

Soft bodies make estimation hard: correlations among body dimensions and weights of multiple species of sea cucumbers
James Prescott A , Shijie Zhou B D and Andhika P. Prasetyo CA Australian Fisheries Management Authority, PO Box 131, Darwin NT 0801, Australia.
B CSIRO Oceans and Atmosphere Flagship, GPO Box 2583, Brisbane, Qld 4001, Australia.
C Research Center for Fisheries Management and Conservation, Ministry for Marine Affairs and Fisheries, Jl. Pasir Putih II, Ancol Timur 14430, Jakarta, Indonesia.
D Corresponding author. Email: shijie.zhou@csiro.au
Marine and Freshwater Research 66(10) 857-865 https://doi.org/10.1071/MF14146
Submitted: 6 June 2014 Accepted: 19 November 2014 Published: 19 March 2015
Journal Compilation © CSIRO Publishing 2015 Open Access CC BY-NC-ND
Abstract
Tropical sea cucumbers are commonly exploited by small-scale, poorly managed fisheries. A fundamental problem in managing sea cucumber fisheries is the lack of basic knowledge of important life history characteristics for most species. As a result of plastic body dimensions, biological research on this group of animals becomes exceptionally challenging. To improve our understanding of essential biological parameters, we conducted a study to investigate correlations among various body measurements. We analysed a total of 18 sea cucumber species and more than 6600 individuals collected at Scott Reef in the Timor Sea, north-west Australia. We used hierarchical Bayesian errors-in-variables models to specifically take into account measurement errors that are obviously unavoidable. The measures included three types of weights (wet weight, gutted weight and dry weight) and two body dimensions (length and width). The modelling reveals that using both body length and width as independent variables, wet weight increases approximately linearly with body length, but is a power function (~1.6) of body width, although variability exists among species. Dry weight tends to increase more slowly with body length, but has a similar power function of body width. Linear relationships are established between the three types of weights. On average, ~11% of a live specimen and ~16% of a gutted specimen is processed to the commercially traded dry body wall. Our results can be applied to sea cucumbers in other areas and can be useful for data standardisation and size-based fisheries management.
Additional keywords: beche-de-mer, Indonesia, small-scale fisheries, trepang.
Introduction
Commercially valuable sea cucumbers are distributed over much of the world’s shallow coastal seas and they are often intensively exploited. In many developing countries, where livelihood alternatives are limited and governance structures are often less well developed, many sea cucumber stocks have been depleted and are in need of better management (Purcell et al. 2013). However, most sea cucumber fisheries are lacking assessments which are typically part of the management process and dialogue that have helped produce positive outcomes in other fisheries where success has been achieved. Assessments have been constrained and management compromised by the absence of important biological information (Conand 1990). Assessments are generally made using fishery independent methods that are independent of the fishers who then have no ownership of the process or outcomes.
There are several reasons for the limited progress in applied fisheries research of sea cucumbers. These animals have plastic size, shape and weight which makes their measurement very difficult (Perry et al. 1999; Laboy-nieves and Conde 2006; Yamana and Hamano 2006). They also have highly variable water content which affects their size, shape and weight (Hannah et al. 2012). Tropical fisheries are characteristically composed of multiple species which compounds the difficulty of the research task. In addition, in many areas stocks in these small scale fisheries are so depleted that collecting enough data to even derive basic morphometric relationships for each species is an imposing challenge. Given these difficulties, biological measurements have been identified as one of the critical gaps that undermine the expected success of management strategies (Purcell et al. 2013). For example, biomass estimates are often an important variable for fisheries management and visual transect surveys of one form or another are the most commonly recommended method for quantifying sea cucumber stocks (e.g. Purcell and Cheng 2010). These surveys provide information on the apparent density and abundance of the observed species. If mean weight of the relevant species is known their apparent biomass can be estimated. Yet, sea cucumbers encountered in situations where weight can be readily measured (e.g. fishing boats and markets) are commonly found in different forms (e.g. live, gutted or dried), and the relationships between alternative measurements and between weight and body dimensions have not been studied for many species.
On reefs off north-western Australia sea cucumbers, locally called trepang (English) or teripang (Indonesian) have been harvested by Indonesian fishers to varying degrees for several centuries (Fox and Sen 2002). Over this long history of exploitation the fishery has undergone many changes including the species that are harvested, with many commercially less attractive species ‘filling the gap’ left as higher value species became more scarce, a common feature of many such fisheries (Toral-Granda et al. 2008; Branch et al. 2013). From a research and management perspective this has introduced a sizeable number of little studied species to the fishery as past research focussed on a smaller number of high value species.
