

Fine-scale spatial and seasonal partitioning among large sharks and other elasmobranchs in south-eastern Queensland, Australia
Stephen Taylor A C , Wayne Sumpton B and Tony Ham AA Fisheries Queensland, Department of Employment, Economic Development and Innovation, Primary Industries Building, 80 Ann Street, Brisbane, Qld 4001, Australia.
B AgriScience Queensland, Department of Employment, Economic Development and Innovation, Ecosciences Precinct, 41 Boggo Road, Park, Qld 4102, Australia.
C Corresponding author. Email: stephen.taylor@deedi.qld.gov.au
Marine and Freshwater Research 62(6) 638-647 https://doi.org/10.1071/MF10154
Submitted: 18 June 2010 Accepted: 2 April 2011 Published: 24 June 2011
Journal Compilation © CSIRO Publishing 2011 Open Access CC BY-NC-ND
Abstract
Our understanding of the ecological role of larger elasmobranchs is limited by a lack of information on their spatial and seasonal abundance. Analysis of 14 years of gill-net catch data in south-eastern Queensland, Australia, revealed that the species composition of large sharks and other elasmobranchs significantly differed among beaches and seasons. Spinner sharks (Carcharhinus brevipinna) and hammerhead sharks (Sphyrna spp.) comprised nearly half the catch of all elasmobranchs. Although the distribution of these sharks overlapped, spatial variation existed in their abundance. Spinner sharks characterised the catch at Sunshine Coast beaches, whereas the catch at Gold Coast beaches was dominated by hammerhead sharks. Seasonal differences in elasmobranch community structure were also apparent, driven largely by a lower abundance of many species during the winter and the predominance of species such as spinner sharks and hammerheads in spring and summer. The present study provides the first quantitative data for numerous species of Carcharhiniformes in south-eastern Queensland and demonstrates that analysis of catch-rate data can improve our understanding of how larger sharks partition resources.
Additional keywords: community structure, hammerhead shark, partitioning, shark-control program, spatial patterns, spinner shark, temporal patterns.
Introduction
Despite the ecological significance of large sharks (Heithaus et al. 2010) and their importance to both commercial and recreational fisheries (Walker 1998), quantitative data on their spatial and seasonal abundance are scarce. Studies that have examined spatial and temporal partitioning among elasmobranchs have typically focussed on juvenile sharks or smaller sharks in shallow-water habitats (Simpfendorfer and Milward 1993; Speed et al. 2010). Elevated catches of juvenile sharks are often reported in subtropical waters during the summer, which is linked to the nursery role that inshore waters provide (Castro 1993; Knip et al. 2010) and the role that water temperature appears to play in the onset and conclusion of shark pupping and nursery seasons (Pratt and Carrier 2001).
While quantitative data on smaller sharks in coastal waters have assisted management, the maintenance of a sustainable shark population also requires information on larger, older individuals outside of nursery areas (Kinney and Simpfendorfer 2009). Movements of larger sharks have been assessed through acoustic tracking (Simpfendorfer and Heupel 2004), and developments in passive-monitoring technology (Heupel et al. 2006) have led to more studies focussing on habitat use among sharks. However, these approaches are expensive and appropriate only for tracking individual animals or relatively small numbers of the population. Relative catch rates using gill-nets and longlines can provide useful information on population-level differences in the abundance and species composition of elasmobranchs (Simpfendorfer and Heupel 2004), although capturing larger sharks in gill-nets and longlines is logistically difficult. As such, many scientific studies that have used gill-nets to catch sharks have not provided reliable estimates of abundance for larger sharks because they used mesh sizes that tended to select for smaller sharks.
The long-term nature of shark-control programs (Dudley 1997), and the fact that they use specialist gear which selects for larger sharks, make them a suitable source for inferring the abundance of large sharks in inshore waters. Annual, seasonal and spatial trends in large sharks have been reported from bather-protection nets in KwaZulu-Natal, South Africa (Cliff and Dudley 1991; de Bruyn et al. 2005; Dudley and Simpfendorfer 2006). For example, analysis of KwaZulu-Natal catch data from 1978 to 2003 examined the catch rate and size of 14 commonly caught shark species. The analysis revealed that the catch rate of bull sharks (Carcharhinus leucas), common blacktips (C. limbatus), scalloped hammerheads (Sphyrna lewini) and great hammerheads (S. mokarran) showed a significant decline (Dudley and Simpfendorfer 2006).
In the Hawaiian Islands, analysis of shark-control program data suggested that interspecific competition influenced the distribution of carcharhinid sharks throughout the Hawaiian Archipelago (Papastamatiou et al. 2006). Tiger sharks (Galeocerdo cuvier) and Galapagos sharks (C. galapagensis) were caught at all Hawaiian Islands but were more abundant in the north-western Hawaiian Islands, whereas an inverse relationship in distribution was found between grey reef sharks (C. amblyrhynchos) and sandbar sharks (C. plumbeus) (Papastamatiou et al. 2006). In New South Wales (NSW), Australia, differences in the species composition of larger sharks were observed from netted beaches in the NSW Shark Meshing Program from 1972 to 1990 (Krogh 1994); however, the lack of species resolution in the dataset did not allow inferences to be made about sharks in the Carcharhinus genus, which comprised the bulk of the catch.
