as well as increased ‘patchiness’ of the pastures and increased numbers of thistles.
Introduction
The botanical composition of a pasture is critically important for livestock enterprises as it contributes, together with soil fertility and climate, to the productivity of grazing enterprises, by influencing the quality and quantity of herbage available to grazing animals which, in turn, impact on the pasture composition through grazing and nutrient transfer. As changes in pasture botanical composition occur over periods of years, in response to soil fertility and climate, as well as livestock and pasture management, managing change is a complex dynamic issue.
Large changes to the botanical composition of pastures on the Northern Tablelands of New South Wales (NSW) began in the 1950s when early pasture development efforts commenced. These coincided with generally favourable climatic conditions which created relatively little moisture stress on sown pasture species and especially on perennial legumes such as white clover (Blair 1997), which was associated with significant bloat problems at that time (Wolfe and Lazenby 1972).
This early phase of pasture improvement in the temperate higher rainfall zone of NSW resulted in a marked improvement in productivity (Jones et al. 2006) of premium livestock products (Michalk et al. 2003). However, since the mid 1970s, cost and price pressures on grazing enterprises on the Northern Tablelands have led to significant overgrazing and deterioration of both native and sown pastures (Wheeler et al. 1987; Michalk et al. 2003). As a consequence, between 1970 and 1990, stocking rates have declined and are now well below potential (Kemp and Dowling 2000).
A desirable botanical composition for a pasture in the New England region is suggested to be one which supports profitable livestock production while having characteristics associated with sustainability. These have been suggested by Scott et al. (2000) to comprise pastures dominated by deep-rooted, fertiliser-responsive perennial grasses (such as the sown perennial grasses in this study) combined with a persistent perennial legume and few weeds.
During recent drier climatic conditions, there has been a noticeable decline in white clover presence on the Northern Tablelands of NSW and this has had consequences for animal growth and stocking rate. Graham et al. (2003) found that, by increasing legume content from 15 to 40% and soil Olsen phosphorus (P) from 6 to 28 mg/kg at the same levels of rainfall, the carrying capacity could be increased from 14.4 to 22.2 dry sheep equivalents (DSE)/ha.
Enhancing the botanical composition of a pasture can be achieved most quickly by establishing a new pasture, or renovating a degraded, previously sown pasture. However, frequent pasture renovation may not be financially viable due to the high costs of re-establishing pastures (Lees and Reeve 1994), the limited persistence of current cultivars (Waller et al. 2001) and the time frame of 5–8 years to recover costs (Vere et al. 1993). Many producers have reported difficulties in getting sown pastures to persist for 10 years or more (Reeve et al. 2000) and thus are often reluctant to invest in more sown pastures.
Environmental factors, particularly soil moisture and soil fertility are key factors influencing changes in species composition (Garden et al. 2001; Sanford et al. 2003) and this is particularly so in pasture systems where species commonly exhibit differences in growth rate under moisture stress which affects their competitive ability (Harris and Lazenby 1974). Soil fertility also influences botanical composition and in sown perennial grass–legume pastures, the application of phosphorus helps to maintain the more desirable, nutrient-responsive species (Hill et al. 2004) and contribute to higher production, in some cases irrespective of grazing management (Saul et al. 1998).
Hill et al. (2005) found that pasture species varied considerably in the response to soil P levels. In grasslands dominated by native perennial grasses considered to be adapted to low nutrient conditions, fertiliser applications may increase pasture production but not necessarily competitiveness of these grasses as they can be displaced by invasive ‘weedy’ annual species (Garden et al. 2003).
Several researchers have hypothesised that grazing management strategies can be used to manipulate pasture composition and at the same time improve the overall profitability and sustainability of grazing enterprises (Harris and Ayres 1997; Mason and Kay 2000; Mason et al. 2003). Grazing management affects the ability of the animals to be selective in their choice of intake; this selectivity has been linked to changes in pasture composition and appears more pronounced as the herbage on offer increases in age and maturity (Arnold 1960).
A prominent feature of Northern Tablelands native pastures grazed by Merino sheep is the development of sheep camps, which commonly occur at or near the highest point of the paddock or under preferred shade trees (Rogers and Whalley 1989). These sheep camps have characteristic plant groupings and represent the outcome of a complex set of interactions between grazing, trampling and the input of plant nutrients from faeces and urine (Rogers and Whalley 1989).
The aim of the study reported here was to examine the changes in botanical composition that occurred under various pasture and grazing management regimes on three farmlets managed in different ways over 6 years. The trial was part of the Cicerone Project’s whole-farmlet experiment which aimed to examine the profitability and sustainability of three different management systems on the Northern Tablelands region of NSW. While much has been written in the scientific literature about improving botanical composition, often based on small-scale trials, little has been reported on how this can be achieved across multiple paddocks within whole farms under realistic constraints such as the need for extended periods of destocking of paddocks during renovation together with having to support existing livestock on those paddocks not being renovated.
The first hypothesis addressed by this trial was that the sowing of perennial grasses and legumes and the maintenance of higher soil fertility (Farmlet A) would result in improved botanical composition with more deep-rooted, sown perennial grasses and a greater level of persistent legumes compared with the typical district practice based on moderate levels of inputs (Farmlet B). The second hypothesis was that an intensive rotational grazing system with short grazing and long rest periods (Farmlet C) would result in improved botanical composition compared with the typical district practice of flexible rotational grazing with relatively long graze periods and relatively short rest periods (Farmlet B).
Materials and methods
The trial was located on 200 ha of land leased from CSIRO on the Pastoral Research Laboratory ‘Chiswick’ near Uralla, NSW, Australia, and was run from July 2000 to December 2006. Over this period, the site received below median rainfall over most seasons and years, resulting in soil moisture constraints on pasture production over much of the experimental period (Behrendt et al. 2013c; Shakhane et al. 2013a). Full details of the study location, experimental layout, grazing and pasture management strategies, including detailed pasture renovation activities within the farmlets are presented in a related paper (Scott et al. 2013b). Prior to this trial, many of the pastures had been sown and/or fertilised over variable periods however station records confirmed that, at the start of this trial (July 2000), all pastures were at least 10 years old.
Farmlet and pasture treatments
The procedures used to allocate the same areas of equivalent capacity land to each farmlet, including a description of the distribution of soil types, have been described by Scott et al. (2013c). The three farmlet systems (each of 53 ha) were managed according to the treatment guidelines developed by the Cicerone Board for each system (Scott et al. 2013b). In brief, Farmlet A aimed, through pasture renovation, towards a botanical composition comprising 100% of sown species with high target levels of soil P (60 mg/kg Colwell P) and sulfur (10 mg/kg KCl40 S) to be achieved by soil testing and subsequent applications of appropriate fertilisers to each paddock. Farmlets B and C both had moderate target levels of soil P (20 mg/kg Colwell P) and S (6.5 mg/kg KCl40 S) with a low level of pasture renovation, considered typical of the region. More details of the soil fertility aspects of the trial have been provided by (Guppy et al. 2013).
The grazing management of Farmlets A and B both employed flexible rotational grazing with attention paid to pasture mass and quality and livestock condition using PROGRAZE principles (Bell and Allan 2000). In contrast, Farmlet C employed intensive rotational grazing over its 17 major paddocks for the first 15 months and thereafter, following further subdivision, 37 subpaddocks. The target stocking rates for Farmlets A and C were set at 15 DSE/ha whereas that for Farmlet B was 7.5 DSE/ha.
