

Effects of Australian strains of infectious bronchitis virus on internal and external quality of hen eggs
K. K. Chousalkar A B and J. R. Roberts AA Animal Science, School of Rural Science and Agriculture, University of New England, Armidale, NSW 2351, Australia.
B Corresponding author. Email: kchousal@une.edu.au
Animal Production Science 49(2) 162-169 https://doi.org/10.1071/EA08167
Submitted: 30 May 2008 Accepted: 16 September 2008 Published: 20 January 2009
Abstract
The effects of two Australian strains of infectious bronchitis virus (IBV-T and N1/88) on the internal and external quality of eggs were studied in unvaccinated Isa Brown hens in full lay. Overall, there was no decline in egg production in either of the infected groups. However, there were some long-lasting negative effects on the egg internal quality of T-infected hens. Negative effects on internal quality in the N1/88-infected group were relatively short term. Yolk colour score was lower only in T-infected hens. Egg shell quality was affected only in terms of loss of egg shell colour. IBV infection resulted in paler egg shells in both the infected groups. Paler egg shells may not be regarded well by consumers. The egg shape index was lower in both infected groups.
Additional keyword: oviduct.
Introduction
In the commercial egg industry, eggs must be strong enough to withstand the additional pressures of automated collection, sorting, storage and packaging in order to reach the consumer in an adequate condition (Hamilton 1982). Also, internal and external quality of the egg is an important issue for the end consumer. In the reproductively active hen, the oviduct is a complex organ, which undergoes a series of daily cellular changes (Chousalkar and Roberts 2008).
Infectious bronchitis virus (IBV) is a ‘corona virus’ that causes respiratory disease in poultry and is one of the factors responsible for deterioration in egg production and quality. The disease is a major potential threat to the egg industry as egg quality problems currently cost millions of dollars each year. In Australia, IBV was first reported by Cumming (1962). Earlier studies were undertaken to study the effects of IBV on egg quality after infection of chickens at an early age (Crinion 1972). However, little, if any, research has been conducted to document the changes in egg quality parameters after IBV infection of hens in lay. Sevoian and Levine (1957) observed changes in internal and external egg quality after infecting adult laying hens with the Massachusetts strain of IBV. Surprisingly, since this study, little effort has been made to study the interaction of IBV with the fully functional oviduct. Australian strains of IBV are thought by some to be a cause of deterioration of egg internal quality and egg shell quality but documentation to prove or disprove this opinion is lacking. The effects of Australian strains of IBV on the oviduct of laying hens have received little attention by researchers.
Vaccination of laying hens in the rearing phase offers some protection for the oviduct (Chousalkar et al. 2007a) and for egg shell quality (Jolly et al. 2005) when laying hens are challenged with the nephropathogenic T strain of IBV. In addition, both the T strain and N1/88 strain of IBV can cause pathology in the fully functional oviduct of White Leghorn hens (Chousalkar et al. 2007b).
Both the virus strains used in this study are able to replicate in albumen-forming (Chousalkar and Roberts 2007a) and shell-forming regions (Chousalkar and Roberts 2007b) of the oviduct of Isa Brown hens.
The present study was undertaken to study the effects of IBV on egg internal quality and egg shell quality in unvaccinated laying hens challenged by the T and N1/88 strains of IBV. Strains T and N1/88 were isolated by Cumming (1962) and Ignjatovic and McWaters (1991), respectively. The N1/88 strain was isolated from a vaccinated broiler flock and is considered to be more pathogenic to the respiratory system. The T strain of IBV was isolated from cases of uraemia in 3-week-old pullets and adult laying hens and is nephropathogenic.