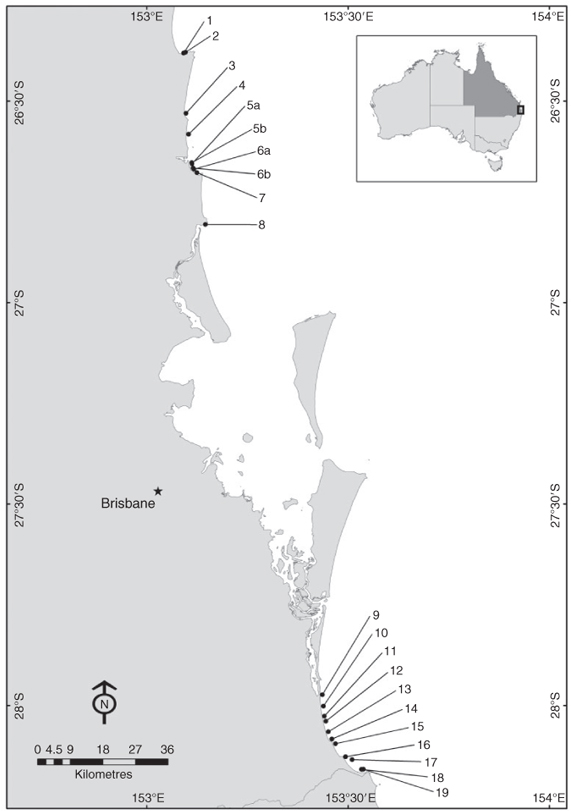
Although these studies on shark-control programs have increased our knowledge of the abundance of large sharks in inshore waters, there is still a lack of information on fine-scale spatial patterns in abundance. To address this, catch data were analysed from gill-nets used in the Queensland Shark Control Program (QSCP) from 1996 to 2009, near Brisbane in south-eastern Queensland. Catch data were examined from 19 netted beaches covering a distance of ∼220 km from north to south. South-eastern Queensland is an area of known high elasmobranch diversity (Kyne et al. 2005) and yet the present study is the first to provide data on resource partitioning among larger sharks within the region.
The aim of the present study was to determine whether the catch composition of elasmobranchs significantly differed among beaches and times of year (4 seasons) and to identify the elasmobranchs responsible for dissimilarity between Gold Coast and Sunshine Coast beaches and among seasons. The catch composition is compared with that in other studies on larger sharks in subtropical and tropical waters. We also report on the average size and sex ratio for the two most abundant sharks and analyse monthly trends in overall catch. Elsewhere, studies have observed spatial partitioning among sharks in inshore waters (White and Potter 2004; Pikitch et al. 2005; DeAngelis et al. 2008). Given the large number of netted beaches in the present study and the fact that environmental conditions surrounding the nets do differ, we expected spatial differences to occur in the elasmobranch assemblage, particularly between Gold Coast and Sunshine Coast beaches. Seasonal differences in the elasmobranch assemblage were also expected, on the basis of the monthly fluctuations in water temperature that occur in this and other subtropical waters.
Materials and methods
Sampling
Surface gill-nets used by the QSCP were permanently in place during the study apart from short periods of storm and cyclonic conditions during which they were sometimes lost or removed. Gill-nets were constructed from three 62-m net panels, joined end to end, with a stretched mesh size of 50 cm. Each panel was joined by a common footrope and float-line but adjacent panels were not stitched together at the two joins. Meshes were attached to a 12-mm polypropylene headline rope with 2.7-mm polypropylene cord at a two-thirds hanging ratio. The headline rope was fitted with 230-mm-long torpedo floats positioned every 2 m. The foot-rope was braided 12-mm polypropylene weighted with 250-g leads every 2 m. Nets were anchored to the seabed with 32-kg Danforth or Clyde Quick Release (CQR) anchors attached to the net bridles. Nets were set parallel to the shore in water between 5 and 12 m in depth, ∼500 m from the shore, although this distance varied from 450 to 800 m depending on local bathymetry (Table 1). Physical conditions surrounding each net were not reported for each day of sampling. They were subjectively assessed by the contractors and independently verified by one of the contractors who had worked on both coasts. Nets were checked about 15 and 20 days per month on the Sunshine Coast and Gold Coast, respectively, and were replaced and cleaned approximately every 20 days.

|
Contractors returned daily catch logs containing information on shark species and size (total length in cm), and by-catch caught. Although the program has been active since 1962, species identifications have been considered reliable only since the mid-1990s, following a training program on species identification in 1992. Before that time, many of the identifications were grouped into categories such as ‘unidentified whaler sharks’. In the present study, we chose to use the data from 1996 to 2009 because the same fishing contractor operated on the Gold Coast throughout this entire period and there was only one change (in 2007) in contractor on the Sunshine Coast. All contractors also had considerable previous experience in species identification and shark-fishing experience before becoming QSCP contractors. They were also collaborating with several elasmobranch researchers during the study who have confirmed species identifications on numerous occasions.
Environmental conditions and gear characteristics at each netted location
All nets at Gold Coast beaches had a 6-m drop (12 meshes) and were set in water of relatively constant depth (10–12 m) over sandy substrate between 600 and 700 m from the shoreline (Table 1). Nets at Sunshine Coast beaches had a varied net-drop size. Caloundra, Mooloolaba, Noosa Beach and Noosa Headland nets had a 3-m drop (6 meshes) and nets at all remaining beaches had a 6-m drop. Sunshine Coast nets were generally set in slightly shallower water (5–12 m) between 450 and 650 m from the shoreline (Table 1). With the exception of Caloundra, all Sunshine Coast nets were set over harder substrate, comprising coffee rock and sand, or over rock (boulders). Visibility at all Gold Coast beaches was clear to moderate at all times, apart from after storms. Water visibility at Sunshine Coast beaches was more varied, ranging from clear to murky. Human disturbance (i.e. number of boats, jet skis and swimmers adjacent to the net) was assessed as high for 7 of the 21 nets and was assessed as extremely high at Mooloolaba. Only three nets (Bilinga, Marcoola and Coolum) were characterised as low disturbance.
