

Residual forest structure influences behaviour of Pacific marten (Martes caurina) on post-fire landscapes
Logan A. Volkmann A * and Karen E. Hodges AA Department of Biology, University of British Columbia Okanagan, Science Building, 1177 Research Road, Kelowna, BC V1V 1V7, Canada.
International Journal of Wildland Fire 31(4) 329-349 https://doi.org/10.1071/WF21075
Submitted: 29 May 2021 Accepted: 18 February 2022 Published: 6 April 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing on behalf of IAWF. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Wildfires are broad-scale disturbances in North American forests, with impacts that persist for many decades. Further disturbance from post-fire salvage logging is extensively modifying burned landscapes. The removal of habitat structure by fire and salvage logging may affect the persistence of forest-specialist wildlife such as Pacific marten (Martes caurina). However, it is unclear which resources are important to marten on burned landscapes. We used snow tracking and habitat surveys to examine marten habitat selection after three large fires in north-central Washington, USA (10–13 years post-fire), and central British Columbia, Canada (1–2 and 6–9 years post-fire). We developed site-scale habitat models to explain marten foraging and scent-marking post-fire, and assessed further structural changes from salvage logging. Foraging marten chose sites with lower burn severity, greater canopy closure, more vertical structures (trees, snags, saplings, and shrubs), and greater moss/lichen cover than what was generally available. When scent-marking, marten selected structurally-complex sites with abundant deadfall or saplings. Marten moved more quickly when canopy cover was sparse, and rarely used salvage-logged areas. Our results suggest that marten rely on residual habitat structure within large burns, and that secondary disturbance from salvage logging is substantially more harmful to marten than the original fire.
Keywords: carnivores, fire ecology, forest structure, habitat use, landscape management, montane forest, salvage logging, wildfire.
Introduction
Wildfires are increasingly prevalent on North American landscapes. Large fires (>1000 ha) have become more common in conifer forests of Canada and the United States (Hessburg et al. 2005; Miller et al. 2009), and climate change models forecast substantial fire-driven shifts in forest composition and connectivity in the coming decades (McKenzie et al. 2004; Westerling et al. 2011). Fires have long-term consequences for forest-associated birds (Schieck and Song 2006), amphibians (Pilliod et al. 2003), and mammals (Fisher and Wilkinson 2005). Although some wildlife benefit from these disturbances (Fontaine and Kennedy 2012), the resilience of many species has not been determined (Hutchen et al. 2017; Volkmann et al. 2020).
Landscape structure matters to wildlife post-fire (Saab and Powell 2005; Fontaine and Kennedy 2012). Fire initially alters forests by killing trees, opening the canopy, and removing understory features such as groundcover and coarse woody debris (hereafter ‘deadfall’), which often creates a mosaic of lightly-to-severely-burned habitats (Agee 1993, 1998). Burns are therefore heterogeneous in terms of small-scale structures, such as green trees, that wildlife use for foraging (Covert-Bratland et al. 2006; Wood et al. 2007), shelter (Magoun and Vernam 1986; Simanonok and Burkle 2019), and reproduction (Meehan and George 2003; Cunningham and Ballard 2004). Residual structures, such as trees within lightly-burned stands or snags in more heavily-burned areas, influence wildlife distributions on burned landscapes (Barton et al. 2014; Swan et al. 2015). For species that select mature stands (‘forest specialists’), post-fire residuals can provide islands of suitable resources amid low-quality habitat (Robinson et al. 2014; Vanbianchi et al. 2017).
Further disturbances from post-fire salvage logging – the harvest of fire-damaged trees – often removes or reconfigures these residual habitats. Salvage logging is a major component of forest harvest in regions such as British Columbia (BC), where mountain pine beetles (Dendroctonous ponderosae) are projected to kill >50% of merchantable lodgepole pine (Pinus contorta) by the mid-2020s (Dhar et al. 2016), and where record-setting fires burned ~2.5 million ha in 2017–2018 (BC Wildfire Service 2020). However, fire and salvage logging create fundamentally different conditions for wildlife (Franklin et al. 2002; Lindenmayer et al. 2008). Conventional timber harvest influences animal distributions (Hargis et al. 1999; Nelson et al. 2019), home range structure (Lambert and Hannon 2000; Potvin et al. 2000), and habitat connectivity (Cushman et al. 2011; Popescu and Hunter 2011). These effects may be more pronounced on salvage-logged landscapes, where resources were already altered from burning. However, long-term impacts of salvage logging on wildlife remain uncertain (Nappi et al. 2004; Thorn et al. 2018).
Forest specialists such as marten (M. americana and M caurina; Hargis et al. 1999) are sensitive to disturbance and may respond negatively to fire and post-fire salvage logging. These carnivores select specific habitat features at multiple spatial scales (Buskirk and Powell 1994; Thompson et al. 2012), and are indicators of forest health in Canada and the United States (Watt et al. 1996; Guppy 2008). Understanding marten behaviour post-fire would help managers maintain marten populations in fire-prone regions.
Marten occupy burned landscapes (Magoun and Vernam 1986; Latour et al. 1994; Paragi et al. 1996; Volkmann and Hodges 2021), but many details of their post-fire habitat use are unclear. Like other carnivores, marten select habitats based on vegetation (Porter et al. 2005; Seip et al. 2018), prey (Vigeant-Langlois and Desrochers 2011), and conspecifics (Hutchings and White 2000). Identifying spatial changes in marten behaviour can thus provide insight into habitat quality (Powell 2004). For example, marten movements vary with small-scale changes in overhead cover (Nams and Bourgeois 2004; Moriarty et al. 2016). The spatial distribution of marten scent-marks (urine and scat) reflects the distribution of high-quality resources encountered while foraging (Hargis and McCullough 1984; Porter et al. 2005).
In unburned forests, high-quality marten habitats have abundant vertical structure (trees and snags), horizontal structure (uneven-aged stands), understory cover (saplings, shrubs, and deadfall), and ground vegetation (Thompson et al. 2012). Marten avoids meadows and cut-blocks, and choose direct routes across low-quality habitats (Cushman et al. 2011). Trees and snags provide cover for resting and denning (Buskirk et al. 1989; Ruggiero et al. 1998), hunting prey (Coffin et al. 1997; Andruskiw et al. 2008), and avoiding predators (Herman and Fuller 1974). Deadfall creates overwinter habitat for prey such as southern red-backed voles (Myodes gapperi; Sullivan et al. 2011) and shelter for marten (Taylor and Buskirk 1994). Saplings, shrubs, and ground cover offer favourable habitat for snowshoe hares (Lepus americanus; Lewis et al. 2011), northern red-backed voles (Myodes rutilus; Vanderwel et al. 2010), and other small mammals. These habitat features likely matter to marten on post-fire landscapes as well.
We studied marten habitat use in Washington, USA, and BC, Canada, where wildfires have caused substantial landscape changes over the past 15 years (2005–2020). We developed habitat selection models to identify small-scale structural features influencing marten behaviour post-fire, and to assess further impacts of post-fire salvage logging on marten habitat quality. Given the strong links between marten and structurally-complex habitats, we hypothesised that marten would respond to residual structure on burned landscapes. We expected foraging marten to select sites with relatively abundant residual trees, deadfall, understory cover, and ground vegetation. Likewise, we expected marten to move more quickly through sparsely-covered (low-quality) habitats. Finally, we expected marten to scent-mark at sites with relatively abundant deadfall and rocks – structures suitable for broadcasting scent (Hargis and McCullough 1984; Porter et al. 2005). Given the importance of scent for marking resources, we anticipated strong selection for high-quality sites along foraging paths. For both behaviours, we expected marten to avoid salvage-logged areas due to loss of forest structure.
Methods
Study areas
We examined marten populations on post-fire landscapes in north-central Washington (48.790°N, −119.953°W) and central BC (52.071°N, −122.436°W), approximately 400 km apart. We chose study areas representative of regional forest types, management strategies, and disturbance events. Because numerous large, recent fires have occurred in Washington and BC (BC Wildfire Service 2020; National Interagency Fire Center 2020), we also selected landscapes based on overall accessibility.
The 2006 burn (Tripod Complex wildfire, 70 575 ha), is situated within the Okanogan-Wenatchee National Forest (Fig. 1, Table 1). This landscape has rugged topography typical of the northeastern Cascade Range, with alpine parkland above ~2100 m and shrub-steppe communities below ~1000 m (Koehler et al. 2008). Pre-fire, much of the study area was mature, highland conifer forest composed of lodgepole pine, Engelmann spruce (Picea engelmannii), and subalpine fir (Abies lasiocarpa), with Douglas fir (Pseudotsuga menziesii), ponderosa pine (Pinus ponderosa), and quaking aspen (Populus tremuloides) at lower elevations. The regenerating landscape is dominated by snags, deadfall, conifer saplings, and willows (Salix spp.). We sampled elevations of 1500–2000 m.

|

|
The Okanogan-Wenatchee National Forest is not actively managed for timber harvest; a single major access road runs north-south through the study area. Aside from a ~10 m road buffer cleared of hazard trees, the 2006 burn has seen little human disturbance post-fire. State forests east of the burn contain cut-blocks of various ages, but were not sampled in this study.
The 2010 burn (Meldrum Creek wildfire, 15 553 ha) and the 2017 burn (Hanceville-Riske Creek wildfire, 239 340 ha) are situated on the Chilcotin Military Reserve and multi-use (Crown) land near Williams Lake (Fig. 1, Table 1). This landscape has gentle topography at elevations of 900–1000 m, and straddles the Interior Douglas-fir and Sub-Boreal Pine-Spruce biogeoclimatic zones (Hope et al. 1991). Pre-fire, the study area contained mature lodgepole pine, Douglas fir, hybrid Engelmann-white spruce (Picea glauca), and quaking aspen. The regenerating landscape is composed of cut-blocks, natural meadows, snags, and residual trees with a patchy understory of conifer saplings, aspen, and willows. Approximately 15% of the 2010 burn re-burned in 2017.