During our first field studies at Scott Reef in 2008, observations of catch onboard fishing vessels and from fishing activities on the reef top indicated that fishers harvested not just a wide range of species, but also all sizes of sea cucumbers that they detected. Consequently the catch consisted of all age groups present on the reef flats, including what appeared to be the young-of-the-year for several species. As growth rates are one of the most important life history parameters in most fisheries’ assessments, using ‘size’ data that could be collected in the fishery to estimate growth was a highly attractive proposition. In this fishery, where it has been possible to estimate fishing mortality rates for several of the important species (Prescott et al. 2013), the asymptotic size and growth parameter have a potential to be used as a proxy for natural mortality (M) and management reference points, such as FMSY (Quinn and Deriso 1999; Zhou et al. 2012).
In this paper our main objective is to investigate the relationships between several metrics of size of live sea cucumbers and their corresponding wet, gutted and dried weights (the latter is the metric with commercial importance as it is typically the marketed form of sea cucumbers). This study was the first step towards estimating the growth of the harvested species at Scott Reef. The results could be also useful for fisheries management more broadly, such as to monitor size limits that could be set to protect juveniles and recently matured adults to allow individuals one or more seasons to spawn before they can be fished (Purcell et al. 2009b). Further, the conversions from wet weight to dry weight examined in this paper are needed to standardise catch in either form (Purcell et al. 2009a). This represents a substantial improvement as it allows data to be collected and compared from multiple sources regardless of the form the sea cucumbers are in when they are encountered. These relationships are reported on for the first time for several widespread Indo-Pacific species.
Materials and methods
Study area and sample procedure
Scott Reef comprises north and south reefs that sit atop the Scott Plateau that rises from the continental slope in water depths of ~800 m (Collins 2011). The reefs are ~270 km off the coast of Australia in the Timor Sea.
Scott Reef is fished by Indonesian fishermen under a 40-year-old Memorandum of Understanding between Australia and Indonesia. Numbers of vessels visiting the reefs is variable but has ranged from 11 to more than 50 at one time during recent years of field work and catches of sea cucumbers are estimated to have ranged from ~100 to 300 tonnes liveweight (J. Prescott, unpubl. data). Most fishing occurs at low tide when the extensive reef flats are exposed so spring tide periods are generally the periods of highest activity. Fishing is conducted during both day and night but many of the species that currently make of the majority of the catch and are important in our study are predominantly harvested at night when they are more readily found.
Scientists from Australia and Indonesia have been working together on research and management of the traditional Indonesian sea cucumber fishery at Scott Reef for 5 years and the observations and measurements used in this paper were made in 2009, 2011 and 2012. During this period more than 6600 individual sea cucumbers, belonging to 17 species and one putative hybrid and were measured for their body size (length and width) and weight. We recorded three types of weights: wet weight (with gut-in), gutted weight (wet weight with gut-out) and dry weight (always gut-out). The dry weight measurement was as processed by the fishers, as opposed to a stable dry weight obtained in a laboratory. All sampling at Scott Reef was undertaken during the months of August and September, the Austral cool months at this latitude. During this period no developing gonads were observed among any of the species, although Holothuria whitmaei (formerly H. nobilis) is known to spawn during the cool months in New Caledonia (Conand 1990). A few other species in our sample, or near their near relatives, are known to spawn during the warmer months where this has been studied (Conand 1990).
Sea cucumbers were measured and weighed on the traditional fishing vessels by prior arrangement with the crews who were asked to retain their catches in their natural state until the survey team could carry out the sampling. This meant that most sea cucumbers were unavoidably harvested several hours before they were measured and weighed, during which time they were kept in various containers available to crews. All sea cucumbers in each catch were measured unless this was prevented by the particular circumstances, for example Stichopus chloronotus was not usually held for sampling because the body wall would break down if they were not processed soon after landing.