Analysis
Catch data were collated as the number of elasmobranchs in each net per month for each of the 19 netted beaches (Fig. 1). Individual species were combined into elasmobranch groups when numbers caught were low or where species identification was uncertain. The latter was the case for the Australian blacktip (C. tilstoni) and the common blacktip, which are notoriously difficult to separate in the field (Last and Stevens 1994). Although scalloped hammerheads (Spyrhna lewini) and great hammerheads were also recorded in the data, catch of hammerheads was not always reported to the species level because of difficulties in species identification. For this reason, all hammerhead sharks were grouped into one category for multivariate analysis.
|
Three locations (Noosa, Alexandra Headland and Maroochydore) each had two nets in close proximity (within 800 m of each other). For the latter two locations, the total catch of the two nets was halved because it was not possible to consistently separate individual catches of these paired nets in the database. In the case of Noosa, catch information was recorded separately in the database and so all subsequent analysis treated the two Noosa nets separately.
Analysis of the mean size and sex ratio of sharks was restricted to the two most abundant species. The hypothesis that the sex ratio of these sharks was 1 : 1 was tested with the chi-square statistic (χ2). Monthly trends in overall catch were displayed graphically for the five most abundant species. Multivariate analysis to identify spatial and seasonal patterns in the species assemblage was conducted using Primer 6.0 (Clarke and Gorley 2006). Before analyses, catch data were square root-transformed and similarity matrices were constructed by using the Bray–Curtis similarity coefficient (Clarke and Warwick 2001). A two-way crossed analysis of similarities (ANOSIM) was used to examine changes in the catch composition, with location (21 nets across 19 beaches) and time of year (4 seasons) as the two factors (Warwick et al. 1990). This test examined the following two null hypotheses: first, that there was no location effect, allowing for the fact that there may have been a seasonal effect; and second, that there was no seasonal effect, allowing for the fact that there may have been a location effect. Inter-annual variations in catch were not investigated and data from each location were pooled across all 14 years. This led to a total of 42 replicates per netted beach for each season (14 years × 3 months in each season).
Ordination of the average monthly catch per netted beach (pooled across all years) was conducted using non-metric multi-dimensional scaling (MDS, Clarke and Warwick 2001). Similarity percentages (SIMPER) were also used to determine which elasmobranchs characterised the assemblage at each coast (Gold Coast: 11 nets at 11 beaches; Sunshine Coast: 10 nets at 8 netted beaches) and which elasmobranchs were responsible for dissimilarities between the two coasts (Clarke and Warwick 2001). SIMPER was also used to determine which elasmobranchs characterised the assemblage within each season and which elasmobranchs were responsible for dissimilarities among seasons.
Results
Overall species composition
In total, 2027 elasmobranchs, 68% of which were sharks, were caught at all 19 netted beaches from 1996 to 2009. The three most abundant elasmobranchs were spinner sharks, hammerheads and Australian cownose rays (Rhinoptera neglecta), which comprised 25%, 23% and 10% of the catch, respectively (Fig. 2). Sharks from the Carcharhinus genus represented 40% of all elasmobranchs caught. In total, 35 separate species of sharks and rays were reported and were combined into 26 elasmobranch groupings for subsequent multivariate analysis.

|
The mean (s.d.) total length of spinner sharks caught during the study was 198 cm (0.61), and the median total length was 210 cm. The sex ratio of male to female spinner sharks was 0.5 : 1, which differed significantly (P = 0.0001) from 1 : 1. The mean (s.d.) total length of scalloped hammerheads caught was 179 cm (0.63), and the median total length was 160 cm. The sex ratio of male to female scalloped hammerheads was 0.9 : 1, which did not differ significantly (P = 0.43) from 1 : 1.
Spatial and temporal variations in the elasmobranch composition
Spinner sharks were the most abundant elasmobranch at Sunshine Coast beaches, comprising 21–48% of all sharks and rays caught at each netted beach (Fig. 3). Hammerheads were the second-most abundant group, comprising 5–27% of all elasmobranchs caught. Nets at Gold Coast beaches had a different species composition, with hammerheads, spinner sharks and Australian cownose rays representing 15–33%, 14–25% and 7–24%, respectively, of the elasmobranchs at each netted beach.

|
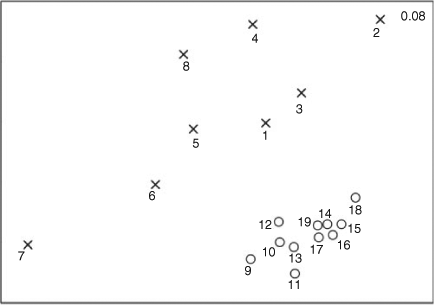
The MDS ordination plot showed that Sunshine Coast and Gold Coast beaches formed two separate non-overlapping clusters (Fig. 4). Gold Coast beaches were more tightly clustered and beaches in closer proximity tended to be closer together within the Gold Coast cluster. Sunshine Coast beaches did not form a tight cluster and the nets at Mooloolaba and Noosa Headland differed considerably from each other and the remaining six Sunshine Coast netted beaches.
|
Analysis of similarities demonstrated that the elasmobranch composition was significantly different among netted beaches (Global r = 0.09, P = 0.001) and among seasons (Global r = 0.07, P = 0.001). In both these instances, the R-statistic value was low, indicating that some similarities in the species composition occurred among beaches and seasons, respectively. The large number of beaches made it impractical to report on the significance levels for all possible pairwise comparisons; however, a significant (P < 0.05) difference in the elasmobranch composition was observed in 90 of all possible 171 pairwise comparisons. Some of these pairwise comparisons may be spurious, given there was a 1 in 20 possibility of committing a Type 1 error when α was set as 0.05. Comparisons between Sunshine Coast and Gold Coast beaches were responsible for 70% of all significant pairwise comparisons. A significant difference in the elasmobranch composition was also observed between spring and summer (Global r = 0.09, P = 0.001), spring and autumn (Global r = 0.06, P = 0.001), summer and autumn (Global r = 0.04, P = 0.001) and summer and winter (Global r = 0.13, P = 0.001).
