

Close human presence reduces avoidance behaviour in commercial caged laying hens to an approaching human
L. E. Edwards A C D E , G. J. Coleman B and P. H. Hemsworth AA Animal Welfare Science Centre, The Melbourne School of Land and Environment, University of Melbourne, Parkville, Vic. 3010, Australia.
B Animal Welfare Science Centre, School of Psychology and Psychiatry, Monash University, Clayton, Vic. 3800, Australia.
C Past student of the former Australian Poultry Cooperative Research Centre, PO Box U242, University of New England, Armidale, NSW 2351, Australia.
D Present address: Animal Welfare and Biodiversity Research Group, Department of Natural Sciences, Unitec Institute of Technology, Carrington Road, Mt Albert, Private Bag 92025, Auckland Mail Centre, Auckland 1142, New Zealand.
E Corresponding author. Email: ledwards@unitec.ac.nz
Animal Production Science 53(12) 1276-1282 https://doi.org/10.1071/AN12342
Submitted: 28 September 2012 Accepted: 16 February 2013 Published: 21 May 2013
Journal Compilation © CSIRO Publishing 2013 Open Access CC BY-NC-ND
Abstract
The quality of human contact that hens are exposed to will determine the degree of fear of humans that they experience. This has consequences for the welfare of commercial laying hens, as hens that are afraid of humans will be regularly exposed to a fear-provoking stressor. Hens can be habituated to human presence using positive or neutral human–animal interactions, although the specific human behaviours that are considered positive or neutral by the hens are still being determined. This experiment investigated whether the proximity or duration of visual contact with a human affected fear of humans in commercial caged laying hens (n = 216). Commercial laying hens were exposed to daily visual human contact at one of three proximities (0, 0.75 or 1.50 m) and one of three durations (2, 30 or 90 s) in a 3 by 3 factorial design for a period of 28 days. Avoidance behaviour was assessed on Days –5, 15 and 30, and the plasma corticosterone response to handling was assessed on Days –5 and Day 30. Visual contact with a stationary human at close proximity (0 m) significantly (P = 0.03) reduced the avoidance response of commercial laying hens to an approaching human, although there was no clear effect of proximity on corticosterone response to handling. The duration of human contact had no effect on avoidance behaviour or corticosterone response. Stockpeople may consider working more closely to the cages in a non-threatening manner to reduce fear of humans in their flock.
Additional keywords: corticosterone, egg farm, human–animal relationship, welfare.
Introduction
Fear is an emotional reaction (Janczak 2010) and is generally considered a motivational state that normally gives rise to defensive behaviour or escape (McFarland 1981; Hogan 2008). Poultry are innately fearful of humans (Murphy and Duncan 1978), and farm animals that experience fear in the presence of stockpeople also experience reduced welfare and productivity due to repeated exposure to this stressor (Hemsworth and Coleman 2011). However, poultry are able to habituate to human presence and previous handling studies have repeatedly demonstrated that human contact of a positive nature can reduce fear of humans in poultry (Barnett et al. 1994; Hemsworth et al. 1994; Zulkifli and Siti Nor Azah 2004; GramL et al. 2008), as well as improving production and immune function, which is indicative of a reduced stress response (Jones and Hughes 1981; Gross and Siegel 1982; Hemsworth and Barnett 1989; Barnett et al. 1994; Hemsworth et al. 1994; Zulkifli et al. 2002; Zulkifli and Siti Nor Azah 2004). For example, an additional 12–15 min of daily visual contact with a stationary or slow-moving human improved egg production and reduced the avoidance behaviour and corticosterone response to human contact in caged laying hens (Barnett et al. 1994; Edwards et al. 2010).