The BC study area has an extensive road network and a complex history of timber management. Approximately 6000 ha (26%) of the 2010 burn was salvage-logged in large blocks in 2010–2012. Some selective harvest also occurred following post-fire insect outbreaks. Salvage logging has occurred at similar intensity in the 2017 burn, beginning in winter 2017–2018.
Field methods
Snow tracking
In winter (December–March, 2016–2019), we located marten trails weekly along ~75 km of snowmobile-accessible forestry roads chosen ad hoc to cover burned, unburned, and salvage-logged habitats representative of each study area (Volkmann 2021). Suitable trails were 1–4 days old with clearly-identifiable tracks for >50 m. We recorded each trail in straight-line 5 m segments (‘steps’) marked with pin flags and georeferenced at every sixth step (30 m). We also georeferenced all scent-marking (urine or scat) unambiguously associated with these trails. Whenever possible, we back-tracked trails to avoid influencing marten behaviour; forward-tracking was limited to trails >2 days old. We prioritised back-tracking >1 km from previously-sampled trails to maximise our coverage of habitats and marten home ranges.
Marten leave easily-identifiable footprints in snow (see Supplementary material; Elbroch 2003). We counted footprints per 5-m step to index relative movement speed (‘track count’), where a lower track count implies faster movement (Hodges et al. 2014). In some cases only a minimum track count was possible due to poor tracking conditions or subnivean travel. We applied linear interpolation to these segments to determine a most likely track count; missing entries at trail endpoints were assigned a nearest-neighbour value.
Habitat measurements
In summer (July 2017 and July–August 2018 for Washington; April–July 2018 and March 2019 for BC), we documented habitats along marten trails and at randomly-located sites available to marten. For habitats used by marten (‘foraging sites’), we sampled a random subset of 3–6 sites from each trail, excluding sites <20 m from one previously sampled (if a trail doubled back). We also sampled all sites where marten scent-marked (‘marking sites’). To characterise habitats available to marten but not necessarily used, we randomly selected sites on each landscape via ArcMap where no post-fire salvage logging had occurred (‘burned sites’), sites with salvage logging in the 2010 burn (‘salvaged sites’), and sites unaffected by fire for >50 years (‘unburned sites’). To mimic the spatial autocorrelation of marten trails, we sampled available habitats in sets of 3–6 sites separated by a randomly-generated number of 10 m increments (30–200 m) and a randomised bearing; we excluded sites <20 m from previously sampled sites.
At each site, we established a 10 m radius circular plot containing a randomly-oriented 20 m transect for measuring deadfall. For vertical structure, we recorded diameter at breast height (DBH) to the nearest 0.1 cm for all trees and snags within the plot; we counted trees and snags <7.5 cm DBH as saplings, and any snags with a lean angle >45° from vertical as deadfall. For deadfall, we recorded diameter to the nearest 0.1 cm where each piece crossed the transect; we excluded pieces <7.5 cm in diameter at the transect or <1 m long. We determined decay class for each snag and piece of deadfall: (1) dead foliage present, (2) foliage absent but bark intact, (3) bark absent but heartwood intact, or (4) heartwood soft and crumbling (Resources Information Standards Committee (RISC) 2007). We recorded height to the nearest 50 cm (0–200 cm) for each sapling and shrub within a 2 m radius circle at the centre of each site. For groundcover, we visually assessed total grass/forb, moss/lichen, bare soil, and bare rock to the nearest 5%, using three 1 m radius circles centred at 5, 10 and 15 m along the deadfall transect. Finally, we measured canopy closure with a spherical densiometer from the centre of each 10 m plot, averaging four readings.
Analyses
Data preparation
We mapped burn perimeters in ArcMap using shapefiles from the USGS Geosciences and Environmental Change Science Centre (http://rmgsc.cr.usgs.gov/outgoing/GeoMAC) and DataBC (https://catalogue.data.gov.bc.ca/dataset). Burn severity layers were derived from Landsat mapping of near-infrared landscape reflectance at ~12 months post-fire; these datasets came from the Monitoring Trends in Burn Severity project and the BC Ministry of Forests, Lands, Natural Resource Operations and Rural Development. Mapping of the 2006 and 2010 burns used the differenced Normalised Burn Ratio (Key and Benson 2006), and the 2017 burn used the more recently developed Relativised Burn Ratio; in practice the two metrics have a ~2% difference in accuracy (Parks et al. 2014). We analysed burn severity layers as rasters at a 30 × 30 m (0.09 ha) resolution.
To estimate deadfall volume, we used the formula:

where V is deadfall volume in m2/ha, k is a unit conversion constant, a is the lean angle correction factor, c is the slope correction factor, L is the survey transect length, and d is deadfall diameter at the transect. We used k = 1.234 and set both a and c at 1 based on recommended methods for North American montane forests (Woodall and Williams 2007).
Our final dataset contained 28 habitat attributes (Table 2). To maximise the dataset available for models, we used predictive mean matching and polytomous regression with the R package ‘mice’ to impute missing values due to recording errors (van Buuren and Groothuis-Oudshoorn 2011). These entries represented <3% of the original dataset and were deemed to be missing at random. We converted continuous variables representing DBH, diameter, decay class, and height class into categorical variables to capture biologically meaningful missing data – for example, a treeless site would have no entry for tree DBH on a continuous scale, but this result is still relevant for marten habitat use. For each attribute, we used the R package ‘boot’ to bootstrap means and 95% confidence intervals with the bias-corrected and accelerated method (Canty and Ripley 2019). Each bootstrap procedure took 10 000 replicates. We pooled habitat data for all years.

|
Habitat influences on marten movement
We built generalised linear mixed models with the R package ‘MASS’ (Venables and Ripley 2002) to determine whether heterogeneous cover affected marten movements. Marten trail characteristics vary in response to local habitat quality, particularly small-scale differences in overhead cover (Moriarty et al. 2016; Slauson et al. 2017). Thus, we expected faster movement (fewer tracks per 5 m) in open areas relative to closed-canopy areas along marten trails. For our models, canopy closure was a fixed-effect predictor of movement speed, and year/trail ID were nested random predictors. We used a Penalised Quasi-Likelihood model fit with an identity link based on inspection of quantile-comparison plots, since track count distributions showed a strong positive skew (Wolfinger and O’Connell 1993). We tested for differences in track counts among burns using an analysis of deviance against a null model of burn ID ~ 1.
Habitat influences on site selection
For each burn, we built generalised linear models to separately examine marten selection of foraging and marking (‘used’) sites compared to burned (‘available’) sites. We a priori excluded salvaged and unburned sites from models because the 2006 and 2017 burns lacked salvage-logged areas, and because marten in the 2010 burn avoided salvage-blocks (Volkmann and Hodges 2021). We pooled habitat data for all years. To reduce the likelihood of nonsense models, we first screened for individually insignificant habitat attributes using single-factor logistic regressions for continuous variables and Fisher’s exact test for categorical variables. We then used the R package ‘Hmisc’ to calculate Pearson correlations for habitat attributes in each dataset (Harrell and Dupont 2020). From correlograms, we identified a subset of individually significant habitat attributes with pairwise correlations of r2 < 0.25; this relatively strict cutoff ensured that only <10 strongly independent variables were retained for each model run (Table 2; Schober et al. 2018).
We applied multi-model inference with the R package ‘glmulti’ to build and rank multivariate logistic regressions linking habitat attributes to marten foraging and marking sites, and to assess the individual importance of each attribute across candidate models (Calcagno 2019). We used differences in Akaike’s Information Criterion (ΔAIC), IC weight, and model diagnostics to judge model performance, retaining models with ΔAIC < 2 and individual variables occurring in >50% of models (Anderson and Burnham 2002). We estimated effect sizes using Cohen’s d. Finally, to assess the quality of salvage-logged areas for marten, we compared habitat attributes of foraging and marking sites in the 2010 burn to those of salvaged and unburned sites, using two-tailed Welch’s t-tests.
Results
Trail characteristics
We back-tracked 100 marten trails (65.5 km total): 50 trails (24.3 km) in the 2006 burn, 33 trails (25.2 km) in the 2010 burn, and 19 trails (16.0 km) in the 2017 burn. Individual trails were 70–2200 m long with 42–440 5-m steps. We recorded 70 scent-marks: 46 sites with urine, 21 with scat, and 3 with both.
We measured track counts from 2317 trail segments: 871 in the 2006 burn, 888 in the 2010 burn, and 558 in the 2017 burn (Fig. 2). Mean track counts were 10.2 ± 0.2 per 5-m step in the 2006 burn, 9.9 ± 0.2 in 2010 burn, and 10.5 ± 0.3 in the 2017 burn; track counts did not differ among burns (residual deviance = 229.52, d.f. = 2314, P = 0.517) or across years within each burn. Track counts varied with canopy closure in the 2006 burn (est. = 0.020 ± 0.005, t820 = 4.22, P < 0.001) and the 2010 burn (est. = 0.015 ± 0.004, t854 = 3.48, P < 0.001). Marten movement speed increased 15–20% overall as canopy closure decreased. We did not see this behavioural shift in the 2017 burn (est. = 0.009 ± 0.006, t538 = 1.59, P = 0.119).