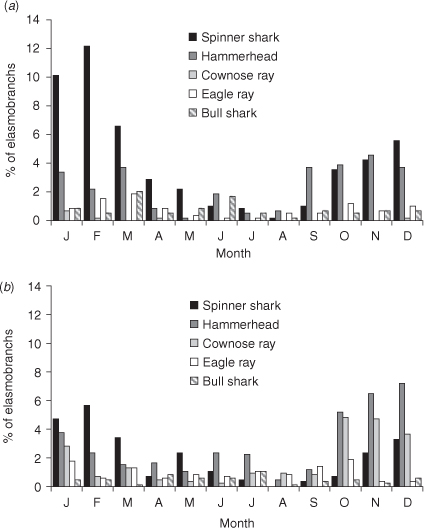
A monthly pattern of abundance was apparent for spinner sharks at both Sunshine Coast (Fig. 5a) and Gold Coast beaches (Fig. 5b), although more spinner sharks were caught at Sunshine Coast netted beaches. Combined total catch during winter (June–August) was low, representing only 5% of the total number of spinner sharks caught. Catch of spinner sharks was highest in the summer when they comprised 43% and 28% of the species assemblage at Sunshine Coast and Gold Coast beaches, respectively.
|
Monthly catch of hammerheads and their contribution towards the total elasmobranch catch differed between Sunshine Coast and Gold Coast netted beaches. At Sunshine Coast beaches catch was lower, ranging from 3 to 27 sharks per month for all years combined. Contribution to overall catch at Sunshine Coast netted beaches was highest in spring. An obvious seasonal cycle was observed at Gold Coast beaches, with an elevated catch of hammerheads from October to January. However, hammerheads comprised a large part of the elasmobranch catch throughout the year at Gold Coast beaches, comprising 12–40% of the species assemblage for each month.
Australian cownose rays were very rarely caught in nets on the Sunshine Coast (7 rays in 14 years), whereas at Gold Coast beaches, a seasonal pattern of abundance was observed, with higher catches from October to January. A defined seasonal pattern was not apparent for eagle rays at Sunshine Coast or Gold Coast beaches, although catches in October, January and March comprised 37% of the total catch of eagle rays from all netted beaches. Catches of bull sharks at Sunshine Coast and Gold Coast beaches remained fairly constant in all months, apart from August when only two sharks were caught from all netted beaches.
Elasmobranchs characterising and distinguishing between the fauna at Gold Coast and Sunshine Coast beaches
SIMPER revealed that Gold Coast beaches were characterised by hammerheads, spinner sharks, Australian cownose rays and eagle rays, which accounted for 1.86 (47%), 1.03 (26%), 0.61 (16%) and 0.16 (4%), respectively, of the average within-group similarity of 3.95. Sunshine Coast beaches were characterised by spinner sharks, hammerheads and bull sharks which accounted for 2.54 (62%), 1.07 (26%) and 0.13 (3%), respectively, of the average within-group similarity of 4.12. Spinner sharks, hammerheads, Australian cownose rays, eagle rays and bull sharks distinguished between Gold Coast and Sunshine Coast beaches, accounting for 25.22 (26%), 22.47 (23%), 8.10 (8%), 7.45 (8%) and 6.13 (6%), respectively, of the average dissimilarity of 96.60.
Elasmobranchs characterising and distinguishing between the fauna in seasons
SIMPER demonstrated that the catch in summer was characterised by spinner sharks, hammerheads and cownose rays, which accounted for 5.11 (63%), 2.17 (27%) and 0.29 (4%), respectively, of the average within-group similarity of 8.16. Catch in winter was characterised by hammerheads, bull sharks, spinner sharks and eagle rays, which comprised 0.94 (56%), 0.24 (15%), 0.18 (10%) and 0.16 (10%), respectively, of the within-group similarity of 1.69. Spinner sharks were the main elasmobranch group responsible for the dissimilarity between spring and summer (26%), summer and autumn (32%), autumn and winter (23%) and summer and winter (30%). Dissimilarity between spring and autumn (25%) and spring and winter (31%) was driven mainly by hammerheads.
Discussion
The present study identified spatial and seasonal partitioning among larger sharks in the coastal subtropical waters of south-eastern Queensland, Australia. The catch of sharks and rays from 19 netted beaches significantly differed among locations and seasons. These differences in the elasmobranch assemblage were largely driven by variations in the relative abundance of spinner sharks and hammerheads. There is a lack of information on partitioning among larger sharks and the present study demonstrated that in subtropical coastal waters, partitioning among larger sharks can occur over relatively fine spatial scales. Although the capture of 35 species of elasmobranchs highlights the high elasmobranch diversity, the overall catch in the shark nets was dominated by spinner sharks and hammerheads, which comprised 25% and 23%, respectively, of the total number of elasmobranchs caught.
