

Conifer encroachment increases foliar moisture content in a northwestern California oak woodland
Jeffrey M. Kane A * , Lucy P. Kerhoulas A and Gabriel S. Goff AA Department of Forestry, Fire, and Rangeland Management, California State Polytechnic University, Humboldt, One Harpst Street, Arcata, CA 95521, USA.
International Journal of Wildland Fire 32(5) 728-737 https://doi.org/10.1071/WF22184
Submitted: 16 August 2022 Accepted: 2 February 2023 Published: 2 March 2023
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing on behalf of IAWF. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Background: Foliar moisture content influences crown fire ignition and behaviour. Some spatial variation in foliar moisture is attributable to differences in stand conditions but evidence describing this role is lacking or contradicting.
Aims: To examine the role of stand conditions on tree physiology and foliar moisture content in Oregon white oak (Quercus garryana) and Douglas-fir (Pseudotsuga menziesii).
Methods: We monitored foliar moisture content in both species, and tree physiology (stomatal conductance and leaf water potential) in oak, across three stand conditions, including intact stands (unencroached), stands invaded by Douglas-fir (encroached), and thinned stands with Douglas-fir removed.
Key results: Encroached stands had higher foliar moisture content than intact or thinned stands. Higher stand density was associated with higher foliar moisture content in both species and foliage ages. Encroached stands also had higher midday leaf water potential compared with intact or thinned stands.
Conclusions: These findings provide strong evidence that stand conditions and thinning treatments can influence foliar moisture content in Oregon white oak ecosystems, with likely implications for other ecosystems.
Implications: Better understanding of the role of stand conditions on foliar moisture content may contribute to improved spatial and temporal prediction of foliar moisture content and modelling of potential crown fire behaviour.
Keywords: fuel moisture, leaf water potential, live fuel moisture, Pseudotsuga menziesii, Quercus garryana, tree physiology, wildfire, wildland fuels.
Introduction
Foliar moisture content (i.e. live fuel moisture in trees and shrubs) is a contributing factor in the ignition and behaviour of crown fires in shrublands and forests (Xanthopoulos and Wakimoto 1993; Dimitrakopoulos and Papaioannou 2001; Weise et al. 2005; Pellizzaro et al. 2007; Kuljian and Varner 2010). Higher foliar moisture content dampens ignition and spread due to the high specific heat of water and the energetic need to evaporate water prior to combustion (Simms and Law 1967). Foliar moisture content is included in all crown fire models and fire danger ratings used by managers (Van Wagner 1977; Bradshaw et al. 1983; Scott and Reinhardt 2001). Although some have questioned the predictive ability of foliar moisture content under field conditions (Alexander and Cruz 2013), others suggest that observed discrepancies between lab and field studies likely reflect an oversimplified consideration of foliar moisture content (Rossa and Fernandes 2018). Still, studies have clearly identified critical thresholds in foliar moisture content that are highly informative in predicting area burned (Dennison et al. 2008; Nolan et al. 2016), but see Pimont et al. (2019) for a critique of this approach. Future projections indicate warming associated with climate change may contribute to reduced foliar moisture content and longer periods below critical threshold levels (Ma et al. 2021). These circumstances suggest that consideration of foliar moisture content can be a useful part of the fuels and fire management process (Keyes 2006).
Foliar moisture content within and among species is known to substantially vary temporally and spatially. Temporal fluctuation in foliar moisture content occurs at diel, seasonal, and interannual scales (Jameson 1966; Van Wagner 1967; Agee et al. 2002; Keyes 2006). Spatial variation in foliar moisture content reflects differences in species composition, environmental gradients, and their interactions (Yebra et al. 2008; Krix and Murray 2018; Nolan et al. 2018). The mechanistic drivers contributing to this variation are still being refined, but likely reflect the interactive influence of climate, edaphic conditions, plant traits, and vegetation structure. Early research by fire scientists primarily treated live fuel moisture as dead fuels with typically higher fuel moisture content, but plant physiologists have long known that water and carbon flows and storage are dynamic (Kozlowski and Clausen 1965), with important implications to plant flammability and fire behaviour (Jolly et al. 2014; Krix and Murray 2018). Advances in understanding and predicting foliar moisture content will inevitably require integration of knowledge and approaches across ecophysiology and fire science disciplines, especially under warming and drying conditions (Jolly and Johnson 2018; Nolan et al. 2020).
Previous research has identified that variations in foliar moisture content among species are attributable to trait differences that correspond to water and carbon acquisition and regulation strategies. Deeper-rooted species with more reliable access to water resources can be decoupled from site conditions and thus maintain higher foliar moisture content (Brown et al. 2022). Plant–water relations (i.e. leaf water potential, transpiration) can also influence foliar moisture content both within and among species (Nolan et al. 2018; Pivovaroff et al. 2019). Differences in foliar moisture content between species have been associated with leaf traits, such as leaf mass area (Krix and Murray 2018).
Foliar moisture content within a species can vary due to tree- and stand-level factors (Ruffault et al. 2022). Tree-level factors such as leaf morphology and tree size (i.e. diameter and height), as well as their corresponding effects on leaf area and rooting depth, likely contribute to variation in foliar moisture content. Tree-level influences on foliar moisture content are also modified by stand-level factors, including stand structure and composition, and their effects on microclimate and soil moisture availability. However, the limited available evidence on the role of stand-level factors on foliar moisture content is seemingly contradictory or dependent on local context (Soler Martin et al. 2017; Krix and Murray 2018).
Based on previous research findings and ecophysiological reasoning, there are three conceivable outcomes in the relationship between stand density (e.g. stand basal area) and foliar moisture content. Some evidence suggests that stand density, and more directly stand-level leaf area index, would be negatively associated with foliar moisture content. Prior studies have shown that higher density stands were associated with greater water stress as measured by leaf water potential (McDowell et al. 2006; Moreno and Cubera 2008), a circumstance that may explain observed decreases in foliar moisture content with higher stand density (McNamara et al. 2019). Conversely, reducing competition through thinning treatments can increase soil moisture availability and throughfall precipitation, contributing to increased diameter growth (Devine and Harrington 2006, 2013; Gould et al. 2011; Sohn et al. 2016), with presumable increases in foliar moisture content.
Stand density could also be unrelated to foliar moisture content in some species due in part to plant water regulation strategies. Many species regulate water storage and loss through numerous mechanisms, such as stomatal control, osmotic potential, and modulus elasticity, among others (Pallardy 2008). Thus, changes in stand density may not result in a strong influence on foliar moisture content due to internal physiological regulation. Observations of relatively static foliar moisture content over time have been observed in some species (Viegas et al. 2001; Pellizzaro et al. 2007), and in one study, foliar moisture content did not differ between thinned and unthinned stands (Soler Martin et al. 2017).