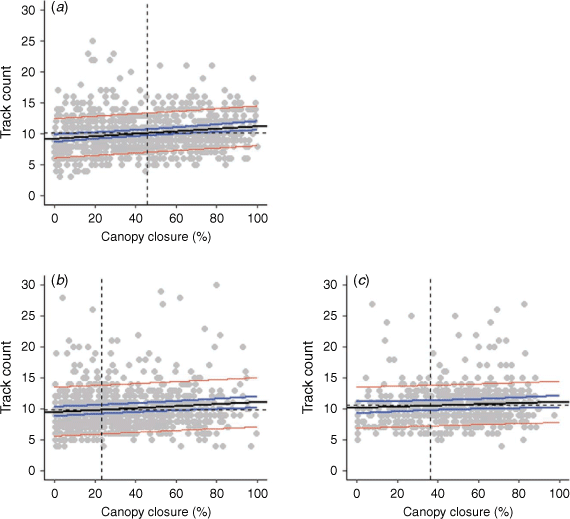
|
Site selection
We surveyed habitats at 476 sites in Washington and 995 sites in BC. Marten responded to both overstory and understory habitat attributes when foraging (Table 3) and scent-marking (Table 4), and had stronger patterns of selection in the 2006 and 2017 burns compared to the 2010 burn (see Supplementary material). Foraging marten consistently selected mesic sites with greater moss/lichen cover, less bare soil, and less-decayed snags. Marten scent-marks were associated with sheltered sites with greater canopy closure and a greater basal area of trees. Neither behaviour was strongly influenced by the presence of large trees or snags.

|

|
Foraging sites
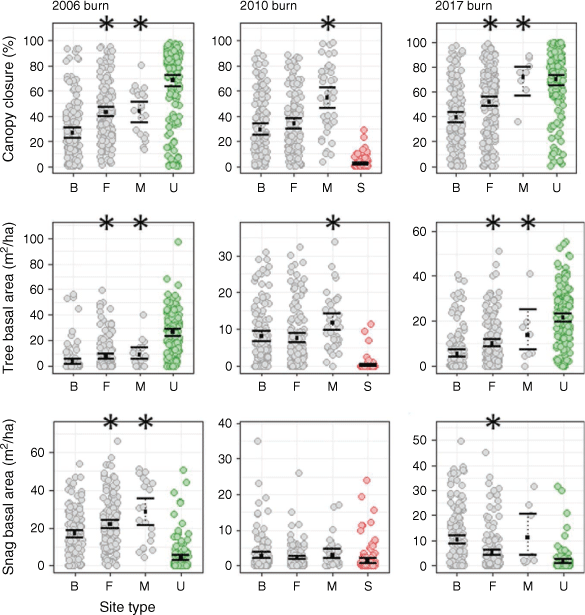
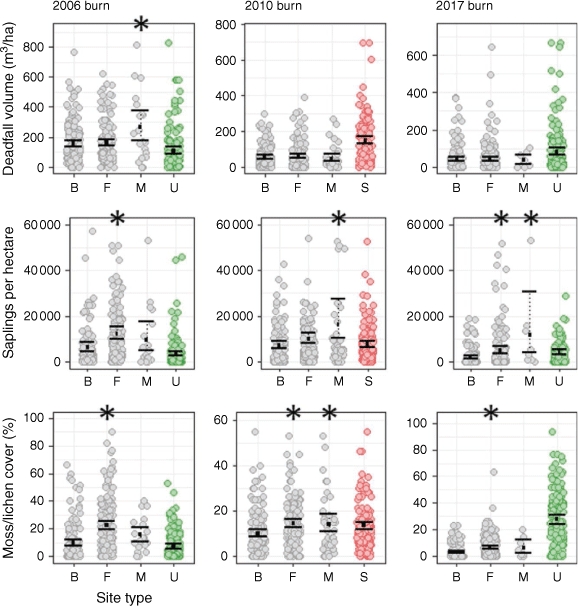
In the 2006 and 2017 burns, foraging marten selected relatively intact sites with lower burn severity, greater canopy closure, and more trees compared to availability; selection was weaker in the 2010 burn (Table 3). Ten habitat attributes best explained marten foraging post-fire (Table 5). In the 2006 burn, marten chose foraging sites with lower burn severity (X2 = 27.51, d.f. = 3, P < 0.001), a greater basal area of snags (21.9 [19.7–24.1] vs 16.9 [14.9–19.0] m2/ha), more vertical structures (1035 [918–1160] vs 622 [528–750] per ha), and greater moss/lichen cover (22.5 [19.7–25.7] vs 9.5 [7.5–12.1]%; Figs 3, 4; see Supplementary material). Marten in the 2010 burn similarly chose foraging sites with greater moss/lichen cover (14.5 [12.8–16.3] vs 10.1 [8.6–11.9]%), along with more saplings (9937 [8146–12 775] vs 7042 [5771–8966] per ha) and fewer shrubs (15 389 [13 215–17 847] vs 19 397 [16 293–23 351] per ha). Marten in the 2017 burn chose foraging sites with greater canopy closure (52.1 [48.4–55.9] vs 39.3 [35.5–43.5]%), fewer snags (300 [266–338] vs 463 [402–530] per ha), more saplings (4726 [3551–6642] vs 1870 [1366–2575] per ha), and less bare rock (1.9 [1.5–2.5] vs 4.1 [3.3–5.1]%).

|
|
|
Marten foraging sites in burns differed from unburned areas in terms of habitat structure. Compared to unburned sites, marten in the 2006 burn selected foraging sites with a greater basal area of snags (21.9 [19.7–24.1] vs 3.9 [2.8–5.6] m2/ha), more saplings (12 027 [9834–15 337] vs 3354 [2520–4647] per ha), and greater moss/lichen cover (22.5 [19.7–25.7] vs 7.1 [5.7–8.9]%; Fig. 3, 4; see Supplementary material). Marten in the 2010 burn similarly chose foraging sites with more saplings (9937 [8146–12 775] vs 3842 [3048–5487] per ha) than at unburned sites (but less moss/lichen cover – 14.5 [12.8–16.3] vs 27.7 [24.3–31.2]%), while marten in the 2017 burn chose foraging sites with a greater basal area of snags (5.0 [4.1–6.3] vs 1.5 [0.9–2.4] m2/ha) and less moss lichen cover (6.7 [5.7–8.0] vs 27.7 [24.3–31.2]%). Foraging sites in all three burns had significantly less canopy closure compared to unburned sites.
Marking sites
Marten in the 2006 and 2010 burns chose structurally-complex sites for scent-marking, with greater canopy closure and more vertical structures; selection was weaker in the 2017 burn, likely because of low sample sizes (Table 4). Marten chose marking sites with abundant deadfall in the 2006 burn, but avoided bare rock in the 2006 and 2010 burns. Eight habitat attributes best explained marten scent-marking post-fire (Table 6). Marten in the 2006 burn chose marking sites with lower burn severity (X2 = 11.25, d.f. = 3, P = 0.01), a greater basal area of vertical structures (37.4 [30.6–43.4] vs 20.0 [17.6–22.7] m2/ha), and greater deadfall volume (263.5 [177.6–375.2] vs 153.4 [132.8–178.4] m3/ha; Fig. 3, 4; see Supplementary material). In the 2010 burn, marten chose marking sites with greater canopy closure (54.8 [46.7–62.7] vs 29.3 [25.3–33.6]%), more snags (224 [173–292] vs 119 [98–145] per ha) and saplings (16 374 [10 186–27 477] vs 7042 [5771–8966] per ha), and less bare rock (2.5 [1.3–4.2] vs 6.8 [5.4–8.4]%). Similarly, marten in the 2017 burn chose marking sites with greater canopy closure (71.8 [56.6–80.0] vs 39.3 [35.5–43.5]%), more saplings (11 597 [4078–32 428] vs 1870 [1366–2575] per ha), and less bare soil (30.3 [20.0–54.0] vs 54.2 [50.1–58.2]%).

|
Marking sites in burns also differed from unburned areas in terms of habitat structure. Compared to unburned sites, marten in the 2006 burn selected marking sites with a greater snag basal area (28.2 [21.2–35.4] vs 3.9 [2.8–5.6] m2/ha) and greater deadfall volume (263.5 [177.6–375.2] vs 109.2 [90.1–134.8] m3/ha; Fig. 3, 4; see Supplementary material). In the 2010 burn, marten chose marking sites with a greater snag basal area (3.1 [2.2–4.6] vs 1.5 [0.9–2.4] m2/ha) and more saplings (16 374 [10 186–27 477] vs 3842 [3048–5487] per ha). Marking sites in the 2010 and 2017 burns had significantly less deadfall volume compared to unburned sites.
Salvaged sites
In the 2010 burn, marten selected habitats substantially different from post-fire salvage-logged areas (Fig. 3, 4). Marten foraged at sites with significantly greater canopy closure (33.7 [29.6–37.8] vs 1.9 [1.4–2.8]%), more trees (258 [221–301] vs 5 [2–10] per ha) and snags (132 [113–154] vs 26 [20–34] per ha), and greater grass/forb cover (65.8 [63.0–68.5] vs 52.6 [50.1–55.0]%) than what was available at salvaged sites (Table 7; see Supplementary material). Scent-marking sites had significantly higher sapling density (16 374 [10 186–27 477] vs 7535 [6414–9067] per ha) compared to salvaged sites. Conversely, marten foraged at sites with significantly less deadfall (60.1 [49.0–73.7] vs 146.2 [128.3–167.2] m3/ha) and less bare rock (4.9 [3.8–6.4] vs 11.5 [9.8–13.6]%) compared to salvaged sites.

|
Discussion
Marten responses to fire
Residual forest structure matters to marten post-fire. In Washington and BC, marten altered their movements in response to canopy closure. Marten selected structurally-complex habitats in burns, foraging and scent-marking at sites with closed canopies, abundant vertical structures, and dense regeneration. These features correspond to lightly-burned stands less altered by fire.