Many scientists in studying fear have adopted a functional view and assessed the animal’s fear of humans on the basis of the avoidance behaviour of the animal to an approaching experimenter in a standard testing situation (Hemsworth and Coleman 2011). The majority of the poultry-handling studies described above have successfully used either tactile contact, such as stroking and gentle handling, or visual contact, such as standing still or moving slowly among the birds, to reduce fear of humans. In fact, visual contact has been shown to be equally effective in reducing fear of humans in poultry as is tactile contact, if not more so (Jones 1993; Zulkifli and Siti Nor Azah 2004). However, the effect of varying the duration of human contact on poultry has not been reported in the literature. In addition, the proximity of the visual contact with humans has been shown to influence the avoidance response of caged hens under experimental conditions, with visual contact at close proximity (<0.75 m) found to be more effective in reducing fear of humans than visual contact from further away (1.50 m) (Edwards et al. 2010). The present experiment investigated whether the habituation of caged laying hens to humans was influenced by the proximity or duration of visual human contact in a commercial situation.
Materials and methods
Treatment and experimental design
This experiment was conducted in four environmentally controlled laying sheds at a large commercial farm in central Victoria, Australia. The hens housed in these sheds were of the Hyline Brown strain. Each shed contained three rows of back-to-back cages, creating four aisles. Details of the sheds are presented in Table 1.

|
Nine human-contact treatments were imposed on the hens, each of them being a combination of one level of duration (2 s, 30 s or 90 s) and one level of distance (0 m, 0.75 m, 1.5 m) in a 3 by 3 factorial design. The treatments were imposed on focal cages that were located in the second tier of cages (~1 m in height) at evenly spaced intervals (~3.5 m) along one side of each row. Each of the nine human-contact treatments were blocked twice per row, creating a total of six treatment replicates per shed (n = 54 focal cages per shed). The experimental design in one shed is presented in Fig. 1.
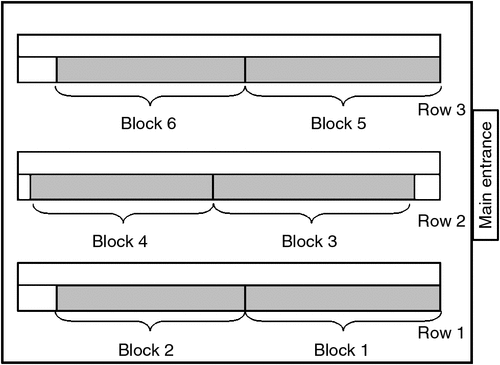
|
Focal cages on the left side of the row were used in Sheds 9 and 10, and the right side of the row was used in Sheds 11 and 12. This design was used to allow all locations in the shed to be sampled, avoiding any location-specific effects on the birds’ response to humans. The positions of the focal cages were staggered between rows to prevent birds in one experimental cage being able to see directly through the row to the experimental cage and its treatment in the adjacent row. However, the birds were able to see the handling treatments being applied to other cages within the same row.
The treatments were imposed daily for 28 days (from Day 2 to Day 29) between 0800 hours and 1300 hours on weekdays, and between 0900 hours and 1700 hours on weekends. The treatments took ~1 h per shed to impose. The treatments were imposed with the researcher standing directly in front of the experimental cage, or slightly to one side for the 1.50 m treatments, looking directly at the birds in that cage. The treatments were timed with a stopwatch, and all movements were made in a slow and predictable manner to minimise startling the hens. The direction of movement through the shed was randomised daily so that the direction of approach within the aisle and order in which the aisles were visited was varied. The researcher dressed in a standard manner for the daily treatments, wearing blue overalls and yellow gumboots. This attire was unfamiliar to the birds, as the stockpeople dressed in casual attire while at work.
Behavioural assessment
Birds were tested for fear of humans at the beginning (Day –5), middle (Day 15) and end (Day 30) of the experiment, using the approaching-human test and the stroll test, which were adapted from the behavioural tests used by Edwards (2009) to assess fear of humans in commercial caged laying hens. The human-contact treatments were not imposed on the testing days.