Species composition in comparison to other subtropical and tropical waters
Spinner sharks are abundant in tropical and warm-temperate Atlantic and Indo-West Pacific waters (Compagno 1984). Previous analysis of spinner shark-catch data from the QSCP has revealed that catch rates off Gold Coast and Sunshine Coast beaches are over five times higher than those at the more northern areas of Mackay and Cairns where bather-protection nets are also used (Sumpton et al. 2010). The abundance of spinner sharks in the current study was also higher than that in KZN, South Africa, where spinner sharks comprised 10% of the total shark catch in the protective gill-nets from 1978 to 1997 (Allen and Cliff 2000) and also appears to be higher than that in central NSW where bather-protection nets are used (Dennis Reid, former New South Wales Department of Primary Industries, pers. comm.).
The abundance of hammerhead sharks in the present study concurs with that in other studies conducted in inshore waters by using large mesh-size gill-nets (Reid and Krogh 1992; Krogh 1994; Cliff 1995; de Bruyn et al. 2005). In KZN, South Africa, the ratio of scalloped hammerheads to great hammerheads was 12.8 : 1 between 1978 and 1993 (Cliff 1995; de Bruyn et al. 2005). In the current study, hammerheads were not always identified to the species level; however, of the 474 hammerheads identified to the species level (73% of all hammerheads caught), the ratio of scalloped hammerheads to great hammerheads was 4.2 : 1. Although this result is best viewed as preliminary, it suggests that scalloped hammerheads are the most abundant species of hammerhead in the coastal waters of south-eastern Queensland.
Although the present study has provided valuable insight into the abundance of pelagic sharks, the fact that nets were not bottom set suggests that the results are not representative for sharks and rays that spend large periods of time close to the benthic substrate. Furthermore, the catch composition from drumlines in the same study region is different, with tiger sharks dominating the catch and low numbers of spinner sharks and hammerheads caught on baited hooks (Sumpton et al. 2011). The difference between gill-net and drumline catch is most likely to be due to gear selectivity. Whereas catch in gill-nets is a function of the mesh size, the size of the animal and its rate of movement, drumlines catch only animals that prey on the bait type used.
Spatial partitioning
Differences in the elasmobranch assemblage were identified among netted beaches, spanning a distance of ∼220 km. Spinner sharks and hammerheads formed a key part of the catch at most locations; however, their abundance did vary among beaches, particularly between the beaches on the Sunshine Coast and those on the Gold Coast. Spinner sharks characterised the catch at the Sunshine Coast beaches, whereas hammerheads dominated the catch at the Gold Coast beaches. These results suggest that although the distribution of these sharks overlaps in the coastal waters of south-eastern Queensland, relatively small-scale spatial variations exist in their abundance. These variations are primarily responsible for discriminating between the elasmobranch fauna at Sunshine Coast beaches and those at Gold Coast beaches.
Although Australian cownose rays were the third-most abundant elasmobranch caught overall, they were rarely encountered in Sunshine Coast nets. These rays predominantly feed on molluscs and other shellfish (Last and Stevens 1994) and are fairly abundant in shallow sandy and seagrass regions in Moreton Bay, a large subtropical embayment that separates the Gold Coast from the Sunshine Coast (Johnson 1999; Schluessel 2008). The fact they were rarely caught in nets on the Sunshine Coast is somewhat surprising and future research is needed to elucidate patterns of habitat use in Australian cownose rays.
Generally, netted beaches in close proximity tended to have a similar catch composition, although the abundance of sharks caught at beaches in close proximity were sometimes very different, particularly for Sunshine Coast netted beaches. Although the nets at Noosa Beach and Noosa Headland caught similar species of elasmobranchs, the latter net caught over three times more sharks and rays. Surprisingly, only 21 elasmobranchs were caught in the net at Mooloolaba throughout the 14-year time period. Human disturbance at this beach was higher than in all other beaches, with a large number of recreational boat and jet-ski users passing close to the net. Krogh (1994) also observed lower catch rates of sharks in more urbanised beaches in the NSW Shark Meshing program. Higher catches were encountered at less urbanised beaches, which tended to be at either end of the netted regions (Krogh 1994). Although these observations do not imply a cause–effect relationship between human disturbance and catch rate, future behavioural studies could evaluate the effect of boat traffic on the distribution of larger sharks and their prey source, the results of which would be particularly relevant to ongoing bather-protection programs.
The selection of habitat by sharks in coastal waters is believed to be influenced by environmental characteristics, resource abundance and presence or absence of other competing species (Knip et al. 2010). Environmental characteristics that have been associated with the distribution of coastal elasmobranchs include tide, water temperature, salinity, dissolved oxygen, substratum type and depth (Speed et al. 2010). In the current study, differences in substratum type were recorded between the netted beaches on the Gold Coast and those on the Sunshine Coast. All nets at the Gold beaches were set over sandy substrate, whereas all but one of the nets at the Sunshine Coast beaches were set over coffee rock and sand or over rock (boulders).
In the absence of correlative data on habitat type, location and catch composition, we are unable to assess the extent to which differences in habitat type were responsible for spatial differences in the species assemblage between the netted beaches on the Sunshine Coast and those on the Gold Coast. Further acoustic-monitoring studies on spinner sharks and scalloped hammerheads would be particularly useful and would allow movement patterns to be compared with known biotic and abiotic variables. Measuring environmental characteristics in situ during each checking of the QSCP nets would allow some of these variables to be correlated against the catch composition.
There are interesting parallels between the present study and studies on the NSW Shark Meshing Program that also reported differential resource use in the catch of large sharks (Krogh 1994). In the NSW study, hammerheads had significantly higher catch rates on long open beaches and significantly higher catches of whaler sharks (Carcharhinus spp.), white sharks (Carcharodon carcharias) and tiger sharks were observed when deeper water was closer to the beach (Krogh 1994). On the basis of the analysis of QSCP from the Townsville region, Simpfendorfer (1992) also suggested that higher catches of tiger sharks are observed when channels of deeper water are adjacent to netted beaches.
