

Evaluation of the productivity and feed value of Wondergraze and Redlands leucaena cultivars under grazing
E. Charmley A * , C. S. McSweeney B , G. J. Bishop-Hurley B , J. Simington A , J. Padmanabha B and P. Giacomantonio B
A * , C. S. McSweeney B , G. J. Bishop-Hurley B , J. Simington A , J. Padmanabha B and P. Giacomantonio B
A CSIRO Agriculture and Food, Private Mail Bag PO, Aitkenvale, Townsville, Qld 4814, Australia.
B CSIRO Agriculture and Food, 306 Carmody Road, St Lucia, Qld 4067, Australia.
Animal Production Science 63(5) 450-462 https://doi.org/10.1071/AN22341
Submitted: 2 September 2022 Accepted: 24 November 2022 Published: 4 January 2023
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Leucaena leucocephala (leucaena) is a leguminous shrub used for beef grazing in low-rainfall regions (<600–700 mm). Newer cultivars have the potential to extend adoption of the species to higher rainfall (>600–700 mm), frost-free areas of Australia.
Aim: We compared productivity, nutritional value and animal performance of two leucaena cultivars, new psyllid-resistant Redlands and the 2010-released Wondergraze, under continuous grazing management in a higher rainfall environment.
Methods: Growing steers were allocated to replicated established stands of Wondergraze or Redlands with inter-row mixed grass–legume pasture from January to July 2021. Pasture and leucaena were characterised for biomass and nutritive characteristics. Botanical composition was measured. Liveweight gain, rumen fermentation, and leucaena mimosine breakdown products were measured in grazing steers.
Key results: At the beginning of the study, leucaena edible biomass was similar for both cultivars (P > 0.05), but at subsequent samplings, biomass of Redlands was lower than of Wondergraze (P < 0.01). Biomass of both cultivars declined rapidly over the grazing period. Pasture biomass increased between February and July and was significantly higher in Wondergraze paddocks (P < 0.05). Animal performance was not significantly different between cultivar treatments, averaging 0.8 kg/day, but declined over time. Patterns of mimosine conversion to DHP isomers and their conjugation were similar for the two cultivars, suggesting that effectiveness of detoxification did not differ between them.
Conclusion: The results demonstrate that leucaena can sustain high levels of animal performance when included in tropical grass pastures in a higher rainfall environment if present in a sufficient quantity (>2 t leucaena edible dry matter/ha established leucaena).
Implications: Grazing leucaena–grass pastures is an effective means of increasing animal productivity in parts of subtropical Australia. However, managing grass and/or leucaena growth to match animal requirements can be challenging.
Keywords: beef cattle, growth rate, legumes, leucaena, mimosine, pasture utilisation, steers, tropical pastures.
Introduction
Leucaena leucocephala (leucaena) is a leguminous shrub adapted to higher rainfall (>600 mm), frost-free areas of northern Australia. It can provide a source of high-quality forage for cattle grazing tropical grass-based pastures (Tomkins et al. 2019). These C4 grasses are typically deficient in nutrient content for adequate levels of performance, particularly in the dry season (Poppi and McLennan 2010). Shelton and Dalzell (2007) estimate that ~13.5 Mha of land is suitable for planting leucaena in the state of Queensland; this represents 9.2% of the grazing area in the state. Beutel et al. (2018) estimated that, in 2018, leucaena was present in 123 500 ha of pastures. Leucaena has a deep and well-developed taproot facilitating rapid growth and greater access to water and nutrient reserves in lower soil horizons than grasses. As a result, leucaena is a productive, high-quality forage, typically having a crude protein (CP) content of 15–25% (Shelton and Brewbaker 1998; Shelton and Dalzell 2007; Radrizzani et al. 2011). Early research with established cultivars of L. leucocephala subsp. glabrata established that liveweight (LW) gains of ~0.7 kg/day could be achieved when leucaena was included in grass pasture diets over 6 months (Quirk et al. 1990; Petty et al. 1998; Petty and Poppi 2012). More recently, Bowen et al. (2018) compared six forage types in central Queensland and showed annual LW gain of 198 kg/ha for grass–leucaena mixes, which exceeded the LW gain from other forages in the study, including oats (Avena sativa), sorghum (Sorghum spp.), lablab (Lablab purpureus), and C4 grass species–butterfly pea (Clitoria ternatea) mixtures. Harrison et al. (2015) compared cattle grazing Rhodes grass (Chloris gayana) pastures with and without leucaena and showed a 50% increase in LW gain over 14 months when leucaena was included in the pasture.
Recently new leucaena cultivars have become available, specifically selected for yield, quality and psyllid resistance (Dalzell 2019), but they have not been thoroughly evaluated for their impact on animal performance. According to Lemin et al. (2019), the susceptibility of leucaena to attack from the psyllid Heteropsylla cubana leads to extensive leaf loss, and this has restricted adoption. Wondergraze is a relatively new cultivar, released in 2010, derived from L. leucocephala subsp. glabrata and specifically bred for forage yield and a branched tree form (Dalzell 2019). Redlands is a newer cultivar, released in 2017; psyllid resistance has been bred into the line through backcrossing with L. pallida (Dalzell 2019). A field grazing trial of these two cultivars was established in North Queensland and has demonstrated the psyllid resistance of Redlands and its similar animal performance to Wondergraze (Lemin et al. 2019).
Leucaena contains mimosine, which is broken down in the rumen to 3-hydroxy-4(1h)-pyridone (3,4-DHP) and 2,3-DHP (Dalzell et al. 2012). Mimosine and its breakdown products are toxic to ruminants and can result in a range of symptoms ranging from poor thrift to, in extreme cases, death (Jones and Hegarty 1984). Mimosine is anti-mitotic, disrupting cell division, often characterised by alopecia (Halliday et al. 2013). The DHP breakdown products are goitrogenic, reducing iodine availability, and they can chelate the metal ions zinc, copper and iron. The resultant deficiencies can be exhibited as a range of non-specific symptoms that collectively can reduce intake, performance and fertility (Halliday et al. 2013). Consequently, cattle are rumen-inoculated with a mixed culture of Synergistes jonesii, which metabolises 3,4-DHP to 2,3-DHP before degrading the latter isomer to non-toxic end products. However, it has become apparent that the detoxification process may be more complex, and doubt has been cast on the ability of the inoculum to break down DHP completely and protect against toxicity (Halliday et al. 2014). It has been postulated that hepatic conjugation of the 2,3-DHP isomer also contributes to reducing toxicity (Halliday et al. 2013, 2014).
The present study was designed to evaluate leucaena cvv. Wondergraze and Redlands in terms of agronomic performance, nutritive value, animal performance and rumen characteristics, including the fate of mimosine in rumen fluid and urine. We hypothesised that there would be no cultivar effect on any of these indexes of productivity or toxicity.