A third possible relationship is that foliar moisture content could be positively associated with stand density. In more open or recently thinned stands, higher wind speeds and greater penetration of solar radiation can lead to increased transpiration and evaporative water loss compared with higher density stands (Lagergren et al. 2008; Brooks and Mitchell 2011). Furthermore, the larger leaf area of more open-growing trees, and greater soil exposure in more open (lower leaf area index) stands, can increase stand-level transpiration and evaporation that may negatively affect soil water availability (Ruffault et al. 2022) and reduce foliar moisture content. Greater overstorey cover associated with higher density stands in more humid environments can reduce surface temperature and increase relative humidity under the canopy (Kane 2021), leading to increased leaf water potential compared with lower density stands due to greater soil moisture availability and higher predawn water potential (Goff 2021). These findings are consistent with observed differences in foliar moisture content between sheltered (higher density stands) and exposed (lower density stands) habitats in a fire-prone landscape of southeastern Australia (Krix and Murray 2018). There is also the possibility that positive, negative, and neutral relationships between plant water stress and foliar moisture content can occur within a given stand, depending on interactions between climate and stand conditions. Differences in the directional effect of plant water stress and foliar moisture content may reflect shifts along a curvilinear relationship (Tyree and Hammel 1972).
Oregon white oak ecosystems provide a framework to examine the role of stand density on tree physiology and seasonal patterns of foliar moisture content. Many Oregon white oak woodlands have substantial encroachment from Douglas-fir (Pseudotsuga menziesii) and other conifers from adjacent forests due to the prolonged absence of fire (Agee 1993; Tveten and Fonda 1999). Oregon white oak woodlands and associated prairies have declined by as much as 30% from their historical extent in some areas due to encroachment (Fritschle 2008; Knight et al. 2020). Low-intensity prescribed fire is a commonly used method to maintain and improve conditions (Underwood et al. 2003; Hamman et al. 2011). However, prescribed fire can be insufficient in killing invading Douglas-fir (Engber and Varner 2012), partly due to reduced herbaceous fuel loads and higher dead surface fuel moisture that lowers fire intensity in encroached stands (Engber et al. 2011). In these instances, managers often opt to remove Douglas-fir by thinning or girdling treatments to improve oak vigour and restore oak woodlands (Devine and Harrington 2013; Kane et al. 2019). Removal of Douglas-fir can better facilitate prescribed fire by increasing temperature and vapour pressure deficit (Devine and Harrington 2007; Kane 2021) that aids in reducing dead surface fuel moisture content compared with encroached stands (Engber et al. 2011; Kane 2021). However, research on the impacts of conifer encroachment and removal on foliar moisture content have not been examined.
The aim of this case study was to characterise the seasonal patterns of foliar moisture content of Oregon white oak and Douglas-fir under varying stand conditions in northwestern California. Specifically, the objectives of the research were to: (1) determine the influence of Douglas-fir encroachment and removal on seasonal changes in foliar moisture content; and (2) compare the effect of stand density on tree physiology (leaf water potential and stomatal conductance) and foliar moisture content across stand conditions. Based on field observations and prior related research at the study site, our hypothesis for this study was that encroached stands would have higher foliar moisture content in Oregon white oak and Douglas-fir, and that this trend would be driven by denser stands having higher leaf water potential and lower stomatal conductance. Results of this study will provide much-needed information about the influence of stand density and tree physiology on foliar moisture content that can better assist in predicting foliar moisture content and modelling of potential crown fire behaviour.
Methods
Study site and experimental design
Our study was conducted in Kneeland, California, USA at the Mountain View site (40.71°N 123.90°W) on the Yager Creek tract of Humboldt Redwood Company. The climate at the study site is characterised as mediterranean, with a maritime influence that experiences coastal summer fog. Between June and November, temperature ranges between 10.1 and 25.4°C, and precipitation ranges between 169 and 664 mm, based on the 30-year mean climate (1989–2018) for the study site (PRISM Climate Group 2020). Based on the Kneeland remote automated weather station data that is located about 2 km from the study site (40.72°N 123.93°W), mean monthly temperatures ranged from 11.2 to 21.9°C (mean 16.6°C), minimum relative humidity ranged from 7 to 86% (mean 42%), and precipitation totalled 252 mm (64% of the 30-year mean) from June to November 2018.
The bedrock of the study site is of sedimentary and meta-sedimentary origin and is part of the Franciscan Complex in the Coast Range Province. Shallow, fine loamy alfisol soils in the Elkcamp series and fine mixed mollisol soils in the Kinman series characterise the site. Prior observations at the study site did not detect differences in soil texture and water holding capacity between encroached and intact stands (S. Marshall, unpubl. data). The elevation of the site ranged between 680 and 820 m, with moderately steep southern (from southeast to southwest) slopes ranging from 19 to 35°. Fire has not been recorded at the site since 1900 (CALFIRE 2020), although the site was prescribed burned the year following data collection of the current study in 2019. Prior to fire exclusion and forced removal of Indigenous communities from the region, fire return interval within the study area was estimated to be between 6 and 10 years (LANDFIRE 2020).
Intact Oregon white oak woodlands with limited Douglas-fir encroachment are generally more common along the mid- to upper-slope portions of the site. Oak woodlands with encroachment are generally more prevalent in lower slope positions that are adjacent to Douglas-fir forests. Oregon white oak and Douglas-fir were the dominant two species, but California black oak (Q. kelloggii) and Pacific madrone (Arbutus menziesii) were also present. We intentionally minimised the potential influences of topographic differences among stand conditions by selecting sites with overlapping elevation, aspect, and slope position (Table 1). Based on a previous study at the same site, tree age did not differ among stand conditions (Schriver et al. 2018). Most Oregon white oak and California black oak occurred between 1850 and 1900, while almost all Douglas-fir established after 1950. Thinning and harvest treatments were implemented at the site in 2017 using a combination of variable-retention harvesting and group-selection harvesting approaches. Treatments primarily focused on the removal of Douglas-fir trees >25 cm diameter at breast height (DBH; 1.37 m). Retention of some Douglas-fir that were growing intertwined within the crowns of large oak trees was necessary to limit damage to oaks and potential injury to operators.