Each sea cucumber was identified to species (or hybrid) and measured in two dimensions (length and width) using fish measuring board with back-board perpendicular to the longitudinal measuring surface. Width at the widest point of the animal was measured quickly after its length while the animal was held against the backboard in order to avoid changes in width relative to length. Then the sea cucumber was weighed using a Salter spring balance of an appropriate scale for the weight of the sea cucumber such that the scale units were approximately a constant proportion of the animal’s weight, i.e. measurement precision was proportional to weight. Based on their appearance and feel of ‘fullness’, or obvious processing status samplers classified each sea cucumber as not eviscerated (gut-in), auto eviscerated (gut-out) or gutted. Gut (intestinal) fullness varied from animal to animal but there was no way to quickly quantify this. A subsample of sea cucumbers were individually marked with numbered plastic ties so they could be identified for re-weighing after gutting and subsequently after the drying process was completed to the satisfaction of the fishers. Some were reweighed at sea after drying, using a spring scale while others were reweighed in Indonesia at the point of landing using a HL-3000 LWP electronic balance with a precision of 1 g.
Clearly, for the reasons described above, field measurements of body dimensions and weight in sea cucumbers incur some measurement error. In classical statistical methods such as regression it is difficult to include errors in both dependent and independent variables. Another issue in this study was how to handle multiple species that belong to the same broader taxonomic group of animals and have similar life history traits. Classical methods typically assume that there is no link among species and analyse species by species separately. To take these two fundamental facts into consideration, we proposed to use a hierarchical Bayesian errors-in-variables approach (Richardson and Best 2003; Zhou et al. 2012).
Relationship between bodyweight and body dimension
We analysed the relationship between two types of bodyweight and two types of measurement of body dimension for each species: the dry weight and wet weight, and body length and width of living wet specimens. The general model is:
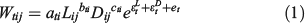
where Wtij is the weight of type t, species i and individual j, L is the observed (wet) body length and D is observed the (wet) body width. There are three parameters, a, b and c. In addition, there are three random errors: observation error in length ϵtL, observation error in width ϵtD and model process error et. These errors were respectively assumed to be normally distributed with a mean of 0 and standard deviation of σtL, σtD and σtW. This model specifically takes uncertainty in measuring the soft sea cucumbers into consideration. Model (1) indicates that the unobserved true variable z (i.e. length or width) is:

where yij is the observed body length or width, and ϵ is a normal random variable with mean 0 and standard error σϵ. This simple model assumes a multiplicative error structure because variance in these measurements often increases with the variables themselves. We also examined the relationship between bodyweight and one measurement of body dimension, either body length or width by excluding the other dimension in Model (1).
Instead of analysing each species independently, we assumed that underlying similarities exist in the relationship and the three types of errors across all species. The hierarchical Bayesian modelling allows sharing and borrowing information across species, hence increasing the accuracy of parameters, particularly for species that have fewer data or larger variance. This is achieved by drawing the parameters of the individual species from a common statistical distribution. One of the alternative hierarchical structures is:
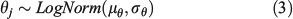
where θj is parameters aj, bj or cj, μθ and σθ are the mean and standard deviation of the priors. The two hyper-parameters were modelled as μθ ~Normal(0, 0.01) and σθ ~Inverse-gamma(0.01, 0.01). These specifications provide reasonably non-informative priors and hyper-priors. There were 17 sea cucumber species and one hybrid that had sufficient data for wet weight v. body dimension analysis, but only 13 species had enough data for dry weight v. body dimension analysis because of missing data and low sample sizes.
Correlations between three types of bodyweights
We examined three types of correlations: dry weight v. wet weight, dry weight v. gutted weight and gutted weight v. wet weight. The gutted weight refers to wet specimens without guts which occurred naturally if the animals had auto eviscerated or they had been gutted by the fishers. Similar hierarchical Bayesian measurement error models are used for analysing correlations between different types of bodyweight measurements. The general model is expressed as.
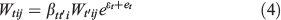
where Wtij is the weight of type t, species i, individual j as in model (1), β is the parameter to be estimated, t′ is the type of weight other than t, ϵ is the measurement error and e is the model (process) error. For each species, parameter βi is assumed as.

where μβ is the prior mean and σβ is the standard deviation. These two hyper-parameters have their own distributions. Here we assumed μβ ~ N(0, 10) and σβ ~ IG(0.01, 0.01).
The analyses were implemented in WinBUGS. Three Markov chain Monte Carlo (MCMC) chains were run for 60 000 iterations. The first 10 000 samples were excluded for parameter inference. We tested alternative model structure and priors which included normal and lognormal distributions, hierarchical and non-hierarchical structures, individual species and all species combined, additive and multiplicative errors, homogeneous and heterogeneous variance across species, etc. Some models converged quickly while in others convergence was more problematic. We used deviation information criterion (DIC) to compare the alternative models.
