

Grazing systems and worm control in sheep: a long-term case study involving three management systems with analysis of factors influencing faecal worm egg count
S. W. Walkden-Brown A F , A. F. Colvin A B , E. Hall C , M. R. Knox D , D. F. Mackay E and J. M. Scott AA School of Environmental and Rural Science, University of New England, Armidale, NSW 2351, Australia.
B Current address: ‘Pine Grove’, 6248 New England Highway, Ben Lomond, NSW 2365, Australia.
C 111 Margaret Street, Launceston, Tas. 7250, Australia.
D CSIRO Livestock Industries, Armidale, NSW 2350, Australia.
E 3 Jayne Close, Armidale, NSW 2350, Australia.
F Corresponding author. Email: swalkden@une.edu.au
Animal Production Science 53(8) 765-779 https://doi.org/10.1071/AN13037
Submitted: 30 January 2013 Accepted: 25 March 2013 Published: 10 July 2013
Journal Compilation © CSIRO Publishing 2013 Open Access CC BY-NC-ND
Abstract
Managing infections of sheep with gastrointestinal nematode parasites (worms) and problems of resistance to anthelmintic treatments continue to be major challenges for graziers on the Northern Tablelands of New South Wales, Australia. The whole-farmlet study of grazing enterprises undertaken by the Cicerone Project tested the broad hypotheses that compared with typical management (farmlet B), internal parasites can be more effectively managed with improved nutrition (farmlet A) or by intensive rotational grazing (farmlet C). Further aims were to identify the major sources of variation in faecal worm egg count (WEC) over the 6-year period and to examine the efficacy of the various anthelmintic treatments used during the experiment. This paper describes the management of sheep worms at the whole-farmlet level during the experiment, and analyses data from the routine WEC monitoring (5644 records) and larval differentiation tests (322 records) carried out on behalf of the Cicerone Management Board and by a doctoral candidate. It complements more detailed investigations published elsewhere.
Over the period from July 2000 to December 2006, worm infections in ewes, lambs, hoggets and wethers were, with some exceptions, successfully controlled on the farmlets through a combination of regular monitoring of WEC, treatment with a wide array of anthelmintics and grazing management. Farmlet C had lower mean WEC (444 epg) and annual anthelmintic treatment frequency (3.1 treatments/year) over the whole experimental period than farmlets B (1122 epg, 4.3 treatments/year) or A (1374 epg, 4.7 treatments/year). The main factors influencing WEC were the time since the last anthelmintic treatment, and the anthelmintic used at that treatment. The magnitude of these effects dwarfed those of climatic and management factors that might be expected to influence the epidemiology of gastrointestinal nematode infections via environmental or host-mediated mechanisms. Nevertheless management factors associated with stocking rate and grazed proportion (proportion of each farmlet grazed at any one time), and climatic indicators of both temperature and moisture availability had significant effects on WEC.
The results show that, in a region with Haemonchus contortus as the major sheep nematode, improved host nutrition in a higher input system (farmlet A) did not provide more effective control of gastrointestinal nematodes than typical management (farmlet B); however, it was observed that gastrointestinal nematode control was no worse on farmlet A than on farmlet B in spite of farmlet A supporting a 48% higher stocking rate by later in the trial period (2005). The study provided strong support for the proposition that intensive rotational grazing (farmlet C) provides more effective control of gastrointestinal nematodes than typical management (farmlet B) as evidenced by significantly lower WEC counts and anthelmintic treatment frequency. Tactical worm control based on routine monitoring of WEC provided adequate control of worms on all three farmlets for much of the experimental period but failed to prevent significant spikes in WEC to values associated with significant production loss on multiple occasions, and significant ewe mortality on farmlets A and B on one occasion.
Additional keywords: anthelmintic, Haemonchus, integrated parasite management, nutrition, resistance, rotational grazing, Trichostrongylus.
Introduction
Gastrointestinal nematode (roundworm) parasites of sheep are an ongoing and important problem for graziers on the Northern Tablelands of New South Wales (NSW), Australia. Resistance to anthelmintic drenches is a major and widespread problem that is getting worse (Love 2012). Gastrointestinal worm infection is the most economically important disease of sheep in Australia (McLeod 1995; Sackett et al. 2006) and the impact has recently been described in detail for the Northern Tablelands of NSW by Kelly et al. (2010). The major worm species in this region are the abomasal Haemonchus contortus (Barber’s pole worm) and the small intestinal Trichostrongylus colubriformis (Black scour worm) with some presence of other species including the abomasal Teladorsagia circumcincta (small brown stomach worm). The former induces significant blood loss with the major impact being elevated mortality. The latter two species are ‘scour worms’, which are associated with lower reproductive rates and a more chronic course of infection leading to reduced production as the major economic impact. Severe infections of any of these worm species can result in animal deaths and these occur most commonly under moist summer conditions (Gordon and Whitten 1941; Roe et al. 1959b).
As part of the producer-led Cicerone Project (Sutherland et al. 2013), a survey of the research needs of livestock producers in this region was undertaken by Kaine et al. (2013). This revealed considerable concern about control strategies for sheep intestinal worms and the potential for drench resistance as well as interest in alternative control strategies including that of intensive rotational grazing (IRG). Even though short-term rotational grazing, using weekly rotation across four paddocks, was found not to affect worm burdens (Roe et al. 1959b), there has been anecdotal evidence of low drenching requirements on some properties practising more IRG or ‘cell grazing’ with shorter graze and longer rest periods. Producer members of the Cicerone Project were thus keen to compare grazing strategies that used short graze periods and long rest periods with more typical management strategies. A decision was taken by the Cicerone Board to explore several questions, including gastrointestinal worm control, within three whole farmlets that would test strategies at a scale viewed as credible by livestock producers.
Thus, three 53-ha farmlets were established to allow the effects of management differences to be quantified over time. One farmlet (B) was managed as a typical, or control, system based on moderate levels of pasture inputs and soil fertility with eight paddocks and flexible rotational grazing. A second farmlet (A) was managed to have a high level of pasture renovation with a higher level of soil fertility but with the same number of paddocks and grazing management as farmlet B. The third farmlet (C) was subdivided into 17 paddocks and, after a year, into a total of 37 subpaddocks to allow for IRG while receiving the same level of inputs as farmlet B. The management of worms on the farmlets was essentially tactical, based upon regular (approximately monthly) worm faecal egg count (WEC) monitoring of different mobs on the different farmlets. Worm control interventions were made on the advice of an expert veterinary consultant who also sat on the Cicerone Board of Management.
Some of the results of detailed parasitological studies on the Cicerone farmlet experiment have already been published. Healey et al. (2004) examined the WEC data for the first 3 years of the experiment and reported that clear farmlet differences emerged in 2003, the third year of the experiment, with the main finding being reduced WEC and frequency of treatment on farmlet C. Colvin et al. (2008), using monthly samples from lambs, hoggets and ewes over 2 years (2004–05) on each farmlet, demonstrated that IRG of farmlet C can indeed assist in the control of gastrointestinal nematodosis, especially where H. contortus is the most common parasite. A companion paper (Colvin et al. 2012) reported that the observed effects of IRG were due to disruption of the parasite lifecycle outside of the host, rather than differences in host immunity. Indeed, sheep on farmlet C were significantly more susceptible to worms than those on farmlets A or B during spring and summer.
As a complement to these detailed studies, this paper provides a longer-term overview of the management of internal worm parasites across mobs of various ages on the three farmlets over the entire 6.5 years of the farmlet experiment. Its main aim is to test two broad hypotheses that producer members of the Cicerone Project wanted investigated at a ‘credible’ scale. These were that compared with typical management (farmlet B), internal parasites can be more effectively managed with (1) improved nutrition (farmlet A) or (2) by IRG (farmlet C). Further aims were to identify the major sources of variation in WEC over the experimental period and to examine the efficacy of the various anthelmintic treatments used.
