

Performance of steer progeny of sires differing in genetic potential for fatness and meat yield following postweaning growth at different rates. 2. Carcass traits
W. A. McKiernan A B , J. F. Wilkins A C F , J. Irwin A D , B. Orchard A C and S. A. Barwick A EA Cooperative Research Centre for Beef Genetic Technologies, University of New England, Armidale, NSW 2351, Australia.
B NSW Department of Primary Industries, Locked Bag 21, Orange, NSW 2800, Australia.
C NSW Department of Primary Industries, Wagga Wagga Agricultural Institute, Private Mail Bag, Wagga Wagga, NSW 2650, Australia.
D NSW Department of Primary Industries, Private Mail Bag, Yanco, NSW 2703, Australia.
E Animal Genetics and Breeding Unit, University of New England, Armidale, NSW 2351, Australia.
F Corresponding author. Email: john.wilkins@dpi.nsw.gov.au
Animal Production Science 49(6) 525-534 https://doi.org/10.1071/EA08267
Submitted: 31 October 2008 Accepted: 9 December 2008 Published: 13 May 2009
Abstract
The steer progeny of sires genetically diverse for fatness and meat yield were grown at different rates from weaning to feedlot entry and effects on growth, carcass and meat-quality traits were examined. The present paper, the second of a series, reports the effects of genetic and growth treatments on carcass traits. A total of 43 sires, within three ‘carcass class’ categories, defined as high potential for meat yield, marbling or both traits, was used. Where available, estimated breeding values for the carcass traits of retail beef yield (RBY%) and intramuscular fat (IMF%) were used in selection of the sires, which were drawn from Angus, Charolais, Limousin, Black Wagyu and Red Wagyu breeds, to provide a range of carcass sire types across the three carcass classes. Steer progeny of Hereford dams were grown at either conventional (slow: ~0.5 kg/day) or accelerated (fast: ~0.7 kg/day) rates from weaning to feedlot entry weight, with group means of ~400 kg. Accelerated and conventionally grown groups from successive calvings were managed to enter the feedlot at similar mean feedlot entry weights at the same time for the 100-day finish under identical conditions. Faster-backgrounded groups had greater fat levels in the carcass than did slower-backgrounded groups. Dressing percentages and fat colour were unaffected by growth treatment, whereas differences in ossification score and meat colour were explained by age at slaughter. There were significant effects of sire type for virtually all carcass traits measured in the progeny. Differences in hot standard carcass weight showed a clear advantage to European types, with variable outcomes for the Angus and Wagyu progeny. Sire selection by estimated breeding values (within the Angus breed) for yield and/or fat traits resulted in expected differences in the progeny for those traits. There were large differences in both meat yield and fatness among the types of greatest divergence in genetic potential for those traits, with the Black Wagyu and the Angus IMF clearly superior for IMF%, and the European types for RBY%. The Angus IMF progeny performed as well as that of the Black Wagyu for all fatness traits. Differences in RBY% among types were generally reflected by similar differences in eye muscle area. Results here provide guidelines for selecting sire types to target carcass traits for specific markets. The absence of interactions between growth and genetic treatments ensures that consistent responses can be expected across varying management and production systems.
Additional keywords: carcass quality, cattle growth path, compensatory growth, estimated breeding value, intramuscular fat, retail beef yield, sire carcass type.
Introduction
The ‘Regional Combinations’ project was a multi-site experiment designed to quantify the effects of altering growth rate in animals with diverse genetic potential for fatness and meat yield on production, carcass traits and meat quality, and thus on output and profitability in beef-production systems, in different environments across southern Australia (McKiernan et al. 2005). Beef producers in southern–central New South Wales (NSW) and Victoria have a variety of grass-finished or feeder markets to target, with the short to medium feedlot-finished product now a large part of the beef production from these regions. Experiments here examined the effects of genetic and growth treatments on carcass traits when steers were managed to reach targeted feedlot entry at approximately equal liveweights but differing in age, in a crossbred production system supplying feeder steers for 100-day grain finish. The overall design and methodology of the experiment was described in detail by McKiernan et al. (2005), and some preliminary results from the various sites have been previously reported by Wilkins et al. (2002, 2004), Tudor et al. (2004), Graham et al. (2005), McKiernan et al. (2006) and Irwin et al. (2006). Specific effects on growth and live-body composition for the NSW site have been presented in a previous paper (Wilkins et al. 2009).
Supplying slaughter cattle with consistently high compliance to targeted specifications for both the domestic and export beef trade is a major industry challenge. There is wide variation among breeds in the genetic value of bulls for various growth and carcass attributes. There is similarly large variation within breeds, but decisions can be aided by comparing estimated breeding values (EBV) for specific traits as generated by BREEDPLAN (Graser et al. 2005). It is also necessary to determine the growth pathways that best achieve targeted specifications when sires with differing potential for carcass and/or growth traits are used. Thus, selection of sires and growth regimens remains a daunting task for many producers and will be assisted by the development of decision aids facilitated by the results here.