Previous work hinted at the importance of structure for marten post-fire. Marten evolved with wildfire and probably benefit from the shifting mosaic of resources on burned landscapes (Koehler and Hornocker 1977; Fisher and Wilkinson 2005). Trappers in Alaska associated marten with deadfall and shrubs <3 years post-fire (Stephenson 1984). Radio-collared marten used an Alaskan burn extensively at 7 years post-fire, selecting sites with abundant trees and deadfall (Magoun and Vernam 1986). Our data from landscapes burned in 2006 (10–13 years post-fire), 2010 (6–9 years post-fire) and 2017 (1–2 years post-fire) broadly agree with these earlier studies.
Marten in the 2006 and 2017 burns selected lightly-burned sites (‘unchanged’ to ‘low’ burn severity) over severely-burned ones. Prior studies did not link marten behaviour to burn severity (Volkmann et al. 2020), but our results resemble observations of other forest specialists including Canada lynx (Lynx canadensis; Vanbianchi et al. 2017), squirrels (Sciurus arizonensis and S. aberti; Ketcham et al. 2017) three-toed woodpeckers (Picoides dorsalis; Murphy and Lenhausen 1998), spotted owls (Strix occidentalis; Eyes et al. 2017), and saproxylic insects (Nappi et al. 2010). Since burn severity relates to tree mortality (Key and Benson 2006), lightly-burned sites often resemble pre-fire conditions in terms of canopy closure, tree abundance, and understory structure.
Vertical structure influenced marten behaviour post-fire. Marten avoided open areas, consistent with these habitats having greater exposure to predators (Herman and Fuller 1974), fewer preferred prey (Hutchen and Hodges 2019; Kelly and Hodges 2020), and fewer subnivean access points (Corn and Raphael 1992). Relative importance of residual trees and snags differed among landscapes. Foraging marten selected for abundant snags in the 2006 burn, had no clear patterns of use in the 2010 burn, and chose densely treed sites with few snags in the 2017 burn. Snags provide less cover for marten, and prey such as red squirrels (Tamiasciurus hudsonicus) are more abundant in treed stands (Allard-Duchêne et al. 2014; Kelly and Hodges 2020). Marten in the 2006 burn may have used snag-rich stands as movement corridors, weakening the selection for trees that we expected (Volkmann and Hodges 2021). Alternatively, subnivean access from snags and a regenerating understory may have supported more prey in the 2006 burn.
Marten also responded to understory features, choosing sites with more saplings and moss/lichen cover. Marten select similar mesic features on other landscapes (Bull et al. 2005; Proulx et al. 2006). Mesic sites regenerate more rapidly post-fire (Dwire and Kaufmann 2003; Halofsky and Hibbs 2009), creating structural complexity sooner for marten and prey. Understory complexity benefits snowshoe hares (Hutchen and Hodges 2019) and meadow voles (Microtus pennsylvanicus; Sullivan et al. 2000), which marten eat when red-backed voles are scarce (Paragi et al. 1996).
Combined understory and overstory features influenced marten scent-marking. Marten chose marking sites with greater coverage from trees and saplings. Structurally-complex sites with abundant deadfall were important to marten in the 2006 burn, but use of deadfall in the 2010 and 2017 burns was ambiguous. Mustelids use scent to signal the presence of prey (Hutchings and White 2000). Although marten frequently investigated deadfall in subnivean forays, we could not determine prey abundance at these sites. Marten also avoided bare rock when scent-marking, contrary to expectations (Hargis and McCullough 1984). Elevated deadfall and boulders help broadcast scent (Pulliainen 1982; Porter et al. 2005), but these structures may have been too low or scarce for selection in BC, compared to densely interlocking deadfall in the 2006 burn (Magoun and Vernam 1986).
Marten showed weaker habitat selection in the 2010 burn compared to the 2006 and 2017 burns, implying greater similarity between used and available sites. We suspect that this similarity was due to salvage logging homogenising large areas of the burn. In effect, marten in the 2010 burn had fewer suitable post-fire habitats (Volkmann and Hodges 2021).
Surprisingly, we did not detect marten in unburned forests bordering either study area. Marten similarly selected a 6-year-old burn over mature forest in Alaska, likely due to differences in habitat structure and prey abundance (Paragi et al. 1996). Prey densities in unburned and lightly-burned areas of the 2010 burn were similar during our study (Kelly and Hodges 2020; Kelly 2021); this combined with greater snag density may have offered better habitat in the 2010 burn (Payer and Harrison 2003; Bull et al. 2005). Alternatively, marten may have chosen burns to avoid larger carnivores (fishers Pekania pennanti and lynx; Volkmann 2021). A broader survey of burned and unburned landscapes would help determine how these factors influence marten habitat use.
Impacts of post-fire salvage logging on wildlife
Post-fire salvage logging severely degraded marten habitat. Marten in the 2010 burn chose sites with substantially more structural complexity than salvaged sites. When crossing open areas, marten took direct routes between available trees and saplings, and rarely approached salvage-blocks (Volkmann and Hodges 2021). Although deadfall was abundant at salvaged sites, these structures were usually stripped of branches and flush with the ground, making subnivean access difficult (Wilbert et al. 2000). Notably, salvage-blocks in the 2010 burn also lacked debris piles to provide cover for marten and microtine prey (Sullivan and Sullivan 2021; Sullivan et al. 2021).
Marten have not been directly studied on post-fire salvage-logged landscapes. Indirectly, Steventon and Daust (2009) modelled marten habitat in response to post-insect salvage logging in BC and forecast substantial marten declines in the next 20–40 years, even with lower quotas for conventional timber harvest. More recent work suggests that marten use residual structures in salvage blocks for resting and denning, but these features are a poor long-term substitute for forest cover (Wilk and Raphael 2018). GPS-tracked marten in the 2010 burn avoided salvage-blocks (Volkmann and Hodges 2021), and the relative abundances of both marten (Volkmann 2021) and preferred prey (Kelly and Hodges 2020; Kelly 2021) were markedly lower in salvage-logged areas. Given that marten avoid meadows (Spencer et al. 1983) and conventional clear-cuts (Soutiere 1979), avoidance of post-fire salvage-logged areas is unsurprising.
Wildfire and post-fire salvage logging are not equivalent disturbances for marten, and create different landscapes (Fig. 5). Salvage logging homogenises burned forests, removing critical resources for marten and prey (Kelly and Hodges 2020; Kelly 2021). Loss of structures suitable for foraging, resting, and denning will likely hinder marten persistence on salvage-logged landscapes. Female marten are particularly intolerant of disturbance (Cheveau et al. 2013; Slauson et al. 2017) and require specific structures for reproduction (Wynne and Sherburne 1984; Ruggiero et al. 1998). For marten, post-fire salvage logging has secondary impacts more severe than high-severity wildfire.

|
Landscape alterations from salvage logging threaten marten and other forest specialists. Salvage-blocks support fewer snowshoe hares (Thomas et al. 2019; Kelly and Hodges 2020), an important prey for many carnivores including fishers (Powell 1979; Raine 1987). Like marten, female fishers require large trees and snags for denning (Weir et al. 2012). Individual snags can persist for >80 years post-fire (Chambers and Mast 2005; Russell et al. 2006), particularly in severely-burned areas (Angers et al. 2011), and provide both structural complexity and a long-term source of deadfall on regenerating landscapes. Sparse, evenly-spaced snags left by salvage logging are less suitable to black-backed woodpeckers (Picoides arcticus) compared to undisturbed stands (Hutto and Gallo 2006; Saab et al. 2009), prompting specific snag-retention guidelines for this species (Nappi and Drapeau 2009, 2011). Our results underscore the need for clearer retention guidelines to support other species on salvage-logged landscapes (Thorn et al. 2020).
Climate change, fire regimes, and wildlife conservation
Climate change is promoting larger, more severe fires in North American forests (McKenzie et al. 2004; Girardin and Mudelsee 2008). Many fire regimes are also shifting towards more frequent disturbance (Kasischke and Turetsky 2006; Westerling et al. 2006; Dennison et al. 2014), which may permanently alter forested landscapes (Adams 2013; Busby et al. 2020). Biodiversity losses from salvage logging are occurring in Australian forests (Lindenmayer et al. 2008; Blair et al. 2016), but fewer data exist for North American wildlife despite mounting pressure to harvest in burns. Amid rapid landscape change, it is critical to identify post-fire habitat features that wildlife need (Hutchen et al. 2017; Volkmann et al. 2020) and reassess forest management strategies accordingly (Hutto 2006; Millar et al. 2007; Lindenmayer et al. 2014).
For marten, we recommend that managers prioritise the retention of residual live trees, corresponding to areas burned at low to moderate severity. Future work should determine how snags and deadfall influence habitat quality and landscape connectivity in severely-burned areas (Seip et al. 2018; Sullivan and Sullivan 2021), and the extent to which debris piles improve salvage-logged areas for marten and prey (Wilk and Raphael 2018; Sullivan et al. 2021). Because post-fire landscapes are diverse and complex, we call on researchers to study a broader range of wildfire ages, severities, and sizes (Volkmann et al. 2020). We also recommend linking post-fire habitat selection with prey abundance (Coffin et al. 1997). More data on wildfire and salvage logging will improve our ability to maintain suitable landscapes for wildlife.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
This project was supported by grants from the Habitat Conservation Trust Foundation and the Forest Enhancement Society of British Columbia (Project #8-400), the Skagit Wildlife Research Grant Program (City Light Grant #2016-01), the Natural Sciences and Engineering Research Council of Canada (Discovery Grant #31222), and by fellowships from Killam Trusts and the University of British Columbia.