Materials and methods
Experimental design and location
The background to the Cicerone Project and its design are detailed by Sutherland et al. (2013) and Colvin et al. (2008). Briefly the farmlet experiment was located on the CSIRO property, ‘Chiswick’, 18 km south of Armidale, NSW, Australia (950 m altitude 3031′S, 15139′E). Armidale has a cool, temperate climate with the majority of rain falling in the summer months. Winters are generally dry, cold and frosty with occasional light snowfall. Summer days are warm but nights are generally cool. The average annual rainfall is ~790 mm. The weather experienced during the trial period was mostly drier than average with more severe frost in winter than average (Behrendt et al. 2013). The farmlet experiment commenced on 1 July 2000 following the subdivision of the three farmlets, each of 53 ha, which were managed under three different management systems described in detail by Scott et al. (2013b). Stock was first introduced just before the start date and the experiment was completed by the end of 2006.
This paper encompasses a longitudinal study of WEC between January 2001 and May 2006 under the three management systems described in the Introduction. The key factors in the design were:
-
Three farmlets each representing a different management system: farmlet A (higher inputs), farmlet B (typical) and farmlet C (IRG).
-
Four classes of sheep: lambs (0–12 months), hoggets (~12–24 months), mature ewes and wethers.
-
Year and season. Years were calendar years and seasons were spring (Sept.–Nov.), summer (Dec.–Feb.), autumn (March–May) and winter (June–Aug.).
Sheep, management and worm control
All sheep were fine wool Merinos. Ewes were generally mated for 5 weeks in mid–late autumn to lamb in spring. Lamb marking was generally in mid November and weaning from late December to January. Shearing of all sheep took place at the end of July each year in an off-site shearing shed. Managing worm burdens was largely tactical, based upon WEC testing at 1–2-month intervals of the various sheep classes on the different farmlets, followed by larval culture and differentiation to species or genus level is a common practice in this region. In addition, two fixed treatments were given each year: the first was a quarantine treatment given to all sheep as they moved off the property for shearing in July and the second was a pre-mating treatment for all ewes in March–April before being grouped together in a paddock external to the experimental farmlets over the joining period, to ensure equivalent genetics between farmlets. Interpretation of the WEC and larval differentiation data and decisions about treatment and the anthelmintic to be used were made by a Veterinary consultant (E. Hall) in consultation with the Cicerone Board of Management where necessary. A wide range of anthelmintic treatments was used as summarised in Table 1; all were used according to the manufacturers’ recommendations.
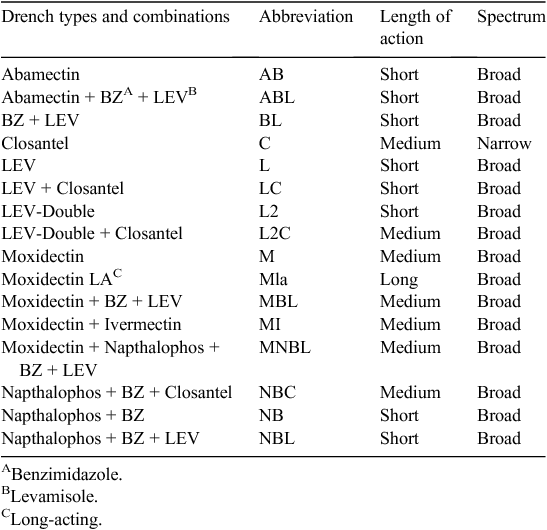
|
Measurements
Individual faecal samples were either collected per rectum (May 2002–November 2003) or collected from each mob by mustering the mob into a paddock corner and waiting for defecation followed by collection of fresh samples from the ground. Ten individual faecal samples, considered adequate by Morgan et al. (2005), were sent to one of three laboratories (Laboratory A, August 2000–May 2002; Laboratory B, May 2002–November 2003 and after October 2005; Laboratory C, November 2003–October 2005) for assessment of WEC using a modified McMaster method (Whitlock 1948). The remaining faeces were pooled, cultured for 7 days followed by L3 recovery and differentiated into species (H. contortus, Teladorsagia circumcincta) or genus (Trichostrongylus, Oesophogostomum or Cooperia) in the same laboratory. The total number of WEC records was 5644 and the number of larval differentiations was 322.
A faecal egg count reduction test (FECRT) was carried out on farmlet C lambs in May 2004 to determine the effectiveness of four anthelmintics against H. contortus. Due to the small number of lambs available, only four treatment groups plus a control group were used. The control lambs were sampled at the beginning and the end of the experiment to determine the change of WEC over time and FECRT was calculated using the method of Kemper and Walkden-Brown (2004). The FECRT commenced when the WEC of the lambs reached a threshold of 400 eggs/g testing the following anthelmintics: albendazole (3.8 mg/kg Alben, Virbac, Australia), levamisole (8.0 mg/kg, Levamisole Gold, Virbac, Australa), ivermectin (0.2 mg/kg Ivomec Oral Sheep Liquid, Merial, Australia), closantel (7.5 mg/kg, Closicare, Virbac, Australiag) at full dose rate. On Day 0, 75 lambs were randomly allocated to five groups of 15, all lambs were sampled for individual faecal WEC and pooled group larval cultures, bodyweights were recorded and each lamb dosed using a stomach tube, with the correct dose of anthelmintic, or 30 mL of water for the control group. On Day 13 individual WEC and group cultures were carried out and faecal WEC reduction calculated for H. contortus as this was the only nematode for which WEC was sufficient to enable accurate determination of FECR. The presence of resistance to an anthelmintic was assumed if (i) the percentage reduction in WEC was less than 95% and (ii) the lower 95% confidence interval was less than 90%. Emerging resistance was suspected when only one of these criteria was met (Coles et al. 1992).
Rainfall and temperature data were accessed from the nearest Bureau of Meteorology station (Armidale) some 17 km north of the experimental site. As described in a related paper by Behrendt et al. (2013), the climate experienced over the experimental period was in general drier than average, especially over the autumn–spring period, and experienced more severe frosts than average. Climatic data over the long-term from 1890 to 2008 were used as inputs to the model Ausfarm, which permitted the estimation of changes in available soil water (ASW) over both the long-term and the experimental period (Behrendt et al. 2013). The same climatic data were used to estimate the potential effects of temperature on gastrointestinal nematodes and pasture growth as the average of growing-degree days (GDD) calculated using base temperatures of 4°C for grasses and 10°C for clovers [((max. temp + min. temp)/2) – (base temp)] (Hutchinson et al. 2000).
Statistical methods
WEC data were not normally distributed and so were treated in two ways. To illustrate general patterns and trends, untransformed means are presented as this is the format in which the farming and advisory community receive and have to interpret such data. In addition to this descriptive presentation, formal analyses of WEC were carried out to explore the sources of variation in this variable and their contribution to total variation. For these analyses WEC was transformed [Log10 (WEC + 20)] (LogWEC) before fitting least-squares models as this transformation best stabilised the variances and distribution of the residuals. WEC data were coded by a range of fixed and variable effects as shown in Table 2.

|
For this analysis all data from wethers and the years 2000 and 2006 were excluded as they were sparse and not well balanced across the other fixed effects. This reduced the number of WEC samples analysed from 5644 to 5183. The importance of various factors in the WEC models was evaluated by their contribution to the R2 value for the model; that is, their contribution to total sums of squared deviations from the mean (SSQ).
Larval differentiation data (% of each genus) are presented without formal analysis. Annual drench frequency was calculated within classes, farmlets and years on the reduced dataset and analysed by fitting these effects and their two-way interactions in a least-squares model.
All analyses were performed using JMP 10 statistical software (SAS Institute, Cary, NC, USA) and a significance level of P < 0.05 has been used throughout.
Results
Mean length of graze and rest periods
The relative lengths of graze and rest periods on each farmlet over each year of the trial are shown in Table 3 as averages over all major paddocks and subpaddocks on each farmlet. The mean number of graze days were lower on farmlet A than on farmlet B, due presumably to the higher level of green digestible herbage (Shakhane et al. 2013) and stocking rate (SR, Hinch et al. 2013a) on this farmlet with both of these farmlets having substantially longer graze periods than on farmlet C. The lengths of the grazing rest periods were similar on both farmlets A and B but were some 50% longer on farmlet C. It should be noted that, as grazing on farmlet C was usually within ‘cells’, which were smaller than subpaddocks, the actual days of grazing on this farmlet were lower and the days of rest higher than indicated in Table 3.