The traits specifically examined in the present study are expressed in the carcass and include retail beef yield (RBY%) and proportion of intramuscular fat (IMF%). Although current payment systems that are essentially driven by carcass weight provide no immediate incentive to select for these traits, RBY% and IMF% are expected to be major contributors to the value of beef carcasses as the industry moves into value-based trading along with more exacting specifications of the end product. Yield of saleable meat has considerable effects on the profitability of processing (Cargill Beef Australia, pers. comm.), and fatness traits have important implications for meat quality affecting palatability (Thompson 2002). Yield and fatness, along with related traits, were important components of the design and analysis of the earlier Beef CRC studies (Bindon 2001; Upton et al. 2001). Experiments in the present study were designed to examine responses to genetic and growth-path treatments, providing the opportunity to further validate the EBV for the carcass traits of RBY% and IMF% in particular, and to assess the likelihood of genetic interactions with varying environmental scenarios (‘G × E’ effects). Understanding the effects of genetics and growth paths provides the biological basis for management strategies that will reliably achieve targeted end points.
Results from this experiment reported in a companion paper (Wilkins et al. 2009) showed clearly the effects of slower backgrounding growth rate resulting in a significant compensatory gain (+5.4% faster growth) during finishing. Animals that grew faster either during the backgrounding phase due to treatment, or while compensating during finish, had significantly greater fat deposition during both phases (8–50% increases in P8 fat depth). The ultimate effects on traits in the carcass following feedlot finishing are now examined because these determine market acceptability and value of the end product.
Materials and methods
The overall design and methodology, covering all sites in the Regional Combinations project, was described in detail by McKiernan et al. (2005). Further description of the experiment discussed in the current paper was given by Wilkins et al. (2009), which reported results up to the end of feedlotting in the live animals. We refer the reader to Wilkins et al. (2009) for details of the location and timing of the experiment, progeny generation, calving and weaning, and for a more complete description of the genetic and growth treatments. The experiment was conducted at Darlington Point (southern NSW), with all but the final feedlot intake groups run on the one property, as previously described. There were five calvings in total during 3 years (May 2001 to November 2002). Progeny were generated from matings by artificial insemination (AI), the dams all previously calved pure-bred Herefords (a self-replacing herd), and the sires as described below.
Sire genotypes
In total, 43 sires were chosen to provide wide diversity in genetic potential in their progeny for the carcass traits of IMF% and RBY%, selected on trait EBV where available, and otherwise on performance expected as a characteristic of their breed. Three carcass ‘classes’ were established, to be known as RBY, IMF and RBY&IMF, representing propensity for high retail meat yield, high marbling/IMF% (fatness) or high for both traits. Within these categories, seven sire ‘types’ were drawn from different breeds (3 Angus, 2 Euro and 2 Wagyu), with RBY drawn from Charolais (‘Char’ – 5 sires), Limousin (‘Lim’ – 4 sires) and Angus with high EBV for RBY% (‘Angus RBY’ – 9 sires); IMF drawn from Black Wagyu (4 sires) and Angus with high EBV for IMF% (Angus IMF – 8 sires); and RBY&IMF drawn from Red Wagyu (5 sires) and Angus with high EBV for both traits (Angus RBY&IMF – 8 sires). The individual sires used at the NSW and other sites were listed in the paper by McKiernan et al. (2005).
Backgrounding growth paths and finishing
The aim of the (backgrounding) growth treatments was to create divergent growth paths to a common point. Steer progeny were grown at either conventional (slow: ~0.5 kg/day) or accelerated (fast: ~0.7 kg/day) rates from weaning to feedlot entry, targeting group means of ~400 kg for feedlot entry weight. The faster growth path was achieved by supplying better nutrition; however, the result rather than the specifics of feed quality is the issue of importance here. Accelerated and conventionally grown groups from successive calvings (~5 months apart) were managed to achieve mean feedlot entry weights at the same time. Thus, we contrived groups with near-equivalent mean liveweights and with a designed age difference. Following the backgrounding period, steers were managed together in the feedlot, having identical treatment during the 100-day commercial feedlot finishing phase (at ‘Jindalee’, near Temora, NSW; Cargill Beef Australia) before slaughter. At feedlot induction, all animals were weighed (empty) and given standard health treatments and growth-promotant implants (Revalor; Intervet, Bendigo, Vic., Australia). They were weighed at exit from the feedlot (full) just before despatch to the abattoir. The overall average age at feedlot entry was 491 days (range 412–538 days for different intake cohorts) for the fast groups, and 649 days (range 573–728 days) for the slow groups. The overall average age at feedlot exit (and kill) was 595 days (range 516–642 days) for the fast groups, and 753 days (range 677–827 days) for the slow groups. Differences in these ages between carcass types within cohorts were minimal, and were accounted for in the analysis model.