Results
Relationship between bodyweight and body dimension
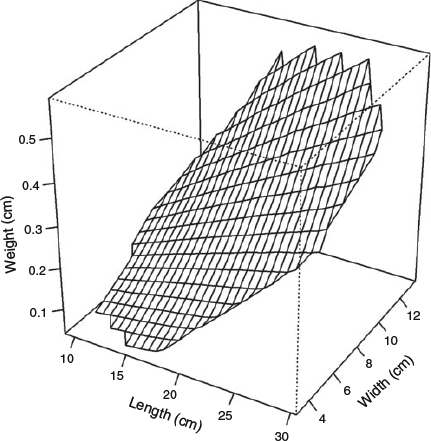
Sea cucumbers typically exhibit increasing bodyweight with both body length and width. The effect of length and width appears to be similar, i.e. both are important in accurately describing the weight of an individual (Fig. 1). This visual inspection was confirmed by modelling. Based on DIC, the best model was the one that included both body length and width as predictors, used species specific and hierarchically structured parameters a, b and c, and assumed multiplicative and homogeneous variances in measurement error and process error. However, a model that combined all species together and assumed a common set of parameters a, b and c was difficult to run and converge. Hence, we only report the results of hierarchical models that did not combine all species into one single group.
|
For the wet weight, the median parameter b ranged from 0.83 to 1.17 and parameter c ranged from 1.37 to 1.82 for the 18 ‘species’ (Fig. 2, Table 1). The narrow 95% CI indicates a strong relationship between weight and body dimension. The estimated bs are close to 1, meaning one unit increase in length transfer to approximately one unit of increase in weight. However, the estimated cs are larger than one, resulting in a non-linear power increase in weight. The scale parameter a depends on the units used (i.e. grams or kilograms for weight, and centimetres or millimetres for dimension), but is fairly consistent among species. Correlation between b and c was moderately high, with an estimated correlation coefficient of –0.76, compared with –0.39 between a and b, and –0.16 between a and c. The measurement error, expressed as standard deviation, was σtL = 0.06 for length and σtD = 0.15 for width. As a comparison, the model process error was σtW = 0.08

|

|
When only one body dimension (either length or width) was used as a predictor, the model fits the data much more poorly. With only length or width, the model had a DIC 5998 or 5366 higher than that from the model including both length and width, indicating a significantly poorer fit. The estimated parameters a, b or c are more diverse across species than the model with both length and width (Table 2). Compared with σtW = 0.08 when the wet weight was a function of both body dimension (length and width), the model process error was 0.45 and 0.32 when only body length or body width was used as a predictor respectively.

|
For the dry weight, the model incorporating both length and width as predictors produced good predictions (Fig. 3). The three model parameters for dry weights were more consistent across species than the model for wet weights. The median parameter b ranges from 0.78 to 0.86 and parameter c ranges from 1.46 to 1.66 for the 13 species (Table 3). Correlation between b and c was similar to wet weight estimates, with an estimated correlation coefficient of –0.75. The standard deviation of measurement error was respectively σtL = 0.23 and σtD = 0.13 for the body length and width. The model process error was σtW = 0.19. These errors were larger than in the wet weight, as there were much smaller sample sizes for dry weight.

|

|
Similar to the wet weight, when only one body dimension (either length or width) is used as a predictor, the model fitted the data much poorly. With only length or width, the model had a DIC 128 or 165 higher than that from the model that included both length and width, again, indicating a significantly poorer model fit. The estimated parameters a, b or c were also more constant across species than in the case of wet weight (Table 4).