Materials and methods
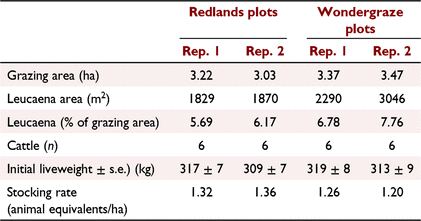
A 20-ha block of leucaena was established in February 2017 at the Lansdown Research Station (19°39′S, 146°50′E), 45 km south of Townsville, Queensland. The intention was to establish half of the area with the new psyllid-resistant cultivar, Redlands, and half with Wondergraze. Limited availability of seed restricted the area sown to Redlands to ~7 ha, comprising 12 double rows each 12 m apart and 400 m in length. The remaining area (13 ha) was seeded to Wondergraze, using the same spacing and row configuration. However, only 14 of the 25 rows of Wondergraze were used for the study. Effective row width of leucaena was 1 m, as assessed on canopy spread of the double-row planting configuration. Therefore, the row area was 400 m2. Establishment of both cultivars was sporadic, with estimated establishment of 62.4% and 72.9% of seeded area for Redlands and Wondergraze, respectively. Soil samples were taken in March 2018 and analysed for pH, phosphorus (P), calcium, potassium, magnesium, sodium and trace elements. Soil was found to be deficient in P (7 mg/kg) and marginal for sulfur (7 mg/kg). Single superphosphate (250 kg/ha) and muriate of potash (150 kg/ha) were applied in September 2018. Repeat soil sampling in 2019 revealed only modest increases in P, with levels still below requirements for leucaena (20 mg/kg; Dalzell et al. 2006). The grazing study described here was conducted during 2021. In previous years, leucaena was slashed to a height of 30 cm above ground level at the end of the dry season, and regrowth was subsequently grazed or manually harvested to provide the legume for indoor studies (Stifkens et al. 2022).
The pasture grasses comprised Indian couch (Bothriochloa pertusa), Queensland bluegrass (Dichanthium sericeum) black speargrass (Heteropogon contortus) and sabi grass (Urochloa mosambicensis). Other legumes, seca stylo (Stylosanthes scabra) and Desmanthus spp., were also present in the pasture, as well as weeds including sicklepod (Senna obtusifolia), snakeweed (Stachytarpheta spp.), sida (Sida acuta) and soft khakiweed (Gomphrena celosioides).
Animals and paddocks
The grazing study complied with the Australian Code for the Care and Use of Animals for Scientific Purposes and was approved by the CSIRO Queensland Animal Ethics Committee (AEC Number: 2020-06).
The grazing trial commenced on 29 January and continued through to 14 July 2021 (a 166-day grazing season). Rainfall in the 4 months to the end of April 2021 totalled 834 mm. The block was divided into four paddocks (two replicates for each cultivar) ranging in size from ~3.0 to 3.5 ha (Table 1), and total leucaena area accounted for ~6–8% of each paddock/grazing area. Paddocks were serviced by a waterpoint situated on the fence line between two adjacent leucaena replicates. Total length of established leucaena was 3699 m for Redlands and 4985 m for Wondergraze. Twelve cattle were allocated per cultivar, divided between the two replicates; each replicate was stocked with six Droughtmaster steers (initial bodyweight 314 ± 4 (s.e.) kg). Cattle were introduced to leucaena paddocks on 28 January 2021. After a 6-day adaption to leucaena, cattle were individually weighed and drenched with an inoculum (100 mL) of a bacterial suspension comprising S. jonesii specifically adapted to the respective cultivars (D. Ouwerkerk, pers. comm.) according to the recommendations of FutureBeef (Leucaena inoculum for cattle see https://futurebeef.com.au/resources/leucaena-inoculum/). Animal performance was assessed by weighing cattle at 4-week or 6-week intervals.
|
Sampling and analysis
Diet characteristics were assessed using techniques based on the BOTANAL method of Tothill et al. (1992). Sampling was conducted on four occasions at intervals of ~6–8 weeks: 17 February, 31 March, 2 June, 19 July 2021. Pasture biomass was assessed using the BOTANAL method with quadrats (0.25 m2) placed at 40-m intervals in pasture inter-rows between leucaena rows. The proportions of grass, weeds and legumes in pasture, bare ground and green material were estimated by visual assessment of quadrats. Mean data from 10 quadrat observations were used as the experimental unit, this representing pasture from one inter-row. One quadrat in 10 within each inter-row was randomly sampled for subsequent near infrared spectroscopy (NIRS) assessment of nutritive value. Leucaena biomass was estimated using BOTANAL principles with the following modifications, similar in approach to Andrew et al. (1979). Five 3-m lengths of leucaena rows were selected to represent lowest to highest biomass, based on visual appraisal. These were used as reference areas (akin to the A, B, C, D, E quadrat samples used for BOTANAL in pasture) for visual assessments taken along all leucaena rows at 40-m intervals. Reference areas were then harvested, and leaves, stems <10 mm and green pods collected, dried and weighed to give a dry yield of edible dry matter (DM) per m. Dried samples were subsequently analysed for nutritive value using NIRS (five samples per paddock).
The nutritive value of pasture and leucaena was characterised from NIR estimates of nitrogen (N), acid detergent fibre (ADF), neutral detergent fibre (NDF), and DM and organic matter (OM) digestibility. CP was assumed to be N × 6.25, and hemicellulose calculated as the difference between NDF and ADF. A full description of the NIR method is given by Stifkens et al. (2022).
The nutritive value of ingested forages was estimated from NIR analysis of faecal samples collected at intervals of ~12 weeks on 17 February, 27 April and 19 July and analysed for dietary N, faecal N, ADF, NDF, and DM and OM digestibility. Faecal samples were dried at 65°C to constant weight and ground to pass a 1-mm sieve before being analysed using NIR methods as described by Stifkens et al. (2022). The ratio of C3:C4 plants was determined using faecal NIR based on the δ13C method (Coates and Dixon 2008; Norman et al. 2009). C3 plants were assumed to be legumes or weeds (non-grass) and were assigned a δ13C value of −28.3‰, whereas C4 plants were assumed to be tropical grasses and assigned a value of −14.4‰ (Bowen et al. 2018). The mean mimosine and 3,4-DHP concentration in leucaena samples was estimated in DM using methods described below. Rumen samples were taken on three occasions at the same time as faecal samples for assessment of rumen pH, ammonia-N and volatile fatty acids according to methods described by Stifkens et al. (2022). Rumen samples were collected by using an oral stomach tube while the animal was restrained in a commercial cattle crush. Mimosine, 3,4-DHP and 2,3-DHP concentrations in rumen fluid were analysed as described below for non-hydrolysed urine samples.