Abattoir processing and carcass measurements
Cattle were processed at the Cargill Beef Australia abattoir in Wagga Wagga. Comprehensive carcass data, according to standard AUS-MEAT and Meat Standards Australia (MSA) procedures, were collected at the kill floor and in the chiller (AUS-MEAT 1996, 1998). The left side of each carcass was described by qualified MSA graders at 18–24 h after slaughter, providing feedback carcass data and input to the MSA model for predicting the eating quality of various meat cuts (see Perry et al. 2001; Thompson 2002). This is the ‘MQ4’ score referred to by those authors – we refer to this in our results as ‘predicted eating quality’ (PEQ). Carcasses were also examined with VIAscan imaging equipment (Sastek, Eagle Farm, Qld, Australia) in the chiller to estimate retail beef yield as well as other traits (Ferguson et al. 1995; Tong et al. 1999). Samples (~2.5 kg) of meat were taken from the striploin (M. longissimus lumborum) and stored frozen after 6 days ageing for later objective analyses (laboratory measured traits, Perry et al. 2001) and sensory eating quality tests (Polkinghorne et al. 1999). Objective quality traits measured in the laboratory included IMF%, shear force, compression, cooking loss, meat colour and final pH, as described by Perry et al. (2001). Consumer palatability tests (sensory eating quality) were conducted by using the MSA taste panel protocol (Polkinghorne et al. 1999). Data for the abattoir kill floor and chiller measurements, as well as the laboratory measurement of IMF% (chemical fat – by using the NIR technique described by Perry et al. 2001), are reported in the present paper. All other objective measurements and the eating-quality data will be reported in a companion paper (J. F. Wilkins, W. A. McKiernan, J. Irwin, B. Orchard and S. A. Barwick, unpubl. data).
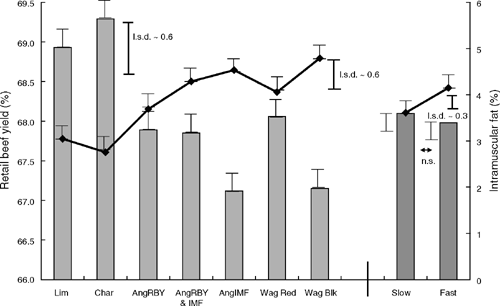
The measurements reported in the present paper are hot standard carcass weight (HSCW), dressing percentage (calculated), P8 fat depth, rib fat depth, marble scores (AUS-MEAT and USDA), eye muscle area (EMA), ossification score, meat colour, fat colour, VIAscan meat yield, predicted meat eating quality (PEQ – equivalent to ‘MQ4’ score calculated by the MSA model) and IMF% (by laboratory assay). The above chiller assessments are all part of standard abattoir procedures as referred to above.
Statistical analyses
Data were analysed with the PC software package Genstat 9 (Release 9.1 for Windows XP; Lawes Agricultural Trust, UK), by using a linear mixed model REML procedure. The procedure and models involved are described in full in Wilkins et al. (2009). HSCW was used as a covariate in the models, and significance for effects on individual carcass traits is indicated in footnotes to results tables. Significance of effects in results and discussion assumes P < 0.05 unless otherwise stated.
Results
Effects on the final carcass characteristics at slaughter
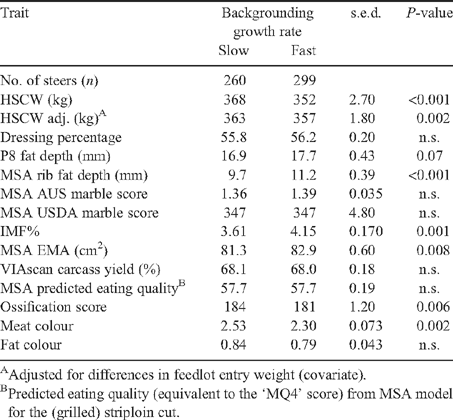
The effects of genetic (sire type) and growth treatments on carcass characteristics are shown in Tables 1–3, with results for classification on sire class shown in Table 4. The treatment responses for the main traits of interest, fatness and meat yield are illustrated in Fig. 1, showing predicted means for IMF% and RBY%. This figure provides a visually obvious demonstration of the expression of the genetic merit of the sires in the outcome measured in their progeny, which is discussed below.

|

|
|

|
|
Sire-type effects
The effects of sire type for all traits measured are shown in Tables 1 and 2. There were significant effects on HSCW, but differences were reduced when feedlot induction weight was applied as a covariate. The Red Wagyu group was significantly lighter than all others. There were also significant differences in dressing percentage in favour of the Limousin and Black Wagyu, with the Angus IMF the lowest.
The differences in both P8 and rib-fat depths among sire-type groups showed the higher-yielding European types to be lower than most other groups for P8, with the Wagyu and Angus types similar to each other. The pattern was the same for rib fat, with the ranking of the values for fat sites almost identical across the range of types.
The differences in intramuscular fat deposition, as shown in visual marble scores and laboratory assay of chemical fat, were the clearest example of the expression of genetic merit. There were dramatic differences in both marble scores and measured IMF% between the groups of highest and lowest IMF% potential (Tables 1, 2 and 4). Additionally there was a well defined ranking in the IMF% levels in particular that was consistent with the expectations on EBV within the Angus types. However, there were differences in IMF% across types that were not apparent in visual marble scores. The Black Wagyu had the highest IMF%, and although it was not significantly different from the two highest Angus types, it was clearly well above the IMF% of the other types.
The results for EMA were also consistent with the expected genetic merit. The high-yielding European types were significantly higher than all others, although most of the differences among other sire types were not significant (Table 2). These differences in EMA were reflected in the estimates of retail beef yield from VIAscan imaging. It is noted that the progeny of Angus sires selected for both RBY% and IMF% had a mean for yield equivalent to those of the RBY% only group, and a lower (non-significant) mean for IMF% than for those of the IMF% only group.
There were significant differences among types in the predictions from the MSA model of eating quality for the striploin cut. This will be discussed further in a separate paper focussed on effects on meat quality.