The approaching-human test (AHT) was administered to the focal cages in a consecutive order along each aisle, with the aisle order randomised within each shed. Prior to the commencement of each test, the researcher walked slowly along the aisle and stood on the opposite side of the aisle and adjacent to the cage before the focal cage to be tested. The researcher stood stationary in this position for 5 s, with her hands in her pockets for the entire test. This allowed the birds in the focal cage to adjust to the presence of the researcher, reducing the possibility of startling the birds as the test commenced. An assistant waited at a distance of 1 m from the researcher (visible to the hens in the focal cage) and timed the test in the following manner: after 5 s of the researcher waiting next to the focal cage, the assistant quietly notified the researcher verbally to commence the test; the researcher then stepped sideways so that she was standing directly in front of the focal cage but on the opposite side of the aisle (~0.9 m away); after 5 s, the researcher stepped forward, standing directly in front of the focal cage, with her torso contacting the feed trough (~0 m away); after a further 5 s, the researcher stepped backward across the aisle and stood opposite the focal cage; after 5 s, the researcher stepped forward again, to the position directly in front of the focal cage, after which the test ended. The total test took 20 s, consisting of two 5 s periods with the researcher standing on the opposite side of the aisle and two 5 s periods with the researcher standing directly in front of the cage. During each 5 s period, the researcher counted the maximum number of birds that had their head within the front 5 cm of the cage and the maximum number of birds that had their head out of the cage concurrently. At the end of each 5 s period, the researcher made a final point count of the number of birds that still had their heads in the front 5 cm of the cage. Thus, at the end of the AHT, the researcher had recorded three variables, measured every 5 s for 20 s. The resulting 12 variables were all significantly correlated with each other for each test day (Day –5, P ≤ 0.02, Day 15, P < 0.01, Day 30, P < 0.01), and were thus deemed sufficiently similar to convert to a single value for each focal cage. All 12 values for each focal cage were converted to z-scores and summed to create a single behavioural value (SumZ). A high value for the variable ‘SumZ’ indicates a greater number of birds at the cage front during testing and low avoidance of humans.
During the stroll test, the researcher walked along the middle of each aisle once at a standard speed (1 step/s), filming the birds in the second tier containing the focal cages, with a small hand-held video camera from ~0.60 m away. During video analysis, the number of birds with their heads out of each focal cage was counted as the researcher approached within a 1-cage distance of the focal cage. The focal cages were marked to allow easy identification during video analysis, and the number of birds in each focal cage was counted on each day of behavioural testing (Day –5, Day 15 and Day 30) to allow the proportion of hens with their head out of the cage as the researcher passed to be calculated accurately. This resulted in the variable ‘proportion of heads out’ (Prop HO), with a higher value indicating low avoidance of a passing human.
Physiological assessment
Six birds from each treatment (n = 54) were blood sampled on Day –5 and Day 30 of the experiment, to assess the corticosterone response of the hens to 4 min of restraint by a human.
All birds were sampled from one shed (Shed 10), with one bird sampled from each experimental cage. Shed 10 had three rows of cages, and all three were sampled simultaneously using three researchers. The focal cages were sampled in a sequential order from the front to the back of the shed. All birds were sampled after 1300 hours to reduce the effects of the diurnal increase in plasma corticosterone concentrations that occur in relation to egg laying (Beuving and Vonder 1977).
One bird from each focal cage was selected on the basis of its position in the cage as the researcher approached. Hens were alternately selected from the front and back of the cage. Once a bird was selected, the researcher caught only that hen. This avoided consistently selecting hens that were positioned at the cage front, and thus avoiding unintentionally selecting birds that may have a particular behavioural disposition. For example, birds at the cage front are closer to the feed trough and a dominant hen would potentially be positioned near the feed trough more often (Mankovich and Banks 1982; Keeling 1995). Also, birds closer to the front are less likely to be fearful of humans. This selection order was repeated along each row for all 18 focal cages.