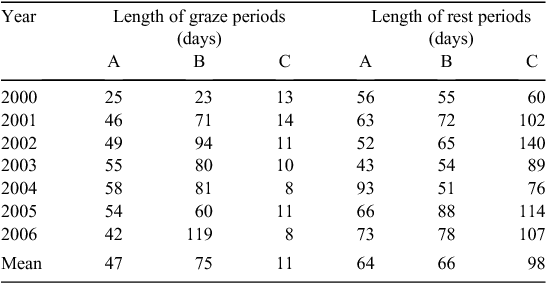
|
Mean percentage of farmlets grazed
Fig. 1 shows the mean percentage of each farmlet grazed together with the daily rainfall over the experimental period. After the grazing treatments were implemented in late 2000, the percentage of farmlets A and B grazed at any one time varied from 20 to 40% whereas that of farmlet C varied from ~5 to 10%.
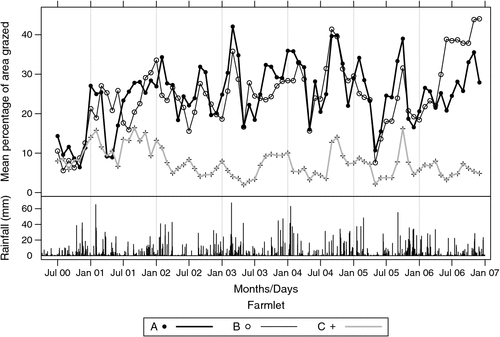
|
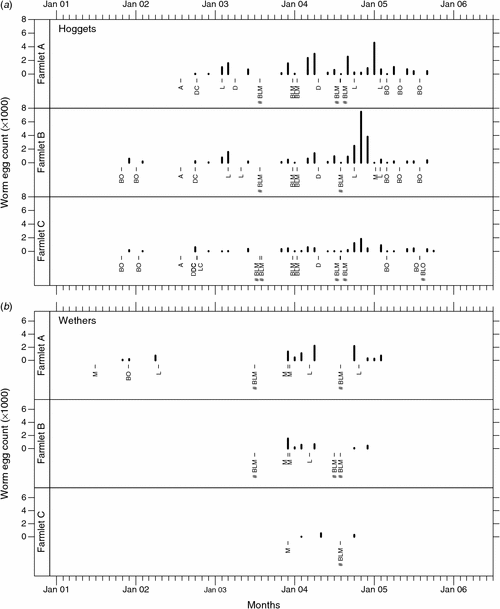
Mean faecal WEC within sheep classes and farmlets over time
Untransformed mean WEC by farmlet over the experimental period are shown in Fig. 2 (ewes and lambs) and Fig. 3 (hoggets and wethers). These data show clearly that major spikes in WEC occurred on farmlets A and B in 2003, 2004 and 2005, but that such extreme fluctuations in WEC were not evident on farmlet C. WEC on farmlets A and B tended to be somewhat higher than those on farmlet C except in early 2006 when farmlet C ewes had the highest counts following a period of shorter rest days.
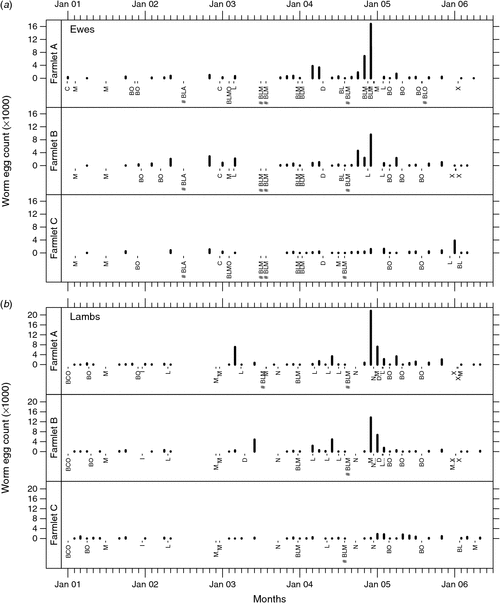
|
|
Overall raw mean WEC for farmlets A, B and C over the entire experimental period were 1374, 1122 and 444, respectively, for ewes; 1162, 910 and 419, respectively, for lambs; 974, 879 and 372, respectively, for hoggets; and 800, 585 and 291, respectively, for wethers. Over all years and classes the mean WEC for farmlets A, B and C were 1178, 985, 415, respectively. Over all years and farmlets, the mean WEC for ewes, lambs, hoggets and wethers were 988, 846, 735 and 666, respectively.
Nematode genera involved
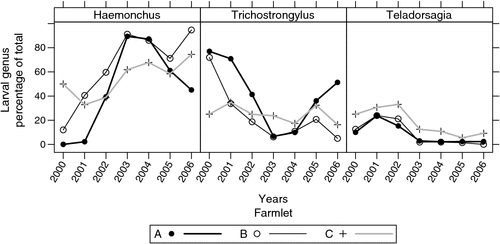
For the 322 recorded larval differential tests carried out during the experiment, the overall means for genus identification were 66.4% Haemonchus, 23.1% Trichostrongylus, 8.2% Teladorsagia, 2.1% Oesophagostomum and 0.2% Cooperia. These varied with time and farmlet as shown in Fig. 4. In the early stages of the experiment, Trichostrongylus was the dominant parasite, but it declined rapidly as Haemonchus increased and, by 2002, Haemonchus had become dominant. Farmlet C had a higher proportion of Teladorsagia than the other farmlets throughout the experiment and a lower proportion of Haemonchus and higher proportion of Trichostrongylus than farmlet B in all years apart from 2000 (Fig. 4). Farmlet B had a higher proportion of Haemonchus and lower proportion of Trichostrongylus than farmlet A in years other than 2003 and 2004 when they were similar. Proportions of Teladorsagia were similar in these two farmlets (Fig. 4).
|
Frequency of anthelmintic treatment
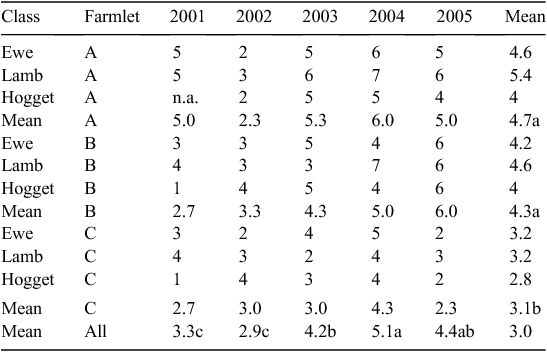
The number of anthelmintic treatments by year, sheep class and farmlet for the core dataset between 2001 and 2005 are summarised in Table 4. Overall lambs had significantly more annual treatments (4.4) than hoggets (3.4) with ewes having an intermediate level of treatment (4.0), with some year-to-year variation (Table 4).
|
Types of anthelmintic treatment used and WEC following treatment
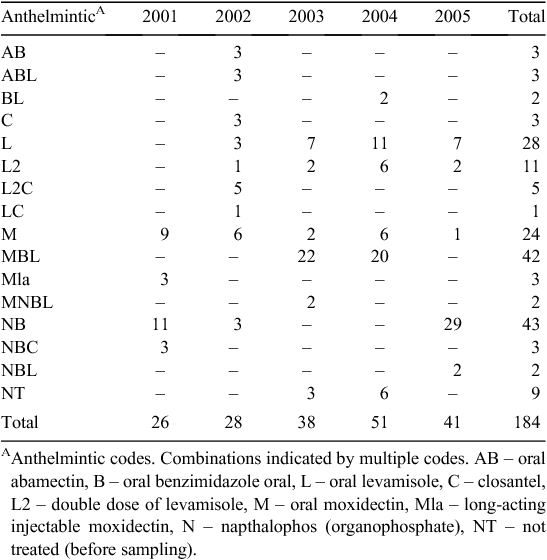
The types of anthelmintic used between 2001 and 2005 in ewe, lamb and hogget mobs and their frequency of use by year are shown in Table 5. The number of treatments is slightly higher than might be inferred from Table 4 because on a small number of occasions different mobs within a class of sheep on a farmlet were treated with different anthelmintics on the same day or week. These would represent a single treatment in Table 4 but two treatments in Table 5. The anthelmintics used varied over time with the broad spectrum combination drenches MBL and NB being the most commonly used, largely for the quarantine drenches, followed by the single active L and M.
|
Examination of WEC profiles following treatment with various anthelmintics failed to show widespread anthelmintic resistance, apart from closantel, for which significant Haemonchus WEC were often observed within the projected WEC-free period for this Haemonchus-specific anthelmintic (Fig. 5). At the commencement of the faecal WEC reduction test in 2003 on farmlet C mean WEC of the 75 lambs was 817 epg with larval differentiation of 92.6% Haemonchus, 3.4% Trichostrongylus and 4% Teladorsagia. Thus FECRT could only be calculated for Haemonchus. Levamisole was the only anthelmintic treatment that returned a 100% susceptibility of H. contortus (Table 6). Closantel, which was suspected to have reduced efficacy against this parasite showed efficacy of 96.8% at Day 13 after treatment. Closantel is a persistent anthelmintic with a recommended protection period against H. contortus of 42 days, thus emergence of resistance is suspected as there was breakthrough WEC only 13 days after treatment. Haemonchus contortus showed resistance to both ivermectin and albendazole.