Materials and methods
Day-old Isa Brown hens (n = 150) were reared on the floor in poultry houses under strict isolation at the University of New England’s Armidale campus in New South Wales, Australia. At 25 weeks of age, hens were divided into three groups and transferred to individual cages in different isolation poultry houses where they remained until the termination of the experiment. Isolation and strict biosecurity were maintained at all times. Complete physical separation of the three experimental treatment groups was essential to the experimental design to prevent cross-infection among the groups. At 30 weeks of age, hens were exposed to either the T or the N1/88 strain of IBV and one group was left unchallenged as a control. The two strains of virus used were obtained from Dr Jagoda Ignatovic, CSIRO, Geelong, Australia. Each challenged bird received the virus at the dose rate of 2 × 105 50% embryo infective dose (EID50) intraocularly and the control hens were sham inoculated with normal saline. During the experimental period, each treatment group was attended by a different researcher, to avoid problems of cross-contamination but all procedures were kept constant across all treatment groups.
Egg production records were maintained daily throughout the experiment. Because it was necessary to maintain the different treatment groups in separate poultry houses, all eggs were collected at 3 and 2 weeks before challenge, to determine any inherent differences among the groups that may have been due, at least in part, to the separate housing and personnel and then daily during the week immediately before infection. Eggs were collected in the morning (0800 hours), brought to the laboratory and analysed on the same day. Eggs were then collected and analysed daily for 5 weeks after infection and again at weekly intervals of 6, 7, 8, 9 and 10 weeks after infection. All egg internal and external quality analyses were performed as described in Roberts (2004). All eggs were analysed for the internal quality parameters – albumen height, Haugh units and yolk colour score. However, the albumen height and Haugh unit data are not reported for the sixth week after infection because there was an unavoidable delay during that week between the collection of eggs and the internal quality analysis which, in itself, affected albumen viscosity in all treatment groups. Egg shell quality was measured as egg weight, shell reflectivity (indicator of lightness of shell colour), egg shell breaking strength and deformation (measured by quasistatic compression), shell weight, shell thickness and percent shell (calculated as the ratio of shell weight to egg weight expressed as a percentage). All the equipment used for egg quality analysis except egg shell thickness was supplied by Technical Services and Supply, UK. Egg shell thickness was measured by taking shell pieces from three equidistant points on the equator with shell membrane intact and measuring their thickness using a gauge constructed from a Mitutoyo Dial Comparator gauge Model 2109-10 (Kawasaki, Japan).
All hens were bled at 2 weeks before infection to check for the presence of IBV antibodies. From each hen, a 3-mL blood sample was collected from either the jugular or wing vein, into heparinised syringes and stored on ice. The blood samples were centrifuged at 4800g for 10 min at 4°C and blood plasma was collected and stored at −20°C until analysis. Plasma samples were analysed for IBV antibody titres using an IDEXX IBV antibody kit (IDEXX Laboratories, Westbrook, ME). Feed intake from 24 hens of each group was measured 1 week before infection and at weekly intervals from 1 to 4 weeks after infection. The same hens were used at all times for the feed intake measurements.
During the trial, it was noticed that the eggs from both infected groups appeared to be more elongated than the control eggs. Therefore, the length and breadth of eggs was measured weekly between 6 and 10 weeks after infection. When differences in shape index among the groups were confirmed, a follow-up trial was conducted to study the time frame of changes in length and breadth of eggs laid by infected hens. The 36 Isa Brown laying hens from the control group of the main trial, at the age of 40 weeks, were divided into three groups, 15 hens per challenge treatment and 6 hens as a control. Each challenged bird received virus at the dose rate of 2 × 105 EID50 intraocularly and the control hens were sham inoculated with normal saline. Eggs were collected daily for 3 weeks after infection. Eggs were analysed for internal and external egg quality parameters as mentioned above, up to 11 days after infection. Length and breadth were measured using a digital Vernier calliper (Mitutoyo digimatic calliper) and the shape index (breadth × 100/length) was calculated.
The experimental design imposed constraints on the statistical analysis of the data. Not all hens laid eggs every day, and two hens were removed at 2-day intervals up to day 24 after infection for studying the effects of virus on the oviduct and other tissues (Chousalkar and Roberts 2007a, 2007b, 2008). Therefore, it was not possible to apply a repeated-measures analysis to the data without exclusion of data. As a result, data were analysed by two-way factorial ANOVA and Student’s t-test were used to distinguish differences between means. Significance was assumed at P < 0.05. The experiment was conducted under approval number AEC05/082 from the University of New England Animal Ethics Committee.
