

Examining the usefulness of a Y-maze choice method to measure the preferences of laying hens
N. A. Arnold A and P. H. Hemsworth A BA Animal Welfare Science Centre, University of Melbourne, Parkville, Vic. 3010, Australia.
B Corresponding author. Email: phh@unimelb.edu.au
Animal Production Science 53(12) 1283-1290 https://doi.org/10.1071/AN12390
Submitted: 7 November 2012 Accepted: 9 July 2013 Published: 24 September 2013
Journal Compilation © CSIRO Publishing 2013 Open Access CC BY-NC-ND
Abstract
Measurement of animal preferences can be used as an indirect, but persuasive, method for assessing animal welfare on the basis that preferences may identify resources and behaviours that might be important to animals. The present experiment examined the usefulness of a Y-maze methodology, incorporating alteration of motivational state through prior restriction of resources of potentially differing value in assessing and understanding animal preferences. The choice behaviour of laying hens for feed, a dustbath substrate (sawdust; ‘dust’) and social contact was measured under three pairwise comparisons in a Y-maze apparatus. In each of the three experiments, 48 birds (HyLine Brown Strain; n = 24 per experiment) were offered a choice of two resources in a Y-maze test, with one of the three possible resource pairings. In each experiment, after Y-maze training, an equal number of birds was deprived in a factorial design of Resource 1, Resource 2, both, or neither resource. Analysis of choices over 24 trials per bird in each experiment revealed that birds preferred feed over social contact or dust, irrespective of restriction of any of these resources, and further, were quicker (P < 0.01) to make feed choices than dust or social-contact choices. In the social contact and dust comparison, restriction of dust significantly (P < 0.05) increased choice for dust in the 24 trials (38 vs 53% dust choice), suggesting that dust restriction increased the birds’ motivation to access dust. This result potentially highlights the impact of the resource of comparison in pairwise tests on overall choice response to restriction. The inclusion of measurements of speed of movement through the Y maze proved a useful aspect of the methodology, providing results consistent with choice behaviour in all three experiments. Although overall choices for dust and social contact were not significantly (P = 0.328) different from random, birds were quicker (P < 0.05) to make dust choices than social-contact choices, suggesting that speed of choice may be a particularly sensitive correlate of current motivation levels. The consistency of results across these experiments, together with results reported in the literature on laying-hen preferences, suggests that the methodology is a promising option for assessing the relative preference for resources for laying hens. Additional evidence, particularly on the occurrence of abnormal behaviour, stress physiology and health, when restricted of the resource of interest is necessary to provide a more comprehensive assessment of the impact of the restriction on animal welfare.
Introduction
The housing of laying hens in conventional cages is internationally one of the most controversial animal-welfare issues and has a high public profile. This negative public sentiment towards cages has led to major changes in their design. For example, under European poultry welfare Directives, cage-layer systems must provide ‘furniture’, which includes a nest box, dust bath, perch and claw shorteners. Public interest in the welfare of hens housed in indoor systems is likely to increase and, clearly, science has an important role in providing an understanding of the welfare implications of these systems. From an animal-welfare perspective, there is increasing interest in the measurement of the preferences of farm, laboratory and captive animals in relation to requirements within these environments. Preferences can be measured as a means to determine what resources are important to an animal. Initial use of preference methodologies appeared in the literature in the 1970s (e.g. Hughes and Black 1973; Dawkins 1976). These early investigations created much scientific debate relating to conceptual and methodological difficulties (e.g. Dawkins 1977; Duncan 1977; van Rooijen 1982). Nevertheless, preference testing using a Y-maze apparatus that allows a choice between access to two different resources has been used to provide information about specific features in the animal environments, such as flooring on raceways (Hutson 1981), restraint methods (e.g. Pollard et al. 1994), handling treatments (Rushen 1986) and ramp design (Phillips et al. 1988), with the overriding objective of optimising captive environments for animal inhabitants. Essentially, these tests are designed to answer the question ‘what is the relative importance of this feature for this animal?’