Supplementary material
Supplementary material is available online.
Acknowledgements
We thank our field assistants Sarah Bird, Taylor Brannock, Jeff Brown, Jordan Cormack, Matt Danielson, Anna Heiter, Angie Kelly, Daniel Laszlo, Sara Laursen, Kelly Likos, Erin Morrison, Laura Platt, Sam Shappas, and Alec Spencer. We are grateful to the Department of National Defence for providing safety training and access to the Chilcotin Military Reserve in BC. We thank Becky Cadsand, Mauro Calabrese, Larry Davis, Jeff Lewis, Matt Marsh, and Rich Weir for their local knowledge and guidance on this project.
References
Adams MA (2013) Mega-fires, tipping points and ecosystem services: managing forests and woodlands in an uncertain future. Forest Ecology and Management 294, 250–261.| Mega-fires, tipping points and ecosystem services: managing forests and woodlands in an uncertain future.Crossref | GoogleScholarGoogle Scholar |
Agee JK (Ed.) (1993) ‘Fire ecology of Pacific Northwest forests.’ (Island Press: Washington, DC)
Agee JK (1998) The landscape ecology of western forest fire regimes. Northwest Science 72, 24–34.
Allard-Duchêne A, Pothier D, Dupuch A, Fortin D (2014) Temporal changes in habitat use by snowshoe hares and red squirrels during post-fire and post-logging forest succession. Forest Ecology and Management 313, 17–25.
| Temporal changes in habitat use by snowshoe hares and red squirrels during post-fire and post-logging forest succession.Crossref | GoogleScholarGoogle Scholar |
Anderson DR, Burnham KP (2002) Avoiding pitfalls when using information-theoretic methods. Journal of Wildlife Management 66, 912–918.
| Avoiding pitfalls when using information-theoretic methods.Crossref | GoogleScholarGoogle Scholar |
Andruskiw M, Fryxell JM, Thompson ID, Baker JA (2008) Habitat-mediated variation in predation risk by the American marten. Ecology 89, 2273–2280.
| Habitat-mediated variation in predation risk by the American marten.Crossref | GoogleScholarGoogle Scholar | 18724737PubMed |
Angers VA, Gauthier S, Drapeau P, Jayen K, Bergeron Y (2011) Tree mortality and snag dynamics in North American boreal tree species after a wildfire: a long-term study. International Journal of Wildland Fire 20, 751–763.
| Tree mortality and snag dynamics in North American boreal tree species after a wildfire: a long-term study.Crossref | GoogleScholarGoogle Scholar |
Barton PS, Ikin K, Smith AL, MacGregor C, Lindenmayer DB (2014) Vegetation structure moderates the effect of fire on bird assemblages in a heterogeneous landscape. Landscape Ecology 29, 703–714.
| Vegetation structure moderates the effect of fire on bird assemblages in a heterogeneous landscape.Crossref | GoogleScholarGoogle Scholar |
BC Wildfire Service (2020) Wildfire Season Summary. Available at https://www2.gov.bc.ca/gov/content/safety/wildfire-status/about-bcws/wildfire-history/wildfire-season-summary [Verified 25 May 2021]
Blair DP, McBurney LM, Blanchard W, Banks SC, Lindenmayer DB (2016) Disturbance gradient shows logging affects plant functional groups more than fire. Ecological Applications 26, 2280–2301.
| Disturbance gradient shows logging affects plant functional groups more than fire.Crossref | GoogleScholarGoogle Scholar | 27755744PubMed |
Bull EL, Heater TW, Shepherd JF (2005) Habitat selection by the American marten in northeastern Oregon. Northwest Science 79, 37–43.
Busby SU, Moffett KB, Holz A (2020) High‐severity and short‐interval wildfires limit forest recovery in the Central Cascade Range. Ecosphere 11, e03247
| High‐severity and short‐interval wildfires limit forest recovery in the Central Cascade Range.Crossref | GoogleScholarGoogle Scholar |
Buskirk SW, Powell RA (1994) Habitat ecology of fishers and American martens. In ‘Martens, sables, and fishers: biology and conservation’. (Eds Buskirk SW, Harestad AS, Raphael MG, Powell RA) pp. 283–296. (Cornell University Press: Ithaca, NY)
Buskirk SW, Forrest SC, Raphael MG, Harlow HJ (1989) Winter resting site ecology of marten in the central Rocky Mountains. Journal of Wildlife Management 53, 191–196.
| Winter resting site ecology of marten in the central Rocky Mountains.Crossref | GoogleScholarGoogle Scholar |
Calcagno V (2019) glmulti: Model Selection and Multimodel Inference Made Easy. R package version 1.0.8. Available at https://cran.r-project.org/web/packages/glmulti/index.html [Verified 25 May 2021]
Canty A, Ripley BD (2019) boot: Bootstrap R (S-Plus) Functions. R package version 1.3-28. Available at https://cran.r-project.org/web/packages/boot/index.html [Verified 25 May 2021]
Chambers CL, Mast JN (2005) Ponderosa pine snag dynamics and cavity excavation following wildfire in northern Arizona. Forest Ecology and Management 216, 227–240.
| Ponderosa pine snag dynamics and cavity excavation following wildfire in northern Arizona.Crossref | GoogleScholarGoogle Scholar |
Cheveau M, Imbeau L, Drapeau P, Belanger L (2013) Marten space use and habitat selection in managed coniferous boreal forests of eastern Canada. Journal of Wildlife Management 77, 749–760.
| Marten space use and habitat selection in managed coniferous boreal forests of eastern Canada.Crossref | GoogleScholarGoogle Scholar |
Coffin CW, Kujala QJ, Douglass RJ, Irby LR (1997) Interactions among marten prey availability, vulnerability, and habitat structure. In ‘Martes: taxonomy, ecology, techniques, and management’. (Eds Proulx G, Bryant HN, Woodward PM) pp. 199–210. (Provincial Museum of Alberta: Edmonton, AB)
Corn JG, Raphael MG (1992) Habitat characteristics at marten subnivean access sites. Journal of Wildlife Management 56, 442–448.
| Habitat characteristics at marten subnivean access sites.Crossref | GoogleScholarGoogle Scholar |
Covert-Bratland KA, Block WM, Theimer TC (2006) Hairy woodpecker winter ecology in ponderosa pine forests representing different ages since wildfire. Journal of Wildlife Management 70, 1379–1392.
| Hairy woodpecker winter ecology in ponderosa pine forests representing different ages since wildfire.Crossref | GoogleScholarGoogle Scholar |
Cunningham SC, Ballard WB (2004) Effects of wildfire on black bear demographics in central Arizona. Wildlife Society Bulletin 32, 928–937.
| Effects of wildfire on black bear demographics in central Arizona.Crossref | GoogleScholarGoogle Scholar |
Cushman SA, Raphael MG, Ruggiero LF, Shirk AS, Wasserman TN, O’Doherty EC (2011) Limiting factors and landscape connectivity: the American marten in the Rocky Mountains. Landscape Ecology 26, 1137–1149.
| Limiting factors and landscape connectivity: the American marten in the Rocky Mountains.Crossref | GoogleScholarGoogle Scholar |
Dennison PE, Brewer SC, Arnold JD, Moritz MA (2014) Large wildfire trends in the western United States, 1984-2011. Geophysical Research Letters 41, 2928–2933.
| Large wildfire trends in the western United States, 1984-2011.Crossref | GoogleScholarGoogle Scholar |
Dhar A, Parrott L, Hawkins CDB (2016) Aftermath of mountain pine beetle outbreak in British Columbia: stand dynamics, management response and ecosystem resilience. Forests 7, 171
| Aftermath of mountain pine beetle outbreak in British Columbia: stand dynamics, management response and ecosystem resilience.Crossref | GoogleScholarGoogle Scholar |
Dwire KA, Kauffman JB (2003) Fire and riparian ecosystems in landscapes of the western USA. Forest Ecology and Management 178, 61–74.
| Fire and riparian ecosystems in landscapes of the western USA.Crossref | GoogleScholarGoogle Scholar |
Elbroch M (Ed.) (2003) ‘Mammal tracks and sign: a guide to North American species.’ (Stackpole Books: Mechanicsburg, PA)
Eyes SA, Roberts SL, Johnson MD (2017) California spotted owl (Strix occidentalis occidentalis) habitat use patterns in a burned landscape. The Condor 119, 375–388.
| California spotted owl (Strix occidentalis occidentalis) habitat use patterns in a burned landscape.Crossref | GoogleScholarGoogle Scholar |
Fisher JT, Wilkinson L (2005) The response of mammals to forest fire and timber harvest in the North American boreal forest. Mammal Review 35, 51–81.
| The response of mammals to forest fire and timber harvest in the North American boreal forest.Crossref | GoogleScholarGoogle Scholar |
Fontaine JB, Kennedy PL (2012) Meta-analysis of avian and small-mammal response to fire severity and fire surrogate treatments in US fire-prone forests. Ecological Applications 22, 1547–1561.
| Meta-analysis of avian and small-mammal response to fire severity and fire surrogate treatments in US fire-prone forests.Crossref | GoogleScholarGoogle Scholar | 22908713PubMed |
Franklin JF, Spies TA, Van Pelt R, Carey AB, Thornburgh DA, Berg DR, Lindenmayer DB, Harmon ME, Keeton WS, Shaw DC, Bible K, Chen J (2002) Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. Forest Ecology and Management 155, 399–423.
| Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example.Crossref | GoogleScholarGoogle Scholar |
Girardin MP, Mudelsee M (2008) Past and future changes in Canadian boreal wildfire activity. Ecological Applications 18, 391–406.
| Past and future changes in Canadian boreal wildfire activity.Crossref | GoogleScholarGoogle Scholar | 18488604PubMed |
Guppy CS (2008) Guide to species of management concern. BC Ministry of Forests and Range, Report prepared for BC Timber Sales, Skeena Business Area. (Terrace, BC)
Halofsky JE, Hibbs DE (2009) Controls on early post-fire woody plant colonization in riparian areas. Forest Ecology and Management 258, 1350–1358.
| Controls on early post-fire woody plant colonization in riparian areas.Crossref | GoogleScholarGoogle Scholar |
Hargis CD, McCullough DR (1984) Winter diet and habitat selection of marten in Yosemite National Park. Journal of Wildlife Management 48, 140–146.
| Winter diet and habitat selection of marten in Yosemite National Park.Crossref | GoogleScholarGoogle Scholar |
Hargis CD, Bissonette JA, Turner DL (1999) The influence of forest fragmentation and landscape pattern on American martens. Journal of Applied Ecology 36, 157–172.
| The influence of forest fragmentation and landscape pattern on American martens.Crossref | GoogleScholarGoogle Scholar |
Harrell FE, Dupont C (2020) Hmisc: Harrell Miscellaneous. R package version 4.5-0. Available at https://cran.r-project.org/web/packages/Hmisc/index.html [Verified 25 May 2021]
Herman T, Fuller K (1974) Observations of the marten, Martes americana, in the Mackenzie District, Northwest Territories. Canadian Field-Naturalist 88, 501–503.