Depth immediately adjacent to the nets is unlikely to explain the spatial partitioning in the current study, given that all nets were set in water between 6 and 12 m; however, the fact that the continental shelf off Sunshine Coast beaches extends further seaward, whereas the shelf of Gold Coast beaches is narrower, resulting in deeper water closer to the shore, may have also been partly responsible for some of the dissimilarity among the beaches, and in particular between the Gold Coast and Sunshine Coast beaches. Proximity to headlands could also be a factor responsible for the spatial disparity in species composition observed in the study, because the majority of netted beaches on the Sunshine Coast were close to headlands, whereas the netted beaches on the Gold coast tended to be more open and further from headlands.
Seasonal partitioning
The abundance of large sharks in shallow coastal waters (<12 m) in south-eastern Queensland was low during the winter and high during the summer. Although spinner sharks and hammerheads, the two most abundant sharks, both displayed elevated catches during the warmer months, spinner sharks were comparably less abundant during the winter.
Although seasonal patterns in the abundance of smaller sharks have been fairly well studied in Australian waters (Simpfendorfer and Milward 1993; Blaber et al. 1995; White and Potter 2004), less is known about the seasonal distribution of larger Carcharhiniformes. The reduced catch of sharks in subtropical waters in the cooler months is often linked to a seasonal migration to warmer coastal waters (Castro 1993; Last and Stevens 1994); however, Sumpton et al. (2010) identified low catches of spinner sharks during the winter from all of Queensland’s QSCP netted beaches, which extend ∼1000 km north from the current study region. This suggests that spinner sharks may migrate to deeper, offshore waters during winter (Sumpton et al. 2010).
On the basis of the average size of spinner sharks in the present study (198 cm TL) and the fact that both sexes of spinner sharks are assumed to mature at between 190 and 200 cm TL in Australian waters (Last and Stevens 1994), a large component of the catch in the present study would have been mature. The abundance of larger sharks in coastal waters during the summer has been associated with the nursery role that inshore waters provide for neonate and juvenile Carcharhiniformes (Springer 1967; Heupel et al. 2007). Neonate scalloped hammerheads, neonate blacktips (C. limbatus/tilstoni), neonate dusky sharks (C. obscurus), neonate nervous sharks (C. cautus) and neonate pigeye sharks (C. amboinensis) have been observed in shallow regions of Moreton Bay (<3-m depth at low tide) (Taylor 2008) and neonate spinner sharks have been reported from Hervey Bay in spring and summer, ∼250 km north of Moreton Bay (Adrian Gutteridge, University of Queensland, pers. obs.). Sumpton et al. (2010) observed near-term embryos from spinner sharks caught in Queensland waters from January to March. In parallel, these results suggest that the abundance of large female spinner sharks during the warmer months may be related to the nursery role that inshore waters provide for young spinner sharks in south-eastern Queensland.
Future stock assessments and risk assessments of sharks and rays in Queensland could benefit from the time series of standardised catch data reported in the present study and should consider the seasonal and spatial heterogeneity in the species composition of larger sharks between Sunshine Coast and Gold Coast coastal waters. The present study has demonstrated that even larger sharks that are known to migrate long distances vary in their spatial and seasonal patterns of abundance when they occupy shallow coastal waters. Although advances in passive-monitoring technology and satellite tags have increased our understanding of the ecology and behaviour of larger sharks, there is still a need for information on population-level differences in abundance and distribution. Bather-protection programs can provide this information and while they continue to exist, their catch data should be further analysed to improve our understanding of partitioning among larger sharks.
Acknowledgements
We acknowledge the contributions made by the various QSCP contractors since the program began in 1962. We are particularly grateful to Craig Newton (Gold Coast), Noel Walker (Sunshine Coast, 1996–2007) and Paddy Dimond (Sunshine Coast, 2007 to present) who operated the QSCP fishing equipment during the study period. The guidance of Baden Lane who was the manager of the QSCP for much of the period of investigation is appreciated. Bob Clarke, Ian Halliday and James Webley provided statistical and analytical advice. We thank Katherine Zahmel for assistance with GIS. Comments by Simon Pierce also greatly improved the manuscript. The map in Fig. 1 incorporates data that are copyright Commonwealth of Australia 2010 and have been used with the permission of the Commonwealth. The map also incorporates data that are copyright Pitney Bowes Mapinfo. Finally, we are very grateful to the reviewers and editors for their insightful comments.
References
Allen, B. R., and Cliff, G. (2000). Sharks caught in the protective gill nets off KwaZulu-Natal, South Africa. 9. The spinner shark Carcharhinus brevipinna (Muller and Henle). South African Journal of Marine Science 22, 199–215.| Sharks caught in the protective gill nets off KwaZulu-Natal, South Africa. 9. The spinner shark Carcharhinus brevipinna (Muller and Henle).Crossref | GoogleScholarGoogle Scholar |
Blaber, S. J. M., Brewer, D. T., and Salini, J. P. (1995). Fish communities and the nursery role of the shallow inshore waters of a tropical bay in the Gulf of Carpentaria, Australia. Estuarine, Coastal and Shelf Science 40, 177–193.
| Fish communities and the nursery role of the shallow inshore waters of a tropical bay in the Gulf of Carpentaria, Australia.Crossref | GoogleScholarGoogle Scholar |
Castro, J. I. (1993). The shark nursery of Bulls Bay, South Carolina, with a review of the shark nurseries of the southeastern coast of the United States. Environmental Biology of Fishes 38, 37–48.
| The shark nursery of Bulls Bay, South Carolina, with a review of the shark nurseries of the southeastern coast of the United States.Crossref | GoogleScholarGoogle Scholar |
Clarke, K. R., and Gorley, R. (2006). ‘PRIMER v6: User Manual/Tutorial.’ (PRIMER-E: Plymouth, UK.)