All hens were held for a total of 4 min before blood sampling, because this duration has been previously demonstrated by Beuving and Vonder (1978) to be a sufficient amount of time to cause a substantial increase in the concentration of plasma corticosterone. The first 2 min consisted of the experimenter holding the bird and slowly walking (1 step/s) along a non-experimental aisle in the shed, and the final 2 min consisted of sitting quietly with the bird next to the blood-sampling area. A 4–5 mL blood sample was then withdrawn via venipuncture of the wing vein, and the hen was returned to its home cage. The blood samples were stored on ice until they could be centrifuged later that day. The plasma fraction was removed, frozen, and stored at –20°C. The plasma was later thawed and analysed by a third party, using a competitive enzyme immunoassay (EIA) kit from IDS Ltd (Fountain Hills, AZ, USA).
Statistical analyses
Behavioural assessment
All data were checked for normality before analysis. A univariate ANOVA found significant (P < 0.01) differences among sheds for both behavioural variables (SumZ and PropHO) on each testing day and shed was included as a factor in further analyses. A repeated-measures analysis was conducted using test day, shed, duration and proximity as factors to examine the effects of the human-contact treatments on bird behaviour. The pre-treatment data collected on Day –5 for the AHT were significantly correlated with the data collected on Days 15 and 30, and were thus used as a covariate. Comparisons between pairs of means were tested using post hoc l.s.d. tests (main effects) and post hoc Scheffe tests (interaction effects).
Physiological assessment
All data were checked for normality before analysis. A repeated-measures analysis was conducted to examine the effects of the human-contact treatments on the plasma corticosterone response to handling of the hens. While the majority of samples taken on Day 30 would have been from birds different from those sampled on Day –5, a repeated-measures analysis was considered appropriate because it was the cage, rather than individual birds, that was the experimental unit. Comparisons between pairs of means were tested using post hoc l.s.d. tests (main effects) and post hoc Scheffe tests (interaction effects).
Results
Approaching-human test
There was a significant (F = 3.56, P = 0.03) effect of proximity on hen behaviour, with birds in the 0 m treatments showing significantly (P < 0.05) less avoidance than those in the 1.50 m treatments (Table 2).

|
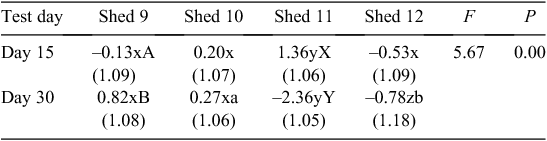
There was also a significant (F = 5.67, P = 0.00) test day × shed interaction, presented in Table 3. The birds in Shed 11 showed a significant (P < 0.01) increase in avoidance behaviour from Day 15 to Day 30. The birds in Shed 11 showed the least avoidance behaviour of birds in all sheds on Day 15 (P < 0.01), but the most avoidance behaviour of birds in all sheds on Day 30 (P < 0.01). The birds in Shed 9 showed a significant (P < 0.05) decrease in avoidance behaviour from Day 15 to Day 30.
|
There were no other significant (P < 0.05) interactions or main effects for duration, proximity, shed or test day for this behavioural variable (SumZ).
Stroll test
There was a significant (F = 4.00, P = 0.01) effect of shed on hen behaviour, with birds in Shed 12 displaying significantly more avoidance behaviour than birds in Shed 10 (P < 0.01) and Shed 11 (P < 0.05) (Table 2).
No other significant (P < 0.05) main effects or interactions for duration, proximity, shed or test day were found for the proportion of heads out during the stroll test (Prop HO) (Table 2).