The significant effect of sire type on ossification score was due to the Black Waygu group having a higher score than most others.
There were no significant effects of type on either meat colour or fat colour.
Table 4 shows the results based on carcass class. HSCW and dressing percentage were not significantly affected by sire class. Differences in the subcutaneous fat depots at the P8 and rib sites were not significant, although the trend was as expected: the highest values in the IMF group and the lowest values in the RBY group.
There were significant differences in both AUS-MEAT and USDA visual marble scores as well as in the laboratory-measured IMF%, in accordance with the grouping by higher or lower marbling potential.
Differences in EMA owing to class of sire bordered on significance (P = ~0.052), with the RBY group having the highest EMA. Estimated meat yield followed the same pattern as for EMA, and in this case the yield of the IMF group was significantly lower than that of the others.
Predictions of eating quality from the MSA model were significantly better in the groups of higher IMF% or marbling potential.
Growth-treatment effects
Carcasses were heavier in the slow groups, both with and without feedlot entry weight as a covariate (Table 3). There was no significant difference in dressing percentage as a result of growth treatment.
Fast-backgrounded groups had greater fat depths, significantly for the rib site, and almost so for P8 (P = ~0.07). There was little difference owing to backgrounding growth in the visual AUS-MEAT or USDA marble scores on chiller assessment. However, the chemical IMF% measured in the laboratory was significantly greater in the fast groups. This is illustrated in Fig. 1.
EMA was significantly greater in the fast groups, even after the carcass-weight covariate adjustment.
There was no significant difference between the growth-treatment groups in retail beef yield as estimated by VIAscan imaging. The larger values for fat depths and EMA owing to faster growth counteracted each other, resulting in little absolute difference in yield.
The predictions of eating quality for the striploin cut from the MSA model did not differ between groups, with both having means in the range for ‘3 Star’ MSA rating (‘good everyday’ quality; Thompson 2002).
The ossification scores were higher for slow groups because of their greater age at slaughter. The difference of three units was significant and corresponded to a mean of 5.4 months difference in age.
The predicted mean for meat colour was higher for the slow treatment, whereas there was no significant effect on fat colour. However, the significant effect on meat colour was removed when age at kill was included as a covariate in the model to account for the difference in age between growth-treatment groups.
Treatment interactions
There were no significant interactions of growth treatment with sire carcass type for any of the carcass traits as shown in Tables 1 and 2. Thus, interactions affecting final carcass composition are unlikely, despite some inconsistencies with the ranking of subcutaneous fat depots at the end of the backgrounding period, as previously reported (Wilkins et al. 2009).
Discussion
There were no significant interactions between genetic and growth treatments for any of the carcass traits, although the interaction was close to significance for marble score, as discussed below. The few interactions that were reported previously, seen in the live composition for P8 and rib fat before feedlot entry (Wilkins et al. 2009), were small in magnitude and since they did not persist in the carcass, do not appear to be important to the final outcome of the study.
The present study focussed on how carcass traits were affected by altering growth rate in offspring of sires differing in genetic merit for IMF% or marbling and retail beef yield. Although direct payment to the producer for these traits under current pricing systems is uncommon, they are major determinants of carcass value to the processor and retailer. Studies within this and other Beef CRC projects have assumed that the industry will soon see rapid changes in the supply chain and payment systems, incorporating value-based trading, and that meat-quality traits will become increasingly important. It would no doubt be ideal if fatness and yield traits could be specifically manipulated to maintain or increase beef output while improving meat quality.
The results presented here demonstrated clearly that choosing sires on genetic merit for specific traits was effective in producing the targeted attributes in carcass traits in their progeny. Scope to alter carcass traits by nutritional management (manipulating growth rate after weaning) was also demonstrated. Although this produced fewer and smaller responses than by genetic means, it may have a large effect on enterprise profitability as discussed by Davies et al. (2009) in a companion paper.
An important issue emerging within the industry is age at slaughter. There are likely to be increased penalties in price/kg of carcass weight for animals that exceed age limits for nominated slaughter weights. This may be determined by dentition and ossification at slaughter, and an upper limit of 30 months of age is already proposed by processors (Cargill Beef Australia, pers. comm.) for the heavier-carcass markets (300–380 kg). Processing efficiency is also improved by heavier carcasses, so the message is clear that high growth rates will become increasingly important to improve profitability for both the supplier (as payment/kg for live or carcass weight) and the processor (efficiency and end-product value). Thus, it is important to know the effects of faster growth for a variety of genetically diverse animals and management systems.
Effects of growth treatments
It has been reported that faster growth resulted in increased fatness in both paddock grazing and feedlot phases of this experiment (Wilkins et al. 2009). Despite the reversal of growth rates from the backgrounding phase to the feedlot because of compensatory growth, the fast-backgrounded groups still showed significant advantage for fatness traits in the carcass reported here – in rib fat depth and IMF%, with a strong trend (P = 0.07) for P8 fat depth. As previously reported (Wilkins et al. 2009), the fast-backgrounded animals were fatter than the slow-backgrounded animals at both feedlot entry and exit, which the present study demonstrates, as expressed in the fat traits in the carcasses. However, the difference between the growth-treatment groups was smaller at exit because of the greater deposition of fat in the slow-backgrounded group in association with their faster (compensatory) growth in the feedlot. This re-enforces a conclusion of faster growth promoting greater fat deposition at any time in the postnatal growth of the animal as supported by results of others for growth at various stages before finish (Robinson et al. 2001; Cafe et al. 2006; Greenwood et al. 2006; Greenwood and Cafe 2007).