|
Correlations between dry weight, wet weight and gutted weight
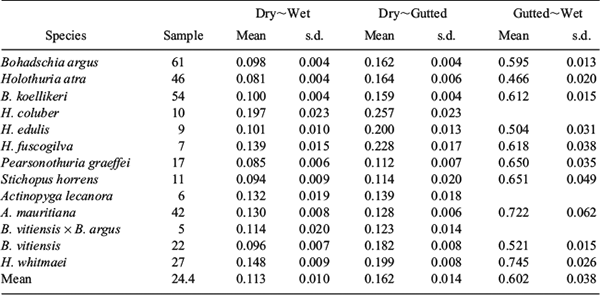
Clearly, there is a linear relationship between any two types of weight (Fig. 4). The models for correlation between different types of weight were very simple. Hence, they converged very quickly without any abnormal behaviour. Among the alternative models, the structure in the form of Model (4) has a lowest DIC (best model fitting) for all three types of correlations. This model is a species specific model with a common measurement error ϵ and a common process error e. If all species were combined together, the model with a single parameter β produced a higher DIC (poor fitting). For the correlation between dry weight and wet weight, the mean parameter βdry–wet,i ranges from 0.085 for Pearsonothuria graeffei to 0.197 for Holothuria coluber (Table 5). The overall mean for the 13 species has a μβ,dry–wet = 0.113 and σβ,dry–wet = 0.010. This means that on average ~89% of these species of live sea cucumber was water and gut. For the dry weight v. gutted weight, a μβ,dry–gutted = 0.162 and σβ,dry–gutted = 0.014; and for the gutted weight v. wet weight, a μβ,gutted–wet = 0.602 and σβ,gutted–wet = 0.038. Hence, water constituted ~84% of gutted specimens and the gut (the intestine and its contents, and respiratory tree) is ~40% of the total wet weight.

|
|
Discussion
This paper examined the relationship between sea cucumber bodyweight and body dimension, and relationship between different types of bodyweight. Weight was measured in three ways: wet weight (with gut in), gutted weight (wet) and dry weight (gut out); body dimension was measured by length and width. We applied hierarchical Bayesian measurement error models to deal with the challenging plasticity sea cucumbers exhibit in body dimensions and weight. The results show that when both body length and width were simultaneously used as predictors, wet weight typically increased with body length approximately linearly, but increased with body width in a power function of ~1.6 on average, although variability existed among species, reflecting the range of shapes they exhibit. Compared with wet weight, dry weight tends to increase more slowly with body length, but as a power function with body width. The relationships between the three types of weight are linear. On average, only ~11% of the weight of live sea cucumbers and ~16% of weight of gutted sea cucumber converted commercially valuable dried body wall weight in the species studied.
Measuring sea cucumber body dimensions and weight and determining their relationship is notoriously difficult (Perry et al. 1999; Laboy-nieves and Conde 2006). This research was important because it shed light on body dimension and weight relationships of several species that have not been reported on previously. By working with the fishermen large samples of the most common species were possible while even comparatively uncommon species in the catch were sampled well enough to estimate the various parameters in combination with the hierarchical Bayesian modelling approach.
Although sampling sea cucumbers in situ would have been preferable for biological studies, this was impractical at Scott Reef because of their low density and the need to carry equipment on several kilometre-long fishing excursions over the expansive low tide reef flat. Similarly, methods that might reduce measurement error including the use of anaesthesia or cold water shock to deal with the plastic dimensions (Guzman and Guevara 2002; Laboy-nieves and Conde 2006; Yamana and Hamano 2006) or weighing in water to overcome water content effects (Hannah et al. 2012) were impossible under the fishery conditions. Instead the catch was measured on the fishing vessels that were accessible to the survey team and this contributed to measurement error in this study. However, for the purpose of fisheries management, samples taken from a fishery, such as in our study, should reflect more accurately the various body dimension relationships that would be observed under common fishery conditions. Our estimated relationships between dry and wet weight can be used to make more accurate estimates of the liveweight harvests for a wider range of species that comprise the catch of many contemporary fisheries in the Indo Pacific region. Seeing as most exports of sea cucumber product tend to be centralised in most jurisdictions, sampling at this convenient point in the value chain can help to improve national liveweight harvest statistics.
Measurement errors, if not accounted for, will bias the parameter estimation (Su and Peterman 2012). In this paper, we used Bayesian hierarchical measurement error model technique to analyse correlations between bodyweight and body dimension and among different types of bodyweight for over a dozen species of sea cucumbers. The relationship between various body measurements is strong and unique to each species, though similarities exist across species. The improved results contrast with studies showing a weak relationship between bodyweight and length (Guzman and Guevara 2002). The key seems to be the simultaneous measurement of length and width to characterise weight, and the use of modelling techniques that adequately deal with inherent measurement errors.