Farmlet A paddocks A1–A6 were renovated in 2000 (A3, A4, A5), 2001 (A6), 2002 (A1, A2), 2003 (A5, A6) and 2004 (A1) whereas paddocks A7 and A8, which had high levels of sown perennial grasses at the start, were not renovated at all over the experimental period (2000–06). The second sowing of paddock A5 in 2003 was due to the inadequate survival of sown species following its initial renovation in June 2000. The need for re-sowing of paddocks A6 and A1 were both due to the failure of the short-term Lolium multiflorum pastures to persist beyond 18 months after being sown in 2001 and 2002, respectively. Paddocks A3, A4 and A5 were initially sown to four perennial grasses (Phalaris aquatica cv. Australian; Festuca arundinacea cv. Quantum; Lolium perenne cv. Lincoln and Dactylis glomerata cv. Porto) and two legumes (Trifolium repens cv. Huia and Trifolium pratense cv. USA) both of which were lime pelleted and treated with molybdenum. Thereafter, most Farmlet A paddocks were renovated with simple pasture mixtures of persistent, perennial grasses and a legume (as for paddocks B6 and C17 below). The exception to this was paddock A1 which, in an effort to increase the percentage of a persistent perennial legume on this farmlet, was resown in 2004 to what was considered a novel pasture mix for the area comprising Medicago sativa cv. Aurora, Phalaris aquatica, cv. Atlas PG (chosen because of its high degree of summer dormancy, to minimise competition over summer with lucerne), Cichorium intybus cv. Puna and Trifolium repens cv. Huia.
In 2004, one paddock of each of Farmlets B and C (B6 and C17) was renovated with a simple pasture mix consisting of persistent, perennial grasses and a perennial legume (Phalaris aquatica cv. Australian; Festuca arundinacea cv. Quantum and Trifolium repens cv. Huia). No other paddocks of these two farmlets were renovated during the trial period.
In an effort to enhance legumes across all three farmlets over the duration of the trial, Trifolium repens (cvv. Huia, Haifa or Nusiral), was lime pelleted and treated with molybdenum before broadcasting whenever fertiliser was applied to any paddock.
Measurements
The botanical composition of all paddocks was assessed approximately annually at a time when pasture species identification was most reliable, when pastures had visible inflorescences (from early to late summer). The assessments were carried out using the BOTANAL procedure for visually estimating the dry weight rank of pasture species (Tothill et al. 1978).
The assessments were made along diagonal transects chosen for each paddock such that any sheep camp areas were avoided; subsequent measurements were taken along the same transects. When sampling, a 0.5 by 0.5-m quadrat was randomly thrown at intervals of ~10–20 m along each transect such that ~20 quadrats were assessed across each paddock at each sampling time. For each quadrat, species were ranked 1st, 2nd, or 3rd according to their estimated contribution to pasture DM. To facilitate calculations, a data entry form in a Microsoft Access database was used to convert the rankings to percentages of each species expressed on a dry weight basis using the constants 70.2, 21.1 and 8.7 for ranks 1, 2, and 3, respectively. Throughout the study, although several operators often worked collaboratively in conducting the botanical composition assessments, one of the authors (CM), with more than 30 years technical experience with the pastures of the CSIRO property ‘Chiswick’, participated in all measurements, thus ensuring consistency in the methodology used.
The initial measurement was conducted in March 2000 (early autumn) to provide a baseline measurement of the state of pastures before the farmlet treatments commenced on 1 July 2000. During the first 30 months of the experiment, assessments were conducted in early summer (December 2000, 2001 and 2002) after flowering of most species had commenced. However, for subsequent assessments of botanical composition, the sampling time was modified to late summer (February 2003, 2004, 2005 and January, 2006) as it was found that greater definition of the majority of species, the warm-season native grasses, was possible at this time.
The individual pasture species were categorised into seven functional groups determined by their seasonal growth patterns, perenniality and plant type: sown perennial grasses (SPG), warm-season grasses (WSG), year-long green grasses (YGG), cool-season perennial grasses (CPG), cool-season annual grasses (CAG), herbs and legumes in a manner similar to the classification used by Whalley et al. (1976). Although categories such as response to fertiliser and grazing, forage value and drought resistance used by Lodge and Whalley (1989) were considered, they were not used for this study as it was difficult to find unambiguous literature which would have supported such categorisation of all 49 observed species.
Statistical analyses
The trends in botanical composition of all functional groups were compared between farmlets by fitting curves with local regression, likelihood and density estimation using the locfit package of R (R Development Core Team 2009), which shows individual data points for each paddock as well as mean values and confidence limits.
The significance of differences between functional groups as affected by farmlet treatment and time were determined using General Estimating Equations (GEE) using the statistics package R (R Development Core Team 2009). In these analyses, the BOTANAL data were considered as repeated-measurements from field plots of botanical compositions, which were represented as percentages of the total vegetation in the sampling unit (the paddock).
The statistical model used had systematic components of farmlet, time and the interaction between farmlet and time. There were two sources of correlation among the residuals: those due to the multivariate nature of the response and those due to repeated-measures from the same plot. The distribution of the errors included terms to account for these correlations.
The statistical analyses used included the use of pairplots to estimate the Pearson correlation coefficients between response and explanatory variables, generalised linear modelling (GLM) and redundancy analysis (RDA) (Zuur et al. 2007) using the software program ‘Brodgar’ (version 2.6.6, Highland Statistics Ltd, Newburgh, UK), which provides a graphical user interface to the statistical package R (R Development Core Team 2009).
In order to extend the GLM to accommodate significantly correlated species groups, the data were modelled using a GEE (Liang and Zeger 1986). The working correlation among residuals was estimated by a 2-step process. At each sampling, the multivariate set of proportions (out of 100) were analysed using a GEE with unstructured correlation. Statistical inferences between the treatments were based on log-odds ratios with corresponding confidence intervals for either Farmlet A or C compared with the control, Farmlet B.
The multivariate RDA comprised botanical composition data for all seven functional groups as the response variables and the explanatory variables consisting of farmlet, paddock, soil nitrogen (N), P and S levels, pasture phase, pasture age, recent stocking rates, growing degree days and modelled available soil water. RDA were also carried out using all of the categories indicated in Table 1 in order to determine the most important interactions between these categories and farmlet treatments.

|
Results
Over the experimental period, a total of 49 species were recorded across the three farmlets comprising, in decreasing order of numbers of species, 19 WSG, 7 CAG, 6 legumes, 5 CPG, 4 YGG, 4 SPG and 4 herbs (Table 1). To aid in understanding the factors governing the changes of pasture species over time, the species were further categorised using criteria such as the species establishment status (sown/volunteer), origin (native/introduced), seasonality, physiology (C3/C4), perenniality (annual/perennial) and plant type (grass, legume, herb or rush/sedge) (Table 1).
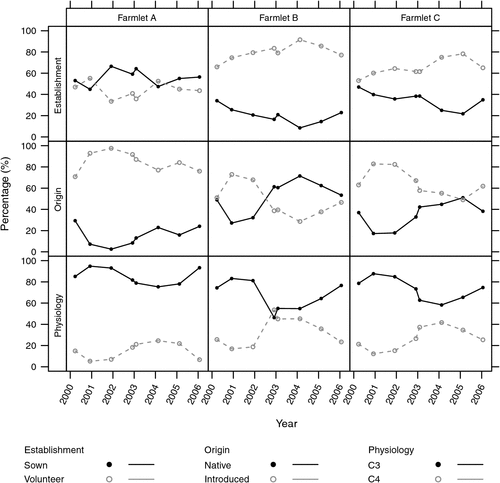
Figure 1 shows an overall summary of the mean composition across all paddocks of each farmlet categorised as seven functional groups over the eight sampling times (mid to late summer). The most obvious feature is that Farmlet A appears to be quite different to the other two farmlets across a range of functional groups while Farmlets B and C do not differ substantially in any of the groups over time.