|
Our experimental design for the study included sampling a total of nine plots – three plots for each stand condition (intact, high encroached, and thinned). The intact woodland and high encroached plots were randomly selected as part of a previous study (Schriver et al. 2018). Intact stands represent relatively open oak woodland stands that had limited or no encroachment by Douglas-fir. Yet, these stands may have slightly increased in oak density over the years due to reduced fire frequency (Fritschle 2008; Schriver et al. 2018). High encroached stands contained moderate-to-large Douglas-fir that partially overtopped the oaks. We intentionally selected thinned plots that were located close to heavily encroached stands and had clear evidence of Douglas-fir removal. All plots contained at least three oaks greater than 15 cm DBH within a 10 m radius of plot centre. Within each plot, we identified each species and measured the DBH of all trees within 10 m (area = 78.5 m2) of plot centre. From these data, we calculated stand basal area and tree density for each plot and stand condition. As intended, stand structure varied by stand condition (F2,7 = 5.2, P = 0.048; Table 1). Encroached stands had higher tree density and mean basal area compared with intact and thinned stands. Variation in stand basal area in thinned stands was higher than intact or encroached stands because one plot was not as heavily thinned due to the inability to remove some Douglas-fir that had crowns intertwining with oaks.
Data collection
Foliar moisture samples were haphazardly collected monthly from three trees of Oregon white oak and Douglas-fir in the co-dominant or dominant canopy position within the 10 m radius of each plot to account for spatial variability at the plot level. DBH for all trees sampled for foliar moisture content ranged between 19.0 and 36.2 cm, and did not differ among stand conditions (P > 0.246). However, the crown base heights of Oregon white oaks were substantially lower in intact stands (~2–6 m) than in encroached and thinned stands (~6–12 m). Sampling of foliage was conducted every month from June until November in 2018, using pole pruners to cut one or two branches from the lower one-third of the crown. We separately collected new foliage (formed in 2018) of both Douglas-fir and Oregon white oak and old foliage (≥2 years old) in Douglas-fir only. Because Oregon white oak is deciduous, only new foliage was collected. All sampling occurred on sunny or lightly overcast days between 1100 and 1500 hours. The collection time of each sample was recorded on the sample bag. Each samples had an average live leaf dry mass of approximately 10 g (s.e. = 0.28 g). Collected samples were separately placed in a prelabelled and preweighed sealable plastic bag. Bags were then placed in a cooler, and were immediately weighed with the plastic bag to 0.01 g on an analytical balance upon arrival at the Cal Poly Humboldt Fire Lab in Arcata, California, USA. Foliage samples were removed from the plastic bag, placed into separate paper bags, and oven-dried for at least 3 days at 60°C. Higher drying temperatures have been suggested (Matthews 2010), but we intentionally selected a lower drying temperature to limit volatile mass loss with the trade-off of retaining some (~1–3%) cellular bound moisture. Each sample was removed from the paper bag and weighed after drying. We calculated percentage of foliar moisture content for each sample by subtracting the wet weight from the dry weight, dividing by the dry weight, and multiplying by 100.
On 25 August 2018, we measured leaf water potential and stomatal conductance of ten Oregon white oak trees within each of five stand conditions: (1) intact (no Douglas-fir encroachment); (2) moderate encroached without thinning; (3) moderate encroached with thinning; (4) high encroached without thinning; and (5) high encroached with thinning. Sampling on this date was chosen because we knew soil moisture would be around its lowest and wildfires most commonly expected. Moderate encroached plots were still dominated by Oregon white oak, but contained some Douglas-fir in the canopy. For each sampled tree, leaf water potential was measured at predawn (0200–0500 hours) and at midday (1030–1400 hours) using a pressure chamber (Model 600, PMS Instruments, Corvallis, OR, USA), and stomatal conductance was measured using a leaf porometer (SC-1, Decagon Devices Inc., Pullman, WA, USA). All samples were collected from the lower crown using a pole pruner, with midday samples taken from fully illuminated portions of the crown when possible. Once a small branch was pruned from the crown, three to five leaf water potential measurements were taken from different sub-branchlets and averaged into one value. Three stomatal conductance measurements were taken at midday from different leaves and averaged into one value. For each midday water potential sample, a subset of the sample was used to calculate foliar moisture content.
Data analysis
We used a linear mixed effect modelling approach using the lmer function in the lme4 package (Bates et al. 2015) in the R statistical environment (version 4.0.2, R Development Core Team 2022). This approach was used to determine if foliar moisture content significantly varied by foliage age (new and old foliage), species (Oregon white oak and Douglas-fir foliage), stand condition (intact, high encroached, and thinned), and time (June through November). Foliar moisture content values were log transformed to meet the normality and equal variance assumptions of residuals in each analysis. The random effect structure of the mixed effects models included a random intercept term for each unique plot. All candidate models included the fixed effect of time. Additional models included foliage age, species, or stand condition, and an additive model with foliage age, species, and stand condition. All candidate models were evaluated for multicollinearity using the vif function in the package car (Fox and Weisberg 2019). Explanatory variables with variance inflation factors >4 were excluded from the top models (Hair et al. 2018). Model selection was determined based on the lowest Akaike information criterion (AIC) value, and in cases where the top models were within 2 AIC, the model with the fewest parameters was selected (Burnham and Anderson 1998). We report the significance of each variable considered using Type III analysis of variance. To examine the effects of stand condition on foliar moisture content, linear regression modelling was used to compare mean foliar moisture content by foliage age, foliage species, total stand basal area, and stand condition of each of the nine plots. A linear regression modelling approach was also used to examine the relationship between Oregon white oak tree physiology (midday and predawn leaf water potential, stomatal conductance). We also used a linear mixed effect modelling approach as described above to examine if foliar moisture content in Oregon white oak was significantly related to leaf water potential (midday and predawn), stomatal conductance, stand condition (intact, moderate encroached, and high encroached), and thinning treatment (thinned or unthinned).
Results
Stand influences on foliar moisture content
An additive model including time, species, foliage age, and stand condition best explained variation in foliar moisture content (Fig. 1). Species explained the most variation in mean foliar moisture content over the duration of the study (F = 251.2, P < 0.0001), with Douglas-fir foliage having 39% more foliar moisture content than Oregon white oak foliage. Foliage age also explained much of the variation in foliar moisture content (F = 237.2, P < 0.0001), with new foliage having 25% more foliar moisture content than old foliage. Foliar moisture content was also strongly associated with time (F = 122.9, P < 0.0001). Generally, foliar moisture content of new leaves was highest in June and decreased through late September. Older Douglas-fir needles tended to increase slightly in foliar moisture content over time. Stand conditions differed in foliar moisture content (F = 61.2, P < 0.0001). Mean foliar moisture content across all foliage ages and species over the duration of the study was about 65 and 35% higher in encroached stands than in intact (P < 0.0001) and thinned stands (P = 0.0012), respectively. Mean oak foliar moisture content was 2.6% higher in intact stands compared with thinned stands, but were not statistically different (P = 0.3945)

|
An additive model of foliage age, species, and stand basal area were related to mean seasonal foliar moisture content (Fig. 2). Species explained the most variation in foliar moisture content (F = 64.1, P < 0.0001). Foliage age explained substantial variation in foliar moisture content (F = 53.1, P < 0.0001), with new foliage having higher foliar moisture content than old foliage. Stand basal area was positively related with foliar moisture content (F = 14.3, P = 0.0015), but the strength of this relationship varied by foliage type, and an interaction between the two terms was not more informative. Although based on univariate analysis, Oregon white oak foliar moisture content had a better relationship with stand basal area (R2 = 0.56, P = 0.02) compared with relationships with either new or old Douglas-fir foliage (R2 < 0.47, P > 0.13).