Urine samples were collected from all steers observed to urinate following mustering and while held in a race prior to faecal sampling and weighing on 17 February and 27 April. Typically, samples were collected from about half of the cattle. Samples were stored at −80°C then filtered and hydrolysed before high performance liquid chromatography (HPLC) analysis, following a similar procedure to Dalzell et al. (2012) with some modifications. Rumen fluid samples were also collected from the same animals. Frozen urine and rumen fluid aliquots were thawed, and particulates were removed by passing through syringe filters (13 mm diameter, with PVDF membrane of pore size 0.45 μm; PhaseSep, Melbourne, Vic., Australia). For non-hydrolysed samples, filtered urine (1.5 mL) was acidified to give a final concentration of ~0.1 M HCl, pH ~3, by adding 15 µL concentrated HCl (32% w/w) in HPLC vials. Separately, duplicate hydrolysis reactions were made by mixing 125 µL filtered urine diluted 1:1 with 125 µL concentrated HCl (32% w/w) in individual 300-µL PCR reaction tubes. Reactions were incubated for 180 min at 98°C in a PCR machine to release DHP conjugates (Hegarty et al. 1964; Dalzell et al. 2012). Following hydrolysis, PCR tubes were twice centrifuged for 20 min at 3200g, with supernatants transferred to fresh tubes between spins to remove particulates. After the second spin, supernatant (~200 µL) was transferred to an HPLC vial with a low-volume insert (Agilent Technologies, Santa Clara, CA, USA). Concentrations of mimosine, 3,4-DHP and 2,3-DHP in the samples were determined using the following HPLC procedure. Each non-hydrolysed sample was run in triplicate (three injections), whereas each hydrolysed sample was run in duplicate using HPLC (four injections). A 10-µL aliquot of each sample was injected on a Luna 3 µm C18(2) 100 Å, 150 mm × 4.6 mm LC Column (Phenomenex, Torrance, CA, USA) with a SecurityGuard guard cartridge (Phenomenex) on a Dionex Ultimate3000 HPLC (Thermo Fisher, Waltham, MA, USA). Samples were run in a corrected mobile phase (Halliday 2017) of 25 mM ammonium dihydrogen phosphate (pH 2.25) at a flow rate of 1 mL/min. Concentrations of analytes were measured using a UV diode array detector at the following wavelengths and retention times (RT): mimosine – λ 280 nm, RT 3.3 min; 3,4-DHP – λ 277 nm, RT 4.42 min; 2,3-DHP – λ 295 nm, RT 11.69 min; 2,3-DHP primary conjugate – λ 295 nm, RT 10.22 min; 2,3-DHP secondary conjugate – λ 300 nm, RT 6.47 min; uric acid – λ 280 nm, RT 7.46 min. Concentrations of mimosine, 3,4-DHP and 2,3-DHP analytes were calculated with standard curves derived from five dilutions each of mimosine (0.375, 0.75, 1.5, 3, 6 mM) (Sigma-Aldrich, St. Louis, MO, USA), 3,4-DHP (0.75, 1.5, 3, 6, 12 mM) (Toroma Organics, Saarbruecken, Germany), and 2,3-DHP (0.75, 1.5, 3, 6 mg/mL) (Sigma-Aldrich). Mimosine concentrations in hydrolysed urine are not reported because losses occurred from the acid hydrolysis.
Statistical analyses
All statistical analyses were performed using general linear models by SAS (SAS Institute, Cary, NC, USA) in a fixed-effect model. The experimental design allowed for comparison between Redlands and Wondergraze leucaena cultivars independently within each sampling period. The effect of paddock replicate and the interaction between paddock replicate and cultivar were included in the analysis. In the majority of cases, these factors were non-significant, so interaction data are not shown; where significant effects were detected, these are indicated in tabulated data. Linear and quadratic contrasts were used to characterise the change in variables over time. The row (leucaena) or inter-row (pasture) was the experimental unit for agronomic data, whereas the steer was the experimental unit for animal data. Probability was considered significant at P = 0.05 for all variables.
Results
Effects on biomass and botanical composition
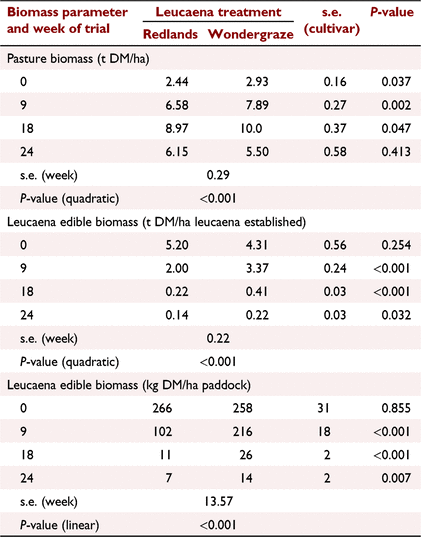
At the beginning of the study, pasture biomass was >2 t/ha and was ~20% greater in Wondergraze than Redlands paddocks (P < 0.04; Table 2). Pasture biomass followed a quadratic response with time (P < 0.001), rapidly increasing early in the season followed by a decline after the Week 18 sampling. At the beginning of the study, leucaena edible biomass was similar for both cultivars (P > 0.05), but at subsequent samplings biomass of Redlands was lower than of Wondergraze (P < 0.01). Unlike the pasture biomass, leucaena biomass declined quickly throughout the grazing period, whether expressed per leucaena area (quadratic effect, P < 0.001) or per paddock area (linear effect, P < 0.001). By Week 18, edible leucaena contributed <0.2% of pasture biomass, which comprised grass, other legumes and weeds. Periodic visual observations failed to find psyllid activity on either cultivar.
|
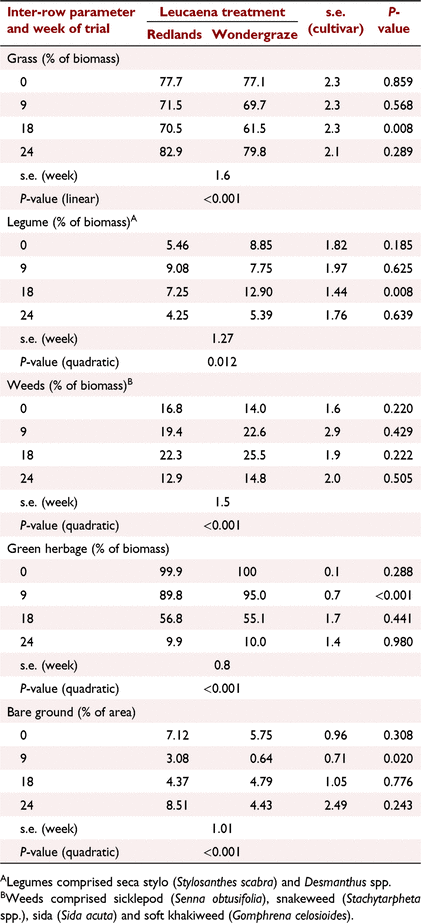
The botanical composition of pastures was similar in Redlands and Wondergraze paddocks, although there were some differences at Week 18 (Table 3), when the proportion of grasses was lower and legumes higher in Wondergraze paddocks (P < 0.01). Grasses contributed >60% of biomass in the inter-rows throughout, although this proportion declined over time (linear effect, P < 0.001). The decline in grass proportion was roughly reflected by increases in proportion of legume and weed components. However, at the conclusion of the study, legume and weed proportions had declined resulting in quadratic responses for these two variables (P < 0.01), and a concomitant numerical increase in grass percentage.
|
At the beginning of the study, pasture was actively growing with almost 100% recorded as green material. As time progressed, this proportion decreased, eventually falling to 10% of the biomass at the conclusion of the study (quadratic effect, P < 0.001). Groundcover was good throughout the study, there being <10% bare ground (Table 3).