Corticosterone concentration
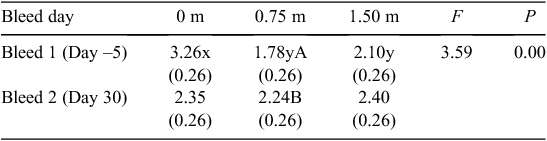
The results of the repeated-measures analysis for the main effects on the plasma corticosterone (ng/mL) response to handling are presented in Table 4. Significant (F = 6.25, P = 0.00) differences were detected for the proximity treatments, with birds in the 0 m treatment having a greater plasma corticosterone response to handling than birds in the 0.75 m treatment (P < 0.01). There was no significant main effect of the duration or bleed day; however, a significant (F = 3.59, P = 0.04) bleed day × proximity interaction occurred (Table 5). The plasma corticosterone response to handling for birds in the 0 m treatment group was significantly (P < 0.01) higher than that in the other groups on Day –5, before the treatments commencing. By Day 30, the corticosterone response for birds in the 0 m group had dropped to similar levels as that for the birds in the 0.75 m and 1.50 m groups, and was not significantly different. In addition, the birds in the 0.75 m treatment group showed a significant (P < 0.05) increase in plasma corticosterone response. There were no significant changes in the plasma corticosterone response for birds in the 1.5 m treatment (P > 0.05).

|
|
Discussion
Fear of humans was significantly reduced for birds receiving visual human contact at 0 m. Birds that had received this treatment remained at the cage front more often when approached by the researcher during the AHT, but not during the stroll test. These results agree with the findings of Edwards et al. (2010), who found that visual human contact at a distance of less than 0.75 m from the cage front was more effective in reducing avoidance behaviour in caged hens than was contact at 1.50 m. However, it was unexpected that the human contact at a medium proximity (0.75 m) did not reduce avoidance behaviour in the present experiment. A possible explanation for this may relate to the visibility of the researcher’s eyes, as this has been previously reported to increase fear and avoidance behaviour in domestic fowl (Jones 1980; Rosa Salva et al. 2007; Scaife 1976a, 1976b). The researcher’s eyes were clearly visible to the birds in the 0.75 m and 1.50 m treatments, but were visible only to some of the birds in the 0 m treatment. This occurred because the researcher was standing with her torso against the feed trough at the front of the cage. If the hens were standing at the back of the cage, the manure belt above them blocked their view of the researcher’s face, leaving only a clear view of the researcher’s body. Birds at the front of the cage were still able to look up and make eye contact; however, an important element of the situation was that they were also able to move out of eye contact if they wished. The birds receiving the 0.75 m and 1.50 m treatments were unable to escape the gaze of the researcher, and may have perceived the treatments as more threatening than the birds in the 0 m treatments. This may have reduced their habituation to the researcher in comparison to the birds receiving the 0 m treatments, and so displayed greater avoidance behaviour during the AHT.
The human-contact treatments did not alter the degree of avoidance behaviour displayed by the birds during the stroll test. It is possible that the human-contact treatments were better suited to habituating the birds to close human presence and approach, and this habituation did not generalise to a human behaving in a novel manner, such as slowly walking past the cages holding a video camera.
The plasma corticosterone response to gentle handling increased for the birds receiving the 0.75 m treatments, although this increase was not associated with a concurrent increase in avoidance behaviour displayed by the birds in this treatment. As discussed above, the visibility of the researcher’s eyes to birds in the 0.75 m and 1.50 m treatments may have influenced how threatening the birds perceived the treatments to be. Potentially, birds receiving the human-contact treatments at the intermediate proximity (0.75 m) found human presence to be more threatening than did the birds receiving human-contact treatments at 1.50 m, hence an increased corticosterone response for birds in the 0.75 m treatment group. However, this hypothesis cannot be verified, particularly as the birds in the 0.75 m treatment group did not show any changes in avoidance behaviour during testing. It is possible that visual human contact at 0.75 m was not sufficient to alter the avoidance response, but may have delayed habituation rates.
The plasma corticosterone response to gentle handling was significantly reduced for birds receiving the 0 m treatments. However, the birds in the 0 m treatments started the experiment with significantly elevated corticosterone responses (Day –5), which then dropped to the same levels as the birds in the other treatments by the conclusion of the experiment. These pre-treatment differences in corticosterone response are an anomaly, as the birds were allocated to the treatments at random. Therefore, it is not clear whether the significant drop in corticosterone response seen in the 0 m birds on Day 30 was due to the effect of the proximity treatments or simply due to the birds ‘returning to normal’. Thus, while it is tempting to attribute this decrease in corticosterone concentrations to a reduction in fear of humans, this relationship cannot be attributed to the human-contact treatments. The high values for the pre-treatment corticosterone data in the 0 m treatment group would also explain why this group had significantly higher overall corticosterone concentration than did the birds in the 0.75 m treatment.