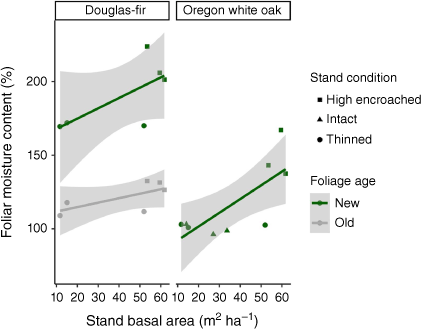
|
Tree physiology influences on foliar moisture content
Leaf water potential was strongly related to Oregon white oak foliar moisture content, but the strength of this relationship varied with stand condition (Fig. 3). The most informative model included midday leaf water potential, stand condition, and an interaction term. Foliar moisture content across all stand conditions was better associated with midday leaf water potential (P < 0.0001) than predawn leaf water potential (P = 0.12). Stand condition (F = 7.5, P = 0.001), and the interaction of stand condition with midday leaf water potential (F = 4.7, P = 0.01), exerted the strongest influence on foliar moisture content. Midday leaf water potential as a factor by itself was not quite significant (F = 3.0, P = 0.08) but was still included in the most informative model. Based on univariate analysis for each stand condition, high encroached stands had the strongest relationship between midday leaf water potential and foliar moisture content (R2 = 0.49, P < 0.0001) compared with moderate encroached or intact stands (R2 < 0.02, P > 0.398). Stomatal conductance and thinning treatment were not considered to be informative in the top model. Mean midday water potential among all stands examined was also strongly positively associated with stand basal area at the plot level (R2 = 0.96, P = 0.0037).

|
Discussion
Our study demonstrates that seasonal foliar moisture dynamics in a northwestern Oregon white oak ecosystem were related to differences in stand condition and its influence on tree physiology. We found that increased stand density associated with Douglas-fir encroachment resulted in higher midday leaf water potential and higher foliar moisture content than in lower density stands (intact or thinned). Given the inclusion of foliar moisture content in crown fire modelling and fire danger rating systems (e.g. Van Wagner 1977; Bradshaw et al. 1983; Scott and Reinhardt 2001), our findings suggest the need to consider the competitive environment (i.e. stand density, leaf area index) when developing predictive models of foliar moisture content, or when assigning foliar moisture content values within crown fire modelling scenarios.
Consistent with our expectations, encroached stands supported higher foliar moisture content in both Douglas-fir and Oregon white oak compared with the more open thinned or intact stands. Our results agree with previous research that found similar differences in foliar moisture content between sheltered and exposed ecosystems in southeastern Australia (Krix and Murray 2018). Our study advances our understanding because we were able to examine differences in stand density within the same species and site conditions, limiting the potential impacts of some confounding factors. Yet, uncertainty remains as to the direct mechanisms that contributed to the observed differences among stand conditions. We suspect that higher density stands (such as the encroached stands observed within our study) had greater canopy cover, which can reduce the amount of solar radiation that reaches the forest floor, resulting in cooler air temperature and higher relative humidity beneath the canopy (Ma et al. 2010; Kane 2021). Additionally, deeper and denser organic soil horizons (i.e. litter and duff) present within encroached stands (Engber et al. 2011) may also impede soil drying. These conditions can reduce evaporative losses of soil moisture and increase leaf water potential, contributing to higher foliar moisture content (Nolan et al. 2018; Balaguer-Romano et al. 2022). Yet, our results run contradictory to observations that increased stand density and greater leaf area index can have negative impacts on plant water status (McDowell et al. 2006). This inconsistency may highlight that our findings are limited to woodlands and forests located within less arid climates or under non-drought conditions. A need exists for additional research to examine these relationships under a broader range of annual and regional climate scenarios.
Although our results are supportive of the role of stand basal area modifying soil moisture conditions and their influence on foliar moisture content, there are other potential mechanisms that could explain the observed differences. An alternative possibility is that thinned and intact stands similarly had higher leaf area at the tree-scale that resulted in lower foliar moisture content compared with encroached stands in both Douglas-fir and Oregon white oak. This may be plausible for Oregon white oak in our study because of their ability to epicormically sprout following thinning treatments (Devine and Harrington 2006); however, we observed similar foliar moisture content differences between thinned and encroached stands for Douglas-fir, a species that is a weak epicormic sprouter (Punches and Puettmann 2018). Similarly, our study had some potentially confounding effects in our comparisons between intact stands and encroached stands because of differences in tree heights and crown characteristics. However, our comparisons between encroached and thinned stands remain robust because of limited tree and crown differences between conditions for both species examined.
Another possible explanation is that intact and thinned stands differed in rooting depth and access to deeper water sources, which were not directly examined in our study. However, a complementary study that examined the depth of water source based on isotopic analysis at the same study site did not find differences between intact and high encroached stands for both Oregon white oak and Douglas-fir (Goff 2021), and thinned stands should not meaningfully differ in water source in either species 1 year after thinning, assuming rooting depth remains somewhat static. Still, future research that more explicitly examines these and other possible mechanisms to explain observed differences in foliar moisture content – and the context (e.g. climate, species, and stand density) in which they may occur – is warranted.
Our results contrasted previous findings that indicated thinning treatments did not influence leaf water potential or foliar moisture content. Where we found lower foliar moisture content in thinned stands compared with encroached stands for both Douglas-fir and Oregon white oak, differences in foliar moisture content between thinned and unthinned stands of maritime pine (Pinus pinaster) were not detected (Soler Martin et al. 2017). We suspect this discrepancy reflects climatic differences between the two studies. Although both studies have mediterranean climates with pronounced summer dry seasons, our site in northwestern California, USA was likely more moderated due to coastal proximity and the influence of summertime fog. Another possibility is that the duration of thinning effects of foliar moisture content declines over time. Our study was implemented 1 year after thinning, whereas the Soler Martin et al. (2017) study was implemented 7 years after thinning. It is possible that the influence of thinning decreases over time because tree species equilibrate foliar moisture content through increased leaf area and compensation of physiological processes (e.g. stomatal control, osmotic potential). The declining influence of thinning has been observed with growth responses in trees (Sohn et al. 2016), but further examination of the role of thinning across differing climates and its impact on leaf area, tree physiology, and foliar moisture content are warranted, with particular attention to how these responses persist or vary with time.