While the consistent choice or preference of one resource over another or others indicates the animal’s relative preference, some have argued that in addition to establishing what an animal prefers, it is important to understand the strength of the preference (Dawkins 1983; Matthews and Ladewig 1994). To address the question of the strength of an animal’s preference, experiments have incorporated varying levels of cost (e.g. work effort, time and relinquishing a desirable resource) associated with gaining access to a resource or avoiding aversive stimulation. These ‘behavioural demand’ studies have been used to study the animal’s level of motivation to access or avoid the situation being tested and the strength of the motivation provides a quantitative measure of how much it matters to the animal (Fraser and Matthews 1997).
One possible way to more closely assess the resource value (for the animal) through preference testing is to experimentally alter motivational state. This can be achieved by restricting access to a resource (such as feed) and observing the impact of prior resource restriction on subsequent choice behaviour in a Y maze offering a choice of two different resources. Barnard (2004) defined motivation as the internal decision-making process an animal uses to make a choice about its behaviour. Motivation to choose a resource in a Y-maze preference test will be affected by the resource utility (or value of the reward), which will be influenced by several factors, such as duration of access to the resource, degree of prior restriction, specific characteristics of the resource (e.g. type of dustbath substrate) and comparative utility value of the alternative resource on offer. Thus, if under either conditions of no prior restriction, birds choose one resource (R1) over the other (R2) in a series of Y-maze trials, then it seems likely that, in the context of the specific utility offered for each resource, birds are less motivated to choose R2 than R1. Preferences can be further investigated by conducting the same choice tests under conditions of higher levels of motivation for access to each resource, achieved through restricting access to the resources before testing.
The present experiments examined soundness of this combination of a Y-maze methodology with resource restriction to alter motivational state in assessing the relative preferences of laying hens for feed, social contact and a dustbath substrate. On the basis of limited literature on the strength of preferences for food and litter, if the combination of preference testing with experimental alteration of motivational state is a sound method in assessing the value of the resource to laying hens, it is proposed that food will be predominantly chosen irrespective of the restrictions on the other two resources. For example, Dawkins (1983) varied the price paid for access to litter by increasing the duration of feed withdrawal before the test. Results indicated that although hens preferred litter to wire floors, their preference for litter was not strong enough to outweigh the attraction of food, and Dawkins (1983) concluded that there was no evidence that hens regarded litter as a necessity. Food can be considered as the ‘gold standard’ in preference testing (Matthews and Ladewig1994), and therefore is generally expected to produce a maximal response or preference.
Materials and methods
Subjects and housing
Forty-eight commercial laying hens (HyLine Brown Strain) were individually housed indoors in furnished cages (57 cm × 50 cm × 48 cm), each containing a dustbath at a floor level. The birds were randomly allocated to three groups of 16 birds, for use in three separate experiments. The birds were 41, 48 and 54 weeks of age at the commencement of the pre-experimental phase for Experiments 1, 2 and 3, respectively, and had not been raised with dustbath access. Commercial layer pellets (15% crude protein) were available ad libitum externally at the front of the cage and water was available ad libitum via a drinker at the rear of the cage. All birds had auditory and olfactory contact with other birds and visual contact with neighbouring birds, as well as some limited physical contact through mesh dividers between pairs of cages. The hens were kept in a constant environmental temperature of 21°C and a light : dark cycle of 16 : 8 h.
Y-maze apparatus
The Y maze was a purpose-built apparatus with solid galvanised steel walls, removable mesh roofing and a floor constructed of mesh identical to flooring in the birds’ home cages (see Fig. 1). The gate between the start box and choice area was also mesh, allowing visual contact with maze arms from the start box. The gates between the choice area and each arm were solid metal. All gates were pulley operated and could be opened and closed by one operator standing in front of the start box. The maze was raised 49 cm off the ground and was positioned in close proximity to the birds’ home cages, in the same temperature-controlled room.
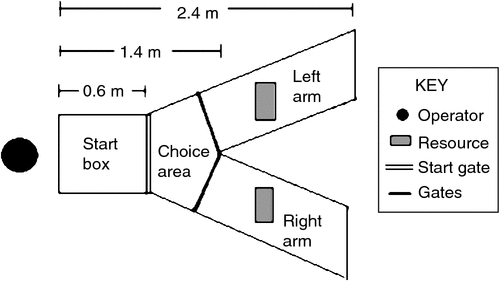
|
Procedure
Pre-experimental phase
Birds were housed in the individual cages with dustbaths (containing sawdust) for 3 weeks before commencement of the experiment. During this period, all birds were familiarised with human handling through incremental exposure to increasing intensity of human contact. Initially, over a period of 3 days, each cage was opened and birds were exposed for 30 s per day to a handler moving a hand around in the cage. This was followed by 5 days of physically touching and stroking the birds on the back for 30 s per day, and finally picking the birds up and holding them for 30 s per day for the 7 days before the experiment commenced.