Interpretation of the results of egg production and egg quality must, through necessity, bear in mind that the three treatment groups were housed in separate poultry houses. This was essential to prevent cross-contamination between the three groups. However, this housing arrangement also introduces the possibility that the poultry house, and/or the different personnel caring for each experimental group, may have influenced egg production or quality. All housing and husbandry practices were maintained the same across all treatment groups.
Results
Clinical findings
All hens were negative for IBV antibodies before infection. There were no significant effects of challenge or time in relation to challenge on egg production, although one hen from the N1/88 group and two hens from the T-infected group stopped laying during the second week after infection. Some hens from both the T- and N1/88-infected groups showed coughing and sneezing from 3 to 9 days after infection.
Feed intake
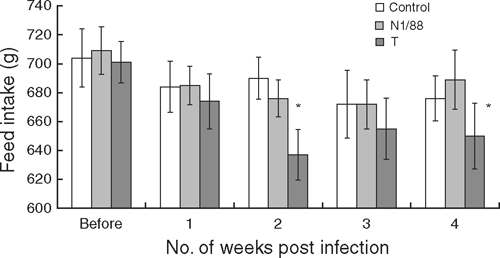
Overall, feed intake did not vary significantly over the weeks after infection or among the groups and there were no significant interactions between treatment group and time after infection. However, when data for each week were analysed separately, feed intake was significantly lower during the second and fourth weeks after infection in the T-infected group (Fig. 1).
|
Egg quality – before challenge
In eggs laid over the 3 weeks before challenge, egg weight, deformation and percent shell varied significantly, mainly as a result of increasing hen age. The only egg quality measurement for which there was a significant difference among treatment groups, before challenge, was egg breaking strength. Breaking strength of eggs laid by hens was higher in the N1/88 strain IBV group than in the control group with the T strain IBV group intermediate between the two and not significantly different from either.
Egg quality – after challenge
There were no visible deformities in egg shells in the N1/88-infected group. Eggs with yolks that separated from the albumen during egg breakout and/or with meat spots were observed mostly between the 10th and 16th day after infection in both the N1/88- and T-infected groups.
For the measures of egg shell quality, there was a significant effect of time after infection as the hens aged (Table 1). However, there was a significant effect of treatment group only for egg shell breaking strength and there were no statistically significant interactions between treatment group and time after infection for any of the measures of egg shell quality. Egg shell breaking strength after infection showed a similar pattern as that recorded before infection: it was highest in the N1/88 group and lowest in the control group, whereas the T group was intermediate.

|
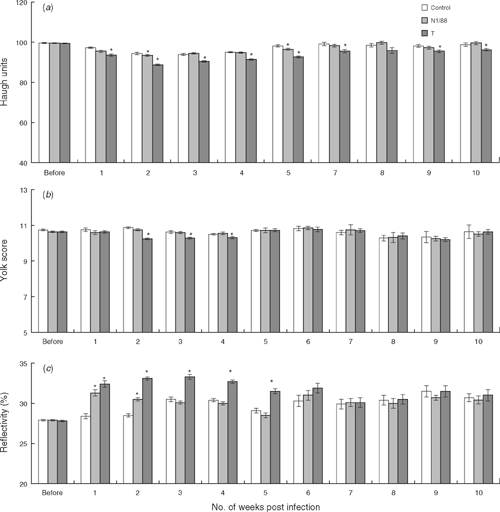
Over the 10 weeks after infection, there were significant main effects of treatment group (P < 0.001) and week after infection (P < 0.001) for albumen height and Haugh units. There were also significant interactions between treatment group and weeks after infection for albumen height (P < 0.001) and Haugh units (P < 0.033). Differences were not recorded within these parameters before challenge. When analysed on a weekly basis and compared with control hens, the albumen height in the N1/88 group was significantly lower at 2 and 5 weeks after infection and Haugh units were significantly lower at 1, and 5 weeks after infection. Except for the eighth week after infection, albumen height and Haugh units were significantly lower from the first week until the end of the experiment in the T-infected group (Fig. 2a). Overall, there were no significant differences among the treatment groups for yolk colour score but there was a significant variation over the weeks in relation to challenge (P < 0.001) and a significant interaction between group and week of experiment (P < 0.001). Yolk colour score was significantly lower in T-infected hens from 2 to 4 weeks after infection. Yolk colour score did not vary significantly among the treatment groups from 5 to 10 weeks after infection (Fig. 2b).