It is possible that a longer period of manual restraint before blood sampling, or a more comprehensive blood sampling schedule, may have yielded different results; however, the practicalities of implementing these changes on farm, combined with the negative effect of blood sampling on the human–animal relationship, means that these options were not investigated. In addition, some birds may have been sampled on both bleed days (Day –5 and Day 30), which may have resulted in an increased corticosterone response in these birds. This potential effect of previous experience may have masked subtle treatment effects and, in hindsight, this aspect of the blood-sampling procedure should have been standardised.
The lack of an effect of the duration treatment was unexpected, as the degree of habituation that occurred would be expected to increase as the duration of exposure to the stimulus increased. Observational research on caged egg farms has found that increased time spent by stockpeople standing stationary close to the cages was associated with less avoidance behaviour to humans in the hens (Edwards 2009). The reason for this lack of effect is unable to be explained; however, the duration of contact with other humans during routine husbandry procedures in the sheds was not quantified, and it is possible that these additional human–animal interactions may have influenced the response of the birds.
Significant differences in avoidance behaviour were recorded among the four sheds used in the present study. The birds in Shed 12 showed a roughly linear reduction in fear during the course of the experiment for both measures of behaviour, while the other sheds displayed large variation in their responses. It is possible that the birds in Shed 12 were responding to the treatments in a manner different from that of the other sheds. The birds in Shed 12 were also significantly more fearful on the basis of the results of the stroll test. Similarly, the birds in Shed 10 were consistently less fearful on the basis of both measures of behaviour throughout the experiment. This was demonstrated particularly well by the stroll test, in which the order of sheds from most fearful to least fearful was Shed 12, Shed 9, Shed 11 and Shed 10. This shed order also represents the ages of the birds, from the oldest in Shed 12 to the youngest in Shed 10. It is possible that the age of the birds may have been influencing the way that the birds responded to the human-contact treatments. This is in accordance with the results of a study by Jones (1995), who found that non-handled chicks became more fearful of humans as they aged. The sheds were also maintained by different stockpeople, perhaps leading to variation in avoidance behaviour due to variation in the human–animal relationship. For example, the stockperson in Shed 12 was observed tapping the cages to move the birds during the daily inspections. In addition, the birds in Shed 12 were housed in larger groups per cage than those in Sheds 9, 10 and 11. Large group size per cage has been significantly correlated with increased avoidance behaviour in commercial caged laying hens (Edwards 2009), and group size may have been influencing the response of the birds in the present study. Thus, it would appear that the differences among sheds may be due to several factors, such as the age of the birds, variation in human contact and the housing conditions of the shed. These factors may have altered the rate at which the hens in different sheds habituated to the researcher, particularly as the experiment was conducted over a relatively short period (30 days).
The close-proximity (0 m) treatment was the only human-contact treatment to reduce avoidance behaviour in commercial laying hens. While it is impractical to regularly impose this type of human contact in a commercial situation, it may be useful for stockpeople to be aware of their proximity to the hens while carrying out their work and moving through the shed. For example, they may consider walking along one side of the aisle rather than walking down the middle so that they are closer to the birds. It should also be recognised that care was taken in this experiment to impose all human-contact treatments in a slow and predictable manner; stockpeople must ensure that their interactions with the birds are also conducted in a slow and predictable manner, preferably avoiding long periods of eye contact, otherwise their close presence to the birds may become threatening.