Greater fatness in faster-growing animals was consistent across most traits measured. Within all sire-types, faster growth produced greater P8 and rib fat depths and increased IMF%. However, the marble scores for the Limousin, Charolais and Black Wagyu were higher in the slower-growth groups, and it is interesting that these types produced the extremes for fatness traits (Limousin and Charolais the lowest and Black Wagyu the highest). This appears to account for the almost significant (P = 0.06) interaction of growth treatment with sire type for MSA AUS marble score (Table 2), with no significant difference between growth treatments for both measures of marble score. Marble scores are visually assessed traits that can be affected by chiller conditions and variation between operators. However, these effects should have been accounted for in the analysis model. Thus, it may be possible that the increases in IMF% resulted from fats that were of lower melting point, and therefore less likely to affect visual marbling. Monounsaturated fatty acids have a lower melting point than saturated fatty acids (Nakahashi et al. 2008), and it has also been shown that fatty-acid composition can be affected by diet, sex, tissue site and genotype (Westerling and Hendrick 1979; Oka et al. 2002; Graham et al. 2006) We have no data to describe fatty-acid profiles in the meat samples from these carcasses to support this hypothesis. However, we suggest that anomalies in marble scores may be due to fat composition, and do not truly reflect the effect of increased fat deposition with faster growth, as shown in all the other measured fatness traits. It is also possible that marble scoring, as a visual assessment, may not be accurate or sensitive enough to reflect the differences present.
As well as increased fatness, faster growth also produced increases in EMA, in agreement with results from previous CRC studies reported by Johnston et al. (2003a). This is consistent with there being no significant difference in estimated yield.
The significantly higher meat colour score due to the slow treatment is explained by the animals being older at slaughter, since the effect was removed when age at slaughter was included as a covariate in the analysis. Differences in fat colour would not be expected following the 100-day grain feeding (Cargill Beef Australia, pers. comm.), which was indeed the case.
It was concluded in the paper on live animal characteristics from this experiment (Wilkins et al. 2009) that in considering the effects of differences in growth rate at various stages of an animal’s life, backgrounding and finishing stages are the most critical when evaluated as effects in the carcass as the final outcome. The results discussed here reinforce that proposal, and are supported by the conclusions of Greenwood and Cafe (2007) and Perry and Thompson (2005). Growth differences up to weaning have relatively small effects subsequently; however, differences during backgrounding will affect later performance during finishing, and both backgrounding and finishing growth rate will have significant effects on final carcass traits. Faster growth promoting greater fatness is a consistent outcome, as also found by Graham et al. (2009).
Effects of differing genetic potential
Results here show significant effects of sire type for virtually all the carcass traits measured. There were no significant interactions between growth treatments and sire types for important production or carcass traits. Responses owing to genotype should be repeatable regardless of nutritional regime, which simplifies management advice for optimising production. Interactions were similarly absent at the other sites in the project (Graham et al. 2009; McIntyre et al. 2009) and in studies on consequences of pre-weaning growth (Greenwood et al. 2006; Greenwood and Cafe 2007). The lack of interaction of growth treatment with sire genotype for phenotypic expression of carcass traits is also in agreement with the conclusions of Johnston et al. (2003a, 2003b) and Reverter et al. (2003) in reporting the heritabilities and genotypic and phenotypic correlations found in earlier Beef CRC studies. Thus, there is strong evidence to support the conclusion that there are likely to be few, if any, interactions between backgrounding growth and genetic potential that affect carcass traits at finish for the range of growth rates observed both here and elsewhere.
All the fatness traits were significantly affected by sire type and showed clear responses to sire genetic merit for fat deposition. The results showed that Angus types with high EBV for IMF% exhibited fatness traits in the progeny equivalent to those of the progeny from the four Black Wagyu sires drawn from a breed widely considered superior for the production of highly marbled beef. This is an important issue for producers in choosing sires, although direct objective comparison is not possible until EBV for the Wagyu breed and across-breed EBV are available. However, it should also be noted that expression of fatness reported here is specific to the finishing system (100-day lot-fed) and does not predict outcomes for longer-fed situations.
The responses to sire type in EMA and retail beef yield were consistent with expectations based on genetic merit, with the two European types clearly and significantly ahead of all others. This is in agreement with a wide body of evidence in the literature reporting results from both overseas and Australian studies covering many diverse breeds (e.g. Gregory et al. 1978, 1994; Cundiff 2006; Afolayan et al. 2007). Similarly, the differences among types for dressing percentage also complied with published data included in the above.
Responses for both IMF% and RBY% within the progeny of the Angus sires were in accordance with their potential on the basis of sire EBV, as clearly shown in Fig. 1. This is a valuable demonstration to the industry of the effectiveness of using EBV-based selection, which is supported by results at the other sites (Graham et al. 2009; McIntyre et al. 2009).