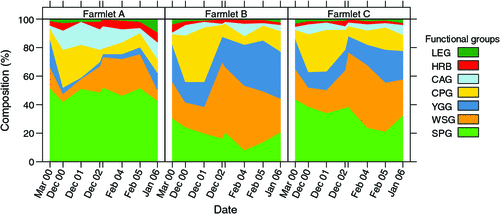
|
More details of each functional group and the variability of the measurements between paddocks and farmlets are presented in Fig. 2. This shows the individual paddock data together with the farmlet mean botanical composition and confidence intervals determined using local regression, likelihood and density estimations using the locfit function in R (R Development Core Team 2009). Results for the paddocks that were re-sown (six on Farmlet A and for one on each of Farmlets B and C) are presented with a different symbol to aid in interpretation.
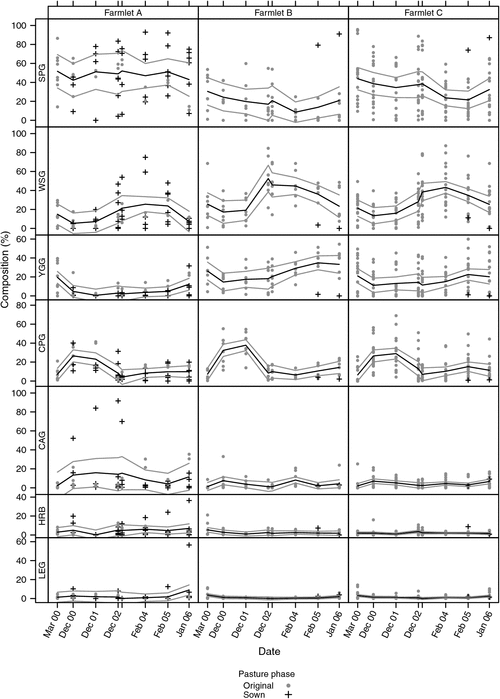
|
The statistical significance of the differences between farmlets for each functional group over time can most readily be seen by viewing the log-odds ratios presented in Fig. 3 in conjunction with the trends in percent composition in Fig. 2. At the first ‘baseline’ assessment in March 2000, before the commencement of farmlet treatments on 1 July 2000, there were no significant differences (P > 0.05) between farmlets.
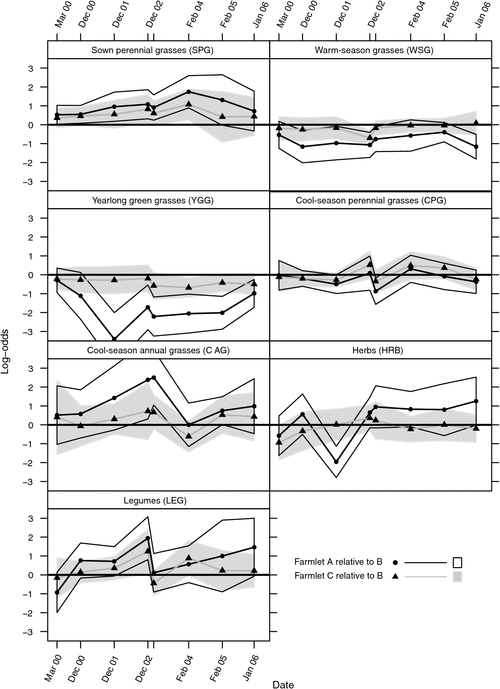
|
Figure 2 shows that differences between Farmlets A (sown and high fertility) and B (typical, control farmlet) were more pronounced and significant (P < 0.05) than those between Farmlets B and C (intensive rotation grazing), which were not significant (P > 0.05). Compared with Farmlet B, Farmlet A had a significantly (P < 0.05) greater level of SPG over most measurement dates. However, it is notable that the only two Farmlet A paddocks that were not renovated (A7 and A8) both had low levels of SPG (less than 20%) at the final measurement having both declined from having over 65% SPG in March 2000.
There was a corresponding significantly (P < 0.05) lower level of WSG and YGG on Farmlet A compared with Farmlet B. In late 2002, Farmlet A had higher levels of CAG, due largely to the sowing of Lolium multiflorum, classified here as an annual because of its limited persistence, on paddocks A1 and A6 over 2 years (2001 and 2002), and of herbs over the latter measurement dates, due largely to the high percentage of chicory which established following being sown (together with lucerne and phalaris) on one paddock only (A1) in 2004. The levels of CPG were not significantly (P > 0.05) different between any of the farmlets.
While the levels of legumes were generally low during the mid to late summer assessments of botanical composition, the percentage of legumes on Farmlet A were generally slightly higher than on Farmlet B, especially following the sowing of lucerne (together with chicory and phalaris), on paddock A1 in 2004.
It is clear that, until one paddock of each of Farmlets B (B6) and C (C17) was renovated in 2004, the percentages of SPG were declining steeply while the percentages of the WSG were rising steeply on both Farmlets B and C. After sowing, those two paddocks recorded high levels of SPG and correspondingly low levels of WSG, YGG, CPG with little or no impact on the sown legumes (Fig. 2).
In order to explore possible relationships behind the differences described above, a series of exploratory analyses were carried out using pairplots to help determine the most important variables and the levels of collinearity between them. These revealed that by far the largest correlations were the substantial curvilinear negative relationships between SPG and WSG (Pearson correlation coefficient of 0.69) and YGG (0.51).
The multivariate RDA conducted on the seven functional groups and their relationships with a wide array of explanatory variables [FarmID, PaddockID, measurement Date, Phase (original or sown), average DSE.days.ha, GrazeRatio (days grazed/period days), soil N, P, S, temperature index and modelled available soil water] showed that the only significant factors affecting percentage composition of functional groups were, in decreasing order of significance, soil phosphorus, Phase, PaddockID and Date (P < 0.01). Nevertheless, the variability of the dataset was such that the majority of variation was not able to be explained, as the sum of all of the canonical eigenvalues for this analysis was only 0.194. Axis-1 and axis-2 explained some 81 and 13%, respectively, of the sum of all canonical eigenvalues.
The triplot (which identifies individual measurements on all Paddocks) and biplot in Fig. 4a, and b, respectively show the strong negative correlation between SPG and both WSG and YGG described above. The triplot also clearly shows the wide separation of Paddocks for the three farmlets with Farmlet A paddocks being associated most closely with SPG, herbs, CAG and legumes as well as P. In contrast, Farmlet C and especially Farmlet B paddocks tended to be scattered towards the left side of axis-1, being more closely associated with WSG and YGG. The exceptions for these two farmlets were paddocks B6 and C17, both of which were sown in 2004, which were more closely aligned with the A paddocks and related explanatory variables towards the right side of axis-1.
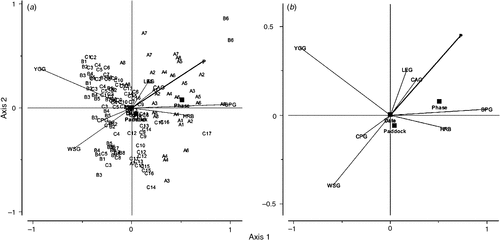
|
The significant effect of P (P < 0.01) was closely correlated with the functional groups CAG, SPG, legumes and herbs. The SPG were also closely related to the renovation of pastures indicated by the symbol for the nominal variable Phase (original or sown pasture).
When the RDA multivariate approach was used to explore the relationship between individual pasture species and explanatory variables, the differences between species were significantly associated with Phase, Paddock and Date (P < 0.01) and Farm and soil S level (P = 0.01). However, there was considerable unexplained variation as the sum of all eigenvalues was only 0.135 (data not presented).
The greatest proportions of variation were explained by species such as the SPG Festuca arundinacea, the volunteer, YGG Poa sieberiana and the volunteer, WSG Themeda australis. The species most closely correlated with sowing (Phase) were Festuca arundinacea, which was sown into paddocks A2–A6 as well as B6 and C17, and also those species only sown in one paddock, namely Medicago sativa (A1) and Cichorium intybus (A1), in two paddocks, such as Lolium multiflorum or in three paddocks, such as Dactylis glomerata (A3–A5). Pasture renovation was also associated with several weedy species such as Cirsium vulgare and Eleusine tristachya.