In conclusion, visual contact with a stationary human at close proximity (0 m) significantly reduced avoidance behaviour in commercial laying hens when approached by a human. The duration of human contact had no effect on avoidance behaviour or plasma corticosterone response. The results of this experiment suggest that regularly exposing caged laying hens to non-threatening human contact at close proximity may improve the human–animal relationship on commercial egg farms. Further research is required to explore the specific mechanisms involved in the observed effect of close human proximity, such as the effect of eye contact, and whether the treatments are similarly effective for different tiers of cages.
Acknowledgements
The authors thank the Australian Poultry Cooperative Research Centre for financial support, the staff and students at the Animal Welfare Science Centre for assistance with data collection and analysis, and Kinross Farm for participating in this research.
References
Barnett JL, Hemsworth PH, Hennessy DP, McCallum TH, Newman EA (1994) The effects of modifying the amount of human contact on behavioural, physiological and production responses of laying hens. Applied Animal Behaviour Science 41, 87–100.| The effects of modifying the amount of human contact on behavioural, physiological and production responses of laying hens.Crossref | GoogleScholarGoogle Scholar |
Beuving G, Vonder GMA (1977) Daily rhythm of corticosterone in laying hens and the influence of egg laying. Journal of Reproductive Sciences 51, 169–173.
| Daily rhythm of corticosterone in laying hens and the influence of egg laying.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE1cXjtlCr&md5=2f0cfa198eb0c4f4d13c13c365a0e399CAS |
Beuving G, Vonder GMA (1978) Effects of stressing factors on corticosterone levels in the plasma of laying hens. General and Comparative Endocrinology 35, 153–159.
| Effects of stressing factors on corticosterone levels in the plasma of laying hens.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE1cXkvVOmsLo%3D&md5=6581e9d44c86ec8b758600938f906df5CAS | 208921PubMed |
Edwards LE (2009) The human–animal relationship in the caged laying hen. PhD Thesis, University of Melbourne, Australia.
Edwards LE, Botheras NA, Coleman GJ, Hemsworth PH (2010) Behavioural and physiological responses of laying hens to humans. Animal Production Science 50, 557–559.
| Behavioural and physiological responses of laying hens to humans.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXnt1yrs7s%3D&md5=2b076f739fd576f240ea9e81e51bc34cCAS |
Graml C, Niebuhr K, Waiblinger S (2008) Reaction of laying hens to humans in the home or a novel environment. Applied Animal Behaviour Science 113, 98–109.
| Reaction of laying hens to humans in the home or a novel environment.Crossref | GoogleScholarGoogle Scholar |
Gross WB, Siegel PB (1982) Socialization as a factor in resistance to infection, feed efficiency, and response to antigen in chickens. American Journal of Veterinary Research 43, 2010–2012.
Hemsworth PH, Barnett JL (1989) Relationships between fear of humans, productivity and cage position of laying hens. British Poultry Science 30, 505–518.
| Relationships between fear of humans, productivity and cage position of laying hens.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK3c%2FlvVWlsg%3D%3D&md5=2d17eb519ccf33e1cfceb8bd8fc5c905CAS | 2819495PubMed |
Hemsworth PH, Coleman GJ (2011) ‘Human–livestock interactions. The stockperson and the productivity and welfare of intensively farmed animals.’ (CAB International: Wallingford, UK)
Hemsworth PH, Coleman GJ, Barnett JL, Jones RB (1994) Behavioural responses to humans and the productivity of commercial broiler chickens. Applied Animal Behaviour Science 41, 101–114.
| Behavioural responses to humans and the productivity of commercial broiler chickens.Crossref | GoogleScholarGoogle Scholar |
Hogan JA (2008) Motivation. In ‘The behaviour of animals: mechanisms, function and evolution’. (Eds JJ Bolhuis, L-C Giraldeau) pp. 41–70. (Blackwell Publishing: Malden, MA)
Janczak AM (2010) Fear. In ‘The encyclopaedia of applied animal behaviour and welfare’. 1st edn. (Eds DM Mills, JN Marchant-Forde, DB Morton, CJC Phillips, PD McGreevy, CJ Nicol, P Sandoe, RR Swaisgood) pp. 251–254. (CABI Publishing: Wallingford, UK)
Jones RB (1980) Reactions of male domestic chicks to two-dimensional eye-like shapes. Animal Behaviour 28, 212–218.