Familiarisation to the Y maze
All birds were exposed to the Y-maze environment over 3 days, with no resource in either maze arm. During this phase, the gates between the choice area and each arm were left open, but the start-box gate was initially closed. Each bird was carried from its home cage and placed in the start box of the maze for 20 s before the start-box gate was opened. Birds were allowed 30 s to move out of the start box before movement was gently encouraged by pushing the bird forward with a flat hand on the tail. Once in the choice area, the start-box gate was closed and birds were allowed another 30 s to move into a maze arm before movement was encouraged by a gentle push using the least amount of pressure necessary to generate movement. In the majority of instances, a light touch on the tail feathers was sufficient to achieve this. Birds were pushed on the tail every 3 s until movement occurred, regardless of the orientation of the bird in relation to the Y-maze arms (i.e. pushes were dictated by time rather than position of the bird). Once in an arm, access to the opposite maze arm was closed using the appropriate gate and birds were left in the maze for 2 min before being carried back to the start box for the next trial (or to the home cage when the sequence of trials was complete). Each bird was exposed to the maze in this way for three consecutive trials each day over the 3 days; thus, each bird received one free choice, followed by two directed exposures (one to each arm of the maze, achieved by blocking access to the opposite arm). Birds were introduced to the maze in a different random order each day.
The selection of a 2-min exposure to the chosen resource was made on the basis of relevant literature. Although 2 min with dustbathing substrate is substantially less than what hens are expected to spend in a dustbathing bout in an unrestricted situation (27 min, Vestergaard 1982), recent research by Laine et al. (2008) found that 2 or 45 min of confinement with peat moss in a series of Y-maze trials, in which choice of peat moss or social contact was studied, did not affect choice behaviour.
Training phase
During the training phase, birds were deprived of feed and dustbath access for 30 min before each Y-maze training session, so as to standardise exposure to these resources. A comparable withdrawal of access to social contact was not practicable in the housing arrangement.
Training took place over 2 days with four consecutive trials per bird per day. In each trial, only one arm of the maze was available, with the other being closed off by the appropriate gate between the choice area and arm. The available maze arm was alternated across the four trials each day, so that each bird was exposed to each arm twice per day.
Resource 1 was presented in one maze arm and Resource 2 in the other (see Table 1 for resource descriptions). Pairing of maze arm and resource was partially randomly assigned to each bird, such that half the birds received each of the two possible configurations and the pairing remained consistent for each bird within each experiment.

|
The procedure for each trial was identical to that described in the familiarisation phase (however, once birds entered the available maze arm there was no need to shut the opposite gate as it was already closed). As with familiarisation, bird order was randomised each day.
Testing phase
In each of three experiments, the effects of restrictions (see Table 1) of the two resources of interest on preferences were studied in a 2 × 2 factorial design over 12 days, such that four birds were deprived of both resources, four were deprived of Resource 1 only, four deprived of Resource 2 only, and four were not deprived of either resource. The description of the two resources and their restrictions studied in each experiment are explained below.
Experiment 1. Feed vs dust
The two main effects studied were (1) feed at two levels, including (i) feed being available in feed tray of the home cage (‘Unrestricted’) and (ii) access to feed being removed from the home cage for 3 h before the first trial each day, by covering the external feed trough in front of the cage (‘Restricted’); and (2) dustbathing substrate (‘dust’) at two levels, including (i) dust in a dustbathing tray being available in the home cage (‘Unrestricted’) and (ii) the dustbath tray being removed from home cage on the night before Day 1 of testing and not being replaced until the end of the experiment (‘Restricted’).
Experiment 2. Dust vs social contact
The two main effects were (1) dustbathing substrate (‘dust’) at two levels, as described for Experiment 1, and (2) social contact at two levels, including (i) visual contact with neighbouring birds at all times while in the home cage (‘unrestricted’) and (ii) visual contact with neighbouring birds being removed on the day before Day 1 of testing, by covering cage divisions with solid rubber partitions that remained in place until the end of the experiment (‘restricted’).