Seasonal patterns of foliar moisture were generally consistent among stand conditions but differed with foliage type. Foliar moisture content of new foliage in Douglas-fir and Oregon white oak was highest in late spring/early summer and steadily decreased over the prolonged dry summer, as has been found in other species (Agee et al. 2002). Old Douglas-fir foliage was lowest at the start of the fire season and generally increased over time. However, we recognise that a portion of the observed seasonal changes in foliar moisture content likely coincided with dry mass changes due to leaf phenology (Kozlowski and Clausen 1965; Jolly et al. 2014), but these phenological changes were not examined as part of our study.
The effects of species (Douglas-fir and Oregon white oak) on foliar moisture content, independent of foliage age, were persistent in our study, where new Douglas-fir foliage maintained higher foliar moisture content than new Oregon white oak foliage. Differences in foliar moisture content between these two species are expected, given the physiological, phenological, and structural (e.g. root depth) differences between them that relate to plant water use and drought tolerance strategies. It is well known that Oregon white oak is much more drought tolerant than Douglas-fir (Niinemets and Valladares 2006; Beckmann et al. 2021). A complementary study conducted at the same site found that Oregon white oak was accessing soil water at deeper soil depths than Douglas-fir (Goff 2021), partially reflecting differences in tree ages between species at our sites (Douglas-fir were around 70 years old whereas Oregon white oak were around 120–170 years old). Interestingly, in our study, Oregon white oak had lower foliar moisture content compared with Douglas-fir, contradicting previous research that has shown shallower rooted species tend to have lower foliar moisture content (Brown et al. 2022). It is therefore more likely that our observed differences in foliar moisture content between these two species reflect anatomical, morphological (e.g. leaf mass area), and physiological differences that relate to plant hydraulic strategies (Kelly 2016; Hahm et al. 2018; Beckmann et al. 2021).
Our observations of foliar moisture content in Douglas-fir ranged between 150 and 300% in new foliage and 100–150% in old foliage between June and November. These values were slightly higher than recorded in a previous study conducted from July to September in Montana, which had values between 120 and 200% and 80 and 120% for new and old Douglas-fir foliage, respectively (Philpot and Mutch 1971). Slight differences in the range of foliar moisture content between studies likely reflect a combination of sample timing and climatic differences between the two sites, with our site in California having relatively milder temperatures during the fire season due in part to coastal proximity. We are unaware of prior studies that have examined foliar moisture content in Oregon white oak. However, foliar moisture content of deciduous broadleaf tree species in Canada ranged between 130 and 250% from spring to summer (Van Wagner 1967), which were slightly higher than our observations in Oregon white oak (90 and 220%) over the same time period.
We found that differences in Oregon white oak foliar moisture content were associated with tree physiological measures that varied with stand condition. Midday leaf water potential (rather than predawn leaf water potential) was significantly related to foliar moisture content in Oregon white oak leaves, but was contingent on stand condition. Previous studies in other fire-prone ecosystems have found predawn leaf water potential to be better associated with foliar moisture content (Nolan et al. 2018). We are uncertain why foliar moisture content was better related to midday leaf water potential and only in the high encroached stands of our study. The stronger role of midday leaf water potential in high encroached stands may indicate that foliar moisture content is more responsive to diel changes in stand microclimate conditions than previously appreciated, but a deeper examination is necessary. Additionally, we only examined leaf water potential among stands on 1 day, and it is possible that this relationship may shift under wetter or drier conditions within or among years.
In our study, we found that tree- and stand-level factors can influence foliar moisture content within an Oregon white oak ecosystem of northern California. Our findings provide further validation of previous calls to better integrate ecophysiological approaches and concepts to understand foliar moisture content dynamics (Jolly and Johnson 2018; Nolan et al. 2020). Improved estimates of foliar moisture content and the role of stand conditions can also provide managers and fire behaviour analysts with better assumed values for modelling potential crown fire behaviour, and for fuel management planning. Given the increased incidence of fire activity and severity in fire-prone ecosystems over the past few decades (Westerling 2016; Parks and Abatzoglou 2020), there is a strong need to better predict crown fire behaviour and its contributing factors, such as foliar moisture content. This need is likely to increase over the coming decades due in part to the potential for climate change to extend the duration of low foliar moisture content below critical thresholds of crown fire ignition and spread (Ma et al. 2021).
Data availability
Data are available at https://doi.org/10.6084/m9.figshare.20485665 (Kane et al. 2022).
Conflicts of interest
The authors declare no conflicts of interest associated with this work.
Declaration of funding
This work was supported by the USDA National Institute of Food and Agriculture, McIntire-Stennis Cooperative Forestry Research Program, Project # CALZ-164.
Acknowledgements
M. Miles and Humboldt Redwood Company provided access to the research site and coordinated the thinning treatments. M. Lopez, C. Nichols, and E. Hoeft collected and processed field and lab data. This manuscript was improved based on comments from two anonymous reviewers.
References
Agee JK (1993) ‘Fire Ecology of the Pacific Northwest.’ (Island Press: Washington, DC, USA.)Agee JK, Wright CS, Williamson N, Huff MH (2002) Foliar moisture content of Pacific Northwest vegetation and its relation to wildland fire behavior. Forest Ecology and Management 167, 57–66.
| Foliar moisture content of Pacific Northwest vegetation and its relation to wildland fire behavior.Crossref | GoogleScholarGoogle Scholar |
Alexander ME, Cruz MG (2013) Assessing the effect of foliar moisture on the spread rate of crown fires. International Journal of Wildland Fire 22, 415–427.
| Assessing the effect of foliar moisture on the spread rate of crown fires.Crossref | GoogleScholarGoogle Scholar |
Balaguer-Romano R, Díaz-Sierra R, De Cáceres M, Cunill-Camprubí À, Nolan RH, Boer MM, Voltas J, Resco de Dios V (2022) A semi-mechanistic model for predicting daily variations in species-level live fuel moisture content. Agricultural and Forest Meteorology 323, 109022
| A semi-mechanistic model for predicting daily variations in species-level live fuel moisture content.Crossref | GoogleScholarGoogle Scholar |
Bates D, Maechler M, Bolker B, Walker S, Christensen RHB, Singmann H, Grothendieck G (2015) ‘Linear mixed-effects models using eigen and S4.’ (R Foundation for Statistical Computing: Vienna, Austria) Available at cran.r-project.org/web/packages/lme4
Beckmann JJ, Sherriff RL, Kerhoulas LP, Kane JM (2021) Douglas-fir encroachment reduces drought resistance in Oregon white oak of northern California. Forest Ecology and Management 498, 119543
| Douglas-fir encroachment reduces drought resistance in Oregon white oak of northern California.Crossref | GoogleScholarGoogle Scholar |
Bradshaw LS, Deeming JE, Burgan RE, Cohen JD (1983) The 1978 National Fire-Danger Rating System: technical documentation. INT-GTR-169. (US Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA)
| Crossref |
Brooks JR, Mitchell AK (2011) Interpreting tree responses to thinning and fertilization using tree‐ring stable isotopes. New Phytologist 190, 770–782.