|
Over the 10 weeks after challenge, there were significant main effects of treatment (P < 0.001), time in relation to challenge (P < 0.001) and a significant interaction between treatment group and time after infection (P < 0.001) for shell reflectivity. As compared with the control group, shell reflectivity was significantly higher in the N1/88 group in the first and second weeks after infection, whereas in the T-infected hens reflectivity was significantly higher from 1 to 5 weeks after infection. From 6 weeks to 10 weeks after infection, no significant variation was recorded among the groups for shell reflectivity (Fig. 2c).
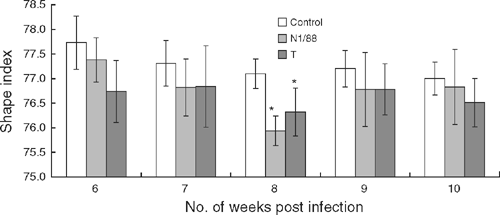
When shape index was measured from 6 to 10 weeks after infection, compared with control hens, it was significantly lower in the eggs laid by hens of the T-infected and N1/88-infected group only at 8 weeks after infection (Fig. 3).
|
Follow-up trial
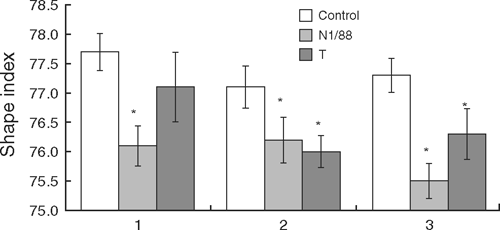
The eggs collected daily from infected hens for 3 weeks after infection, during the follow-up trial, showed the time course of change in shape index. Eggs laid by infected groups were more elongated (lower shape index) when compared with the control group. The shape index varied significantly among the groups (P < 0.009), but not over the weeks after infection and there was no interaction between groups and weeks after infection. In the N1/88-infected hens, shape index was significantly lower from the first to the third week after infection, whereas, in T-infected hens, egg shape index was significantly lower during the second and third weeks after infection (Fig. 4). Overall, egg shape index was lower in the T strain and even lower in the N1/88-infected hens when compared with the control.
|
The effects of IBV challenge on egg shell colour and albumen quality followed the same pattern during the follow-up trial as they did during the main challenge trial. Albumen quality, measured as albumen height (P < 0.001) and Haugh units (P < 0.001), was generally lower in the infected groups from 2 to 10 days after infection (Fig. 5a).
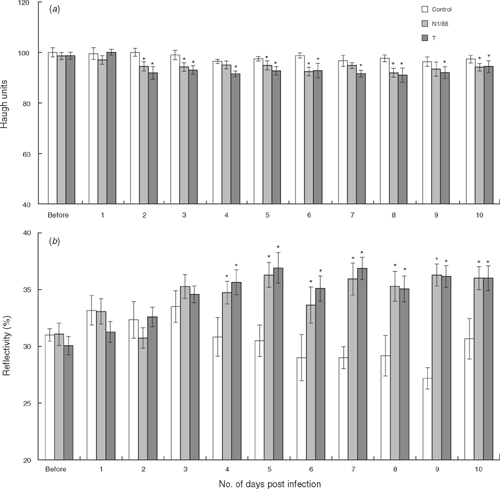
|
Shell colour, measured as shell reflectivity, was significantly lighter for the two challenge groups (P < 0.001) from 4 to 10 days after infection (Fig. 5b).
Discussion
There were no statistically significant effects of IBV infection on egg production over the 10 weeks after infection. It may be that the relatively small number of hens in each treatment group made detection of production effects of IBV more difficult. However, Cook (1971) observed a drop in egg production in a small number of hens after infecting them with the Massachusetts strain of IBV. Reduction in egg production may be an effect of the Massachusetts strain but not necessarily of other strains of IBV.