Hessburg PF, Agee JK, Franklin JF (2005) Dry forests and wildland fires of the inland Northwest USA: contrasting the landscape ecology of the pre-settlement and modern eras. Forest Ecology and Management 211, 117–139.
| Dry forests and wildland fires of the inland Northwest USA: contrasting the landscape ecology of the pre-settlement and modern eras.Crossref | GoogleScholarGoogle Scholar |
Hodges KE, Cunningham JAF, Mills LS (2014) Avoiding and escaping predators: movement tortuosity of snowshoe hares in risky habitats. Ecoscience 21, 97–103.
| Avoiding and escaping predators: movement tortuosity of snowshoe hares in risky habitats.Crossref | GoogleScholarGoogle Scholar |
Hope GD, Mitchell WR, Lloyd DA, Erickson WR, Harper WL, Wikeem BM (1991) Interior Douglas-fir zone. In ‘Ecosystems of British Columbia’. Special Report Series 06. (Eds D Meidinger, J Pojar) pp. 153–166. (BC Ministry of Forests: Victoria, BC)
Hutchen J, Hodges KE (2019) Impact of wildfire size on snowshoe hare relative abundance in southern British Columbia, Canada. Fire Ecology 15, 37
| Impact of wildfire size on snowshoe hare relative abundance in southern British Columbia, Canada.Crossref | GoogleScholarGoogle Scholar |
Hutchen J, Volkmann LA, Hodges KE (2017) Experimental designs for studying small-mammal responses to fire in North American conifer forests. International Journal of Wildland Fire 26, 523–531.
| Experimental designs for studying small-mammal responses to fire in North American conifer forests.Crossref | GoogleScholarGoogle Scholar |
Hutchings MR, White PCL (2000) Mustelid scent-marking in managed ecosystems: implications for population management. Mammal Review 30, 157–169.
| Mustelid scent-marking in managed ecosystems: implications for population management.Crossref | GoogleScholarGoogle Scholar |
Hutto RL (2006) Toward meaningful snag‐management guidelines for postfire salvage logging in North American conifer forests. Conservation Biology 20, 984–993.
| Toward meaningful snag‐management guidelines for postfire salvage logging in North American conifer forests.Crossref | GoogleScholarGoogle Scholar | 16922215PubMed |
Hutto RL, Gallo SM (2006) The effects of postfire salvage logging on cavity-nesting birds. The Condor 108, 817–831.
| The effects of postfire salvage logging on cavity-nesting birds.Crossref | GoogleScholarGoogle Scholar |
Kasischke ES, Turetsky MR (2006) Recent changes in the fire regime across the North American boreal region—spatial and temporal patterns of burning across Canada and Alaska. Geophysical Research Letters 33, L09703
| Recent changes in the fire regime across the North American boreal region—spatial and temporal patterns of burning across Canada and Alaska.Crossref | GoogleScholarGoogle Scholar |
Kelly A (2021) Small mammals and mesomammals in a post-fire and salvage-logged landscape. MSc thesis, University of British Columbia Okanagan, Kelowna, BC.
Kelly AJ, Hodges KE (2020) Post-fire salvage logging reduces snowshoe hare and red squirrel densities in early seral stages. Forest Ecology and Management 473, 118272
| Post-fire salvage logging reduces snowshoe hare and red squirrel densities in early seral stages.Crossref | GoogleScholarGoogle Scholar |
Ketcham SL, Koprowski JL, Falk DA (2017) Differential response of native Arizona gray squirrels and introduced Abert’s squirrels to a mosaic of burn severities. Mammal Study 42, 247–258.
| Differential response of native Arizona gray squirrels and introduced Abert’s squirrels to a mosaic of burn severities.Crossref | GoogleScholarGoogle Scholar |
Key CH, Benson NC (2006) Landscape Assessment: ground measure of severity, the Composite Burn Index; and remote sensing of severity, the Normalized Burn Ratio. In ‘FIREMON: Fire Effects Monitoring and Inventory System’. General Technical Report RMRS-GTR-164-CD: LA 1-51. (Eds DC Lutes, RE Keane, JF Caratti, CH Key, NC Benson, S Sutherland, LJ Gangi) pp. 1–51. (USDA Forest Service: Ogden, UT)
Koehler GM, Hornocker MG (1977) Fire effects on marten habitat in the Selway-Bitterroot Wilderness. Journal of Wildlife Management 41, 500–505.
| Fire effects on marten habitat in the Selway-Bitterroot Wilderness.Crossref | GoogleScholarGoogle Scholar |
Koehler GM, Maletzke BT, Von Kienast JA, Aubry KB, Wielgus RB, Naney RH (2008) Habitat fragmentation and the persistence of lynx populations in Washington State. Journal of Wildlife Management 72, 1518–1524.
| Habitat fragmentation and the persistence of lynx populations in Washington State.Crossref | GoogleScholarGoogle Scholar |
Lambert JD, Hannon SJ (2000) Short-term effects of timber harvest on abundance, territory characteristics, and pairing success of ovenbirds in riparian buffer strips. The Auk 117, 687–698.
| Short-term effects of timber harvest on abundance, territory characteristics, and pairing success of ovenbirds in riparian buffer strips.Crossref | GoogleScholarGoogle Scholar |
Latour PB, Maclean N, Poole KG (1994) Movements of martens, Martes americana, in burned and unburned taiga in the Mackenzie Valley, Northwest Territories. Canadian Field-Naturalist 108, 351–354.
Lewis CW, Hodges KE, Koehler GM, Mills LS (2011) Influence of stand and landscape features on snowshoe hare abundance in fragmented forests. Journal of Mammalogy 92, 561–567.
| Influence of stand and landscape features on snowshoe hare abundance in fragmented forests.Crossref | GoogleScholarGoogle Scholar |
Lindenmayer DB, Burton PJ, Franklin JF (2008) ‘Salvage logging and its ecological consequences.’ (Island Press: Washington, DC)
Lindenmayer DB, Laurance WF, Franklin JF, Likens GE, Banks SC, Blanchard W, Gibbons P, Ikin K, Blair D, McBurney L, Manning AD, Stein JAR (2014) New policies for old trees: averting a global crisis in a keystone ecological structure. Conservation Letters 7, 61–69.
| New policies for old trees: averting a global crisis in a keystone ecological structure.Crossref | GoogleScholarGoogle Scholar |
Magoun AJ, Vernam DJ (1986) An evaluation of the Bear Creek burn as marten (Martes americana) habitat in interior Alaska. US Bureau of Land Management and Alaska Department of Fish and Game, Final Report, Special Cooperative Project AK-950-CAH-0. (Fairbanks, AK).
McKenzie D, Gedalof Z, Peterson DL, Mote P (2004) Climate change, wildfire, and conservation. Conservation Biology 18, 890–902.
| Climate change, wildfire, and conservation.Crossref | GoogleScholarGoogle Scholar |
Meehan TD, George TL (2003) Short-term effects of moderate- to high-severity wildfire on a disturbance-dependent flycatcher in northwest California. The Auk 120, 1102–1113.
| Short-term effects of moderate- to high-severity wildfire on a disturbance-dependent flycatcher in northwest California.Crossref | GoogleScholarGoogle Scholar |
Millar CI, Stephenson NL, Stephens SL (2007) Climate change and forests of the future: managing in the face of uncertainty. Ecological Applications 17, 2145–2151.
| Climate change and forests of the future: managing in the face of uncertainty.Crossref | GoogleScholarGoogle Scholar | 18213958PubMed |
Miller JD, Safford HD, Crimmins M, Thode AE (2009) Quantitative evidence for increasing fire forest fire severity in the Sierra Nevada and southern Cascade Mountains, California and Nevada, USA. Ecosystems 12, 16–32.
| Quantitative evidence for increasing fire forest fire severity in the Sierra Nevada and southern Cascade Mountains, California and Nevada, USA.Crossref | GoogleScholarGoogle Scholar |
Moriarty KM, Epps CW, Zielinski WJ (2016) Forest thinning changes movement patterns and habitat use by Pacific marten. Journal of Wildlife Management 80, 621–633.
| Forest thinning changes movement patterns and habitat use by Pacific marten.Crossref | GoogleScholarGoogle Scholar |
Murphy EC, Lenhausen WA (1998) Density and foraging ecology of woodpeckers following a stand-replacement fire. Journal of Wildlife Management 62, 1359–1372.