Experiment 3. Feed vs social contact
The two main effects were feed at two levels, as described in Experiment 1, and social contact at two levels, as described in Experiment 2.
Birds that were not restricted of feed or dust in the three experiments had feed and dustbath removed for 30 min before testing, as in the training phase, so as to standardise contact with these resources immediately before testing.
Testing was conducted over 12 days and consisted of two consecutive trials per bird per day. Again, bird order was randomly generated for each day of testing and, thus, the time of day each bird was tested was randomly assigned each day. Both maze arms were available on every trial and, thus, birds were able to move into either arm from the choice area. Birds that failed to move after 30 s in the start box and/or after 30 s in the choice area were gently pushed, as described in the familiarisation phase. Identical maze arm and resource pairings were maintained for each bird consistent with those used in the training phase for each experiment. The procedure for each trial was identical to that described for the familiarisation phase, and once a maze arm was entered, access to the opposite arm was closed using the appropriate gate.
Times taken to leave the start box and choice area, number of pushes required to leave the choice area and resource chosen were recorded for each trial.
Statistical analyses
For each experiment, an ANOVA was used to examine the two main effects, levels of Resources 1 and 2, and the interactions of these factors, on choice of resource, latency to leave the start box (s), latency to leave the choice area (s) and number of pushes required to move birds out of the choice area after 30 s. Analyses were conducted on these variables calculated for each of the 24 trials for each experiment (2 per day per bird), and for the first trials of each day only (total of 12 trials). The ‘choice of resource’ variable was calculated for Experiment 1 as the proportion of trials in which feed was chosen, for Experiment 2 as the proportion of trials in which dust was chosen, and for Experiment 3 as the proportion of trials in which feed was chosen.
A further set of ANOVAs was run, both on the set of 24 trials and the set of 12 first trials for each experiment, excluding all trials were birds needed to be pushed in the choice area. Trials in which pushes were recorded were treated as missing data for the purposes of these analyses.
Logistic regression using an estimated binomial model was conducted for each experiment to determine whether overall choice data differed significantly from random (i.e. 50%). A one-way ANOVA was conducted for each experiment to determine whether there was an effect of resource type chosen on time to move into a maze arm.
Results
Experiment 1. Feed vs dust
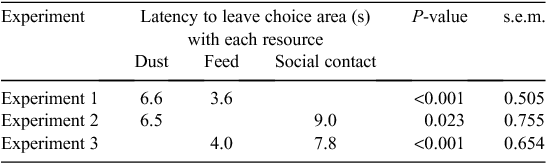
There was a total of 24 trials during the testing phase. The overall proportion of feed choices by hens was 78% of trials, with binomial regression indicating a significant (P < 0.01) difference from 50%. Irrespective of the level of restriction, latency to enter a maze arm from the choice area was greater (P < 0.01) when dust was chosen than when feed was chosen (Table 2).
|
No effects of either feed (P = 0.140) or dust (P = 0.699) restriction were found on choice for feed (Table 3), and there was no interaction effect (P = 0.699). However, when only the first trials of each day were included in the analysis, there was a significant (P = 0.027) effect of feed restriction on choice behaviour (Table 3), with no interaction effect (P = 0.233). Birds that were feed restricted chose feed in the Y maze on a greater proportion of these first trials than did birds that were not restricted.

|
When only trials where no pushes were required were included in the analyses, a similar pattern of results was observed (Table 3), with a significant (P = 0.034) increase in choice for feed by feed restricted birds in the first trials of each day compared with birds that were not feed restricted.
There were no significant treatment effects on mean latency to leave the start box once the gate had been lifted, mean latency to leave the choice area and the number of pushes required (Table 3). However, there was a trend (P = 0.092) indicating a decrease in latency to leave the start box in feed-restricted hens compared with those that were not feed restricted.
Experiment 2. Dust vs social contact
The overall proportion of dust choices by hens was 46%, and this was not significantly (P = 0.377) different from random. Irrespective of the level of restriction, latency to leave the choice area was greater (P < 0.05) when social contact was chosen than when dust was chosen (Table 2).