Apart from the trait responses in the Angus types mentioned above, Fig. 1 also shows the general inverse relationship between IMF% and RBY% across genotypes. This is consistent with the negative genetic correlations between RYB% and fatness traits reported by Reverter et al. (2003) for both temperate and tropically adapted breeds. Despite that association, responses here in the progeny of sires chosen for high EBV for both RBY% and IMF% have demonstrated that selection for these traits need not be antagonistic. They can be improved simultaneously to increase carcass value by the choice of appropriate sires, an important outcome for industry. Multi-trait selection systems based on the value of the end product, such as BREEDOBJECT, as described by Barwick and Henzell (2005), depend on a net gain in the value of the combined effects of the individual traits. Thus, strategies based on our results will be compatible with this approach. Results also suggested that proponents of the higher-yielding European breed types could improve the potential value of carcasses in their progeny by applying some selection pressure for fatness traits in their seed-stock herds, without sacrificing their yield advantage.
Despite only small differences in PEQ, the types chosen for greater marbling potential returned better eating-quality predictions from the MSA model, as would be expected, because fatness and chiller marble score are a significant component of the model (Thompson 2002). It is noted that all carcass types achieved mean ‘3-star’ ratings (good everyday quality), even though the range in PEQ values within groups and actual values from sensory tests have wider implications, such as proportions of samples failing to meet acceptable quality. This was suggested in a preliminary report (McKiernan et al. 2006) and will be discussed more fully in a paper to follow, relating carcass traits and objective measurements of meat quality to sensory tests (J. F. Wilkins, W. A. McKiernan, J. Irwin, B. Orchard and S. A. Barwick, unpubl. data).
There were no significant effects owing to sire type for meat colour or fat colour; however, there was a significant effect on ossification score because the mean for the Black Wagyu-sired progeny was higher than that for most others. This suggests that the sires chosen are earlier-maturing types than others. However, this result was surprising, since similar differences were not seen between Angus and European types, which have considerable differences in maturity patterns (Cundiff 2006).
The grouping of sires by ‘class’ produced effects in reasonable accordance with expectations. However, such grouping may also mask important differences among types, which need to be identified. Thus, the results of type- and class-based analyses need to be assessed in combination when drawing conclusions. This was reported previously in comparing liveweights and liveweight gains among various types (Wilkins et al. 2009), and here we see some ‘dilution’ of effects in the traits of IMF% and RBY%, owing to differences among types within classes. Nevertheless, there was a strong relationship of class to response. Hence, it is suggested that selection of breed and class needs to be balanced, depending on the traits of interest.
There is little information on the Red Wagyu breed, although it has anecdotal support for potential as a dual-purpose type (for yield and marbling). There were only a very limited number of sires, and the progeny had the lowest growth rates, as previously reported (Wilkins et al. 2009), which would be a major concern if confirmed by a wider sampling of the breed. Results of the carcass traits seen here, in relation to the other sire types, showed that propensity to deposit fat was not as high as for the Black Wagyu, although clearly better than for the European types, with the reverse being the case for yield. In comparison to the Angus types, the Red Wagyu progeny were reasonably equivalent for yield and fatness traits, except for marble score, where they were equivalent to the Angus RBY but lower than the others. Thus, the progeny have displayed ‘dual purpose’ responses in carcass traits, although they had lower growth rates than other types. The progeny from these sires may not have high-enough IMF% potential to attract premiums sufficient to compensate for lower production, and as shown in a paper to follow, there also may be doubts about the eating quality of the progeny.
General effects
Results reported in Wilkins et al. (2009) clearly demonstrated compensatory gain following slower backgrounding growth. It was also suggested that the degree of compensation was related to growth potential when steers had nutritional restrictions removed, because the difference in the mean growth rate between fast- and slow-treatment groups for each type (compensatory effect) was related to the mean growth rate for the type (i.e. growth potential). The overall effect of faster growth on increasing fatness is now equally well demonstrated, although the pattern of compensation in fat deposition appears somewhat different from that for liveweight. The differences between growth-treatment groups within sire types in the means for deposition of fat during finishing showed no consistent relationship to the weight gains, nor were the patterns consistent between the P8 and rib sites. This is without explanation at present and requires analyses using the growth patterns of individual animals.
The analyses of carcass traits with carcass weight as a covariate accounted for differences that were induced by experimental design (‘all in–all out’ feedlot management) rather than by treatment response. Thus, responses in the various traits were evaluated at an equivalent carcass weight.
The data here confirm our earlier conclusion that the performance of the progeny of the different sire types was consistent with that reported previously in studies from both Australia and overseas (e.g. Gregory et al. 1978, 1994; Afolayan et al. 2007), as similarly concluded by Graham et al. (2009) for the results from the Victorian site of this project. However, caution is still recommended in making assumptions about the breeds from which these types have been drawn.
Conclusions
The effect of faster growth, resulting in greater fat deposition, was clear from the present and previous experiments, evident in both the backgrounding phase in the paddock and in subsequent feedlot finishing (Wilkins et al. 2009), and finally expressed in the carcass as reported here.
Responses in the progeny clearly reflected the genetic potential for yield and fatness traits in their sires. The absence of interactions between growth and genetic treatments ensures that consistent responses can be expected across varying management and production systems.
An outcome of great practical significance to the beef industry is the demonstration that selection for supposedly ‘antagonistic’ traits (RBY% and IMF%) can be successfully combined in the one sire to capture the benefits of both in the progeny.