Species most negatively correlated with sowing Phase were species such as Bothriochloa macra and Anthoxanthum odoratum. Species unrelated to Phase included species which were sown into several paddocks, such as Phalaris aquatica and Lolium perenne, but were less obviously associated with sowing in this trial as they were also present in considerable proportions in some paddocks from the commencement of the trial, having been sown more than 10 years earlier. Other species unrelated to sowing Phase included Trifolium repens, which would have been broadcast sown many times in the past and was sown or oversown into many paddocks during this trial as well as some native perennial grass species such as Microlaena stipoides or volunteer species such as Paspalum dilatatum.
Investigations of the characteristics of species which were most important to changes in species and their functional group percentages over time were also carried out using multivariate RDA with the mean trends of proportions shown in Fig. 5. The most significant characteristics were establishment status (sown/volunteer) (sum of eigenvalues of 0.463) where the significant explanatory variables were Phase, PaddockID and P (P < 0.01). The next most significant category was origin (native/introduced) (sum of eigenvalues of 0.46) explained significantly by Phase, PaddockID and soil S (P < 0.01). The physiology category (C3/C4) (sum of eigenvalues of 0.21) showed that C3 and C4 species had a significant (P < 0.01) positive and negative correlation, respectively, with soil P. Other categories such as seasonality, life cycle and plant type were also explored but were found to have few significant associations.
|
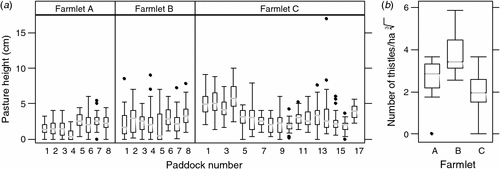
Investigation of the variability in compressed pasture height uniformity across farmlets in late winter (August) 2006 revealed that Farmlet A paddocks tended to have been grazed relatively evenly and close to the ground, whereas Farmlets B and C was considerably more variable suggesting that either ‘patch’ grazing was occurring and/or that more tussocky grasses persisted on these two farmlets (Fig. 6a). Of the two latter farmlets, Farmlet B was observed to be noticeably more ‘patchy’ with some areas within paddocks varying from almost bare ground to areas of large tussocks of ungrazed native species such as Poa sieberiana. In the case of Farmlet C, the average pasture height was much higher than Farmlet A due mostly to the accumulation of substantial quantities of grassy herbage during the long rest periods between grazings.
|
The number of thistles was assessed on each farmlet in August 2006 as an indicator of pasture degradation; this showed that Farmlet B had a significantly higher numbers of thistles (P < 0.01) (mean of 68/ha) compared with both Farmlets A (24) and C (13), which were not significantly different (P > 0.05) from each other (Fig. 6b).
Discussion
This study of three whole farmlets has provided considerable insight into the challenges of attempting to improve, or even to maintain botanical composition across whole farms in this summer-rainfall environment, albeit over 6 years of below median soil moisture conditions (Behrendt et al. 2013c). Of particular note is that the changes observed over the three farmlets occurred under grazing and management conditions which were considered to be realistic by producer members of the Cicerone Project (Edwards et al. 2013). Thus, this trial has demonstrated at paddock and farmlet scale, the effects of pasture and grazing management called for by Kemp and Dowling (2000) in their review of the Temperate Pastures Sustainability Key Program, when they noted the need for further experiments at a paddock scale.
The botanical composition results described here clearly demonstrate the high degree of variability which can occur between paddocks within whole farmlets and across variable seasons. This is to be expected as it is well known that factors such as the original composition, climate, soil type, fertiliser and degree of disturbance, such as sowing of introduced species and grazing management, can all impact on botanical composition (Garden et al. 2000a). As shown by the frequent significance of farm management interventions at the paddock level in this trial, such as sowing and fertilising, botanical composition needs to be managed on a paddock-by-paddock basis (Dowling et al. 2005).
Thus, the results of this farmlet trial support the hypothesis that, the renovation of pastures and maintenance of high levels of soil fertility, practised on the Farmlet A system, was able to result in a higher level of SPG and legumes than was the typical system (Farmlet B), albeit at considerable cost as considered by Scott et al. (2013a). While there was clear evidence of the changes brought about by sowing pastures and fertilising on six of the eight paddocks of Farmlet A, it was also apparent that the two Farmlet A paddocks which were not renovated experienced diminished SPG, presumably as a consequence of having been used to carry higher stocking densities than would have been required had the other paddocks not been renovated. This response to higher soil fertility levels is consistent with the findings of Saul et al. (1998) in Victoria and of Cook et al. (1978) on the Northern Tablelands of NSW who found that superphosphate applications contributed to more stable pastures.
However, in spite of the higher level of inputs on Farmlet A, the proportion of SPG on this farmlet declined from a mean of 52 to 43% over the 6 years of measurement. We suggest that the primary reason for this decline was the gradually increased stocking rate carried by Farmlet A, which reached a maximum annual average stocking rate of 12.6 DSE/ha (48% above Farmlet B) compared with Farmlets B (8.5 DSE/ha) and C (7.7 DSE/ha) (9% below Farmlet B) (Hinch et al. 2013). The influence of farmlet management factors such as pasture renovation, fertiliser and grazing management on the emergent property of stocking rate and the interactions between these factors and pasture composition, herbage mass and livestock production are beyond the scope of this paper; they are considered further in related papers by Behrendt et al. (2013a, 2013b).
The significant relationships found between species presence and their categories relating to establishment, origin and physiology together with the significant explanatory variables suggest strongly that botanical composition was most readily improved by sowing introduced, C3 species while increasing soil P and S levels through the addition of fertiliser. The positive effect of sowing was demonstrated not only on Farmlet A but also on the two paddocks renovated on Farmlets B and C. A qualification is noted that, in some situations, such as where shallow soils limit persistence, replacing persistent native grasses with SPG may not be the best long-term strategy.
The second hypothesis, that the intensive rotational grazing system (Farmlet C) would result in the retention of a better botanical composition compared with the typical district grazing system (Farmlet B), was not supported by the results of this study which showed that on Farmlet C, the proportion of SPG declined from an average of 44 to 32%, a similar reduction to that observed on Farmlet B (from 31 to 21%).
In addition, the low proportion of legumes on Farmlet C, which may have been due in part to the high levels of grass-dominant DM, which accumulated during the long rest periods, is likely to have contributed to a lower quality diet resulting in a lower stocking rate (7.7 DSE/ha) than the initial target stocking rate for that Farmlet (15 DSE/ha). It is noteworthy that, in spite of large differences between farmlet treatments in the length of graze and rest periods and substantial changes in stocking rate, neither of these factors was found to have significantly affected the changes in abundance of either functional group or individual pasture species. This may have been related to the generally drier than average available soil water which was evident during most of the farmlet systems trial (Behrendt et al. 2013c) and which would have limited the growth of pastures during rest periods.
In general, it is known that overgrazing can result in weedy pastures while undergrazing can lead to the exclusion of lower growing legumes (Kemp et al. 1996; Dowling et al. 2006). A limitation of the published evidence on the effects of grazing management is that too often studies have used continuous grazing as the control treatment (Earl and Jones 1996; Garden et al. 2000b), which disregards grazing practices more typical on farms in this region such as the employment of the ‘flexible rotational grazing’ approach chosen by producer members of the Cicerone Board as the ‘control’ grazing treatment for comparison with intensive rotational grazing.
Dowling et al. (2005) conducted an experiment to compare the effects of continuous and time-control grazing (a form of intensive rotational grazing) on grassland components in south-eastern Australia. They reported that time-control grazing did not appear to have any consistent benefits on the perennial grass component but that short-term changes in management caused changes in the composition of C3 perennial grasses. This is supported by the findings of this trial, which showed that intensive rotational grazing did not markedly enhance the SPG, nor other desirable components of the pastures.