Effects on nutritive value and animal performance
Table 4 presents the nutritive value of pasture and leucaena estimated by NIR analysis of plant material. Nutritive value of the two leucaena cultivars was similar. However, as the grazing season progressed, there were small perturbations in nutritive value. CP content varied between ~20% and 25%, being higher mid-season than at the beginning and end (quadratic effect, P < 0.001). DM digestibility was consistently >65% and OM digestibility >60%, with both measures declining over time (P < 0.01), although values were numerically higher at the last sampling week. NDF and ADF concentrations both followed quadratic responses, with values being higher mid-season (P < 0.001). There were cultivar differences at various sampling times for the fibre fractions. ADF concentration was higher in Wondergraze than Redlands early in the season but lower at the end (P < 0.01), whereas end-of-season NDF was higher in Wondergraze than Redlands (P < 0.01). As expected, nutritive value of pasture was much lower than of leucaena. CP content varied between 6% and 10% (quadratic effect, P < 0.001). Digestibility was consistent up to Week 18, but declined markedly thereafter with linear (OM) and quadratic (DM) effects (P < 0.001). Fibre concentrations also remained constant until Week 18, thereafter increasing (quadratic effects, P < 0.001).

|
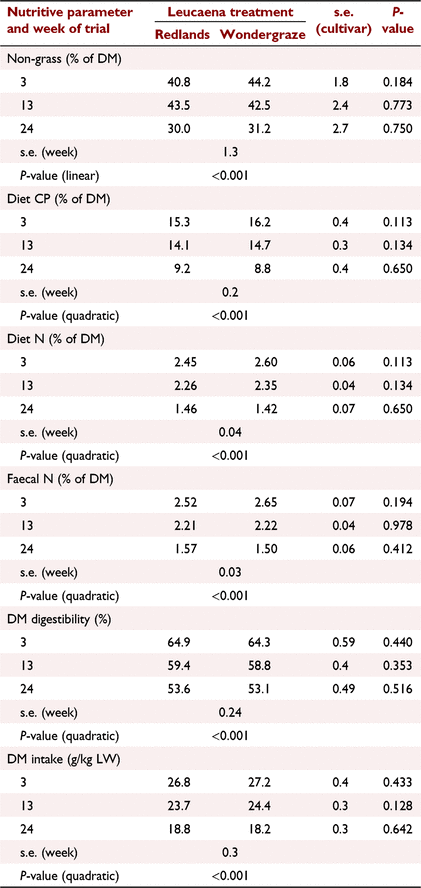
Nutritive value of the consumed diet was quantified by NIR analysis of faecal samples (Table 5). There were no cultivar effects for any observed variable (P > 0.05). However, seasonal effects were apparent suggesting a marked reduction in nutritive value of the diet in the latter half of the study (quadratic effects for all variables, P < 0.001). For example, dietary CP dropped from >14% to <10%, and DM digestibility dropped by >10 percentage units. The proportion of the diet characterised as non-grass according to δ13C ratios was >40% but declined linearly (P < 0.001) as the season progressed to ~30%.
|
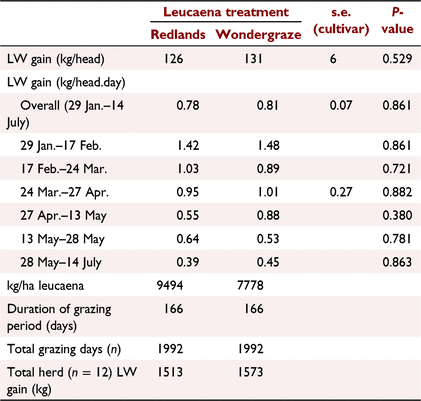
Liveweight gain was not significantly different (P > 0.05) between cultivar treatments, averaging ~0.8 kg/day over the 24 weeks of the study (Table 6). However, Fig. 1 and Table 6 show a marked decline in rate of gain as the study progressed for cattle grazing both cultivars. This decline in LW gain was consistent with the changes in nutritive value and availability of leucaena.
|

|
Effects on rumen fermentation
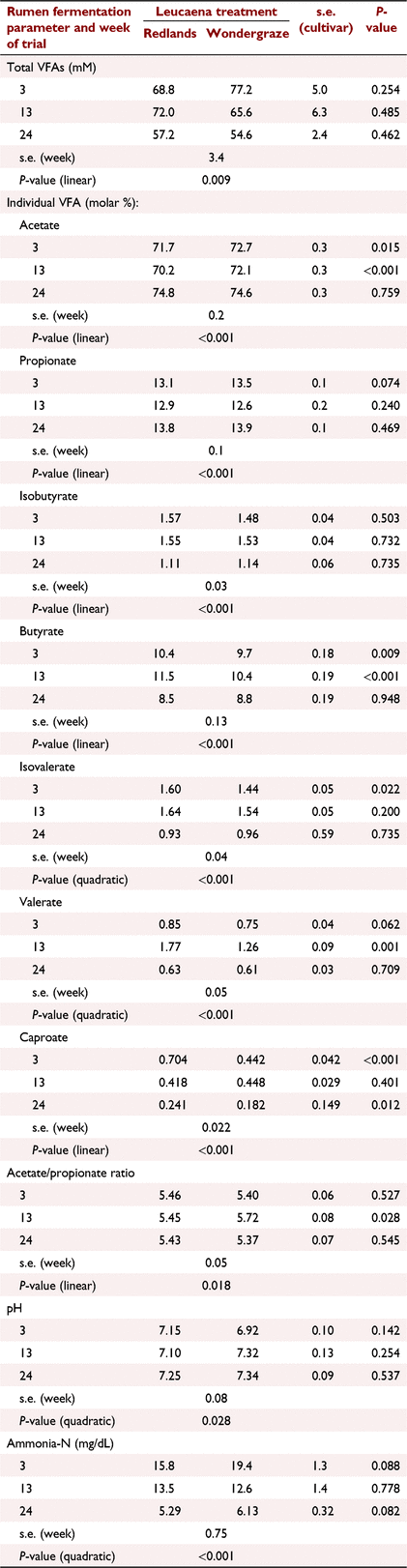
Effects on rumen fermentation were numerically small, reflective of the similar nutritive value of the diets (Table 7). Nevertheless, owing to low variability in the samples, significant cultivar differences were frequently observed for individual volatile fatty acids (VFAs) (e.g. acetate, butyrate, caproate). Consistent with the changes in nutritive value over time, total VFAs were lower at the end of the study than earlier (quadratic effect, P < 0.01). Acetate/propionate ratios were ~5.4, and pH was mostly >7, both results consistent with a forage diet. Rumen ammonia-N concentration was initially high and dropped markedly in concert with the changes in dietary N intake (quadratic effect, P < 0.001).
|
Effect on mimosine metabolism
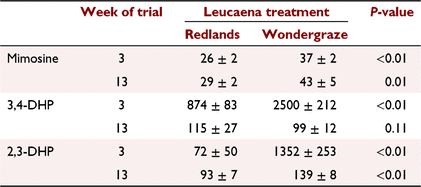
The average concentrations of mimosine and 3,4-DHP in leucaena were 5.38 ± 0.57 and 0.47 ± 0.05 g/kg DM, respectively, for Wondergraze, and 6.34 ± 0.62 and 0.20 ± 0.04 g/kg DM for Redlands. Mimosine, 3,4-DHP and 2,3-DHP concentrations are presented for non-hydrolysed rumen fluid (Table 8), and non-hydrolysed and hydrolysed urine (Table 9). In the rumen fluid, there was a clear cultivar effect, with concentrations of mimosine and DHP generally higher for cattle grazing Wondergraze than Redlands (P ≤ 0.01), except in the case of 3,4-DHP at Week 13, which was not different between treatments (P > 0.1). At Week 3, the predominant isomer of DHP in the rumen was 3,4-DHP, especially in Redlands; by Week 13, concentrations of 3,4 DHP had dropped and were broadly similar to 2,3-DHP.