| Interpreting tree responses to thinning and fertilization using tree‐ring stable isotopes.Crossref | GoogleScholarGoogle Scholar |
Brown TP, Hoylman ZH, Conrad E, Holden Z, Jencso K, Jolly WM (2022) Decoupling between soil moisture and biomass drives seasonal variations in live fuel moisture across co-occurring plant functional types. Fire Ecology 18, 14
| Decoupling between soil moisture and biomass drives seasonal variations in live fuel moisture across co-occurring plant functional types.Crossref | GoogleScholarGoogle Scholar |
Burnham KP, Anderson DR (1998) ‘Model selection and inference: a practical information-theoretic approach.’ (Springer: New York, NY, USA)
CALFIRE (2020) CalFire FRAP-fire perimeters. Available at https://frap.fire.ca.gov/frap-projects/fire-perimeters/
Dennison PE, Moritz MA, Taylor RS (2008) Evaluating predictive models of critical live fuel moisture in the Santa Monica Mountains, California. International Journal of Wildland Fire 17, 18–27.
| Evaluating predictive models of critical live fuel moisture in the Santa Monica Mountains, California.Crossref | GoogleScholarGoogle Scholar |
Devine WD, Harrington CA (2006) Changes in Oregon white oak (Quercus garryana Dougl. ex Hook.) following release from overtopping conifers. Trees 20, 747–756.
| Changes in Oregon white oak (Quercus garryana Dougl. ex Hook.) following release from overtopping conifers.Crossref | GoogleScholarGoogle Scholar |
Devine WD, Harrington CA (2007) Release of Oregon white oak from overtopping Douglas-fir: effects on soil water and microclimate. Northwest Science 81, 112–124.
| Release of Oregon white oak from overtopping Douglas-fir: effects on soil water and microclimate.Crossref | GoogleScholarGoogle Scholar |
Devine WD, Harrington CA (2013) Restoration release of overtopped Oregon white oak increases 10-year growth and acorn production. Forest Ecology and Management 291, 87–95.
| Restoration release of overtopped Oregon white oak increases 10-year growth and acorn production.Crossref | GoogleScholarGoogle Scholar |
Dimitrakopoulos AP, Papaioannou KK (2001) Flammability assessment of Mediterranean forest fuels. Fire Technology 37, 143–152.
| Flammability assessment of Mediterranean forest fuels.Crossref | GoogleScholarGoogle Scholar |
Engber EA, Varner JM (2012) Predicting Douglas-fir sapling mortality following prescribed fire in an encroached grassland. Restoration Ecology 20, 665–668.
| Predicting Douglas-fir sapling mortality following prescribed fire in an encroached grassland.Crossref | GoogleScholarGoogle Scholar |
Engber EA, Varner III JM, Arguello LA, Sugihara NG (2011) The effects of conifer encroachment and overstory structure on fuels and fire in an oak woodland landscape. Fire Ecology 7, 32–50.
| The effects of conifer encroachment and overstory structure on fuels and fire in an oak woodland landscape.Crossref | GoogleScholarGoogle Scholar |
Fox J, Weisberg S (2019) ‘An R companion to applied regression.’ (Sage: Thousand Oaks, CA, USA)
Fritschle JA (2008) Reconstructing historic ecotones using the public land survey: the lost prairies of Redwood National Park. Annals of the Association of American Geographers 98, 24–39.
| Reconstructing historic ecotones using the public land survey: the lost prairies of Redwood National Park.Crossref | GoogleScholarGoogle Scholar |
Goff GS (2021) Conifer encroachment and removal in a northern California oak woodland: influences on ecosystem physiology and biodiversity. Humboldt State University, Arcata, CA, USA.
Gould PJ, Harrington CA, Devine WD (2011) Growth of Oregon white oak (Quercus garryana). Northwest Science 85, 159–171.
| Growth of Oregon white oak (Quercus garryana).Crossref | GoogleScholarGoogle Scholar |
Hahm WJ, Dietrich WE, Dawson TE (2018) Controls on the distribution and resilience of Quercus garryana: ecophysiological evidence of oak’s water-limitation tolerance. Ecosphere 9, e02218
| Controls on the distribution and resilience of Quercus garryana: ecophysiological evidence of oak’s water-limitation tolerance.Crossref | GoogleScholarGoogle Scholar |
Hair JF, Black WC, Babin B, Anderson R (2018) ‘Multivariate data analysis.’ (Cengage: UK)
Hamman ST, Dunwiddie PW, Nuckols JL, McKinley M (2011) Fire as a restoration tool in Pacific Northwest prairies and oak woodlands: challenges, successes, and future directions. Northwest Science 85, 317–328.
| Fire as a restoration tool in Pacific Northwest prairies and oak woodlands: challenges, successes, and future directions.Crossref | GoogleScholarGoogle Scholar |
Jameson DA (1966) Diurnal and seasonal fluctuations in moisture content of pinyon and juniper. RMRS-RN-67. (US Department of Agriculture Forest Service Research Note, Rocky Mountain Research Station: Fort Collins, CO, USA)
Jolly WM, Johnson DM (2018) Pyro-ecophysiology: shifting the paradigm of live wildland fuel research. Fire 1, 8
| Pyro-ecophysiology: shifting the paradigm of live wildland fuel research.Crossref | GoogleScholarGoogle Scholar |
Jolly WM, Hadlow AM, Huguet K (2014) De-coupling seasonal changes in water content and dry matter to predict live conifer foliar moisture content. International Journal of Wildland Fire 23, 480–489.
| De-coupling seasonal changes in water content and dry matter to predict live conifer foliar moisture content.Crossref | GoogleScholarGoogle Scholar |
Kane JM (2021) Stand conditions alter seasonal microclimate and dead fuel moisture in a Northwestern California oak woodland. Agricultural and Forest Meteorology 308–309, 108602
| Stand conditions alter seasonal microclimate and dead fuel moisture in a Northwestern California oak woodland.Crossref | GoogleScholarGoogle Scholar |
Kane JM, Engber EA, McClelland JE (2019) Effectiveness and impacts of girdling treatments in a conifer-encroached Oregon white oak woodland. Forest Ecology and Management 447, 77–86.
| Effectiveness and impacts of girdling treatments in a conifer-encroached Oregon white oak woodland.Crossref | GoogleScholarGoogle Scholar |
Kane JM, Kerhoulas LP, Goff GS (2022) Douglas-fir and Oregon white oak physiology and foliar moisture content data. Figshare.
| Crossref |
Kelly J (2016) Physiological responses to drought in healthy and stressed trees: a comparison of four species in Oregon, USA. Masters thesis, Lund University, Lund, Sweden.