Grazing management can have major effects on the composition of pastures, especially on the Northern Tablelands when high stocking rates are carried throughout winter allowing the selective grazing of preferred species (Cook et al. 1978). During periods of high temperature, the subtropical C4 species (such as Bothriochloa macra) are known to have the potential for invading pastures, especially in lower fertility sites, resulting in lower growth potential during the cooler months of the year (Cook et al. 1978) when the nutritional needs of pregnant ewes are greatest.
Other studies have shown how grazing management can be used to shift the balance from weedy to desirable species (Michael 1968; Lodge and Whalley 1985) but the benefits of any deferment of grazing appear to be dependent on whether the period of rest coincides with favourable seasonal conditions and the reproductive phase of the plant (Dowling et al. 2006). In the case of the Cicerone farmlet experiment, conditions were often unfavourable for growth during periods of rest and this may explain why there were few significant effects of deferred grazing on pasture composition.
The use of plant functional groups, which are commonly based on sets of traits (e.g. morphological, physiological, environmental responses) common to those species (Nicholas et al. 1998), was found to have been useful in this study as the levels of statistical significance that were associated with functional groups were considerably higher than those for analyses of individual species.
The significant relationships determined between functional groups and soil parameters and management interventions are consistent with the results obtained by López et al. (2006) in New Zealand. In a related study in New Zealand (Zhang et al. 2005), soil P and the rate of annual fertiliser P applications were found to be important determinants of fertility-responsive grass and legumes functional groups as was the case in this trial with Farmlet A.
The negative relationship between SPG and WSG and YGG was no doubt influenced by the fact that the latter two groups were mostly killed by herbicide before sowing. The relationship was also influenced to a considerable extent by the influence of soil P on both SPG and WSG. The occasional significance of soil S, in addition to soil P was most likely due to the fact that the target levels of both these nutrients were managed together and the two nutrients were supplied together in the most commonly used fertiliser on this trial, single superphosphate.
Legume persistence was diminished in the face of the dry seasons experienced throughout most of the trial. The attempts made to establish legume on all farmlets over the 6.5 years through over sowing white clover when fertilising and whenever pastures were renovated were less successful than desired. In a study of white clover persistence over 30 years in the summer-rainfall environment of the Northern Tablelands of NSW, moisture deficit during summer was found to be critical in determining clover presence in the following spring (Hutchinson et al. 1995). White clover is known to be a poor competitor with companion grasses (Curll et al. 1985) and persistence can be affected by competition during both the establishment phase and also in developed swards (Lane et al. 2000).
An alternative approach to increasing the proportion of legumes that could have been attempted and may have been more successful than oversowing, is the direct drilling of white clover into the pasture swards in autumn. This practice was found to be highly successful by Scott et al. (2000) who direct drilled white clover in April into a phalaris-dominant pasture after temporarily halting the growth of the dominant perennial grass phalaris with a single application of a non-systemic, knockdown herbicide. In subsequent seasons, this companion clover was associated with much greater persistence of phalaris, even under continuous grazing, compared with a phalaris-dominant pasture without clover which lost much of its phalaris content within just 3 years (Scott et al. 2000).
After experiencing several years of relatively low levels of legumes on Farmlet A, a pasture was sown to a strongly perennial legume, lucerne, in conjunction with a grass and a herb. This was successful in lifting the legume percent on that one paddock (A1) substantially with an overall positive effect on mean legume percent across the farmlet.
The lower proportion of legume in both of the moderate fertility systems (Farmlets B and C), in comparison to Farmlet A, was likely to have been related not only to dry seasons/moisture stress, but also to the strong dominance of native grasses and the lower levels of available P and S. The low legume content on Farmlet C may also have been associated with the greater competition for light imposed by the high herbage mass accumulated (typically 2300 kg DM/ha) during the long rest periods on this farmlet. This is consistent with the shading effect, observed by Dowling et al. (2006), who reported reduced legume persistence under rotational grazing as did Chapman et al. (2003).
While too much inference should not be drawn from the individual species data, it is noteworthy that the retention of SPG, such as Phalaris aquatica and Festuca arundinacea, on Farmlet A, even during years of low soil moisture, and under higher stocking rates, reflects the higher soil fertility of Farmlet A. In addition, the grazing-tolerance (Ayres et al. 2000) and deep-rooting habit of these grasses are likely to have contributed to their superior recovery and tolerance of drought stress. These species can also become dormant under stresses of low soil moisture and high temperature (Kemp and Culvenor 1994; Watson et al. 2001), while the good summer growth of Festuca arundinacea allows it to compete well with warm-season C4 grasses (Oram and Lodge 2003). The short-term persistence of the so-called ‘high performance’ pastures based on Lolium multiflorum, commonly promoted by commercial firms in the region, was to be expected; it is likely that the relatively dry conditions experienced hastened the demise of this species, which was sown in only two paddocks of Farmlet A.
Generally, the proportion of annual weedy grasses such as Vulpia species was low across the farmlets reflecting the strong dominance of perennial species within this trial. One less desirable warm-season perennial grass species that was noticeable in closely grazed paddocks was Eleusine tristachya, which is common in degraded pastures in the region (Scott et al. 2000). Being readily recognised due to a distinctive inflorescence, which is visible over most seasons, this species may serve as a useful indicator of pasture degradation in this region.
Selective grazing by sheep and cattle is known to create a mosaic of different heights, which is most clearly marked during periods when there is surplus herbage. Correll et al. (2003) reported that the rising-plate meter was a suitable method which provided detailed descriptions of the sward structure of both extensively and intensively grazed pastures. The present trial showed that farmlet B pastures were more spatially variable compared with the intensively grazed pastures on Farmlet A and those recently grazed on Farmlet C. The dominance of Themeda australis and Poa sieberiana observed on some paddocks of Farmlets B and C could be an indication of selective avoidance by sheep which is more common in pastures with large herbage masses (Arnold 1960).
The increased numbers of thistles on Farmlet B paddocks, which generally carried low stocking densities over relatively long grazing periods, suggests that sheep grazing on Farmlet B paddocks had a greater chance to repeatedly select species of their preference while avoiding other species such as thistles. Grace et al. (2002), who investigated the control of saffron thistle (Carthamus lanatus) on the Northern Tablelands through the use of strategic grazing with sheep, found that as little as 2 cm of pasture cover reduced seedling emergence by 96% compared with bare ground. They found that pastures had fewer thistles when perennial grasses accounted for over 35% of groundcover and when pastures were rotationally grazed compared with continuous grazing.
It is clear that the findings from this grazed farmlet trial would have been enhanced had it been possible to continue the project at least through to a period of above average rainfall conditions so that the effects of higher moisture conditions on relative farmlet performance could have been measured. Such problems have been acknowledged by Jones et al. (1995) who pointed out the value of long-term grazing trials, especially in relation to the effects of climate, soil fertility and grazing management on botanical composition and the flow-on effects for sustainability.
The levels of SPG in pastures have been related to the sustainability of grazing systems (Graham et al. 2003) as there is a need for the pastures to remain productive and resilient over the long-term if livestock enterprises are to remain viable. Perennial grasses are considered to be more effective at reducing the development of soil acidity (Ridley et al. 1990), compared with annual grasses and broad-leaf weeds. They utilise more soil water, which can reduce the risk of rising watertables, and the ground cover they provide over summer can also reduce the risk of erosion from storms which are commonly experienced on the Northern Tablelands. Scott et al. (2000) found that deep-rooted perennial grasses, combined with a persistent legume, improved water and N capture and thereby contributed to more profitable and sustainable grazed systems.