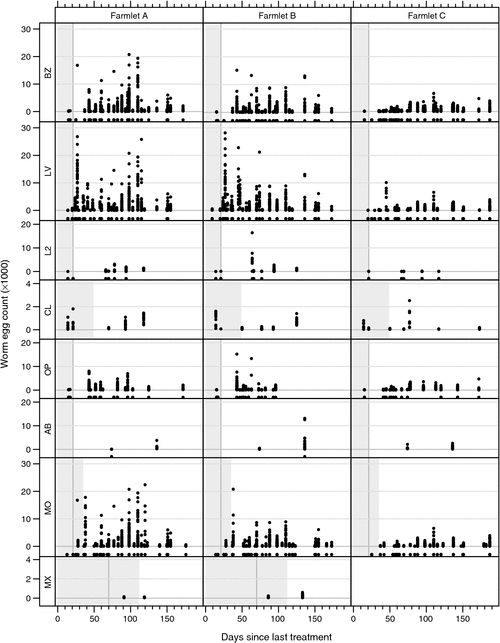
|

|
Sources of variation in WEC and their magnitude
A basic linear model fitting the effects of farmlet, class, year and season and their first-order interactions (Model 1) accounted for only 12.3% (R2) of the variation in LogWEC during the experiment with an Akaike information criterion (AIC) of 11 981. All effects and first-order interactions were statistically significant (P < 0.05), apart from the main effect of farmlet. Addition of the last anthelmintic treatment code (TC) to the model (Model 2) increased the R2 to 22.2% and further addition of the days since last anthelmintic treatment (DSLT) as a covariate (Model 3) increased the R2 to 34.3% with an AIC of 10 484. Interactions with these latter two effects were not fitted. In the last of these models all effects and first-order interactions were highly significant (P < 0.001) other than the interaction between farmlet and sheep class. The major sources of variation explained by the model were DSLT (18%), TC (11.5%), year (5.1%), class by year (3.5%), class by season (2.7%), season (1.2%) and farmlet by year (1.1%) with all other effects in the model accounting for less than 1% of the total sums of squares. The overall effect of year (P < 0.0001) was due to significantly higher LogWEC in 2004 (2.50) than all other years, and significantly higher LogWEC in 2005 (2.20) than 2001 (1.91), 2002 (1.75) and 2003 (1.81), which did not differ among themselves. The overall effect of class was due to significantly higher LogWEC in hoggets (2.23) than both lambs (1.96) or ewes (1.91). The overall effect of season was due to significantly (P < 0.0001) lower LogWEC in winter (1.82) than all of the three other seasons, spring (2.12), summer (2.10) or autumn (2.10). The overall effect of farmlet was due to significantly (P < 0.0001) lower LogWEC on farmlet C (1.89) than farmlets B (2.11) or A (2.10). However, there was significant interaction between the effects of farmlet and year (P < 0.0001) with the lower LogWEC on farmlet C only being evident in 2003, 2004 and 2005. Farmlet A also had significantly lower LogWEC than farmlet B in 2002 only.
To test whether actual measures of temperature, rainfall and evaporation would explain more of the variation than the broad category ‘season’, various combinations of actual and derived measures were used in place of season in Model 1. Replacing season with mean temperature (TMEAN) and sum of rainfall (RAIN) over the preceding 30 days increased the Model 1 R2 marginally from 12.3 to 13.1% (Model 4). Replacing TMEAN with mean minimum temperature (TMIN) did not alter the R2 (13.1%), but replacing it with the mean maximum temperature (TMAX) increased it slightly to 13.6%. Including both TMIN and TMAX increased the R2 to 20.4%. Replacing TMEAN with the mean of GDD over the preceding 30 days had little effect on the Model 4 R2, increasing it marginally to 13.3%. Thus, inclusion of both TMIN and TMAX in place of season provided the best fit of data with regard to temperature.
On the moisture side, replacing RAIN in Model 4 with mean evaporation rate over the preceding 30 days (EVAP) increased the R2 of Model 4 from 13.1 to 14.6%. Inclusion of both RAIN and EVAP increased it further to 19.1%, but replacing them with the Precipitation/Evaporation ratio over the preceding 30 days (P/E) resulted in a decline in R2 to 12.3%. Replacing RAIN with modelled available soil moisture for the 30 days before sampling (ASW) increased the R2 to 14.7%. Thus inclusion of both RAIN and EVAP in place of season provided the best fit of data with regard to moisture.
To test whether actual measures of SR and grazed proportion (GP) for the 30 days before sampling would explain more of the variation in LogWEC than the broad effect of ‘farmlet’, the effect of farmlet was replaced by combinations of these variables in Model 1. Replacement of ‘farmlet’ by SR and GP on their own had little effect on Model 1 R2 (R2 of 12.4 and 11.6%, respectively), but inclusion of both, increased the R2 to 15.0%.
A final model (Model 5) based on Model 3 but replacing farmlet with SR and GP, season with TMIN, TMAX, RAIN and EVAP had an R2 of 36.4% and AIC of 10 360. Thus only a marginal improvement in R2 (from 34.3 to 36.4%) was obtained by replacing the generic effects ‘Farmlet’ and ‘Season’ with measured variables that may have been expected to capture more relevant information about factors influencing WEC in sheep than these. In Model 5, all of the main effects were statistically significant with the exception of TMIN and RAIN but these, as with other main effects were involved in numerous significant two-way interactions. Removal of interactions reduced the R2 to 27.3%.
In Model 5 the main sources of variation were TC (6.6% of SSQ, P < 0.0001) and DSLT (6.9% of SSQ, P < 0.0001), with a strong positive association between DSLT and LogWEC. The effect of TC was difficult to interpret with levamisole, benzimidazoles and closantel in various combinations associated with both high and low LogWEC. The organophosphate napthalophos tended to be associated with lower WEC as did moxidectin, both oral and injectable. The overall effects of SR and GP were highly significant (P < 0.0001) and after the anthelmintic-associated effects, accounted for the largest proportion of variation among the main effects (0.82 and 0.92% of SSQ, respectively). In both cases the association with LogWEC was positive. Regression of the Model 5 predicted Log10 WEC against SR and GP and resulted in the following linear regression equations:
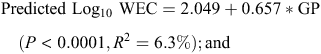

The overall effects of year and sheep class remained highly significant (P < 0.0001) accounting for 0.58 and 0.42% of SSQ, respectively. The effect of year was due largely to significantly lower LogWEC in 2003 than the other years, while the effect of class was due to hoggets having significantly higher WEC than ewes and lambs in this dataset. There was however a significant interaction between the effects of sheep class and year (P < 0.0001, 1.86% of SSQ) such that this effect of higher WEC in hoggets was only evident in 2001 and 2002.
The overall effects of TMAX (P = 0.04, 0.05% of SSQ) and EVAP (P = 0.01, 0.05% of SSQ) but not TMIN and RAIN on LogWEC were significant. For both TMAX and EVAP the association was positive.
Among the many significant first-order interactions it was notable that the climatic variables TMAX and EVAP had differential effects on sheep class with effects seen largely in ewes only (TMAX * Class P < 0.0001, 0.58% of SSQ; EVAP * Class P < 0.0001, 1.05% of SSQ). No other significant interactions accounted for more than 0.7% of SSQ.