Keyes CR (2006) Role of foliar moisture content in the silvicultural management of forest fuels. Western Journal of Applied Forestry 21, 228–231.
| Role of foliar moisture content in the silvicultural management of forest fuels.Crossref | GoogleScholarGoogle Scholar |
Knight CA, Cogbill CV, Potts MD, Wanket JA, Battles JJ (2020) Settlement‐era forest structure and composition in the Klamath Mountains: reconstructing a historical baseline. Ecosphere 11, e03250
| Settlement‐era forest structure and composition in the Klamath Mountains: reconstructing a historical baseline.Crossref | GoogleScholarGoogle Scholar |
Kozlowski TT, Clausen JJ (1965) Changes in moisture contents and dry weights of buds and leaves of forest trees. Botanical Gazette 126, 20–26.
| Changes in moisture contents and dry weights of buds and leaves of forest trees.Crossref | GoogleScholarGoogle Scholar |
Krix DW, Murray BR (2018) Landscape variation in plant leaf flammability is driven by leaf traits responding to environmental gradients. Ecosphere 9, e02093
| Landscape variation in plant leaf flammability is driven by leaf traits responding to environmental gradients.Crossref | GoogleScholarGoogle Scholar |
Kuljian H, Varner JM (2010) The effects of sudden oak death on foliar moisture content and crown fire potential in tanoak. Forest Ecology and Management 259, 2103–2110.
| The effects of sudden oak death on foliar moisture content and crown fire potential in tanoak.Crossref | GoogleScholarGoogle Scholar |
Lagergren F, Lankreijer H, Kučera J, Cienciala E, Mölder M, Lindroth A (2008) Thinning effects on pine–spruce forest transpiration in central Sweden. Forest Ecology and Management 255, 2312–2323.
| Thinning effects on pine–spruce forest transpiration in central Sweden.Crossref | GoogleScholarGoogle Scholar |
LANDFIRE (2020) LANDFIRE-Fire Return Interval. Available at https://landfire.gov/fri.php
Ma S, Concilio A, Oakley B, North M, Chen J (2010) Spatial variability in microclimate in a mixed-conifer forest before and after thinning and burning treatments. Forest Ecology and Management 259, 904–915.
| Spatial variability in microclimate in a mixed-conifer forest before and after thinning and burning treatments.Crossref | GoogleScholarGoogle Scholar |
Ma W, Zhai L, Pivovaroff A, Shuman J, Buotte P, Ding J, Christoffersen B, Knox R, Moritz M, Fisher RA, Koven CD, Kueppers L, Xu C (2021) Assessing climate change impacts on live fuel moisture and wildfire risk using a hydrodynamic vegetation model. Biogeosciences 18, 4005–4020.
| Assessing climate change impacts on live fuel moisture and wildfire risk using a hydrodynamic vegetation model.Crossref | GoogleScholarGoogle Scholar |
Matthews S (2010) Effect of drying temperature on fuel moisture content measurements. International Journal of Wildland Fire 19, 800–802.
| Effect of drying temperature on fuel moisture content measurements.Crossref | GoogleScholarGoogle Scholar |
McDowell NG, Adams HD, Bailey JD, Hess M, Kolb TE (2006) Homeostatic maintenance of ponderosa pine gas exchange in response to stand density changes. Ecological Applications 16, 1164–1182.
| Homeostatic maintenance of ponderosa pine gas exchange in response to stand density changes.Crossref | GoogleScholarGoogle Scholar |
McNamara BA, Kane JM, Greene DF (2019) Post-fire fuel succession in a rare California, USA, closed-cone conifer. Fire Ecology 15, 39
| Post-fire fuel succession in a rare California, USA, closed-cone conifer.Crossref | GoogleScholarGoogle Scholar |
Moreno G, Cubera E (2008) Impact of stand density on water status and leaf gas exchange in Quercus ilex. Forest Ecology and Management 254, 74–84.
| Impact of stand density on water status and leaf gas exchange in Quercus ilex.Crossref | GoogleScholarGoogle Scholar |
Niinemets Ü, Valladares F (2006) Tolerance to shade, drought, and waterlogging of temperate Northern Hemisphere trees and shrubs. Ecological Monographs 76, 521–547.
| Tolerance to shade, drought, and waterlogging of temperate Northern Hemisphere trees and shrubs.Crossref | GoogleScholarGoogle Scholar |
Nolan RH, Boer MM, Resco de Dios V, Caccamo G, Bradstock RA (2016) Large-scale, dynamic transformations in fuel moisture drive wildfire activity across southeastern Australia: transformations in Fuel Moisture. Geophysical Research Letters 43, 4229–4238.
| Large-scale, dynamic transformations in fuel moisture drive wildfire activity across southeastern Australia: transformations in Fuel Moisture.Crossref | GoogleScholarGoogle Scholar |
Nolan RH, Hedo J, Arteaga C, Sugai T, Resco de Dios V (2018) Physiological drought responses improve predictions of live fuel moisture dynamics in a Mediterranean forest. Agricultural and Forest Meteorology 263, 417–427.
| Physiological drought responses improve predictions of live fuel moisture dynamics in a Mediterranean forest.Crossref | GoogleScholarGoogle Scholar |
Nolan RH, Blackman CJ, de Dios VR, Choat B, Medlyn BE, Li X, Bradstock RA, Boer MM (2020) Linking forest flammability and plant vulnerability to drought. Forests 11, 779
| Linking forest flammability and plant vulnerability to drought.Crossref | GoogleScholarGoogle Scholar |
Pallardy SG (2008) ‘Physiology of woody plants.’ (Elsevier: Amsterdam, Netherlands)
Parks SA, Abatzoglou JT (2020) Warmer and drier fire seasons contribute to increases in area burned at high severity in western US forests from 1985 to 2017. Geophysical Research Letters 47, e2020GL089858
| Warmer and drier fire seasons contribute to increases in area burned at high severity in western US forests from 1985 to 2017.Crossref | GoogleScholarGoogle Scholar |
Pellizzaro G, Duce P, Ventura A, Zara P (2007) Seasonal variations of live moisture content and ignitability in shrubs of the Mediterranean Basin. International Journal of Wildland Fire 16, 633–641.