Management can clearly have large effects on botanical composition; the challenge now is to learn how to optimise composition to ensure that the feed supply for grazing animals is enhanced over the long-term while optimising profit. This is discussed further by Behrendt et al. (2013b). Results concerning the effects of farmlet treatment on pasture growth and quality as well as the balance between pasture supply and animal demand are addressed in subsequent papers in this series (Shakhane et al. 2013a, 2013b).
Acknowledgements
The Cicerone Project was generously funded by Australian taxpayers and woolgrowers through Australian Wool Innovation. The Cicerone Board is gratefully acknowledged for providing support and access to the study site. The continuous farm management support provided by Mr Justin Hoad and coordination support from Ms Caroline Gaden was very much appreciated. The financial and in-kind support of the University of New England is acknowledged as is the in-kind support of CSIRO Livestock Industries. Valuable database support was provided by Mr Colin Lord, Mr Jim Cook and Mr Dion Gallagher.
References
Arnold G (1960) Selective grazing by sheep of two forage species at different stages of growth. Australian Journal of Agricultural Research 11, 1026–1033.| Selective grazing by sheep of two forage species at different stages of growth.Crossref | GoogleScholarGoogle Scholar |
Ayres JF, McPhee MJ, Turner AD, Curll ML (2000) The grazing value of tall fescue (Festuca arundinacea) and phalaris (Phalaris aquatica) for sheep production in the northern tablelands of New South Wales. Australian Journal of Agricultural Research 51, 57–68.
| The grazing value of tall fescue (Festuca arundinacea) and phalaris (Phalaris aquatica) for sheep production in the northern tablelands of New South Wales.Crossref | GoogleScholarGoogle Scholar |
Behrendt K, Cacho O, Scott JM, Jones R (2013a) Optimising pasture and grazing management decisions on the Cicerone Project farmlets over variable time horizons. Animal Production Science 53, 796–805.
| Optimising pasture and grazing management decisions on the Cicerone Project farmlets over variable time horizons.Crossref | GoogleScholarGoogle Scholar |
Behrendt K, Scott JM, Cacho O, Jones R (2013b) Simulating the impact of fertiliser strategies and prices on the economics of developing and managing the Cicerone Project farmlets under climatic uncertainty. Animal Production Science 53, 806–816.
| Simulating the impact of fertiliser strategies and prices on the economics of developing and managing the Cicerone Project farmlets under climatic uncertainty.Crossref | GoogleScholarGoogle Scholar |
Behrendt K, Scott JM, Mackay DF, Murison R (2013c) Comparing the climate experienced during the Cicerone farmlet experiment against the climatic record. Animal Production Science 53, 658–669.
| Comparing the climate experienced during the Cicerone farmlet experiment against the climatic record.Crossref | GoogleScholarGoogle Scholar |
Bell AK, Allan CJ (2000) PROGRAZE – an extension package in grazing and pasture management. Australian Journal of Experimental Agriculture 40, 325–330.
| PROGRAZE – an extension package in grazing and pasture management.Crossref | GoogleScholarGoogle Scholar |
Blair GJ (1997) Matching pastures to the Australian environment. In ‘Pasture production and management’. (Eds JV Lovett, JM Scott) pp. 88–109. (Inkata Press: Melbourne)
Chapman DF, McCaskill MR, Quigley PE, Thompson AN, Graham JF, Borg D, Lamb J, Kearney G, Saul GR, Clark SG (2003) Effects of grazing method and fertiliser inputs on the productivity and sustainability of phalaris-based pastures in Western Victoria. Australian Journal of Experimental Agriculture 43, 785–798.
| Effects of grazing method and fertiliser inputs on the productivity and sustainability of phalaris-based pastures in Western Victoria.Crossref | GoogleScholarGoogle Scholar |
Cook S, Blair G, Lazenby A (1978) Pasture degeneration. II. The importance of superphosphate, nitrogen and grazing management. Australian Journal of Agricultural Research 29, 19–29.
| Pasture degeneration. II. The importance of superphosphate, nitrogen and grazing management.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE1cXhtF2qtb4%3D&md5=eb1e445201f5793668a4980db6b8a9a1CAS |
Correll O, Isselstein J, Pavlu V (2003) Studying spatial and temporal dynamics of sward structure at low stocking densities: the use of an extended rising-plate-meter method. Grass and Forage Science 58, 450–454.
| Studying spatial and temporal dynamics of sward structure at low stocking densities: the use of an extended rising-plate-meter method.Crossref | GoogleScholarGoogle Scholar |
Curll ML, Wilkins RJ, Snaydon RW, Shanmugalingam VS (1985) The effects of stocking rate and nitrogen fertilizer on a perennial ryegrass-white clover sward. Grass and Forage Science 40, 129–140.
| The effects of stocking rate and nitrogen fertilizer on a perennial ryegrass-white clover sward.Crossref | GoogleScholarGoogle Scholar |
Dowling PM, Kemp DR, Ball PD, Langford CM, Michalk DL, Millar GD, Simpson PC, Thompson RP (2005) Effect of continuous and time-control grazing on grassland components in south-eastern Australia. Australian Journal of Experimental Agriculture 45, 369–382.
| Effect of continuous and time-control grazing on grassland components in south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Dowling PM, Michalk DL, Kemp DR, Millar GD, Priest SM, King WM, Packer IJ, Holst PJ, Tarleton JA (2006) Sustainable grazing systems for the Central Tablelands of New South Wales. 2. Effect of pasture type and grazing management on pasture productivity and composition. Australian Journal of Experimental Agriculture 46, 457–469.
| Sustainable grazing systems for the Central Tablelands of New South Wales. 2. Effect of pasture type and grazing management on pasture productivity and composition.Crossref | GoogleScholarGoogle Scholar |
Earl J, Jones C (1996) The need for a new approach to grazing management – is cell grazing the answer? The Rangeland Journal 18, 327–350.
| The need for a new approach to grazing management – is cell grazing the answer?Crossref | GoogleScholarGoogle Scholar |
Edwards C, Gaden C, Marchant R, Coventry T, Dutton P, Scott JM (2013) Delivering extension and adult learning outcomes from the Cicerone Project by ‘comparing, measuring, learning and adopting’. Animal Production Science 53, 827–840.
| Delivering extension and adult learning outcomes from the Cicerone Project by ‘comparing, measuring, learning and adopting’.Crossref | GoogleScholarGoogle Scholar |
Garden DL, Dowling PM, Eddy DA, Nicol HI (2000a) A survey of farms on the Central, Southern and Monaro Tablelands of New South Wales: management practices, farmer knowledge of native grasses, and extent of native grass areas. Australian Journal of Experimental Agriculture 40, 1081–1088.
| A survey of farms on the Central, Southern and Monaro Tablelands of New South Wales: management practices, farmer knowledge of native grasses, and extent of native grass areas.Crossref | GoogleScholarGoogle Scholar |
Garden DL, Lodge GM, Friend DA, Dowling PM, Orchard BA (2000b) Effects of grazing management on botanical composition of native grass-based pastures in temperate south-east Australia. Australian Journal of Experimental Agriculture 40, 225–245.
| Effects of grazing management on botanical composition of native grass-based pastures in temperate south-east Australia.Crossref | GoogleScholarGoogle Scholar |
Garden DL, Dowling PM, Eddy DA, Nicol HI (2001) The influence of climate, soil, and management on the composition of native grass pastures on the central, southern, and Monaro tablelands of New South Wales. Australian Journal of Agricultural Research 52, 925–936.
| The influence of climate, soil, and management on the composition of native grass pastures on the central, southern, and Monaro tablelands of New South Wales.Crossref | GoogleScholarGoogle Scholar |
Garden DL, Ellis NJS, Rab MA, Langford CM, Johnston WH, Shields C, Murphy T, Holmberg M, Dassanayake KB, Harden S (2003) Fertiliser and grazing effects on production and botanical composition of native grasslands in south-east Australia. Australian Journal of Experimental Agriculture 43, 843–859.