Discussion
Overall, the results indicate that tactical worm control based on routine monitoring of WEC provided adequate control of worms on all three farmlets for much of the experimental period. However, this approach failed to prevent significant spikes in WEC to values associated with significant production loss on multiple occasions, and ewe mortality up to 5% on one occasion in December 2004, on farmlets A and B. Farmlet C clearly exhibited improved worm control relative to the other farmlets, as evidenced by both lower overall WEC values and fewer anthelmintic treatments required. However, this management system, as implemented, was associated with reduced individual animal productivity overall (Hinch et al. 2013a, 2013b) and by 2005, a SR some 10% lower than the typical farmlet B. The most important factors affecting WEC in the experiment were the time since the last anthelmintic treatment, and the anthelmintic used at that treatment. The magnitude of these effects dwarfed those climatic and management factors that might be expected to influence the epidemiology of gastrointestinal nematode infections via environmental or host-mediated mechanisms. Management factors associated with SR and GP, and climatic indicators of both temperature and moisture availability had significant effects on WEC, confirming that these factors are nevertheless still important.
The main objective of the study was to test whether, compared with typical management (farmlet B), internal parasites can be more effectively managed with improved nutrition (farmlet A) or by IRG (farmlet C). In spite of farmlet A supporting over time a higher SR than farmlet B (48% higher by 2005), there were few differences between farmlets A and B in WEC or the number of drenches applied. The only difference in LogWEC was in 2002 when farmlet A had significantly lower values than farmlet B. On the other hand, slightly more anthelmintic treatments per year were used on farmlet A (4.7) than farmlet B (4.3) although the difference was not statistically significant. The large peaks of WEC in late spring and summer of November and December 2004 on farmlets A and B were both associated with losses of ewes with lambs with acute haemonchosis (4.7 and 5% of ewes on each of these farmlets, respectively). These results suggest that, while intensification of sheep grazing systems in the Northern Tablelands region is unlikely to provide substantial benefits in parasite control, nevertheless the higher levels of green digestible herbage provided by farmlet A (Shakhane et al. 2013) enabled a substantially higher SR to be supported without incurring higher worm burdens than on farmlet B. On the other hand, farmlet C had significantly lower WEC and significantly fewer anthelmintic treatments per year than both of the other farmlets. Importantly, these effects were greatest when WEC and anthelmintic treatment numbers were highest, between 2003 and 2005, and this farmlet also did not have the haemonchosis-associated mortality observed in December 2004 on the other two farmlets. Working with a subset of the data from the present experiment, Colvin et al. (2008) also reported this beneficial effect of IRG on worm control and in a later report (Colvin et al. 2012) showed that the effect of IRG was mediated by reduced larval challenge on pastures rather than improved host resistance to nematode infection. Indeed sheep in the IRG treatment exhibited increased susceptibility to larval challenge in that study.
In the present study, the efficacy of IRG on farmlet C was associated with a shift in the relative abundance of the different parasite genera. As the experiment progressed (years 2002–06) there was a reduction in the proportion of Haemonchus in the larval cultures on farmlet C relative to farmlet B of 20.3% units (range 12.6–29.4% units), an increase in the proportion of Trichostrongylus by 10.6% units (range 6.2–17.5% units) and an increase in the proportion of Teladorsagia by 8.9% units (range 4.1–12.0% units). The Haemonchus and Teladorsagia larvae identified were almost certainly of the species H. contortus and T. circumcincta, while the Trichostrongylus larvae were most likely T. colubriformis, given the predominance of this species of Trichostrongylus in the region (Bailey et al. 2009b).
The reasons why IRG on farmlet C produced this profound effect on WEC and the worm species involved almost certainly relate to its impacts on the free-living ecology of the three main worm species involved. These are likely to operate in three distinct ways as summarised below, with detailed discussion of each following. First, the short grazing periods in the IRG treatments would have largely prevented infection from the current grazing cycle as sheep would have moved to a new paddock by the time significant numbers of infective L3 larvae would have appeared on pasture. Second, the long rest periods between grazings would have ensured significant die-off of infective L3 on pasture before the next grazing. Third, spatial and temporal aggregation of fresh sheep faeces in limited parts of the grazed area on farmlet C ensured that, when rainfall events occurred, which were permissive for development of eggs to L3, only limited parts of the landscape became infective in comparison to farmlets A and B.
On the first of these points, the graze periods on farmlet C were typically less than 5–10 days (Hinch et al. 2013a; Scott et al. 2013a) and tended to be longer in winter and shorter in summer. Under optimal conditions of temperature and moisture, first development of T. colubriformis and H. contortus eggs to the infective L3 stage in faeces takes 3 and 4 days, respectively (Hsu and Levine 1977) with appearance on herbage at Days 5 and 6, respectively (Cheah and Rajamanickam 1997). The timing of this development is strongly temperature-dependent. At 10C, the minimum development time of H. contortus to L3 was ~16 days, and thereafter decreased as temperature increased, ranging from 6.4 days at 20C, 3.5 days at 30C, and 2.5 days at 37C (Smith 1990). Based on these data, it is clear that under the IRG system used on farmlet C, little infection of sheep will occur during a given grazing period due to appearance of L3 on pasture from eggs deposited during that period. Larvae will generally need to survive until the next grazing period to infect a host.
On the second point relating to larval die-off between grazing periods, the factors affecting survival of L3 on pasture have been reviewed by O’Connor et al. (2006). The ensheathed infective L3 is the stage of the lifecycle most resistant to adverse environmental influences but, as the L3 cannot feed, survival is reduced by factors that increase motility or metabolic rate, and thus deplete metabolic reserves, such as rainfall and elevated temperatures. Thus, under tropical conditions in Fiji, complete depletion of L3 on herbage within 5–13 weeks has been reported for H. contortus and T. colubriformis (Banks et al. 1990). Based on these findings, timed, IRG systems that effectively control worms in goats and sheep in the tropics have been devised and successfully implemented (Barger et al. 1994; Gray et al. 2000).
On the other hand, L3 are comparatively resistant to the effects of cold, and survive frosts and cold temperatures in the Armidale region that inhibit hatching of eggs to L3 (Southcott et al. 1976; Bailey et al. 2009a). Larval survival under these conditions is prolonged with Barger et al. (1972) predicting L3 population half-lives of 93 days at 12C and 85% relative humidity (RH), and only 9 days at 28C and 35% RH. At 20C and 65% RH, the half-life was estimated at 33 days. The longer survival period under temperate conditions, coupled with use of rotational grazing schemes that fail to adequately disrupt the nematode lifecycle has resulted in many studies failing to show major benefits of rotational grazing for worm control in sheep in temperate climates (Morgan 1933; Morgan and Oldham 1934; Roe et al. 1959a; Gibson and Everett 1968). Other studies that did show beneficial effects used such long spelling periods that pasture and animal productivity were impaired (Robertson and Fraser 1933; Eysker et al. 2005) as also occurred on farmlet C in this experiment.
Long-term average maximum temperatures for summer, autumn, winter and spring in Armidale are 25.7, 20.1, 13.7 and 20.8C, respectively. Based on these data and a RH of 50%, the model of Barger et al. (1972) would predict half-lives of L3 on pasture of 15.5, 20, 27 and 19.5 days, respectively. Thus, to reduce L3 numbers to 10% of their original value would require spelling times of 49, 64, 88 and 62 days, respectively. On farmlet C, rest periods were rarely below 50 days and sometimes over 150 days, with a mean of 98 days (Table 3) thus allowing time for significant larval die-off in most cases. It should be noted that, in practice, these spelling times will overestimate the decrease in larval populations on pasture as L3 may continue to develop and appear on pasture for several weeks following removal of stock, particularly in the case of T. colubriformis and T. circumcincta (O’Connor et al. 2006). The improved level of control of worms on farmlet C was associated with significantly increased susceptibility to worm infection (Colvin et al. 2012) and reduced animal productivity as shown by reduced liveweights (Hinch et al. 2013a) and fat scores (Hinch et al. 2013b) of livestock on this farmlet relative to those on farmlets A and B. The long rest periods and lower levels of soil fertility on farmlet C were found to be associated with lower levels of both legume and pasture digestibility within this farmlet (Shakhane et al. 2013) and these are likely to have contributed to the reduced productivity.