|

|
Mimosine concentration in non-hydrolysed urine was high (187 µM) at Week 3 following introduction to leucaena but by Week 13 concentrations were very low (5 µM). Cultivar effects were negligible, although statistically significant at Week 13 (P = 0.02). The concentration of 3,4-DHP was similar for both hydrolysed and non-hydrolysed urine, but at Week 3, the respective concentrations were significantly higher in cattle grazing Redlands than Wondergraze (P ≤ 0.03; P < 0.01, respectively). There was no cultivar effect at Week 13, except for 2,3 DHP in hydrolysed urine, which was higher with Wondergraze than Redlands. Hydrolysis resulted in a ~30-fold increase in the concentration of 2,3-DHP at both Weeks 3 and 13, indicating that most 2,3-DHP was in the conjugated form. In the non-hydrolysed analytes, 2,3 DHP concentrations were higher with Redlands than Wondergraze at Week 3 (P < 0.01) but not at Week 13. In the hydrolysed analytes, the opposite was true, with concentrations of 2,3-DHP being higher at Week 13 with Wondergraze (P = 0.03). The concentration of 2,3-DHP present in hydrolysed urine relative to 3,4-DHP increased from ~2–4-fold at Week 3 to 7–8-fold at Week 13, indicating greater microbial conversion of 3,4 DHP to 2,3 DHP over time.
Discussion
Nutritive value of leucaena
Leucaena is recognised for its high protein content and ability to provide quality forage during the dry season typical of northern Australia. In the present study, harvested leaves and green stems <10 mm averaged >20% CP throughout the grazing study for both cultivars, which is within the expected CP range for leucaena. In an international review of leucaena nutritive value, Garcia et al. (1996) found a range of CP content from 10% to 30% of DM in leaves and petioles. In a more recent review, De Angelis et al. (2021) quoted average CP of leucaena leaves and seeds to be 24% and 31%, respectively, whereas Feedipedia (http://www.feedipedia.org/node/282) quotes CP of leucaena pods to vary between 21% and 31% of DM. Under field conditions in the present study where cattle were continuously browsing leucaena, there was no decline in CP content over the grazing period. However, Figueredo et al. (2019) observed a decline in CP content of leucaena leaves and petioles with advancing maturity from 22% at Day 30 of regrowth to 12% at Day 90 of regrowth. The lack of effect on CP content with time in our study was probably due to the continuous removal of leaves through browsing and production of new growth. Prior to this grazing study, the leucaena stands were used in a cut-and-carry experiment with indoor cattle. In that study conducted in 2019, 2 years prior to this study, the mean CP concentration was only 14.8% of DM (Stifkens et al. 2022). Variation in CP content of leucaena, although influenced by maturity (Figueredo et al. 2019), is also influenced by the harvest method and the proportions of leaf, stem and seed pods included in the sample. We concluded that where the harvesting techniques require large amounts of leucaena, there is a tendency to include more stems than when taking small samples by hand harvesting as in the present study.
Concentrations of ADF and NDF were within the expected ranges, averaging 37% and 20% of DM, respectively. However, some authors have observed lower hemicellulose (NDF – ADF) concentration (De Angelis et al. 2021; Stifkens et al. 2022). Variation over time was minimal, with a general observation of higher concentrations mid-grazing. Although this was significant, it was not expected to influence animal performance. Figueredo et al. (2019) observed increases in fibre concentration as the regrowth period increased from 30 to 90 days in non-browsed leucaena. As was noted with CP, it is apparent that continuous removal of edible leucaena through browsing maintains nutritive value.
Animal performance responses
Overall, animal performance was good, with LW gain averaging ~0.8 kg/day over 166 days. However, it was clear that as the availability of leucaena declined in the paddock, so too did the daily rates of gain. Average LW gain up to Week 12 was double the average LW gain between Weeks 12 and 23. If leucaena availability had not been compromised, it is reasonable to assume that gains in the order of 0.9–1.0 kg/day could have been sustained throughout the study because these were observed in the first month of grazing when leucaena was in abundance. Similar results have been reported by Quirk et al. (1990), who observed increased seasonal LW gain in response to the amount of leucaena on offer, particularly in the early growing season. Dixon and Coates (2008), in three sequential grazing studies each over 8–10 months, observed LW gains of 0–1 kg/day. This variation was attributed to changes in the proportion of leucaena in the diet, which varied between <20% and >80%. Mean percentage of leucaena in the diet was between 50% and 60% over the 3 years of study.
We used the same method as Dixon and Coates (2008) to estimate C3/C4 ratios. δ13C indicated that non-grass (in this case mostly leucaena plus stylos and desmanthus in the pasture) contributed >40% of the diet over the first half of the grazing period, dropping to 30% at the end of the season. These levels of legume intake are lower than those seen in earlier studies where leucaena was established on 2–4 m spacing between rows, compared with 12 m in our study (Petty et al. 1998; Dixon and Coates 2008; Graham et al. 2013).
If the CP content of the diet and the dietary components (in this case, pasture and leucaena) are known, it is possible to estimate the component contributions to the diet. Using this method, legumes comprised 60%, 30% and 15% of the diet at the beginning, mid-point and end of the study, respectively. Although these data do not closely agree with the δ13C approach (mean of 39% vs 35% CP method), they nevertheless support the assumption that diet quality declined over time. However, edible leucaena comprised only 10–0.2% of available biomass, indicating marked preference for leucaena over pasture throughout the grazing period, which resulted in leucaena being ‘grazed out’. Thus, the paddocks were no longer able to support the initial high rates of gain observed in the early months of the study. Similar observations have been made by others (Dixon and Coates 2008; Lemin et al. 2019).
Liveweight gain per ha over 6 months in the present study was ~400 kg/ha, which was higher than that achieved by Dixon and Coates (2008) over 10–11 months (124–187 kg/ha) and Bowen et al. (2018) over 12 months (198 kg/ha). Because leucaena growth is markedly seasonal, leucaena availability over long-term continuous grazing varies, as shown by Quirk et al. (1990), Petty et al. (1998) and Dixon and Coates (2008). Thus, higher gains per ha in our study can be attributed to the study terminating as soon as leucaena availability dropped to negligible levels (<15 kg DM edible material/paddock ha). By extrapolating to a year, we can assume that gains per ha would have been similar to other published data (Dixon and Coates 2008; Harrison et al. 2015; Bowen et al. 2018). Managing year-round availability of leucaena in the paddock at levels consistent with gains of ~1 kg/day is difficult under continuous grazing. Closer row spacing to increase leucaena yield per ha may sustain longer periods of good (>0.7 kg/day) animal production (e.g. Petty et al. 1998) but results in wide variation in leucaena intake and diet quality (e.g. Dixon and Coates 2008). Wider spacing, as in the present study, shortens the period during which leucaena is in sufficient supply to maintain good rates of gain (Lemin et al. 2019). Rotational grazing and tactical irrigation offer management options that should sustain annual high rates of LW gain, although these measures will not completely alleviate the problem.