| Reactions of male domestic chicks to two-dimensional eye-like shapes.Crossref | GoogleScholarGoogle Scholar |
Jones RB (1993) Reduction of the domestic chick’s fear of human beings by regular handling and related treatments. Animal Behaviour 46, 991–998.
| Reduction of the domestic chick’s fear of human beings by regular handling and related treatments.Crossref | GoogleScholarGoogle Scholar |
Jones RB (1995) Ontogeny of response to humans in handled and non-handled female domestic chicks. Applied Animal Behaviour Science 42, 261–269.
| Ontogeny of response to humans in handled and non-handled female domestic chicks.Crossref | GoogleScholarGoogle Scholar |
Jones RB, Hughes BO (1981) Effects of regular handling on growth in male and female chicks of broiler and layer strains. British Poultry Science 22, 461–465.
| Effects of regular handling on growth in male and female chicks of broiler and layer strains.Crossref | GoogleScholarGoogle Scholar |
Keeling L (1995) Spacing behaviour and an ethological approach to assessing optimum space allocations for groups of laying hens. Applied Animal Behaviour Science 44, 171–186.
| Spacing behaviour and an ethological approach to assessing optimum space allocations for groups of laying hens.Crossref | GoogleScholarGoogle Scholar |
Mankovich NJ, Banks EM (1982) An analysis of social orientation and the use of space in a flock of domestic fowl. Applied Animal Ethology 9, 177–193.
| An analysis of social orientation and the use of space in a flock of domestic fowl.Crossref | GoogleScholarGoogle Scholar |
McFarland D (1981) ‘The Oxford companion to animal behaviour.’ (Oxford University Press: Oxford, UK)
Murphy LB, Duncan IJH (1978) Attempts to modify the responses of domestic fowl towards human beings. II. The effect of early experience. Applied Animal Ethology 4, 5–12.
| Attempts to modify the responses of domestic fowl towards human beings. II. The effect of early experience.Crossref | GoogleScholarGoogle Scholar |
Rosa Salva O, Regolin L, Vallortigara G (2007) Chicks discriminate human gaze with their right hemisphere. Behavioural Brain Research 177, 15–21.
| Chicks discriminate human gaze with their right hemisphere.Crossref | GoogleScholarGoogle Scholar | 17174412PubMed |
Scaife M (1976a) The response to eye-like shapes by birds. I. The effect of context: a predator and a strange bird. Animal Behaviour 24, 195–199.
| The response to eye-like shapes by birds. I. The effect of context: a predator and a strange bird.Crossref | GoogleScholarGoogle Scholar |
Scaife M (1976b) The response to eye-like shapes by birds II. The importance of staring, pairedness and shape. Animal Behaviour 24, 200–206.
| The response to eye-like shapes by birds II. The importance of staring, pairedness and shape.Crossref | GoogleScholarGoogle Scholar |
Zulkifli I, Siti Nor Azah A (2004) Fear and stress reactions, and the performance of commercial broiler chickens subjected to regular pleasant and unpleasant contacts with human beings. Applied Animal Behaviour Science 88, 77–87.
| Fear and stress reactions, and the performance of commercial broiler chickens subjected to regular pleasant and unpleasant contacts with human beings.Crossref | GoogleScholarGoogle Scholar |
Zulkifli I, Gilbert J, Liew PK, Ginsos J (2002) The effects of regular visual contact with human beings on fear, stress, antibody and growth responses in broiler chickens. Applied Animal Behaviour Science 79, 103–112.
| The effects of regular visual contact with human beings on fear, stress, antibody and growth responses in broiler chickens.Crossref | GoogleScholarGoogle Scholar |