The third, related reason why the IRG used in farmlet C produced such marked worm control is likely to be spatial and temporal aggregation of faecal deposition such that when rainfall events occurred that were permissive for development of eggs to L3, only limited parts of the landscape became infective. This can be visualised in Fig. 6 by selecting a single date and running a perpendicular line down through all the paddocks in all three farmlets to represent it. If rainfall falls on that date, where the line passes through black areas (stocked) it will fall on freshly deposited faeces whereas, where it passes through white areas it will fall on unstocked paddocks, mostly not containing fresh faeces. Under dry conditions, rainfall just before or in the first few days after faecal deposition is a requirement for successful development of H. contortus and T. colubriformis from egg to infective L3 outside the faecal pellet (Khadijah et al. 2013).

|
Thus on farmlet C, periods of successful development to infective L3 following rainfall, will be spatially restricted to the small paddocks containing sheep during or shortly before the rainfall event. Over much of the farmlet C landscape, rainfall will have no beneficial effect on larval development because there are no fresh faeces available. In ‘hot spots’ where development occurs, the L3 that do develop have to survive the long spell period before being exposed to sheep again. This is not the case on farmlets A and B, where rainfall at any time will fall on fresh faeces on a larger proportion of the landscape facilitating development to L3, which will often be able to infect sheep immediately on emergence from the pasture.
At the start of the farmlet experiment, the dominant nematode genus present was Trichostrongylus. On farmlets A and particularly B, there was a rapid decline in the percentage of Trichostrongylus from ~75% to less than 10% between 2000 and 2003, after which is remained at comparatively low levels. Most of the decline was associated with a large increase in the proportion Haemonchus over the same period. This shift probably reflects the greater reproductive potential of H. contortus under the experimental conditions early in the experiment. On farmlet C the situation differed markedly with less ‘displacement’ of Trichostrongylus by Haemonchus over time, such that there were lower proportions of Haemonchus and higher proportions of Trichostrongylus and Teladorsagia on this farmlet than the other farmlets for most of the period 2002–06. In the broadest terms Haemonchus appeared to have been most successful on farmlet B, Trichostrongylus on farmlet A and Teladorsagia on farmlet C. The lower proportion of Haemonchus seen on farmlet C probably reflects its greater susceptibility to lifecycle disruption by grazing management. As noted by Colvin et al. (2008), it is likely that the rapid rotational grazing system of Barger et al. (1994) under tropical conditions was effective against both H. contortus and Trichostrongylus spp. but that this was unlikely to be the case in the cool, temperate climate of the Northern Tablelands, which is more conducive to the survival and development of Trichostrongylus spp. eggs and larvae between grazing events (Anderson et al. 1966; Waller and Donald 1970; Levine and Andersen 1973; Levine et al. 1974). On the other hand, the susceptibility of H. contortus eggs to desiccation and low temperatures is likely to make them just as susceptible to rapid rotational grazing as they would be under tropical conditions, but probably for different reasons. Temperature and moisture are often limiting in the Northern Tablelands for development of H. contortus eggs to L3 (O’Connor et al. 2006, 2007), and failure of development is likely to be a major mechanism operating in this environment. The superior ability of Trichostrongylus and Teladorsagia species to survive in the embryonated egg stage during cool dry conditions relative to Haemonchus could account for the species difference in response to IRG observed in this experiment.
With regard to the objective of investigating sources of variation in WEC during the experiment, it is perhaps not surprising that the majority of the explained variation was due to the last anthelmintic used, and the time since that last anthelmintic treatment. Replacement of generic factors in the model such as season and farmlet with more detailed and proximate measures of environmental and management factors likely to affect WEC failed to explain a great deal more of the observed variation in WEC. On the other hand, they were useful in teasing out the factors that best accounted for the variation. For temperature, a higher proportion of the variation in WEC was explained by including both TMIN and TMAX in the model, than TMEAN or GDD. For moisture, inclusion of both RAIN and EVAP in the model provided a considerably better fit than the inclusion of the derived variables P/E or with ASW providing an intermediate fit. Replacement of the effect of farmlet with both SR and GP resulted in a higher model R2 whereas the inclusion of either alone did not. In both cases there was a significant positive linear association between SR and GP on LogWEC.
Regarding the objective of investigating the efficacy of anthelmintic treatments during the experiment, the tactical approach used, based upon routine WEC monitoring, provided reasonable control of worms on all three farmlets for much of the experimental period. However, this approach failed to prevent significant spikes in WEC to values associated with significant production loss on multiple occasions, and mortality on limited occasions, particularly on farmlets A and B. As the results of monitoring showed increasing WEC numbers later in the experiment, the Cicerone Management Board recognised that the experimental guidelines laid down for farmlets A and B – of eight paddocks and a minimum of five mobs on each farmlet – were overly constraining in terms of flexible grazing options. Thus, a decision was taken late in the trial to increase the number of paddocks on each of these farmlets to 10, and to increase the paddock number on farmlet C to 40, however these changes were implemented too late to observe the effect of such changes.
When Haemonchus is the dominant parasite, effects are likely to be minimal at WEC below 1000, with significant production losses likely between 1000 and 5000, clinical signs and severe production losses between 5 and 10 000 and deaths can be expected in the range 10–30 000 (Brightling 1994). When Haemonchus is not present, the respective values are <200, 200–500, 500–2000 and >2000. The Haemonchus values are consistent with the WEC-associated mortality observed on farmlets A and B in December 2004. To prevent such spikes, the frequency of WEC sampling during the high-risk months in late spring and summer may need to be increased, and more sophisticated treatment thresholds adopted. Adoption of a tactical integrated parasite management program based on routine WEC monitoring has been shown in other studies to reduce the adverse impact of worms and frequency of anthelmintic treatment (Kelly et al. 2010) and such an IPM approach is central to the recommendations of WormBoss, Australia’s national sheep worm control program (http://www.wormboss.com.au/, accessed 1 April 2013). Adoption of the WormBoss summer rainfall IPM program could be expected to reduce the likelihood of the spikes in WEC observed in this experiment.
The overall frequency of anthelmintic treatment of four per annum in the present experiment (Table 3) is well above those reported for the Northern Tablelands region in the IPM 2004 Benchmarking Survey (Reeve and Thompson 2005). In that study the 161 respondents from that region reported a mean treatment frequency of 3.1 in adult ewes in 2003, compared with mean values of 6, 5 and 4 in that year for farmlets A, B and C. This is likely due to the significant numbers of quarantine drenches used on the experiment (two per year for ewes and one per year for other sheep), the tactical approach used in the study, with intensive WEC monitoring taking place, and also the predominant use of short- rather than long-acting anthelmintics. The FECRT conducted on farmlet C in 2003 showed that there was evidence of resistance to ivermectin and albendazole but neither was used alone after this point. There was evidence of resistance to closantel based on a reduced duration of protection, and this anthelmintic was also not used alone after 2002.
In conclusion, the results show that, in a summer rainfall temperate environment with H. contortus as the dominant parasite, improved host nutrition in a higher input system (farmlet A) did not provide more effective control of gastrointestinal nematodes than typical management (farmlet B); however, it was observed that gastrointestinal nematode control was no worse on farmlet A than on farmlet B in spite of farmlet A supporting a 48% higher SR by later in the trial period (2005). The experiment provided strong support for the proposition that IRG (farmlet C) does provide more effective control of gastrointestinal nematodes as evidenced by significantly lower WEC counts and anthelmintic treatment frequency. As reported elsewhere (Colvin et al. 2012) this effect is mediated by reduced larval challenge on pasture, rather than improved host resistance. Indeed host resistance to worms was lower on farmlet C due to reduced exposure. Tactical worm control based on routine monitoring of WEC provided adequate control of worms on all three farmlets for much of the experimental period but failed to prevent significant spikes in WEC to values associated with significant production loss on multiple occasions, and mortality on limited occasions, particularly on farmlets A and B.
The most important factors affecting WEC in the experiment were the time since the last anthelmintic treatment, and the anthelmintic used at that treatment. The magnitude of these effects dwarfed those climatic and management factors that might be expected to influence the epidemiology of gastrointestinal nematode infections via environmental or host-mediated mechanisms.