The faecal NIR method allowed for estimation of feed:gain ratio (kg DM intake:kg LW gain). By employing the mean DM intake across three sampling periods and the mean LW gain to estimate feed:gain ratio, mean values of 11.3 were observed for both cultivars, which was better than the values of 17 observed when 36–48% leucaena was fed with poor-quality Rhodes grass indoors (Stifkens et al. 2022). Although the present trial did not allow for accurate estimation of intake, it is still clear that the higher quality of the non-legume portion of the diet in this study had a positive effect on feed:gain ratio.
Rumen fermentation
Rumen fermentation was characteristic of cattle consuming forage diets. The pH was generally >7, which was consistent with a high acetate:propionate ratio. Although there were some statistical differences between cattle fed the two cultivars, these were considered not to be biologically important. In trials where contrasting levels of leucaena are fed, higher proportions of leucaena can result in increased VFA concentration (Rira et al. 2015; Stifkens et al. 2022) or have no effect (Montoya-Flores et al. 2020). The same three authors have observed little effect on the proportions of VFA. McSweeney and Tomkins (2015) measured rumen VFA in cattle grazing Rhodes grass or Rhodes–leucaena mixtures. They observed lower acetate, higher longer chain VFAs and higher branch-chain FAs when cattle grazed leucaena pastures. Their acetate:propionate ratios were ~6 for Rhodes grass and 5 for Rhodes grass plus leucaena. Our acetate:propionate ratio was similar, averaging 5.5.
Leucaena contains tannins (Jones and Palmer 2002), and depending on the type and concentration, they can have either a negative or positive effect on nutritive value of the diet. However, there is little evidence that tannin-rich leucaena reduces VFA concentration and diet digestibility (McSweeney et al. 1999; Rira et al. 2015) even though it may reduce methane production (Stifkens et al. 2022) and improve N-use efficiency (Montoya-Flores et al. 2020). We conclude that potential anti-nutritional effects of tannins in the leucaena were unlikely to have any major effects on intake or animal performance and would have been overwhelmed by the positive benefits of including a high-quality forage with the accompanying pasture of generally low nutritional value.
Depending on the nature and concentration of tannins in forages, they can either improve N supply to the small intestine through increasing rumen undegraded protein or decrease N availability by rendering it less digestible throughout the digestive tract (Waghorn 2008). However, in the situation where leucaena (a high-CP forage) is added to a low-CP grass, the simple increase in N supply is probably more important than the solubility or degradability of N. Fig. 2 shows the N relationships over time for the forage in the paddock, faecal NIR estimates of dietary and faecal N, and rumen ammonia. Nitrogen percentages in the diet and faeces were very similar, suggesting that N digestibility was not negatively affected by tannins. Estimated N in the forage (from the %N of leucaena and pasture and the %non-grass in the diet) was much lower than in the diet in the early phase of the study, suggesting a clear preference for high-N dietary constituents, presumably leucaena. However, as leucaena availability was reduced approximately mid-way through the study, forage N and dietary N were similar. Throughout the study, %N in the diet declined, consistent with the reduced N in the forages, and this caused a reduction in rumen ammonia-N. By the end of the study, rumen N was approaching 5 mg/dL, a value considered insufficient for optimal rumen microbial protein synthesis (Satter and Slyter 1974).

|
Mimosine metabolites in rumen fluid and urine
Current theory regarding the detoxification of mimosine in ruminants suggests that many Australian cattle possess rumen bacteria (including S. jonesii) that can convert mimosine to secondary compounds which are excreted in urine (McSweeney et al. 2019; Halliday et al. 2013, 2014). There remains debate about the efficacy of the S. jonesii inoculum in an environment where casual inoculation seems to be occurring (McSweeney et al. 2019). Multiple strains of S. jonesii appear to be widely dispersed in the Australian cattle herd and could comprise indigenous organisms and those introduced from Hawaii (McSweeney et al. 2019). In the present trial we used cattle that, to our knowledge, had not grazed leucaena previously. However, they may have been colonised at birth with S. jonesii or casually acquired S. jonesii or other detoxifying bacteria. The first urine sampling, a week after entering the leucaena paddock and immediately prior to inoculation with S. jonesii, showed 2,3-DHP, indicating that the rumen was already colonised with S. jonesii.
The concentrations of mimosine and DHP isomers in rumen fluid are influenced by the concentration of mimosine in the leucaena, the intake of leucaena, and the rate of conversion of mimosine to 3,4-DHP and then conversion to 2,3-DHP by S. jonesii before degradation of the pyridine ring. The average levels of mimosine and DHP in plant samples of Wondergraze and Redlands were relatively similar, but rumen DHP concentration was substantially higher for the Wondergraze treatment at the Week 3 sampling, whereas similar concentrations of mimosine and DHP isomers were observed in the rumen at Week 13 for both leucaena cultivar treatments. This probably indicates that animals initially consumed Wondergraze more readily, but as the trial progressed, intakes of both varieties were similar. At the Week 3 sampling, the very high ratio of 2,3- to 3,4-DHP in hydrolysed urine for the Wondergraze treatment indicated that rumen microbial adaptation to the plant and conversion of 3,4- to 2,3-DHP by S. jonesii occurred sooner than for Redlands.
The primary metabolite of mimosine is 3,4-DHP, so it was predicted that concentrations of 3,4-DHP would be higher of 2,3-DHP shortly after exposure to leucaena, which was confirmed in rumen samples. However, when considering the hydrolysed isomers in urine, it was clear that concentrations of 2,3-DHP were much higher than 3,4-DHP at both Weeks 3 and 13 of exposure to leucaena, which probably indicates that the conjugated 2,3-DHP is excreted more rapidly than 3,4-DHP. The results suggest that there was conversion of 3,4-DHP to 2,3-DHP in the rumen soon after introduction of leucaena, and the rate of conversion in the following weeks increased as the rumen adapted to the leucaena. Hydrolysis of urine resulted in >20-fold increases in detection of 2,3-DHP, whereas 3,4-DHP concentrations did not change, indicating that following absorption into the bloodstream, 2,3-DHP was then conjugated in the liver, potentially rendering it less toxic while 3,4-DHP was excreted in an unconjugated toxic form.
The initial levels of non-hydrolysed 3,4-DHP were below the threshold level of 100 µg/mL for toxicity (Dalzell et al. 2012) and after 13 weeks of leucaena exposure, concentrations of non-hydrolysed 3,4-DHP had declined >6-fold and no overt signs of toxicity were detected.
These results support the findings of Halliday et al. (2013) that conversion of 3,4-DHP to 2,3-DHP occurs soon after exposure to leucaena, but the rate of this microbial transformation increases over several weeks. Furthermore, the majority of excreted 2,3-DHP is in a conjugated form whereas 3,4-DHP is primarily removed in the urine unconjugated. These observations suggest that the detoxification process for mimosine involves both microbial degradation in the gut and conjugation of the 2,3-DHP isomer in the liver.