Changes in weight during processing from wet weight to gutted weight and finally dry weight are not well investigated for many ‘low value’ species which now comprise the bulk of the catch in many fisheries. A previous study that included multiple species was carried out in Torres Strait, north-east Australia (Skewes et al. 2004). In the Torres Strait study seven, six and four species were examined for weight conversion ratios between live v. gutted, gutted v. dry and live v. dry respectively. They reported that the conversion ratio (i.e. parameter β in Model 4 this paper) between wet weight and dry weight varied from 0.048 to 0.126 with a mean of 0.085 for four species (i.e. between 4.8 and 12.6% and a mean of 8.5%), between gutted and dry weight from 0.094 to 0.247 (mean = 0.181 for 6 species) and between wet weight and gutted weight from 0.0480 to 0.633 (mean = 0.528 for 7 species). The results are approximately comparable, but their values for Dry v. Wet and Gutted v. Wet relationships tend to be smaller than our estimates. Purcell et al. (2009b) studied weight change during process stages for six sea cucumber species with 7–15 individuals each. Their results are also similar to ours. The reproductive condition of the animals, handling by the fishermen of the wet product and processing methods as well as the weighing process (at sea or under controlled conditions) are all likely to influence the results and therefore some of the differences that exist in the relationships between different studies should not be unexpected. Nevertheless, the large number of species and specimens examined in our study enrich existing knowledge from limited samples (Purcell et al. 2009a).
Sound fisheries management requires a range of information. Biological parameters, including growth, natural mortality, age or size structure, reproduction, abundance, etc., are typical input parameters for stock assessments. Our results suggest that the relationship between wet weight, dry weight and gutted weight are strong enough to be able to measure any of these forms and convert the weights to a single, consistent measurement. We provide these relationships for a large number of species that are not well studied, but are extensively exploited in waters to the north of Australia. Given the quantification of these relationships, the opportunity now exists to access and translate sea cucumber harvest data for a wide range of exploited species from an equally wide range of access points; fishing boats, processing areas and from the final product at first point of sale and further along the value chain regional markets. The results in this paper will become fundamental information for quantitative stock analysis where biological reference points and associated fisheries measures can be estimated.
Acknowledgements
This research was collaboration between Australia and Indonesia involving the Australian Fisheries Management Authority and the Indonesian Ministry of Marine Affairs and Fisheries. Critical support for the research was provided by Australian Border Protection Command who made available their vessels ACV Ashmore Guardian, ACV Hervey Bay and ACV Corio Bay and their respective crews. Sampling was undertaken at Scott Reef on more than 30 traditional fishing vessels whose crews were universally helpful, including in Indonesia where some crews also assisted with weighing the dried trepang. Captain Arfaksad Halundaka and his crew deserve special mention for their exceptional assistance. James Riwu, Dian Octaviani, Anthony Panggabean, Muhamad Taufik, Andrew Brown, Niki Alber, Sam Hyson and Miranda Carver carried out much of the sampling at Scott Reef and in Indonesia. We thank Marcus Finn and Timothy Skewes for their constructive comments on the earlier versions of the paper.
References
Branch, T. A., Lobo, A. S., and Purcell, S. W. (2013). Opportunistic exploitation: an overlooked pathway to extinction. Trends in Ecology & Evolution 28, 409–413.| Opportunistic exploitation: an overlooked pathway to extinction.Crossref | GoogleScholarGoogle Scholar |
Collins, L. B. (2011). Geological setting, marine geomorphology, sediments and oceanic shoals growth history of the Kimberley region. Journal of the Royal Society of Western Australia 94, 89–105.
Conand, C. (1990) The fishery resources of Pacific island countries: part 2. Holothurians. FAO Fisheries Technical Paper 272.2. FAO, Rome.
Fox, J. J., and Sen, S. (2002) A study of the socio-economic issues facing traditional Indonesian fishers who access the MoU Box. Report to Environment Australia, The Australian National University and FERM, Canberra.
Guzman, H. M., and Guevara, C. A. (2002). Population structure, distribution and abundance of three commercial species of sea cucumber (Echinodermata) in Panama. Caribbean Journal of Science 38, 230–238.
Hannah, L., Duprey, N., Blackburn, J., Hand, C. M., and Pearce, C. M. (2012). Growth rate of the California sea cucumber Parastichopus californicus: measurement accuracy and relationships between size and weight metrics. North American Journal of Fisheries Management 32, 167–176.
| Growth rate of the California sea cucumber Parastichopus californicus: measurement accuracy and relationships between size and weight metrics.Crossref | GoogleScholarGoogle Scholar |
Laboy-nieves, E. N., and Conde, J. E. (2006). A new approach for measuring Holothuria mexicana and Isostichopus badionotus for stock assessments. SPC Beche-de-mer Information Bulletin 24, 39–44.