Acknowledgements
The Cicerone farmlet experiment was generously funded by Australian Wool Innovation with support for postgraduate scholarships from the Australian Sheep CRC and the University of New England. The considerable in-kind contributions from the CSIRO, University of New England, NSW Department of Primary Industries (DPI), Betty Hall Pty Ltd and NSW Technical and Further Education are gratefully acknowledged. The expert livestock extension advice of Mr Bob Marchant of NSW DPI was of great assistance. The support and practical assistance of the Cicerone Farm Manager, Mr Justin Hoad and the Cicerone Executive Office, Ms Caroline Gaden, are gratefully acknowledged.
References
Anderson FL, Wang G, Levine ND (1966) Effect of temperature on survival of the free-living stages of Trichostrongylus colubriformis. The Journal of Parasitology 52, 712–721.Bailey JN, Kahn LP, Walkden-Brown SW (2009a) Availability of gastro-intestinal nematode larvae to sheep following winter contamination of pasture with six nematode species on the northern tablelands of New South Wales. Veterinary Parasitology 160, 89–99.
| Availability of gastro-intestinal nematode larvae to sheep following winter contamination of pasture with six nematode species on the northern tablelands of New South Wales.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD1M7ksVOrtQ%3D%3D&md5=55666b76df4ad4194108a189206cbf27CAS | 19070435PubMed |
Bailey JN, Kahn LP, Walkden-Brown SW (2009b) The relative contributions of T. colubriformis, T. vitrinus, T. axei and T. rugatus to sheep infected with Trichostrongylus spp. on the Northern Tablelands of New South Wales. Veterinary Parasitology 165, 88–95.
| The relative contributions of T. colubriformis, T. vitrinus, T. axei and T. rugatus to sheep infected with Trichostrongylus spp. on the Northern Tablelands of New South Wales.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD1MnmsVKjug%3D%3D&md5=18dee4195c86aacf86008196680872d7CAS | 19632782PubMed |
Banks DJD, Singh R, Barger IA, Pratap B, Le Jambre LF (1990) Development and survival of infective larvae of Haemonchus contortus and Trichostrongylus colubriformis on pasture in a tropical environment. International Journal for Parasitology 20, 155–160.
| Development and survival of infective larvae of Haemonchus contortus and Trichostrongylus colubriformis on pasture in a tropical environment.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK3c3js1GlsQ%3D%3D&md5=df8c6faa57aba6e31467450e4357269bCAS |
Barger IA, Benyon PR, Southcott WH (1972) Simulation of pasture larval populations of Haemonchus contortus. Proceedings of the Australian Society of Animal Production 9, 38–42.
Barger IA, Siale K, Banks DJD, Le Jambre LF (1994) Rotational grazing for control of gastrointestinal nematodes of goats in a wet tropical environment. Veterinary Parasitology 53, 109–116.
| Rotational grazing for control of gastrointestinal nematodes of goats in a wet tropical environment.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK2cznvVCntw%3D%3D&md5=108756037b4c0accdb52b01c5fca634bCAS | 8091608PubMed |
Behrendt K, Scott JM, Mackay DF, Murison R (2013) Comparing the climate experienced during the Cicerone farmlet experiment against the climatic record. Animal Production Science 53, 658–669.
| Comparing the climate experienced during the Cicerone farmlet experiment against the climatic record.Crossref | GoogleScholarGoogle Scholar |
Brightling A (1994) ‘Stock diseases.’ (Inkata Press: Melbourne) 328 pp.
Cheah TS, Rajamanickam C (1997) Epidemiology of gastrointestinal nematodes of sheep in wet tropical conditions in Malaysia. Tropical Animal Health and Production 29, 165–173.
| Epidemiology of gastrointestinal nematodes of sheep in wet tropical conditions in Malaysia.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK2svmtF2mug%3D%3D&md5=e01c75a06ce1770f5809d3ec56ea0133CAS | 9316233PubMed |
Coles GC, Bauer C, Borgsteede FHM, Geerts S, Klei TR, Taylor MA, Waller PJ (1992) World Association for the Advancement of Veterinary Parasitology (W.A.A.V.P.) methods for the detection of anthelmintic resistance in nematodes of veterinary importance. Veterinary Parasitology 44, 35–44.
| World Association for the Advancement of Veterinary Parasitology (W.A.A.V.P.) methods for the detection of anthelmintic resistance in nematodes of veterinary importance.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK3s%2FmvF2hsg%3D%3D&md5=a850be27f95c4ab8a676b2e744a9165aCAS | 1441190PubMed |
Colvin AF, Walkden-Brown SW, Knox MR, Scott JM (2008) Intensive rotational grazing assists control of gastrointestinal nematodosis of sheep in a cool temperate environment with summer-dominant rainfall. Veterinary Parasitology 153, 108–120.
| Intensive rotational grazing assists control of gastrointestinal nematodosis of sheep in a cool temperate environment with summer-dominant rainfall.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD1c3ivVSisA%3D%3D&md5=fa1fe9efa94c6be0d4f342455e6e43b7CAS | 18339483PubMed |
Colvin AF, Walkden-Brown SW, Knox MR (2012) Role of host and environment in mediating reduced gastrointestinal nematode infections in sheep due to intensive rotational grazing. Veterinary Parasitology 184, 180–192.
| Role of host and environment in mediating reduced gastrointestinal nematode infections in sheep due to intensive rotational grazing.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC387mtVGluw%3D%3D&md5=51190433da30d47b954e2b02b48ae9a9CAS | 21924833PubMed |
Eysker M, Bakker N, Kooyman FNJ, Ploeger HW (2005) The possibilities and limitations of evasive grazing as a control measure for parasitic gastroenteritis on small ruminants in temperate environments. Veterinary Parasitology 129, 95–104.
| The possibilities and limitations of evasive grazing as a control measure for parasitic gastroenteritis on small ruminants in temperate environments.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2M7osFCitw%3D%3D&md5=28c07f53410acb100bfb2626711bd074CAS | 15817209PubMed |
Gibson TE, Everett G (1968) A comparison of set stocking and rotational grazing for the control of Trichostrongylosis in sheep. The British Veterinary Journal 124, 287
Gordon HML, Whitten L (1941) A note on variations in the efficiency of the copper sulphate and nicotine sulphate drench against Haemonchus contortus. Australian Veterinary Journal 17, 172–176.
| A note on variations in the efficiency of the copper sulphate and nicotine sulphate drench against Haemonchus contortus.Crossref | GoogleScholarGoogle Scholar |
Gray GD, Anchetta PA, Arguzon JA, Barcelo PM, Casis C, Cerbito WA, Dobson RJ, Dumilon R, Cruz EM, Knox MR, Le Jambre LF, Mateo KS, Suba MS, Villar EC (2000) Better worm control for goats and sheep in the Philippines. Asian-Australasian Journal of Animal Sciences 13, 548
Healey AF, Hall E, Gaden CA, Scott JM, Walkden-Brown SW (2004) Intensive rotational grazing reduces nematode faecal egg counts in sheep on the Cicerone Project. Proceedings of the Australian Society of Animal Production 25, 85–88.
Hinch GN, Hoad J, Lollback M, Hatcher S, Marchant R, Colvin A, Scott JM, Mackay D (2013a) Livestock weights in response to three whole-farmlet management systems. Animal Production Science 53, 727–739.
| Livestock weights in response to three whole-farmlet management systems.Crossref | GoogleScholarGoogle Scholar |
Hinch GN, Lollback M, Hatcher S, Hoad J, Marchant R, Mackay DF, Scott JM (2013b) Effects of three whole-farmlet management systems on Merino ewe fat scores and reproduction. Animal Production Science 53, 740–749.
| Effects of three whole-farmlet management systems on Merino ewe fat scores and reproduction.Crossref | GoogleScholarGoogle Scholar |
Hsu C-K, Levine ND (1977) Degree-day concept in development of infective larvae of Haemonchus contortus and Trichostrongylus colubriformis under constant and cyclic conditions. American Journal of Veterinary Research 38, 1115–1119.
Hutchinson GK, Richards K, Risk WH (2000) Aspects of accumulated heat patterns (growing degree-days) and pasture growth in Southland. Proceedings of the New Zealand Grassland Association 62, 81–85.