Conclusions
The main finding of this study was that the psyllid-resistant cultivar, Redlands, can produce levels of animal performance equal to those achieved with Wondergraze, an older commercial cultivar. In this study, LW gain averaged 0.8 kg/day. Differences between the cultivars were small or non-existent, regardless of which parameter was compared. Early in the grazing period, rates of gain were very high for both treatments, but they declined rapidly as the availability of leucaena declined, even as pasture availability continued to increase well into the study. This highlights a problem with achieving the appropriate balance between leucaena and grass across the season. With 12-m spacing between leucaena rows, it is essential to have good establishment, which was not achieved in the present study. Further, set stocking, although useful for experimental purposes, may not be ideal for commercial management. Matching stock numbers to leucaena availability throughout the season will lead to sustained performance over longer periods. Analysis of mimosine and its breakdown products in rumen fluid and urine indicated that there were no cultivar differences in metabolism of mimosine, and no signs of toxicity were seen in any cattle.
Data availability
The data used to generate the results in the paper will be shared upon reasonable request to the corresponding author.
Conflicts of interest
Ed Charmley is an Associate Editor of Animal Production Science but was blinded from the peer-review process for this paper. The authors declare no other conflicts of interest.
Declaration of funding
Funding for this research was provided by Meat & Livestock Australia Contract B.GBP.0026.
Acknowledgements
Animal husbandry and technical support were provided by Greg Bishop-Hurley, Wayne Flintham, Steve Austin, Mel Matthews and Elizabeth Hulm. Supply of Redlands seed was kindly organised by Max Shelton, University of Queensland. Advice on leucaena establishment and agronomy and on supply of the Synergistes jonesii rumen inoculant was provided by staff of Queensland Department of Agriculture and Fisheries including Diane Ouwerkerk, Kendrick Cox and Craig Lemin.
References
Andrew MH, Noble IR, Lange RT (1979) A non-destructive method for estimating the weight of forage on shrubs. The Australian Rangeland Journal 1, 225–231.| A non-destructive method for estimating the weight of forage on shrubs.Crossref | GoogleScholarGoogle Scholar |
Beutel TS, Corbet DH, Hoffmann MB, Buck SR, Kienzle M (2018) Quantifying leucaena cultivation extent on grazing land. The Rangeland Journal 40, 31–38.
| Quantifying leucaena cultivation extent on grazing land.Crossref | GoogleScholarGoogle Scholar |
Bowen MK, Chudleigh F, Buck S, Hopkins K (2018) Productivity and profitability of forage options for beef production in the subtropics of northern Australia. Animal Production Science 58, 332–342.
| Productivity and profitability of forage options for beef production in the subtropics of northern Australia.Crossref | GoogleScholarGoogle Scholar |
Coates DB, Dixon RM (2008) Development of near infrared analysis of faeces to estimate non-grass proportons in diets selected by cattle grazing tropical pastures. Journal of Near Infrared Spectroscopy 16, 471–480.
| Development of near infrared analysis of faeces to estimate non-grass proportons in diets selected by cattle grazing tropical pastures.Crossref | GoogleScholarGoogle Scholar |
Dalzell SA (2019) Leucaena cultivars: current releases and future opportunities. Tropical Grasslands-Forrajes Tropicales 7, 56–64.
| Leucaena cultivars: current releases and future opportunities.Crossref | GoogleScholarGoogle Scholar |
Dalzell S, Shelton M, Mullen B, McLaughlan K (2006) ‘Leucaena: a guide to establishment and management.’ (Meat and Livestock Australia and The University of Queensland)
Dalzell SA, Burnett DJ, Dowsett JE, Forbes VE, Shelton HM (2012) Prevalence of mimosine and DHP toxicity in cattle grazing Leucaena leucocephala pastures in Queensland, Australia. Animal Production Science 52, 365–372.
| Prevalence of mimosine and DHP toxicity in cattle grazing Leucaena leucocephala pastures in Queensland, Australia.Crossref | GoogleScholarGoogle Scholar |
De Angelis A, Gasco L, Parisi G, Danieli PP (2021) A multipurpose leguminous plant for the Mediterranean countries: Leucaena leucocephala as an alternative protein source: a review. Animals 11, 2230
| A multipurpose leguminous plant for the Mediterranean countries: Leucaena leucocephala as an alternative protein source: a review.Crossref | GoogleScholarGoogle Scholar |
Dixon RM, Coates DB (2008) Diet quality and liveweight gain of steers grazing Leucaena-grass pasture estimated with faecal near infrared reflectance spectroscopy (F.NIRS). Australian Journal of Experimental Agriculture 48, 835–842.
| Diet quality and liveweight gain of steers grazing Leucaena-grass pasture estimated with faecal near infrared reflectance spectroscopy (F.NIRS).Crossref | GoogleScholarGoogle Scholar |
Figueredo ES, Rodrigues RC, Araújo RAd, Costa CdS, Santos FNdS, Silva IR, Jesus APR, Araújo JdS, Cabral LdS, Araújo IGR (2019) Maturity dependent variation in composition and characteristics of potentially digestible tissues of leucaena. Semina: Ciências Agrárias 40, 3133–3142.
| Maturity dependent variation in composition and characteristics of potentially digestible tissues of leucaena.Crossref | GoogleScholarGoogle Scholar |
Garcia GW, Ferguson TU, Neckles FA, Archibald KAE (1996) The nutritive value and forage productivity of Leucaena leucocephala. Animal Feed Science and Technology 60, 29–41.
| The nutritive value and forage productivity of Leucaena leucocephala.Crossref | GoogleScholarGoogle Scholar |
Graham SR, Dalzell SA, Ngu NT, Davis CK, Greenway D, McSweeney CS, Shelton HM (2013) Efficacy, persistence and presence of Synergistes jonesii in cattle grazing leucaena in Queensland: on-farm observations pre- and post-inoculation. Animal Production Science 53, 1065–1074.
| Efficacy, persistence and presence of Synergistes jonesii in cattle grazing leucaena in Queensland: on-farm observations pre- and post-inoculation.Crossref | GoogleScholarGoogle Scholar |
Halliday MJ (2017) Unravelling Leucaena leucocephala toxicity: ruminant studies in eastern Indonesia and Australia. University of Queensland.
Halliday MJ, Padmanabha J, McSweeney CS, Kerven G, Shelton HM (2013) Leucaena toxicity: a new perspective on the most widely used forage tree legume. Tropical Grasslands – Forrajes Tropicales 1, 1–11.
| Leucaena toxicity: a new perspective on the most widely used forage tree legume.Crossref | GoogleScholarGoogle Scholar |
Halliday MJ, Giles HE, Dalzell SA, McSweeney CS (2014) The efficacy of in vitro Synergistes jonesii inoculum in preventing DHP toxicity in steers fed leucaena-grass diets. Tropical Grasslands – Forrajes Tropicales 2, 68–70.
| The efficacy of in vitro Synergistes jonesii inoculum in preventing DHP toxicity in steers fed leucaena-grass diets.Crossref | GoogleScholarGoogle Scholar |
Harrison MT, McSweeney C, Tomkins NW, Eckard RJ (2015) Improving greenhouse gas emissions intensities of subtropical and tropical beef farming systems using Leucaena leucocephala. Agricultural Systems 136, 138–146.
| Improving greenhouse gas emissions intensities of subtropical and tropical beef farming systems using Leucaena leucocephala.Crossref | GoogleScholarGoogle Scholar |
Hegarty MP, Court RD, Thorne PM (1964) The determination of mimosine and 3,4-dihydroxypyridine in biological material. Australian Journal of Agricultural Research 15, 168–179.