Clarke, K. R., and Warwick, R. M. (2001). ‘Change in Marine Communities: an Approach to Statistical Analysis and Interpretation.’ 2nd edn. (PRIMER-E: Plymouth, UK.)
Cliff, G. (1995). Sharks caught in the protective gill nets off Kwazulu-Natal, South-Africa. 8. The great hammerhead shark Sphyrna mokarran (Ruppell). South African Journal of Marine Science 15, 105–114.
| Sharks caught in the protective gill nets off Kwazulu-Natal, South-Africa. 8. The great hammerhead shark Sphyrna mokarran (Ruppell).Crossref | GoogleScholarGoogle Scholar |
Cliff, G., and Dudley, S. F. J. (1991). Sharks caught in the protective gill nets off Natal, South-Africa. 5. The java shark Carcharhinus amboinensis (Muller and Henle). South African Journal of Marine Science 11, 443–453.
| Sharks caught in the protective gill nets off Natal, South-Africa. 5. The java shark Carcharhinus amboinensis (Muller and Henle).Crossref | GoogleScholarGoogle Scholar |
Compagno, L. J. V. (1984). Sharks of the world. An annotated and illustrated catalogue of shark species to date. Part 2 (Carcharhiniformes). FAO Fisheries Synopsis No 125, 251–655.
de Bruyn, P., Dudley, S. F. J., Cliff, G., and Smale, M. J. (2005). Sharks caught in the protective gill nets off KwaZulu-Natal, South Africa. 11. The scalloped hammerhead shark Sphyrna lewini (Griffith and Smith). South African Journal of Marine Science 27, 517–528.
DeAngelis, B. M., McCandless, C. T., Kohler, N. E., Recksiek, C. W., and Skomal, G. B. (2008). First characterization of shark nursery habitat in the United States Virgin Islands: evidence of habitat partitioning by two shark species. Marine Ecology Progress Series 358, 257–271.
| First characterization of shark nursery habitat in the United States Virgin Islands: evidence of habitat partitioning by two shark species.Crossref | GoogleScholarGoogle Scholar |
Dudley, S. F. J. (1997). A comparison of the shark control programs of New South Wales and Queensland (Australia) and KwaZulu-Natal (South Africa). Ocean and Coastal Management 34, 1–27.
| A comparison of the shark control programs of New South Wales and Queensland (Australia) and KwaZulu-Natal (South Africa).Crossref | GoogleScholarGoogle Scholar |
Dudley, S. F. J., and Simpfendorfer, S. A. (2006). Population status of 14 shark species caught in the protective gillnets off KwaZulu-Natal beaches, South Africa 1978–2003. Australian Journal of Marine and Freshwater Research 57, 225–240.
| Population status of 14 shark species caught in the protective gillnets off KwaZulu-Natal beaches, South Africa 1978–2003.Crossref | GoogleScholarGoogle Scholar |
Heithaus, M. R., Frid, A., Vaudo, J. J., Worm, B., and Wirsing, A. J. (2010). Unraveling the ecological importance of elasmobranchs. In ‘Sharks and their Relatives II: Biodiversity, Adaptive Physiology, and Conservation’. (Eds J. C. Carrier, J. A. Musick and M. R. Heithaus.) pp. 611–639. (CRC Press: Boca Raton, FL.)
Heupel, M. R., Semmens, J. M., and Hobday, A. J. (2006). Automated acoustic tracking of aquatic animals: scales, design and deployment of listening station arrays. Australian Journal of Marine and Freshwater Research 57, 1–13.
| Automated acoustic tracking of aquatic animals: scales, design and deployment of listening station arrays.Crossref | GoogleScholarGoogle Scholar |
Heupel, M. R., Carlson, J. K., and Simpfendorfer, C. A. (2007). Shark nursery areas: concepts, definition, characterization and assumptions. Marine Ecology Progress Series 337, 287–297.
| Shark nursery areas: concepts, definition, characterization and assumptions.Crossref | GoogleScholarGoogle Scholar |
Johnson, J. W. (1999). Annotated checklist of the fishes of Moreton Bay, Queensland, Australia. Memoirs of the Queensland Museum 43, 709–762.
Kinney, M. J., and Simpfendorfer, C. A. (2009). Reassessing the value of nursery areas to shark conservation and management. Conservation Letters 2, 53–60.
| Reassessing the value of nursery areas to shark conservation and management.Crossref | GoogleScholarGoogle Scholar |
Knip, D. M., Heupel, M. R., and Simpfendorfer, C. A. (2010). Sharks in nearshore environments: models, importance and consequences. Marine Ecology Progress Series 402, 1–11.
| Sharks in nearshore environments: models, importance and consequences.Crossref | GoogleScholarGoogle Scholar |
Krogh, M. (1994). Spatial, seasonal and biological analysis of sharks caught in the New South Wales Protective Beach Meshing Program. Australian Journal of Marine and Freshwater Research 45, 1087–1106.
| Spatial, seasonal and biological analysis of sharks caught in the New South Wales Protective Beach Meshing Program.Crossref | GoogleScholarGoogle Scholar |
Kyne, P. M., Johnson, J. W., Courtney, A. J., and Bennett, M. B. (2005). New biogeographical information on Queensland chondrichthyans. Memoirs of the Queensland Museum 50, 321–327.