| Density and foraging ecology of woodpeckers following a stand-replacement fire.Crossref | GoogleScholarGoogle Scholar |
Nams VO, Bourgeois M (2004) Fractal analysis measures habitat use at different spatial scales: an example with American marten. Canadian Journal of Zoology 82, 1738–1747.
| Fractal analysis measures habitat use at different spatial scales: an example with American marten.Crossref | GoogleScholarGoogle Scholar |
Nappi A, Drapeau P (2009) Reproductive success of the black-backed woodpecker (Picoides arcticus) in burned boreal forests: are burns source habitats? Biological Conservation 142, 1381–1391.
| Reproductive success of the black-backed woodpecker (Picoides arcticus) in burned boreal forests: are burns source habitats?Crossref | GoogleScholarGoogle Scholar |
Nappi A, Drapeau P (2011) Pre-fire forest conditions and fire severity as determinants of the quality of burned forests for deadwood-dependent species: the case of the black-backed woodpecker. Canadian Journal of Forest Research 41, 994–1003.
| Pre-fire forest conditions and fire severity as determinants of the quality of burned forests for deadwood-dependent species: the case of the black-backed woodpecker.Crossref | GoogleScholarGoogle Scholar |
Nappi A, Drapeau P, Savard JPL (2004) Salvage logging after wildfire in the boreal forest: is it becoming a hot issue for wildlife? Forestry Chronicle 80, 67–74.
| Salvage logging after wildfire in the boreal forest: is it becoming a hot issue for wildlife?Crossref | GoogleScholarGoogle Scholar |
Nappi A, Drapeau P, Saint-Germain M, Angers VA (2010) Effect of fire severity on long-term occupancy of burned boreal conifer forests by saproxylic insects and wood-foraging birds. International Journal of Wildland Fire 19, 500–511.
| Effect of fire severity on long-term occupancy of burned boreal conifer forests by saproxylic insects and wood-foraging birds.Crossref | GoogleScholarGoogle Scholar |
National Interagency Fire Center (2020) Statistics. Available at https://www.nifc.gov/fire-information/statistics [Verified 20 November 2021]
Nelson DL, Kellner KF, Swihart RK (2019) Rodent population density and survival respond to disturbance induced by timber harvest. Journal of Mammalogy 100, 1253–1262.
| Rodent population density and survival respond to disturbance induced by timber harvest.Crossref | GoogleScholarGoogle Scholar |
Paragi TF, Johnson WN, Katnik DD, Magoun AJ (1996) Marten selection of postfire seres in the Alaskan taiga. Canadian Journal of Zoology 74, 2226–2237.
| Marten selection of postfire seres in the Alaskan taiga.Crossref | GoogleScholarGoogle Scholar |
Parks SA, Dillon GK, Miller C (2014) A new metric for quantifying burn severity: the Relativized Burn Ratio. Remote Sensing 6, 1827–1844.
| A new metric for quantifying burn severity: the Relativized Burn Ratio.Crossref | GoogleScholarGoogle Scholar |
Payer DC, Harrison DJ (2003) Influence of forest structure on habitat use by American marten in an industrial forest. Forest Ecology and Management 179, 145–156.
| Influence of forest structure on habitat use by American marten in an industrial forest.Crossref | GoogleScholarGoogle Scholar |
Pilliod DS, Bury RB, Hyde EJ, Pearl CA, Corn PS (2003) Fire and amphibians in North America. Forest Ecology and Management 178, 163–181.
| Fire and amphibians in North America.Crossref | GoogleScholarGoogle Scholar |
Popescu VD, Hunter ML (2011) Clear‐cutting affects habitat connectivity for a forest amphibian by decreasing permeability to juvenile movements. Ecological Applications 21, 1283–1295.
| Clear‐cutting affects habitat connectivity for a forest amphibian by decreasing permeability to juvenile movements.Crossref | GoogleScholarGoogle Scholar | 21774430PubMed |
Porter AD, St Clair CC, de Vries A (2005) Fine-scale selection by marten during winter in a young deciduous forest. Canadian Journal of Forest Research 35, 901–909.
| Fine-scale selection by marten during winter in a young deciduous forest.Crossref | GoogleScholarGoogle Scholar |
Potvin F, Bélanger L, Lowell K (2000) Marten habitat selection in a clearcut boreal landscape. Conservation Biology 14, 844–857.
| Marten habitat selection in a clearcut boreal landscape.Crossref | GoogleScholarGoogle Scholar |
Powell RA (1979) Ecological energetics and foraging strategies of the fisher (Martes pennanti). Journal of Animal Ecology 48, 195–212.
| Ecological energetics and foraging strategies of the fisher (Martes pennanti).Crossref | GoogleScholarGoogle Scholar |
Powell R (2004) Home ranges, cognitive maps, habitat model and fitness landscapes for Martes. In ‘Martens and fishers (Martes) in human-altered environments: an international perspective’. (Eds DJ Harrison, AK Fuller, G Proulx) pp. 135–146. (Springer: New York, NY)
Proulx G, Kariz R, Farkvam J (2006) Using forest inventory data to predict the distribution of potential winter habitats for American martens. In ‘Martes in carnivore communities’. (Eds M Santos-Reis, JDS Birks, EC O’Doherty, G Proulx) pp. 77–88. (Alpha Wildlife Publications: Sherwood Park, AB)
Pulliainen E (1982) Scent-marking in the pine marten (Martes martes) in Finnish forest Lapland in winter. Zeitschrift für Säugetierkunde 47, 91–99.
Raine MR (1987) Winter food habits and foraging behaviour of fishers (Martes pennanti) and martens (Martes americana) in southeastern Manitoba. Canadian Journal of Zoology 65, 745–747.
| Winter food habits and foraging behaviour of fishers (Martes pennanti) and martens (Martes americana) in southeastern Manitoba.Crossref | GoogleScholarGoogle Scholar |
Resources Information Standards Committee (RISC) (2007) Vegetation resources inventory ground sampling procedures. BC Ministry of Forests and Range, Forest Analysis and Inventory Branch for the Terrestrial Ecosystems Task Force Resources Information Standards Committee, Version 4.7. Victoria, BC.
Robinson NM, Leonard SWJ, Bennett AF, Clarke MF (2014) Refuges for birds in fire-prone landscapes: the influence of fire severity and fire history on the distribution of forest birds. Forest Ecology and Management 318, 110–121.
| Refuges for birds in fire-prone landscapes: the influence of fire severity and fire history on the distribution of forest birds.Crossref | GoogleScholarGoogle Scholar |
Ruggiero LF, Pearson DE, Henry SE (1998) Characteristics of American marten den sites in Wyoming. Journal of Wildlife Management 62, 663–673.
| Characteristics of American marten den sites in Wyoming.Crossref | GoogleScholarGoogle Scholar |
Russell RE, Saab VA, Dudley JG, Rotella JJ (2006) Snag longevity in relation to wildfire and postfire salvage logging. Forest Ecology and Management 232, 179–187.
| Snag longevity in relation to wildfire and postfire salvage logging.Crossref | GoogleScholarGoogle Scholar |
Saab VA, Powell HDW (2005) Fire and avian ecology in North America: process influencing pattern. Studies in Avian Biology 30, 1–13.
Saab VA, Russell RE, Dudley JG (2009) Nest-site selection by cavity-nesting birds in relation to postfire salvage logging. Forest Ecology and Management 257, 151–159.
| Nest-site selection by cavity-nesting birds in relation to postfire salvage logging.Crossref | GoogleScholarGoogle Scholar |
Schieck J, Song SJ (2006) Changes in bird communities throughout succession following fire and harvest in boreal forests of western North America: literature review and meta-analyses. Canadian Journal of Forest Research 36, 1299–1318.
| Changes in bird communities throughout succession following fire and harvest in boreal forests of western North America: literature review and meta-analyses.Crossref | GoogleScholarGoogle Scholar |
Schober P, Boer C, Schwarte LA (2018) Correlation coefficients: appropriate use and interpretation. Anesthesia and Analgesia 126, 1763–1768.
| Correlation coefficients: appropriate use and interpretation.Crossref | GoogleScholarGoogle Scholar | 29481436PubMed |
Seip CR, Hodder DP, Crowley SM, Johnson CJ (2018) Use of constructed coarse woody debris corridors in a clearcut by American martens (Martes americana) and their prey. Forestry: An International Journal of Forest Research 91, 506–513.
| Use of constructed coarse woody debris corridors in a clearcut by American martens (Martes americana) and their prey.Crossref | GoogleScholarGoogle Scholar |
Simanonok MP, Burkle LA (2019) Nesting success of wood-cavity-nesting bees declines with increasing time since wildfire. Ecology and Evolution 9, 12436–12445.
| Nesting success of wood-cavity-nesting bees declines with increasing time since wildfire.Crossref | GoogleScholarGoogle Scholar | 31788188PubMed |
Slauson KM, Zielinski WJ, Schwartz MK (2017) Ski areas affect Pacific marten movement, habitat use, and density. Journal of Wildlife Management 81, 892–904.
| Ski areas affect Pacific marten movement, habitat use, and density.Crossref | GoogleScholarGoogle Scholar |
Soutiere EC (1979) Effects of timber harvesting on marten in Maine. Journal of Wildlife Management 43, 850–860.
| Effects of timber harvesting on marten in Maine.Crossref | GoogleScholarGoogle Scholar |
Spencer WD, Barrett RH, Zielinski WJ (1983) Marten habitat preferences in the northern Sierra Nevada. Journal of Wildlife Management 47, 1181–1186.
| Marten habitat preferences in the northern Sierra Nevada.Crossref | GoogleScholarGoogle Scholar |
Stephenson RO (1984) The relationship of fire history to furbearer populations and harvest. Alaska Department of Fish and Game, Final Report, Project W-22-2, Job 7.13 R. (Juneau, AK).