Kaine G, Doyle B, Sutherland H, Scott JM (2013) Surveying the management practices and research needs of graziers in the New England region of New South Wales. Animal Production Science 53, 602–609.
| Surveying the management practices and research needs of graziers in the New England region of New South Wales.Crossref | GoogleScholarGoogle Scholar |
Kelly GA, Kahn LP, Walkden-Brown SW (2010) Integrated Parasite Management for sheep reduces the effects of gastrointestinal nematodes on the Northern Tablelands of NSW. Animal Production Science 50, 1043–1052.
| Integrated Parasite Management for sheep reduces the effects of gastrointestinal nematodes on the Northern Tablelands of NSW.Crossref | GoogleScholarGoogle Scholar |
Kemper K, Walkden-Brown SW (2004) Methods for detecting anthelmintic resistance: a case study on the New England Tablelands. Proceedings of the Australian Sheep Veterinary Society 14, 122–129.
Khadijah S, Kahn LP, Walkden-Brown SW, Bailey JN, Bowers SF (2013) Effect of simulated rainfall timing on faecal moisture and development of Haemonchus contortus and Trichostrongylus colubriformis eggs to infective larvae. Veterinary Parasitology 192, 199–210.
Levine ND, Andersen FL (1973) Development and survival of Trichostrongylus colubriformis on pasture. The Journal of Parasitology 59, 147–165.
| Development and survival of Trichostrongylus colubriformis on pasture.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaE3s7itFKmsA%3D%3D&md5=3fe24a2b3c514bebd4fc4fbd50b0c29aCAS | 4734543PubMed |
Levine ND, Todd KSJ, Boatman PA (1974) Development and survival of Haemonchus contortus on pasture. American Journal of Veterinary Research 35, 1413–1422.
Love S (2012) Drench resistance and sheep worm control. (NSW Industry and Investment) Available at http://www.dpi.nsw.gov.au/__data/assets/pdf_file/0009/111060/Drench-resistance-and-sheep-worm-control.pdf [Verified 19 October 2012]
McLeod RS (1995) Costs of major parasites to the Australian livestock industries. International Journal for Parasitology 25, 1363–1367.
| Costs of major parasites to the Australian livestock industries.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK283ivFaktw%3D%3D&md5=bb8da80e4912eeb782a0f4d894451f92CAS | 8635886PubMed |
Morgan DO (1933) The effect of heavy stocking on the worm burden under a system of rotational grazing. Journal of Helminthology 11, 169–180.
| The effect of heavy stocking on the worm burden under a system of rotational grazing.Crossref | GoogleScholarGoogle Scholar |
Morgan DO, Oldham JN (1934) Further observations on the effect of heavy stocking on the worm burden under a system of rotational grazing. Journal of Helminthology 12, 177–182.
| Further observations on the effect of heavy stocking on the worm burden under a system of rotational grazing.Crossref | GoogleScholarGoogle Scholar |
Morgan ER, Cavill L, Curry GE, Wood RM, Mitchell ESE (2005) Effects of aggregation and sample size on composite faecal egg counts in sheep. Veterinary Parasitology 131, 79–87.
| Effects of aggregation and sample size on composite faecal egg counts in sheep.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2Mzhslahtw%3D%3D&md5=626c1d4e0a73c9faa440f3c81bf7d929CAS | 15921855PubMed |
O’Connor LJ, Walkden-Brown SW, Kahn LP (2006) Ecology of the free-living stages of major trichostrongylid parasites of sheep. Veterinary Parasitology 142, 1–15.
| Ecology of the free-living stages of major trichostrongylid parasites of sheep.Crossref | GoogleScholarGoogle Scholar | 17011129PubMed |
O’Connor LJ, Kahn LP, Walkden-Brown SW (2007) The effects of amount, timing and distribution of simulated rainfall on the development of Haemonchus contortus to the infective larval stage. Veterinary Parasitology 146, 90–101.
| The effects of amount, timing and distribution of simulated rainfall on the development of Haemonchus contortus to the infective larval stage.Crossref | GoogleScholarGoogle Scholar | 17398009PubMed |
Reeve I, Thompson L-J (2005) ‘Integrated parasite management in sheep project: Benchmark Survey.’ (Australian Wool Innovation Limited: Sydney)
Robertson D, Fraser AHH (1933) On the incidence of stomach worms in lambs in the North of Scotland and their control by progressive sectional grazing. Journal of Helminthology 11, 187–194.
| On the incidence of stomach worms in lambs in the North of Scotland and their control by progressive sectional grazing.Crossref | GoogleScholarGoogle Scholar |
Roe R, Southcott W, Turner H (1959a) Grazing management of native pastures in the New England region of New South Wales. I. Pasture and sheep production with special reference to systems of grazing and internal parasites. Australian Journal of Agricultural Research 10, 530–554.
| Grazing management of native pastures in the New England region of New South Wales. I. Pasture and sheep production with special reference to systems of grazing and internal parasites.Crossref | GoogleScholarGoogle Scholar |
Roe R, Southcott WH, Turner HN (1959b) Grazing management of native pastures in the New England region of New South Wales. I. Pasture and sheep production with special reference to systems of grazing and internal parasites. Australian Journal of Agricultural Research 10, 530–554.
| Grazing management of native pastures in the New England region of New South Wales. I. Pasture and sheep production with special reference to systems of grazing and internal parasites.Crossref | GoogleScholarGoogle Scholar |
Sackett D, Holmes P, Abbott K, Jephcott S, Barber M (2006) Assessing the economic cost of endemic disease on the profitability of Australian beef cattle and sheep producers. Final Report of Project AHW.087. Meat and Livestock Australia, Sydney.
Scott JM, Gaden CA, Edwards C, Paull DR, Marchant R, Hoad J, Sutherland H, Coventry T, Dutton P (2013a) Selection of experimental treatments, methods used and evolution of management guidelines for comparing and measuring three grazed farmlet systems. Animal Production Science 53, 628–642.
| Selection of experimental treatments, methods used and evolution of management guidelines for comparing and measuring three grazed farmlet systems.Crossref | GoogleScholarGoogle Scholar |
Scott JM, Gaden CA, Edwards C, Paull DR, Marchant R, Hoad J, Sutherland H, Coventry T, Dutton P (2013b) Selection of experimental treatments, methods used and evolution of management guidelines for comparing and measuring three grazed farmlet systems. Animal Production Science 53, 628–642.
| Selection of experimental treatments, methods used and evolution of management guidelines for comparing and measuring three grazed farmlet systems.Crossref | GoogleScholarGoogle Scholar |
Shakhane LM, Mulcahy C, Scott JM, Hinch GN, Donald GE, Mackay DF (2013) Pasture herbage mass, quality and growth in response to three whole-farmlet management systems. Animal Production Science 53, 685–698.
| Pasture herbage mass, quality and growth in response to three whole-farmlet management systems.Crossref | GoogleScholarGoogle Scholar |
Smith G (1990) The population biology of the free-living phase of Haemonchus contortus. Parasitology 101, 309–316.
| The population biology of the free-living phase of Haemonchus contortus.Crossref | GoogleScholarGoogle Scholar | 2263426PubMed |
Southcott W, Major G, Barger I (1976) Seasonal pasture contamination and availability of nematodes for grazing sheep. Australian Journal of Agricultural Research 27, 277–286.
| Seasonal pasture contamination and availability of nematodes for grazing sheep.Crossref | GoogleScholarGoogle Scholar |
Sutherland H, Scott JM, Gray GD, Woolaston RR (2013) Creating the Cicerone Project: seeking closer engagement between livestock producers, research and extension. Animal Production Science 53, 593–601.
| Creating the Cicerone Project: seeking closer engagement between livestock producers, research and extension.Crossref | GoogleScholarGoogle Scholar |
Waller PJ, Donald AD (1970) The response to desiccation of eggs of Trichostrongylus colubriformis and Haemonchus contortus (Nematoda: Trichostrongylidae). Parasitology 61, 195–204.
| The response to desiccation of eggs of Trichostrongylus colubriformis and Haemonchus contortus (Nematoda: Trichostrongylidae).Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaE3M%2FktlWrtw%3D%3D&md5=c5c7f24b303ad769748e470c8541c619CAS | 5530099PubMed |
Whitlock HV (1948) Some modifications of the McMaster helminth egg-counting technique and apparatus. Journal of the Council for Scientific and Industrial Research 21, 177–180.