IBV clearly has major effects on albumen height and Haugh units and, except for 8 weeks after infection, these parameters in the T-infected group were significantly lower than the control from the first week after infection until the end of the experiment, indicating prolonged effects of IBV. The fluctuation in the N1/88 group could be due to selective removal of hens showing low albumen height for other studies. The hens from both the infected groups, which produced eggs with low Haugh units, were removed from the experiment at periodic intervals to study the ultrastructural pathology of IBV in different parts of the oviduct (Chousalkar and Roberts 2007a, 2007b). The finding regarding low Haugh units is consistent with results from our study of ultrastructural changes in the magnum during T and N1/88 infection and thus indicates that both of these Australian strains of IBV have tropism for the upper reproductive tract (Chousalkar and Roberts 2007a). Our findings support previous suggestions that IBV is associated with the thinning of albumen (Sevoian and Levine 1957; Butler et al. 1972). The reduction in albumen quality, which also occurred during the follow-up trial, suggests that this is a real and consistent effect of challenge with the T and N1/88 strains of IBV.
The significantly lower yolk colour score in T-infected hens from 2 to 4 weeks after infection needs further investigation as, overall, feed intake did not vary significantly among the treatment groups. This finding regarding feed intake was recorded earlier by Roberts (2005) and may be due to altered lipid utilisation. In addition, yolk colour score was not significantly affected by challenge treatment during the follow-up trial.
A reduction in shell colour as the result of IBV challenge was observed in both the main challenge trial and the follow-up trial, suggesting that this is a real and consistent effect of IBV exposure. A significant increase in shell reflectivity in the T-infected group was reported earlier by Jolly et al. (2005). However, these results were found in vaccinated HyLine Brown and HyLine Grey hens with no significant effect on vaccinated Isa Brown hens. Temporary fading of shell colour from N1/88 infection has not been reported previously but a reduction in shell colour during IBV infection with other strains has been reported (Cook and Huggins 1986). In the present experiment, reflectivity was measured using a sensitive shell reflectivity meter and pale shells were visually recorded only between 4 and 8 days after infection. Abnormal shell whitening (increase in shell reflectivity) usually occurs due to the deposition of a superficial calcareous layer on the egg shell cuticle (Hughes et al. 1986) and is observed in stress-related egg retention in the shell gland (Mills et al. 1991). The shell whitening in this study could be attributed to IBV infection-induced stress on hens. Also, porphyrin pigment imparts colour to the egg shell in brown egg-laying hens (Baird et al. 1975) and alteration in pigment deposition during IBV infection is a possible cause of egg shell whitening, although further studies are essential to prove this. Paler egg shells may not be regarded well by consumers.
Other measures of egg shell quality did not vary significantly. Cook (1971) found deterioration in egg shell quality after infecting laying hens with the Massachusetts strain of IBV. We did not observe such findings in the present study, which could be due to differences in the breed of hen or strain of IBV. The trend towards the hens in the N1/88-infected group having lower egg weight and higher breaking strength may be due to inherent differences among hens that were present before challenge, although there was no significant variation in egg weight among the treatment groups. Deformation was not different among the treatment groups, a finding that is in accordance with the study of Roberts (2005). Increase in shell weight and shell thickness towards the end of the experiment could be due to increase in egg weight as hens get older. Such findings were reported earlier in Isa Brown hens by Leary (1999). There was no obvious pattern for shell percentage.
A decrease in egg shape index with progressive increase in hen age was reported earlier by Leary (1999). However, in the present study, the hens were in their early lay period. The change in shape index in infected groups observed in the present study could be attributed to the thinner albumen providing a less firm substrate for the laying down of shell membranes and shell or to alterations in the tone of the musculature of the oviduct wall during IBV infection. However, little attention has been paid to establish the cause of change in shape index during IBV infection and further studies are necessary to clarify this effect of IBV.