Results here provide guidelines for selecting sire types to target carcass traits for specific markets. This information is important to the current development of management tools to improve profitability at all stages of the supply chain.
Acknowledgments
We are pleased to acknowledge the large contribution to the project by the support of our commercial cooperator, AgReserves Australia Ltd – sincere thanks go to all staff and management at ‘Bringagee’ and ‘Kooba’ for their enthusiastic involvement (Tony Abel and Angus Paterson in particular). We are also grateful for the cooperation and assistance of Cargill Beef Australia Ltd (Harry Waddington and Grant Garey, in particular) at the feedlot finishing (‘Jindalee’, near Temora, NSW) and processing stages (Cargill works, Wagga Wagga, NSW). Special thanks go to Matt Wolcott for his expert assistance with ultrasound measurements, to Diana Perry and staff at the Meat Science Laboratory (Armidale), and to Jeffrey House and Greg Meaker for their valuable inputs in field data collection. The authors were supported by the NSW Department of Primary Industries, the CRC for Cattle and Beef Quality and by Meat and Livestock Australia. Many thanks go to all the field and administrative support staff of NSW DPI who assisted with this large experiment and to all our colleagues within the CRC and NSW DPI who provided advice throughout. Finally, we gratefully acknowledge the inputs of Dr Jim Walkley for his helpful advice in the drafting and revision of this paper.
Afolayan RA,
Pitchford WS,
Deland MPB, McKiernan WA
(2007) Breed variation and genetic parameters for growth and body development in diverse beef cattle genotypes. Animal 1, 13–20.
| Crossref | GoogleScholarGoogle Scholar |

Barwick SA, Henzell AL
(2005) Development successes and issues for the future in deriving and applying selection indexes for beef breeding. Australian Journal of Experimental Agriculture 45, 923–933.
| Crossref | GoogleScholarGoogle Scholar |
Bindon BM
(2001) Genesis of the Cooperative Research Centre for the Cattle and Beef Industry: integration of resources for beef quality research (1993–2000). Australian Journal of Experimental Agriculture 41, 843–853.
| Crossref | GoogleScholarGoogle Scholar |
Cafe LM,
Hearnshaw H,
Hennessy DW, Greenwood PL
(2006) Growth and carcase characteristics of Wagyu-sired steers at heavy market weights following slow or rapid growth to weaning. Australian Journal of Experimental Agriculture 46, 951–956.
| Crossref | GoogleScholarGoogle Scholar |
Davies BL,
Alford AR, Griffith GR
(2009) Economic effects of alternate growth path, time of calving and breed type combinations across southern Australian beef cattle environments: feedlot finishing at the New South Wales experimental site. Animal Production Science 49, 535–541.
| Crossref | GoogleScholarGoogle Scholar |
Graham JF,
Byron J, Deland MPB
(2005) Effect on liveweight and carcass traits of progeny sired by bulls selected for extremes in intramuscular fat and retail beef yield and the effect of post weaning growth. Proceedings of the Association for the Advancement of Animal Breeding and Genetics 16, 338–341.
Graham JF,
Bernaud E, Deland MPB
(2006) Sire and dam breed effects on fatty acid profiles in the longissimus dorsi muscle and subcutaneous fat of beef cattle. Australian Journal of Experimental Agriculture 46, 913–919.
| Crossref | GoogleScholarGoogle Scholar |
Graham JF,
Byron J,
Clark AJ,
Kearney G, Orchard B
(2009) Effect of postweaning growth and bulls selected for extremes in retail beef yield and intramuscular fat on progeny liveweight and carcass traits. Animal Production Science 49, 493–503.
| Crossref | GoogleScholarGoogle Scholar |
Graser H-U,
Tier B,
Johnston DJ, Barwick SA
(2005) Genetic evaluation for the beef industry in Australia. Australian Journal of Experimental Agriculture 45, 913–921.
| Crossref | GoogleScholarGoogle Scholar |
Greenwood PL, Cafe LM
(2007) Prenatal and pre-weaning growth and nutrition of cattle: long-term consequences for beef production. Animal 19, 1283–1296.
Greenwood PL,
Cafe LM,
Hearnshaw H,
Hennessy DW,
Thompson JM, Morris SG
(2006) Long-term consequences of birth weight and growth to weaning on carcass, yield and beef quality characteristics of Piedmontese- and Wagyu-sired cattle. Australian Journal of Experimental Agriculture 46, 257–269.
| Crossref | GoogleScholarGoogle Scholar |
Gregory KE,
Cundiff LV,
Smith GM,
Laster DB, Fitzhugh HA
(1978) Characterization of biological types of cattle-Cycle II: I. Birth and weaning traits. Journal of Animal Science 47, 1022–1030.
Gregory KE,
Cundiff LV,
Koch RM,
Dikeman ME, Koohmaraie M
(1994) Breed effects, retained heterosis, and estimates of genetic and phenotypic parameters for carcass and meat traits of beef cattle. Journal of Animal Science 72, 1174–1183.