Steventon JD, Daust DK (2009) Management strategies for a large-scale mountain pine beetle outbreak: modelling impacts on American martens. Forest Ecology and Management 257, 1976–1985.
| Management strategies for a large-scale mountain pine beetle outbreak: modelling impacts on American martens.Crossref | GoogleScholarGoogle Scholar |
Sullivan TP, Sullivan DS (2021) Responses of mustelids and small mammals prey to combined retention on clearcuts: woody debris, green trees, and riparian structures. Forest Ecology and Management 496, 119431
| Responses of mustelids and small mammals prey to combined retention on clearcuts: woody debris, green trees, and riparian structures.Crossref | GoogleScholarGoogle Scholar |
Sullivan TP, Sullivan DS, Lindgren PMF (2000) Small mammals and stand structure in young pine, seed-tree, and old-growth forest, southwest Canada. Ecological Applications 10, 1367–1383.
| Small mammals and stand structure in young pine, seed-tree, and old-growth forest, southwest Canada.Crossref | GoogleScholarGoogle Scholar |
Sullivan TP, Sullivan DS, Lindgren PMF, Ransome DB, Bull JG, Ristea C (2011) Bioenergy or biodiversity? Woody debris structures and maintenance of red-backed voles on clearcuts. Biomass and Bioenergy 35, 4390–4398.
| Bioenergy or biodiversity? Woody debris structures and maintenance of red-backed voles on clearcuts.Crossref | GoogleScholarGoogle Scholar |
Sullivan TP, Sullivan DS, Klenner W (2021) Fate of postharvest woody debris, mammal habitat, and alternative management of forest residues on clearcuts: a synthesis. Forests 12, 551
| Fate of postharvest woody debris, mammal habitat, and alternative management of forest residues on clearcuts: a synthesis.Crossref | GoogleScholarGoogle Scholar |
Swan M, Christie F, Sitters H, York A, Di Stefano J (2015) Predicting faunal fire responses in heterogeneous landscapes: the role of habitat structure. Ecological Applications 25, 2293–2305.
| Predicting faunal fire responses in heterogeneous landscapes: the role of habitat structure.Crossref | GoogleScholarGoogle Scholar | 26910956PubMed |
Taylor SL, Buskirk SW (1994) Forest microenvironments and resting energetics of the American marten Martes americana. Ecography 17, 249–256.
| Forest microenvironments and resting energetics of the American marten Martes americana.Crossref | GoogleScholarGoogle Scholar |
Thomas JP, Reid ML, Barclay RMR, Jung TS (2019) Salvage logging after an insect outbreak reduces occupancy by snowshoe hares (Lepus americanus) and their primary predators. Global Ecology and Conservation 17, e00562
| Salvage logging after an insect outbreak reduces occupancy by snowshoe hares (Lepus americanus) and their primary predators.Crossref | GoogleScholarGoogle Scholar |
Thompson ID, Fryxell J, Harrison DJ (2012) Improved insights into use of habitat by American martens. In ‘Biology and conservation of martens, sables, and fishers: a new synthesis’. (Eds KB Aubry, WJ Zielinski, MG Raphael, G Proulx, SW Buskirk) pp. 209–230. (Cornell University Press: Ithaca, NY)
Thorn S, Bässler C, Brandl R, Burton PJ, Cahall R, Campbell JL, Castro J, Choi C-Y, Cobb T, Donato DC, Durska E, Fontaine JB, Gauthier S, Hebert C, Hothorn T, Hutto RL, Lee E-J, Leverkus AB, Lindenmayer DB, Obrist MK, Rost J, Seibold S, Seidl R, Thom D, Waldron K, Wermelinger B, Winter M-B, Zmihorski M, Müller J (2018) Impacts of salvage logging on biodiversity: a meta-analysis. Journal of Applied Ecology 55, 279–289.
| Impacts of salvage logging on biodiversity: a meta-analysis.Crossref | GoogleScholarGoogle Scholar |
Thorn S, Chao A, Georgiev KB, et al. (2020) Estimating retention benchmarks for salvage logging to protect biodiversity. Nature Communications 11, 4762
| Estimating retention benchmarks for salvage logging to protect biodiversity.Crossref | GoogleScholarGoogle Scholar | 32958767PubMed |
van Buuren S, Groothuis-Oudshoorn K (2011) mice: Multivariate Imputation by Chained Equations in R. Journal of Statistical Software 45, 1–67.
| mice: Multivariate Imputation by Chained Equations in R.Crossref | GoogleScholarGoogle Scholar |
Vanbianchi CM, Murphy MA, Hodges KE (2017) Canada lynx use of burned areas: conservation implications of changing fire regimes. Ecology and Evolution 7, 2382–2394.
| Canada lynx use of burned areas: conservation implications of changing fire regimes.Crossref | GoogleScholarGoogle Scholar | 28405301PubMed |
Vanderwel MC, Malcolm JR, Caspersen JP, Newman MA (2010) Fine‐scale habitat associations of red‐backed voles in boreal mixedwood stands. Journal of Wildlife Management 74, 1492–1501.
| Fine‐scale habitat associations of red‐backed voles in boreal mixedwood stands.Crossref | GoogleScholarGoogle Scholar |
Venables WN, Ripley BD (Eds) (2002) ‘Modern applied statistics with S.’, 4th edn. (Springer: New York, NY)
Vigeant-Langlois C, Desrochers A (2011) Movements of wintering American martens (Martes americana): relative influences of prey activity and forest stand age. Canadian Journal of Forest Research 41, 2202–2208.
| Movements of wintering American martens (Martes americana): relative influences of prey activity and forest stand age.Crossref | GoogleScholarGoogle Scholar |
Volkmann LA (2021) Habitat selection by Pacific marten (Martes caurina) and other carnivores after wildfire and post-fire salvage logging. PhD dissertation, University of British Columbia Okanagan, Kelowna, BC.
Volkmann LA, Hodges KE (2021) Post-fire movements of Pacific marten (Martes caurina) depend on the severity of landscape change. Movement Ecology 9, 49
| Post-fire movements of Pacific marten (Martes caurina) depend on the severity of landscape change.Crossref | GoogleScholarGoogle Scholar | 34627394PubMed |
Volkmann LA, Hutchen J, Hodges KE (2020) Trends in carnivore and ungulate fire ecology research in North American conifer forests. Forest Ecology and Management 458, 117691
| Trends in carnivore and ungulate fire ecology research in North American conifer forests.Crossref | GoogleScholarGoogle Scholar |
Watt WR, Baker JA, Hogg DM, McNicol JG, Naylor BJ (1996) ‘Forest management guidelines for the provision of marten habitat. Version 1.0.’ (Ontario Ministry of Natural Resources: Sault Ste. Marie, ON)
Weir RD, Phinney M, Lofroth EC (2012) Big, sick, and rotting: why tree size, damage, and decay are important to fisher reproductive habitat. Forest Ecology and Management 265, 230–240.
| Big, sick, and rotting: why tree size, damage, and decay are important to fisher reproductive habitat.Crossref | GoogleScholarGoogle Scholar |
Westerling AL, Hidalgo HG, Cayan DR, Swetnam TW (2006) Warming and earlier spring increase western US forest wildfire activity. Science 313, 940–943.
| Warming and earlier spring increase western US forest wildfire activity.Crossref | GoogleScholarGoogle Scholar | 16825536PubMed |
Westerling AL, Turner MG, Smithwick EAH, Romme WH, Ryan MG (2011) Continued warming could transform Greater Yellowstone fire regimes by mid-21st century. Proceedings of the National Academy of Sciences 108, 13165–13170.
| Continued warming could transform Greater Yellowstone fire regimes by mid-21st century.Crossref | GoogleScholarGoogle Scholar |
Wilbert CJ, Buskirk SW, Gerow KG (2000) Effects of weather and snow on habitat selection by American martens (Martes americana). Canadian Journal of Zoology 78, 1691–1696.
| Effects of weather and snow on habitat selection by American martens (Martes americana).Crossref | GoogleScholarGoogle Scholar |
Wilk RJ, Raphael MG (2018) Survival and predators of Pacific marten in a salvage-logged pine forest, south-central Oregon. Northwestern Naturalist 99, 115–123.
| Survival and predators of Pacific marten in a salvage-logged pine forest, south-central Oregon.Crossref | GoogleScholarGoogle Scholar |
Wolfinger R, O’Connell M (1993) Generalized linear mixed models: a pseudo-likelihood approach. Journal of Statistical Computation and Simulation 48, 233–243.
| Generalized linear mixed models: a pseudo-likelihood approach.Crossref | GoogleScholarGoogle Scholar |
Wood DJA, Drake S, Rushton SP, Rautenkranz D, Lurz PWW, Koprowski JL (2007) Fine-scale analysis of Mount Graham red squirrel habitat following disturbance. Journal of Wildlife Management 71, 2357–2364.
| Fine-scale analysis of Mount Graham red squirrel habitat following disturbance.Crossref | GoogleScholarGoogle Scholar |
Woodall C, Williams MS (2007) Sampling protocol, estimation, and analysis procedures for the Down Woody Materials Indicator of the FIA Program.’ USDA Forest Service, North Central Research Station, General Technical Report NC-256. (St. Paul, MN)
Wynne KM, Sherburne JA (1984) Summer home range use by adult marten in northwestern Maine. Canadian Journal of Zoology 62, 941–943.
| Summer home range use by adult marten in northwestern Maine.Crossref | GoogleScholarGoogle Scholar |