Perry, R. I., Walters, C. J., and Boutillier, J. A. (1999). A framework for providing scientific advice for the management of new and developing invertebrate fisheries. Reviews in Fish Biology and Fisheries 9, 125–150.
| A framework for providing scientific advice for the management of new and developing invertebrate fisheries.Crossref | GoogleScholarGoogle Scholar |
Prescott, J., Vogel, C., Pollock, K., Hyson, S., Oktaviani, D., and Panggabean, A. S. (2013). Estimating sea cucumber abundance and exploitation rates using removal methods. Marine and Freshwater Research 64, 599–608.
| Estimating sea cucumber abundance and exploitation rates using removal methods.Crossref | GoogleScholarGoogle Scholar |
Purcell, S., and Cheng, Y. (2010). Experimental restocking and seasonal visibility of a coral reef gastropod assessed by temporal modelling. Aquatic Biology 9, 227–238.
| Experimental restocking and seasonal visibility of a coral reef gastropod assessed by temporal modelling.Crossref | GoogleScholarGoogle Scholar |
Purcell, S. W., Gossuin, H., and Agudo, N. S. (2009a). Changes in weight and length of sea cucumbers during conversion to processed beche-de-mer: filling gaps for some exploited tropical species. SPC Beche-de-mer Information Bulletin 29, 3–6.
Purcell, S. W., Gossuin, H., and Agudo, N. S. (2009b) Status and management of the sea cucumber fishery of La Grande Terre, New Caledonia. Programme ZoNéCo. WorldFish Center Studies and Reviews 1901. The WorldFish Center, Penang, Malaysia.
Purcell, S. W., Mercier, A., Conand, C., Hamel, J.-F., Toral-Granda, M. V., Lovatelli, A., and Uthicke, S. (2013). Sea cucumber fisheries: global analysis of stocks, management measures and drivers of overfishing. Fish and Fisheries 14, 34–59.
| Sea cucumber fisheries: global analysis of stocks, management measures and drivers of overfishing.Crossref | GoogleScholarGoogle Scholar |
Quinn, T. J., and Deriso, R. B. (1999) ‘Quantitative Fish Dynamics.’ (Oxford University Press: New York.)
Richardson, S., and Best, N. (2003). Bayesian hierarchical models in ecological studies of health-environment effects. Environmetrics 14, 129–147.
| Bayesian hierarchical models in ecological studies of health-environment effects.Crossref | GoogleScholarGoogle Scholar |
Skewes, T., Dennis, D., Donovan, A., and Ellis, N. (2004) ‘Conversion Ratios for Commercial Beche-de-Mer Species in Torres Strait.’ (Australian Fisheries Management Authority, Torres Strait Research Program: Canberra.)
Su, Z., and Peterman, R. M. (2012). Performance of a Bayesian state-space model of semelparous species for stock-recruitment data subject to measurement error. Ecological Modelling 224, 76–89.
| Performance of a Bayesian state-space model of semelparous species for stock-recruitment data subject to measurement error.Crossref | GoogleScholarGoogle Scholar |
Toral-Granda, V., Lovatelli, A., and Vasconcellos, M. (Eds) (2008). Sea cucumbers. A global review on fishery and trade. FAO Fisheries Technical Paper 516. FAO, Rome.
Yamana, Y., and Hamano, T. (2006). New size measurement for the Japanese sea cucumber Apostichopus japonicus (Stichopodidae) estimated from the body length and body breadth. Fisheries Science 72, 585–589.
| New size measurement for the Japanese sea cucumber Apostichopus japonicus (Stichopodidae) estimated from the body length and body breadth.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XmsVOjsbw%3D&md5=e49c8abe7269acdb3544cd3f9a37c37cCAS |
Zhou, S., Yin, S., Thorson, J. T., and Smith, A. D. M. (2012). Linking fishing mortality reference points to life history traits: an empirical study. Canadian Journal of Fisheries and Aquatic Sciences 69, 1292–1301.
| Linking fishing mortality reference points to life history traits: an empirical study.Crossref | GoogleScholarGoogle Scholar |