Johnston DJ,
Reverter A,
Burrow HM,
Oddy VH, Robinson DL
(2003a) Genetic and phenotypic characterisation of animal, carcass, and meat quality traits from temperate and tropically adapted beef breeds. Australian Journal of Agricultural Research 54, 107–118.
| Crossref | GoogleScholarGoogle Scholar |
Johnston DJ,
Reverter A,
Ferguson DM,
Thompson JM, Burrow HM
(2003b) Genetic and phenotypic characterisation of animal, carcass, and meat quality traits from temperate and tropically adapted beef breeds. 3. Meat quality traits. Australian Journal of Agricultural Research 54, 135–147.
| Crossref | GoogleScholarGoogle Scholar |
McIntyre BL,
Tudor GD,
Read D,
Smart W,
Della Bosca TJ,
Speijers EJ, Orchard B
(2009) Effects of growth path, sire type, calving time and sex on growth and carcass characteristics of beef cattle in the agricultural area of Western Australia. Animal Production Science 49, 504–514.
| Crossref | GoogleScholarGoogle Scholar |
McKiernan WA,
Wilkins JF,
Barwick SA,
Tudor GD,
McIntyre BL,
Graham JG,
Deland MPD, Davies L
(2005) CRC ‘Regional Combinations’ Project – effects of genetics and growth paths on beef production and meat quality: experimental design, methods and measurements. Australian Journal of Experimental Agriculture 45, 959–969.
| Crossref | GoogleScholarGoogle Scholar |
Nakahashi Y,
Maruyama S,
Seki S,
Hikada S, Kuchida K
(2008) Relationships between monounsaturated fatty acids of marbling flecks and image analysis traits in longissimus muscle for Japanese black steers. Journal of Animal Science 86, 3551–3556.
| Crossref | GoogleScholarGoogle Scholar |
Oka A,
Iwaki F,
Dohgo T,
Ohtagaki S,
Noda M,
Shiozaki T,
Endoh O, Ozaki M
(2002) Genetic effects on fatty acid composition of carcass fat of Japanese Black Wagyu steers. Journal of Animal Science 80, 1005–1011.
Perry D, Thompson JM
(2005) The effect of growth rate during backgrounding and finishing on meat quality traits in beef cattle. Meat Science 69, 691–702.
| Crossref | GoogleScholarGoogle Scholar |
Perry D,
Shorthose WR,
Ferguson DM, Thompson JM
(2001) Methods used in the CRC program for the determination of carcass yield and beef quality. Australian Journal of Experimental Agriculture 41, 953–957.
| Crossref | GoogleScholarGoogle Scholar |
Polkinghorne R,
Watson R,
Porter M,
Gee A,
Scott J, Thompson J
(1999) Meat Standards Australia, a ‘PACCP’ based beef grading scheme for consumers. 1. The use of consumer scores to set grade standards. Proceedings of the 45th International Congress of Meat Science and Technology 45, 14–15.
Reverter A,
Johnston DJ,
Perry D,
Goddard ME, Burrow HM
(2003) Genetic and phenotypic characterisation of animal, carcass, and meat quality traits from temperate and tropically adapted beef breeds. 2. Abattoir carcass traits. Australian Journal of Agricultural Research 54, 119–134.
| Crossref | GoogleScholarGoogle Scholar |
Robinson DL,
Oddy VH,
Dicker RW, McPhee M
(2001) Post-weaning growth in northern New South Wales. 3. carry-over effects on finishing, carcass characteristics and intramuscular fat. Australian Journal of Experimental Agriculture 41, 1041–1049.
| Crossref | GoogleScholarGoogle Scholar |
Thompson J
(2002) Managing meat tenderness. Meat Science 62, 295–308.
| Crossref | GoogleScholarGoogle Scholar |
Tong AKW,
Robinson DJ,
Robertson WR,
Zawadski SM, Liu T
(1999) Evaluation of the Canadian Vision System for beef carcass grading. Proceedings of the 45th International Congress of Meat Science and Technology 45, 374–375.
Tudor GD,
Della Bosca TJ,
McIntyre BL,
Read D,
Smart WL,
Taylor EG, Hiskock AJF
(2004) Optimising feed supply, reproductive efficiency and progeny growth to meet market specifications. 1. Background and experimental design. Animal Production in Australia 25, 334.
Upton W,
Burrow HM,
Dundon A,
Robinson DL, Farrell EB
(2001) CRC breeding program design, measurements and database: methods that underpin CRC research results. Australian Journal of Experimental Agriculture 41, 943–952.
| Crossref | GoogleScholarGoogle Scholar |
Westerling DB, Hendrick HB
(1979) Fatty acid composition of bovine lipids as influenced by diet, sex and anatomical location and relationship to sensory characteristics. Journal of Animal Science 48, 1343–1348.
Wilkins JF,
Irwin J,
McKiernan WA, Barwick SA
(2002) Combining genetics and growth rate to produce high quality beef in south eastern Australia. Animal Production in Australia 24, 370.
Wilkins JF,
Irwin J,
McKiernan WA, Barwick SA
(2004) Effects of altering growth rate on carcass traits in a range of beef genotypes that vary in potential for yield and fat deposition. Animal Production in Australia 25, 337.
Wilkins JF,
McKiernan WA,
Irwin J,
Orchard B, Barwick SA
(2009) Performance of steer progeny of sires differing in genetic potential for fatness and meat yield following post-weaning growth at different rates. 1. Growth and live-animal composition. Animal Production Science 49, 515–524.
| Crossref | GoogleScholarGoogle Scholar |