| Seasonal variations of live moisture content and ignitability in shrubs of the Mediterranean Basin.Crossref | GoogleScholarGoogle Scholar |
Philpot CW, Mutch RW (1971) ‘The seasonal trends in moisture content, ether extractives, and energy of ponderosa pine and Douglas-fir needles.’ (Intermountain Forest & Range Experiment Station, Forest Service, US Department of Agriculture: Ogden, UT, USA)
| Crossref |
Pimont F, Ruffault J, Martin-StPaul NK, Dupuy JL (2019) A cautionary note regarding the use of cumulative burnt areas for the determination of fire danger index breakpoints. International Journal of Wildland Fire 28, 254–258.
| A cautionary note regarding the use of cumulative burnt areas for the determination of fire danger index breakpoints.Crossref | GoogleScholarGoogle Scholar |
Pivovaroff AL, Emery N, Sharifi MR, Witter M, Keeley JE, Rundel PW (2019) The effect of ecophysiological traits on live fuel moisture content. Fire 2, 28
| The effect of ecophysiological traits on live fuel moisture content.Crossref | GoogleScholarGoogle Scholar |
PRISM Climate Group (2020) PRISM Climate Data. Oregon State University, data created 4 Feb 2014, accessed 16 Dec 2020. Available at http://www.prism.oregonstate.edu/
Punches JW, Puettmann KJ (2018) Distribution of epicormic branches and foliage on Douglas-fir as influenced by adjacent canopy gaps. Canadian Journal of Forest Research 48, 1320–1330.
| Distribution of epicormic branches and foliage on Douglas-fir as influenced by adjacent canopy gaps.Crossref | GoogleScholarGoogle Scholar |
R Development Core Team (2022) ‘R: A language and environment for statistical computing.’ (R Foundation for Statistical Computing: Vienna, Austria)
Rossa C, Fernandes P (2018) Live fuel moisture content: the ‘pea under the mattress’ of fire spread rate modeling? Fire 1, 43
| Live fuel moisture content: the ‘pea under the mattress’ of fire spread rate modeling?Crossref | GoogleScholarGoogle Scholar |
Ruffault J, Pimont F, Cochard H, Dupuy J-L, Martin-StPaul N (2022) SurEau-Ecos v2.0: a trait-based plant hydraulics model for simulations of plant water status and drought-induced mortality at the ecosystem level. Geoscientific Model Development 15, 5593–5626.
| SurEau-Ecos v2.0: a trait-based plant hydraulics model for simulations of plant water status and drought-induced mortality at the ecosystem level.Crossref | GoogleScholarGoogle Scholar |
Schriver M, Sherriff RL, Varner JM, Quinn-Davidson L, Valachovic Y (2018) Age and stand structure of oak woodlands along a gradient of conifer encroachment in northwestern California. Ecosphere 9, e02446
| Age and stand structure of oak woodlands along a gradient of conifer encroachment in northwestern California.Crossref | GoogleScholarGoogle Scholar |
Scott JH, Reinhardt ED (2001) Assessing crown fire potential by linking models of surface and crown fire behavior. RMRS-RP-29. (US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ft. Collins, CO, USA)
| Crossref |
Simms DL, Law M (1967) The ignition of wet and dry wood by radiation. Combustion and Flame 11, 377–388.
| The ignition of wet and dry wood by radiation.Crossref | GoogleScholarGoogle Scholar |
Sohn JA, Saha S, Bauhus J (2016) Potential of forest thinning to mitigate drought stress: A meta-analysis. Forest Ecology and Management 380, 261–273.
| Potential of forest thinning to mitigate drought stress: A meta-analysis.Crossref | GoogleScholarGoogle Scholar |
Soler Martin M, Bonet JA, Martínez De Aragón J, Voltas J, Coll L, Resco De Dios V (2017) Crown bulk density and fuel moisture dynamics in Pinus pinaster stands are neither modified by thinning nor captured by the Forest Fire Weather Index. Annals of Forest Science 74, 51
| Crown bulk density and fuel moisture dynamics in Pinus pinaster stands are neither modified by thinning nor captured by the Forest Fire Weather Index.Crossref | GoogleScholarGoogle Scholar |
Tveten RK, Fonda RW (1999) Fire effects on prairies and oak woodlands on Fort Lewis, Washington. Northwest Science 73, 145–158.
Tyree MT, Hammel HT (1972) The measurement of the turgor pressure and the water relations of plants by the pressure-bomb technique. Journal of Experimental Botany 23, 267–282.
| The measurement of the turgor pressure and the water relations of plants by the pressure-bomb technique.Crossref | GoogleScholarGoogle Scholar |
Underwood S, Arguello L, Siefkin N (2003) Restoring ethnographic landscapes and natural elements in Redwood National Park. Ecological Restoration 21, 278–283.
| Restoring ethnographic landscapes and natural elements in Redwood National Park.Crossref | GoogleScholarGoogle Scholar |
Van Wagner CE (1967) ‘Seasonal variation in moisture content of eastern Canadian tree foliage and the possible effect on crown fires.’ (Canada Department of Forestry and Rural Development, Forestry Branch Departmental Publication 1204: Ottawa, Canada)
Van Wagner CE (1977) Conditions for the start and spread of crown fire. Canadian Journal of Forest Research 7, 23–34.
| Conditions for the start and spread of crown fire.Crossref | GoogleScholarGoogle Scholar |
Viegas DX, Piñol J, Viegas MT, Ogaya R (2001) Estimating live fine fuels moisture content using meteorologically-based indices. International Journal of Wildland Fire 10, 223–240.
| Estimating live fine fuels moisture content using meteorologically-based indices.Crossref | GoogleScholarGoogle Scholar |
Weise DR, White RH, Beall FC, Etlinger M (2005) Use of the cone calorimeter to detect seasonal differences in selected combustion characteristics of ornamental vegetation. International Journal of Wildland Fire 14, 321–338.
| Use of the cone calorimeter to detect seasonal differences in selected combustion characteristics of ornamental vegetation.Crossref | GoogleScholarGoogle Scholar |
Westerling AL (2016) Increasing western US forest wildfire activity: sensitivity to changes in the timing of spring. Philosophical Transactions of the Royal Society B: Biological Sciences 371, 20150178
| Increasing western US forest wildfire activity: sensitivity to changes in the timing of spring.Crossref | GoogleScholarGoogle Scholar |
Xanthopoulos G, Wakimoto RH (1993) A time to ignition–temperature–moisture relationship for branches of three western conifers. Canadian Journal of Forest Research 23, 253–258.
| A time to ignition–temperature–moisture relationship for branches of three western conifers.Crossref | GoogleScholarGoogle Scholar |
Yebra M, Chuvieco E, Riaño D (2008) Estimation of live fuel moisture content from MODIS images for fire risk assessment. Agricultural and Forest Meteorology 148, 523–536.
| Estimation of live fuel moisture content from MODIS images for fire risk assessment.Crossref | GoogleScholarGoogle Scholar |