Jones RJ, Hegarty MP (1984) The effect of different proportions of Leucaena leucocephala in the diet of cattle on growth, feed intake, thyroid function and urinary excretion of 3-hydroxy-4(1H)-pyridone. Australian Journal of Agricultural Research 35, 317–325.
| The effect of different proportions of Leucaena leucocephala in the diet of cattle on growth, feed intake, thyroid function and urinary excretion of 3-hydroxy-4(1H)-pyridone.Crossref | GoogleScholarGoogle Scholar |
Jones RJ, Palmer B (2002) Assessment of the condensed tannin concentration in a collection of Leucaena species using 14C-labelled polyethylene glycol (PEG 4000). Tropical Grasslands 36, 47–53.
Lemin C, Rolfe J, English B, Caird R, Black E, Dayes S, Cox K, Perry L, Brown G, Atkinson R, Atkinson N (2019) Tropical Grasslands – Forrajes Tropicales 7, 96–99.
| Crossref | GoogleScholarGoogle Scholar |
McSweeney CS, Tomkins N (2015) Impacts of leucaena plantations on greenhouse gas emissions in northern Australian cattle production systems. Project B.CCH.6510 Final Report. Meat & Livestock Australia, Sydney, NSW, Australia.
McSweeney CS, Palmer B, Bunch R, Krause DO (1999) In vitro quality assessment of tannin-containing tropical shrub legumes: protein and fibre digestion. Animal Feed Science and Technology 82, 227–241.
| In vitro quality assessment of tannin-containing tropical shrub legumes: protein and fibre digestion.Crossref | GoogleScholarGoogle Scholar |
McSweeney CS, Padmanabha J, Halliday MJ, Hubbard B, Dierens L, Denman S, Shelton HM (2019) Detection of Synergistes jonesii and genetic variants in ruminants from different geographical areas. Tropical Grasslands-Forrajes Tropicales 7, 154–163.
Montoya-Flores MD, Molina-Botero IC, Arango J, Romano-Muñoz JL, Solorio-Sanchez FJ, Aguilar-Pérez CF, Ku-Vera JC (2020) Effect of dried leaves of Leucaena leucocephala on rumen fermentation, rumen microbial population, and enteric methane production in crossbred heifers. Animals 10, 300
| Effect of dried leaves of Leucaena leucocephala on rumen fermentation, rumen microbial population, and enteric methane production in crossbred heifers.Crossref | GoogleScholarGoogle Scholar |
Norman HC, Wilmot MG, Thomas DT, Masters DG, Revell DK (2009) Stable carbon isotopes accurately predict diet selection by sheep fed mixtures of C3 annual pastures and saltbush or C4 perennial grasses. Livestock Science 121, 162–172.
| Stable carbon isotopes accurately predict diet selection by sheep fed mixtures of C3 annual pastures and saltbush or C4 perennial grasses.Crossref | GoogleScholarGoogle Scholar |
Petty SR, Poppi DP (2012) The liveweight gain response of heifers to supplements of molasses or maize while grazing irrigated Leucaena leucocephala/Digitaria eriantha pastures in north-west Australia. Animal Production Science 52, 619–623.
| The liveweight gain response of heifers to supplements of molasses or maize while grazing irrigated Leucaena leucocephala/Digitaria eriantha pastures in north-west Australia.Crossref | GoogleScholarGoogle Scholar |
Petty SR, Poppi DP, Triglone T (1998) Effect of maize supplementation, seasonal temperature and humidity on the liveweight gain of steers grazing irrigated Leucaena leucocephala/Digitaria eriantha pastures in north-west Australia. The Journal of Agricultural Science 130, 95–105.
| Effect of maize supplementation, seasonal temperature and humidity on the liveweight gain of steers grazing irrigated Leucaena leucocephala/Digitaria eriantha pastures in north-west Australia.Crossref | GoogleScholarGoogle Scholar |
Poppi DP, McLennan SR (2010) Nutritional research to meet future challenges. Animal Production Science 50, 329–338.
| Nutritional research to meet future challenges.Crossref | GoogleScholarGoogle Scholar |
Quirk MF, Paton CJ, Bushell JJ (1990) Increasing the amount of leucaena on offer gives faster growth rates of grazing cattle in South East Queensland. Australian Journal of Experimental Agriculture 30, 51–54.
| Increasing the amount of leucaena on offer gives faster growth rates of grazing cattle in South East Queensland.Crossref | GoogleScholarGoogle Scholar |
Radrizzani A, Dalzell SA, Shelton HM (2011) Effect of environment and plant phenology on prediction of plant nutrient deficiency using leaf analysis in Leucaena leucocephala. Crop & Pasture Science 62, 248–260.
| Effect of environment and plant phenology on prediction of plant nutrient deficiency using leaf analysis in Leucaena leucocephala.Crossref | GoogleScholarGoogle Scholar |
Rira M, Morgavi DP, Archimède H, Marie-Magdeleine C, Popova M, Bousseboua H, Doreau M (2015) Potential of tannin-rich plants for modulating ruminal microbes and ruminal fermentation in sheep. Journal of Animal Science 93, 334–347.
| Potential of tannin-rich plants for modulating ruminal microbes and ruminal fermentation in sheep.Crossref | GoogleScholarGoogle Scholar |
Satter LD, Slyter LL (1974) Effect of ammonia concentration on rumen microbial protein production in vitro. British Journal of Nutrition 32, 199–208.
| Effect of ammonia concentration on rumen microbial protein production in vitro.Crossref | GoogleScholarGoogle Scholar |
Shelton M, Dalzell S (2007) Production, economic and environmental benefits of leucaena pastures. Tropical Grasslands – Forrajes Tropicales 41, 174–190.
Shelton M, Brewbaker JL (1998) Leucaena leucocephala – the most widely used forage tree legume. In ‘Forage tree legumes in tropical agriculture’. (Eds RC Gutteridge, HM Shelton) pp. 15–29. (Tropical Grassland Society of Australia: Brisbane, Qld, Australia)
Stifkens A, Matthews EM, McSweeney CS, Charmley E (2022) Increasing the proportion of Leucaena leucocephala in hay-fed beef steers reduces methane yield. Animal Production Science
| Increasing the proportion of Leucaena leucocephala in hay-fed beef steers reduces methane yield.Crossref | GoogleScholarGoogle Scholar |
Tomkins N, Harrison M, McSweeney CS, Denman S, Charmley E, Lambrides CJ, Dalal R (2019) Greenhouse gas implications of leucaena-based pastures. Can we develop an emissions reduction methodology for the beef industry? Tropical Grasslands-Forrajes Tropicales 7, 267–272.
| Greenhouse gas implications of leucaena-based pastures. Can we develop an emissions reduction methodology for the beef industry?Crossref | GoogleScholarGoogle Scholar |
Tothill JC, Hargreaves JNG, Jones RM, McDonald CK (1992) ‘BOTANAL - a comprehensive sampling and computing procedure for estimating pasture yield and composition. 1. Field sampling.’ (CSIRO: Brisbane)
Waghorn G (2008) Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production: progress and challenges. Animal Feed Science and Technology 147, 116–139.
| Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production: progress and challenges.Crossref | GoogleScholarGoogle Scholar |