| Fertiliser and grazing effects on production and botanical composition of native grasslands in south-east Australia.Crossref | GoogleScholarGoogle Scholar |
Grace BS, Whalley RDB, Sheppard AW, Sindel BM (2002) Managing saffron thistle in pastures with strategic grazing. The Rangeland Journal 24, 313–325.
| Managing saffron thistle in pastures with strategic grazing.Crossref | GoogleScholarGoogle Scholar |
Graham JF, Cullen BR, Lodge GM, Andrew MH, Christy BP, Holst PJ, Wang X, Murphy SR, Thompson AN (2003) SGS Animal Production Theme: effect of grazing system on animal productivity and sustainability across southern Australia. Australian Journal of Experimental Agriculture 43, 977–991.
| SGS Animal Production Theme: effect of grazing system on animal productivity and sustainability across southern Australia.Crossref | GoogleScholarGoogle Scholar |
Guppy CN, Edwards C, Blair GJ, Scott JM (2013) Whole-farm management of soil nutrients drives productive grazing systems: the Cicerone farmlet experiment confirms earlier research. Animal Production Science 53, 649–657.
| Whole-farm management of soil nutrients drives productive grazing systems: the Cicerone farmlet experiment confirms earlier research.Crossref | GoogleScholarGoogle Scholar |
Harris CA, Ayres JF (1997) The performance of set-stocked pastures on the northern tablelands of New South Wales through severe drought. In ‘Proceedings of the 12th annual conference of the Grassland Society of New South Wales’. pp. 136–137.
Harris W, Lazenby A (1974) Competitive interaction of grasses with contrasting temperature responses and water stress tolerances. Australian Journal of Agricultural Research 25, 227–246.
| Competitive interaction of grasses with contrasting temperature responses and water stress tolerances.Crossref | GoogleScholarGoogle Scholar |
Hill JO, Simpson RJ, Moore AD, Graham P, Chapman DF (2004) Impact of phosphorus application and sheep grazing on the botanical composition of sown pasture and naturalised, native grass pasture. Australian Journal of Agricultural Research 55, 1213–1225.
| Impact of phosphorus application and sheep grazing on the botanical composition of sown pasture and naturalised, native grass pasture.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXhtFWqsLjP&md5=d09b717cd6cf3dd9f1d51be553f23fb5CAS |
Hill JO, Simpson RJ, Wood JT, Moore AD, Chapman DF (2005) The phosphorus and nitrogen requirements of temperate pasture species and their influence on grassland botanical composition. Australian Journal of Agricultural Research 56, 1027–1039.
| The phosphorus and nitrogen requirements of temperate pasture species and their influence on grassland botanical composition.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXhtFChsLbI&md5=d0c8c7cb2c619659424d978a72f77f5cCAS |
Hinch GN, Hoad J, Lollback M, Hatcher S, Marchant R, Colvin A, Scott JM, Mackay D (2013) Livestock weights in response to three whole-farmlet management systems. Animal Production Science 53, 727–739.
| Livestock weights in response to three whole-farmlet management systems.Crossref | GoogleScholarGoogle Scholar |
Hutchinson K, King K, Wilkinson D (1995) Effects of rainfall, moisture stress, and stocking rate on the persistence of white clover over 30 years. Australian Journal of Experimental Agriculture 35, 1039–1047.
| Effects of rainfall, moisture stress, and stocking rate on the persistence of white clover over 30 years.Crossref | GoogleScholarGoogle Scholar |
Jones RM, Jones RJ, McDonald CK (1995) Some advantages of long-term grazing trials, with particular reference to changes in botanical composition. Australian Journal of Experimental Agriculture 35, 1029–1038.
| Some advantages of long-term grazing trials, with particular reference to changes in botanical composition.Crossref | GoogleScholarGoogle Scholar |
Jones RE, Dowling PM, Michalk DL, King WM (2006) Sustainable grazing systems for the Central Tablelands of New South Wales. 5. A bioeconomic framework for assessing the long-term economic benefits of grazing management tactics and implications for sustainability. Australian Journal of Experimental Agriculture 46, 495–502.
| Sustainable grazing systems for the Central Tablelands of New South Wales. 5. A bioeconomic framework for assessing the long-term economic benefits of grazing management tactics and implications for sustainability.Crossref | GoogleScholarGoogle Scholar |
Kemp DR, Culvenor RA (1994) Improving the grazing and drought tolerance of temperate perennial grasses. New Zealand Journal of Agricultural Research 37, 365–378.
| Improving the grazing and drought tolerance of temperate perennial grasses.Crossref | GoogleScholarGoogle Scholar |
Kemp DR, Dowling PM (2000) Towards sustainable temperate perennial pastures. Australian Journal of Experimental Agriculture 40, 125–132.
| Towards sustainable temperate perennial pastures.Crossref | GoogleScholarGoogle Scholar |
Lane LA, Ayres JF, Lovett JV (2000) The pastoral significance, adaptive characteristics, and grazing value of white clover (Trifolium repens L.) in dryland environments in Australia: a review. Australian Journal of Experimental Agriculture 40, 1033–1046.
| The pastoral significance, adaptive characteristics, and grazing value of white clover (Trifolium repens L.) in dryland environments in Australia: a review.Crossref | GoogleScholarGoogle Scholar |
Lees JW, Reeve IJ (1994) Temperate Pasture Sustainability Key Program 1994. Producer Survey. Report to the Meat Research Corporation. Rural Development Centre, University of New England, Armidale, NSW.
Liang KY, Zeger SL (1986) Longitudinal data analysis using generalized linear models. Biometrika 73, 13–22.
| Longitudinal data analysis using generalized linear models.Crossref | GoogleScholarGoogle Scholar |
Lodge GM, Whalley RDB (1985) The manipulation of species composition of natural pastures by grazing management on the Northern Slopes of New South Wales. Australian Rangelands Journal 7, 6–16.
| The manipulation of species composition of natural pastures by grazing management on the Northern Slopes of New South Wales.Crossref | GoogleScholarGoogle Scholar |
Lodge GM, Whalley RDB (1989) ‘Native and natural pastures on the Northern Slopes and Tablelands of New South Wales: a review and annotated bibliography.’ (NSW Agriculture and Fisheries: Tamworth)
López IF, Valentine I, Lambert MG, Hedderley DI, Kemp PD (2006) Plant functional groups in a heterogeneous environment. New Zealand Journal of Agricultural Research 49, 439–450.
| Plant functional groups in a heterogeneous environment.Crossref | GoogleScholarGoogle Scholar |
Mason WK, Kay G (2000) Temperate Pasture Sustainability Key Program: an overview. Australian Journal of Experimental Agriculture 40, 121–123.
| Temperate Pasture Sustainability Key Program: an overview.Crossref | GoogleScholarGoogle Scholar |
Mason WK, Lamb K, Russell B (2003) The Sustainability Grazing Systems Program: new solutions for livestock producers. Australian Journal of Experimental Agriculture 43, 663–672.
| The Sustainability Grazing Systems Program: new solutions for livestock producers.Crossref | GoogleScholarGoogle Scholar |
Michael PW (1968) Control of the biennial thistle Onopordum by Amitrole and five perennial grasses. Australian Journal of Experimental Agriculture and Animal Husbandry 8, 332–339.
| Control of the biennial thistle Onopordum by Amitrole and five perennial grasses.Crossref | GoogleScholarGoogle Scholar |
Michalk DL, Dowling PM, Kemp DR, King WM, Packer IJ, Holst PJ, Jones RE, Priest SM, Millar GD, Brisbane S, Stanley DF (2003) Sustainable grazing systems for the Central Tablelands, New South Wales. Australian Journal of Experimental Agriculture 43, 861–874.