In conclusion, when the hens were challenged with the T and N1/88 strains of IBV, there was deterioration in albumen quality, which occurred for a short period in N1/88-infected hens but was more prolonged in T-infected hens. This finding reflects the uterotropism of Australian strains of IBV for the fully-functional oviduct. Yolk colour score was lower than the control only in T-infected hens, which could be due to transient decreases in feed intake in T-infected hens. Egg shell quality was affected only by changes in shell colour and shape index. However, there does appear to be a significant and consistent effect of challenge with both the T and N1/88 strains of IBV causing lightening of shell colour and deterioration in albumen quality. Paler egg shells may not be regarded well by consumers.
Acknowledgement
The Physiology Teaching Unit, University of New England, provided financial support to K. Chousalkar for this study.
Baird T,
Solomon SE, Tedstone DR
(1975) Localisation and characterization of egg shell porphyrins in several avian species. British Poultry Science 16, 201–208.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |

Butler EJ,
Curtis MJ,
Pearson AW, McDougall JS
(1972) Effect of infectious bronchitis on the structure and composition of egg albumen. Journal of the Science of Food and Agriculture 23, 359–369.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Chousalkar KK, Roberts JR
(2007a) Ultra structural study of infectious bronchitis virus infection in infundibulum and magnum of commercial laying hens. Veterinary Microbiology 122, 223–236.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Chousalkar KK, Roberts JR
(2007b) Ultra structural observations on effects of infectious bronchitis virus in egg shell-forming regions of the oviduct of the commercial laying hen. Poultry Science 86, 1915–1919.
|
CAS |
PubMed |
Chousalkar KK, Roberts JR
(2008) Ultra structural changes in the oviduct of the laying hen during the laying cycle. Cell and Tissue Research 332, 349–358.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Chousalkar KK,
Roberts JR, Reece R
(2007a) Histopathology of two serotypes of infectious bronchitis virus in laying hens vaccinated in the rearing phase. Poultry Science 86, 59–62.
|
CAS |
PubMed |
Chousalkar KK,
Roberts JR, Reece R
(2007b) Comparative histopathology of two serotypes of infectious bronchitis virus (T and N1/88) in laying hens and cockerels. Poultry Science 86, 50–58.
|
CAS |
PubMed |
Cook JKA
(1971) Recovery of infectious bronchitis virus from eggs and chicks produced by experimentally inoculated hens. Journal of Comparative Pathology 81, 203–211.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Cook JKA, Huggins MB
(1986) Newly isolated serotypes of infectious bronchitis virus: their role in disease. Avian Pathology 15, 129–138.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Crinion RAP
(1972) Egg quality and production following infectious bronchitis virus exposure at one day old. Poultry Science 51, 582–585.
|
CAS |
PubMed |
Cumming RB
(1962) The etiology of ‘uraemia’ of chickens. Australian Veterinary Journal 38, 554.
| Crossref | GoogleScholarGoogle Scholar |
Hamilton RMG
(1982) Methods and factors that affect the measurement of egg shell quality. Poultry Science 61, 2022–2039.
Hughes BO,
Gilbert AB, Brown MF
(1986) Categorisation and causes of abnormal egg shells: relationship with stress. British Poultry Science 27, 325–337.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Ignjatovic J, McWaters PJ
(1991) Monoclonal antibodies to three structural proteins of avian infectious bronchitis virus: characterization of epitopes and antigenic differentiation of Australian strains. Journal of General Virology 72, 2915–2922.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
Mills AD,
Nys Y,
Gautron J, Zawadski J
(1991) Whitening of brown shelled eggs: individual variation and relationships with age fearlessness, oviposition interval and stress. British Poultry Science 32, 117–129.
| Crossref | GoogleScholarGoogle Scholar |
Roberts JR
(2004) Factors affecting egg internal quality and egg shell quality in laying hens. Japanese Poultry Science 41, 161–177.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
Sevoian M, Levine PP
(1957) Effects of infectious bronchitis on the reproductive tracts, egg production, and egg quality of laying chickens. Avian Diseases 1, 136–164.
| Crossref | GoogleScholarGoogle Scholar |