Last, P. R., and Stevens, J. D. (1994). ‘Sharks and Rays of Australia.’ (CSIRO Publishing: Melbourne.)
Papastamatiou, Y. P., Wetherbee, B. M., Lowe, C. G., and Crow, G. L. (2006). Distribution and diet of four species of carcharhinid shark in the Hawaiian Islands: evidence for resource partitioning and competitive exclusion. Marine Ecology Progress Series 320, 239–251.
| Distribution and diet of four species of carcharhinid shark in the Hawaiian Islands: evidence for resource partitioning and competitive exclusion.Crossref | GoogleScholarGoogle Scholar |
Pikitch, E. K., Chapman, D. D., Babcock, E. A., and Shivji, M. S. (2005). Habitat use and demographic population structure of elasmobranchs at a Caribbean atoll (Glover’s Reef, Belize). Marine Ecology Progress Series 302, 187–197.
| Habitat use and demographic population structure of elasmobranchs at a Caribbean atoll (Glover’s Reef, Belize).Crossref | GoogleScholarGoogle Scholar |
Pratt, H. L., and Carrier, J. C. (2001). A review of elasmobranch reproductive behavior with a case study on the nurse shark, Ginglymostoma cirratum. Environmental Biology of Fishes 60, 157–188.
| A review of elasmobranch reproductive behavior with a case study on the nurse shark, Ginglymostoma cirratum.Crossref | GoogleScholarGoogle Scholar |
Reid, D. D., and Krogh, M. (1992). Assessment of catches from protective shark meshing of New South Wales beaches between 1950 and 1990. Australian Journal of Marine and Freshwater Research 43, 283–296.
| Assessment of catches from protective shark meshing of New South Wales beaches between 1950 and 1990.Crossref | GoogleScholarGoogle Scholar |
Schluessel, V. (2008). Life history, population genetics and sensory biology of the white spotted eagle ray Aetobatus narinari (Euphrasen, 1790) with emphasis on the relative importance of olfaction. PhD Thesis, University of Queensland, Brisbane.
Simpfendorfer, C. A. (1992). Biology of tiger sharks (Galeocerdo cuvier) caught by the Queensland Shark Control Program off Townsville, Australia. Australian Journal of Marine and Freshwater Research 43, 33–43.
| Biology of tiger sharks (Galeocerdo cuvier) caught by the Queensland Shark Control Program off Townsville, Australia.Crossref | GoogleScholarGoogle Scholar |
Simpfendorfer, C. A., and Heupel, M. R. (2004). Assessing habitat use and movement. In ‘Biology of Sharks and their Relatives’. (Eds J. C. Carrier, J. A. Musick and M. R. Heithaus.) pp. 553–572. (CRC Press: Boca Raton, FL.)
Simpfendorfer, C. A., and Milward, N. E. (1993). Utilisation of a tropical bay as a nursery area by sharks of the families Carcharhinidae and Sphyrnidae. Environmental Biology of Fishes 37, 337–345.
| Utilisation of a tropical bay as a nursery area by sharks of the families Carcharhinidae and Sphyrnidae.Crossref | GoogleScholarGoogle Scholar |
Speed, C. W., Field, I. C., Meekan, M. G., and Bradshaw, C. J. A. (2010). Complexities of coastal shark movements and their implications for management. Marine Ecology Progress Series 408, 275–293.
| Complexities of coastal shark movements and their implications for management.Crossref | GoogleScholarGoogle Scholar |
Springer, S. (1967). Social organisation of shark populations. In ‘Sharks, Skates and Rays’. (Eds P. W. Gilbert, R. F. Mathewson and D. P. Rall.) pp. 149–174. (John Hopkins Press: Baltimore, MD.)
Sumpton, W., Lane, B., and Ham, T. (2010). Characteristics of the biology and distribution of the spinner shark (Carcharhinus brevipinna) in Queensland, Australia based on data collected from the Shark Control Program. Asian Fisheries Science 23, 340–354.
Sumpton, W., Taylor, S., Gribble, N., McPherson, G., and Ham, T. (2011). Gear selectivity of large-mesh nets and drumlines used to catch sharks in the Queensland Shark Control Program. African Journal of Marine Science 33, 37–43.
Taylor, S. M. (2008). Population structure and resource partitioning among Carcharhiniform sharks in Moreton Bay, southeast Queensland, Australia. PhD Thesis, University of Queensland, Brisbane.
Walker, T. I. (1998). Can shark resources be harvested sustainably? A question revisited with a review of shark fisheries. Australian Journal of Marine and Freshwater Research 49, 553–572.
| Can shark resources be harvested sustainably? A question revisited with a review of shark fisheries.Crossref | GoogleScholarGoogle Scholar |
Warwick, R. M., Clarke, K. R., and Gee, J. M. (1990). The effect of disturbance by soldier crabs, Mictyris platycheles H. Milne Edwards on meiobenthic community structure. Journal of Experimental Marine Biology and Ecology 135, 19–33.
| The effect of disturbance by soldier crabs, Mictyris platycheles H. Milne Edwards on meiobenthic community structure.Crossref | GoogleScholarGoogle Scholar |
White, W. T., and Potter, I. C. (2004). Habitat partitioning among four elasmobranch species in nearshore, shallow waters of a subtropical embayment in Western Australia. Marine Biology 145, 1023–1032.
| Habitat partitioning among four elasmobranch species in nearshore, shallow waters of a subtropical embayment in Western Australia.Crossref | GoogleScholarGoogle Scholar |