| Sustainable grazing systems for the Central Tablelands, New South Wales.Crossref | GoogleScholarGoogle Scholar |
Nicholas PK, Kemp PD, Barker DJ (1998) Biodiversity, stability and pasture management – the role of functional groups. In ‘Agronomy, growing a greener future? Proceedings of the 9th Australian agronomy conference’. Charles Sturt University, Wagga Wagga, NSW. (Eds DL Michalk, JE Pratley) (The Regional Institute) Available at http://www.regional.org.au/au/asa/1998/3/067nicholas.htm [Verified 26 June 2012]
Oram RN, Lodge GM (2003) Trends in temperate Australian grass breeding and selection. Australian Journal of Agricultural Research 54, 211–241.
| Trends in temperate Australian grass breeding and selection.Crossref | GoogleScholarGoogle Scholar |
R Development Core Team (2009) R: a language and environment for statistical computing. Available at http://www.R-project.org [Verified 21 June 2012]
Reeve IJ, Kaine G, Lees JW, Barclay E (2000) Producer perceptions of pasture decline and grazing management. Australian Journal of Experimental Agriculture 40, 331–341.
| Producer perceptions of pasture decline and grazing management.Crossref | GoogleScholarGoogle Scholar |
Ridley A, Slattery W, Helyar K, Cowling A (1990) Acidification under grazed annual and perennial grass based pastures. Australian Journal of Experimental Agriculture 30, 539–544.
| Acidification under grazed annual and perennial grass based pastures.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3MXlvVaisr8%3D&md5=359aa2bed69ca7186e914d789049e587CAS |
Rogers R, Whalley R (1989) Relationship Between Diaspore Characteristics and Distribution of Grasses Around Sheep Camps on the Northern Tablelands of New-South-Wales. Australian Journal of Botany 37, 501–510.
| Relationship Between Diaspore Characteristics and Distribution of Grasses Around Sheep Camps on the Northern Tablelands of New-South-Wales.Crossref | GoogleScholarGoogle Scholar |
Sanford P, Cullen BR, Dowling PM, Chapman DF, Garden DL, Lodge GM, Andrew MH, Quigley PE, Murphy SR, King WM, Johnston WH, Kemp DR (2003) Sustainable Grazing Systems Theme: effect of climate, soil factors and management on pasture production and stability across the high rainfall zone of southern Australia. Australian Journal of Experimental Agriculture 43, 945–959.
| Sustainable Grazing Systems Theme: effect of climate, soil factors and management on pasture production and stability across the high rainfall zone of southern Australia.Crossref | GoogleScholarGoogle Scholar |
Saul GR, Waller R, Warn L, Hill R (1998) Grazing management for sheep pastures in Victoria – update on current research results. In ‘Proceedings of the 39th Grassland Society of Victoria conference’. Mornington, Victoria. (Eds R Stockdale, L Warn) pp. 87–95. (Grassland Society of Victoria Inc.: Tooborac, Vic.)
Scott JM, Hutchinson KJ, King K, Chen W, McLeod M, Blair GJ, White A, Wilkinson D, Lefroy RDB, Cresswell H, Daniel H, Harris C, MacLeod DA, Blair N, Chamberlain G (2000) Quantifying the sustainability of grazed pastures on the Northern Tablelands of New South Wales. Australian Journal of Experimental Agriculture 40, 257–265.
| Quantifying the sustainability of grazed pastures on the Northern Tablelands of New South Wales.Crossref | GoogleScholarGoogle Scholar |
Scott JF, Scott JM, Cacho OJ (2013a) Whole-farm returns show true profitability of three different livestock management systems. Animal Production Science 53, 780–787.
| Whole-farm returns show true profitability of three different livestock management systems.Crossref | GoogleScholarGoogle Scholar |
Scott JM, Gaden CA, Edwards C, Paull DR, Marchant R, Hoad J, Sutherland H, Coventry T, Dutton P (2013b) Selection of experimental treatments, methods used and evolution of management guidelines for comparing and measuring three grazed farmlet systems. Animal Production Science 53, 628–642.
| Selection of experimental treatments, methods used and evolution of management guidelines for comparing and measuring three grazed farmlet systems.Crossref | GoogleScholarGoogle Scholar |
Scott JM, Munro M, Rollings N, Browne W, Vickery PJ, Macgregor C, Donald GE, Sutherland H (2013c) Planning for whole-farm systems research at a credible scale: subdividing land into farmlets with equivalent initial conditions. Animal Production Science 53, 618–627.
| Planning for whole-farm systems research at a credible scale: subdividing land into farmlets with equivalent initial conditions.Crossref | GoogleScholarGoogle Scholar |
Shakhane LM, Mulcahy C, Scott JM, Hinch GN, Donald GE, Mackay DF (2013a) Pasture herbage mass, quality and growth in response to three whole-farmlet management systems. Animal Production Science 53, 685–698.
| Pasture herbage mass, quality and growth in response to three whole-farmlet management systems.Crossref | GoogleScholarGoogle Scholar |
Shakhane LM, Scott JM, Hinch GN, Mackay DF, Lord C (2013b) Estimating the balance between pasture feed supply and demand of grazing livestock in a farmlet experiment. Animal Production Science 53, 711–726.
| Estimating the balance between pasture feed supply and demand of grazing livestock in a farmlet experiment.Crossref | GoogleScholarGoogle Scholar |
Tothill JC, Hargreaves JNG, Jones RM (1978) BOTANAL – a comprehensive sampling and computing procedure for estimating pasture yield and composition. CSIRO Division of Tropical Crops and Pastures, Tropical Agronomy Technical Memorandum No. 8, St Lucia, Qld.
Vere DT, Campbell MH, Kemp DR (1993) ‘Pasture improvement budgets for the central and southern tablelands of New South Wales.’ (NSW Agriculture: Orange, NSW)
Waller RA, Sale PWG, Saul GR, Kearney GA (2001) Tactical versus continuous stocking in perennial ryegrass-subterranean clover pastures grazed by sheep in south-western Victoria – 1. Stocking rates and herbage production. Australian Journal of Experimental Agriculture 41, 1099–1108.
| Tactical versus continuous stocking in perennial ryegrass-subterranean clover pastures grazed by sheep in south-western Victoria – 1. Stocking rates and herbage production.Crossref | GoogleScholarGoogle Scholar |
Watson DJ, Avery A, Mitchell GJ, Chenner SR (2001) Grazing management to increase utilisation of phalaris-based pasture in dryland dairy systems. Australian Journal of Experimental Agriculture 41, 29–36.
| Grazing management to increase utilisation of phalaris-based pasture in dryland dairy systems.Crossref | GoogleScholarGoogle Scholar |
Whalley R, Robinson G, Taylor J (1976) General effects of management and grazing by domestic livestock on the rangelands of the Northern Tablelands of New South Wales. The Rangeland Journal 1, 174–190.
| General effects of management and grazing by domestic livestock on the rangelands of the Northern Tablelands of New South Wales.Crossref | GoogleScholarGoogle Scholar |
Wheeler JL, Pearson CJ, Robards GE (Eds) (1987) ‘Temperate pastures: their production, use and management.’ (CSIRO and Australian Wool Corporation: Melbourne)
Wolfe E, Lazenby A (1972) Bloat incidence and liveweight gain in beef cattle on pastures containing different proportions of white clover (Trifolium repens). Australian Journal of Experimental Agriculture 12, 119–125.
| Bloat incidence and liveweight gain in beef cattle on pastures containing different proportions of white clover (Trifolium repens).Crossref | GoogleScholarGoogle Scholar |
Zhang B, Valentine I, Kemp PD (2005) A decision tree approach modelling functional group abundance in a pasture ecosystem. Agriculture Ecosystems & Environment 110, 279–288.
| A decision tree approach modelling functional group abundance in a pasture ecosystem.Crossref | GoogleScholarGoogle Scholar |
Zuur AF, Ieno EN, Smith GM (2007) ‘Analysing ecological data.’ (Springer: New York)