There was a significant effect of dust restriction on choice of dust (Table 4). Birds that were restricted of dust chose dust on a greater proportion (P = 0.048) of trials than did birds that were not restricted. There was no effect of social contact restriction (P = 0.886) and no interaction effect (P = 0.886). When the analysis was conducted on the first trials only, no significant treatment effects were found for either dust (P = 0.123) or social-contact (P = 1.000) restriction and there was no interaction effect (P = 1.00).

|
A similar pattern of results was observed in the analysis excluding trials where pushes were required, except that the significant effect of dust restriction on dust choice (P = 0.022) was confined to the first trials of each day only (Table 4, in parentheses). There was also a tendency (P = 0.062) for choice of social contact to increase with restriction of social contact when trials with pushes were excluded.
There were no significant (P > 0.05) treatment effects observed on latency to leave the start box, latency to leave the choice area, or the number of pushes required (Table 4). However, there was a trend towards social contact-restricted birds moving faster into the maze arms (P = 0.065) and requiring fewer pushes (P = 0.065) than those with unrestricted social contact. There was also a tendency for fewer pushes being required for dust-restricted birds than for those not restricted of dust (P = 0.090).
Experiment 3. Feed vs social contact
The overall proportion of feed choices was 88%, which significantly (P < 0.001) differed from random. Latency to leave the choice area was reduced (P < 0.01) when feed was chosen than when social contact was chosen, irrespective of the level of restriction (Table 2).
There were no significant effects of either feed (P = 0.634) or social-contact (P = 0.327) restriction on choice for feed (Table 5) and there was no interaction effect (P = 0.615). Similarly, when only the first trials of each day were included in the analysis, no significant effects of either treatment were found (Table 5). A similar pattern of results was observed in the analysis excluding trials where pushes were required (Table 5).

|
There were no significant treatment effects on latency to leave the start box, latency to leave the choice area, or the number of pushes required (Table 5).
Discussion
The present study examined the usefulness of a combination of a Y-maze methodology with experimental alteration of the motivational state in assessing hen preferences for the resources of food, a dust bath and social contact for laying hens. This was achieved by examining the effects of restriction of these resources on choice behaviour relative to the expectation that food would be predominantly chosen regardless of restrictions of the other two resources.
The results of Experiments 1 and 3 showed that birds consistently chose feed over dust or social contact, irrespective of restriction of any of these resources. The proportion of choices for feed in each of these experiments was significantly greater than that for a random choice pattern (i.e. 50%). The results agreed with those of Dawkins (1983) using a behavioural demand approach that showed that although hens preferred litter to wire floors, their preference for litter was not strong enough to outweigh the attraction of food. While some studies have indicated that domestic hens have a strong preference to be near familiar hens over unfamiliar hens (Hughes 1977; Bradshaw 1992), the strength of the preference for social contact has not been studied. However, if, as mentioned earlier, food is generally assumed to generate the maximal response or preference in most cases (i.e. the ‘gold standard’ in preference testing; Matthews and Ladewig 1994), we would expect hens to be highly motivated to choose food over social contact, even with restriction of visual and tactile contact with conspecifics.
It is important to note that the levels of restriction studied in these experiments were arbitrary and the utility (i.e. reward value) of the resource for restricted birds is likely to vary, both due to the levels of these arbitrary restrictions, but also due to other characteristics of the resources, such as the type of dustbath substrate, level of prior experience, and duration of access to the resources on offer. Therefore, any interpretation of the relative attractiveness of the resources, based on the somewhat contrived conditions of the present study, must be made conservatively.
In general, the overall choice for dust and social contact in Experiment 2 was similar (46% vs 54% for dust and social contact, respectively); however, restriction of dust but not social contact increased choice for the restricted resource (53% vs 38% dust choices for dust-restricted and unrestricted, respectively, and 45% vs 46% social-contact choices for social contact-restricted and unrestricted, respectively), suggesting that dust restriction increased the birds’ motivation to access the dustbath substrate. Furthermore, since the overall choice for dust or social contact in Experiment 2 was not significantly different from random (i.e. 50%), the result of an effect of dust restriction, but not social-contact restriction, cannot be attributed to any difference in preference for dust over social contact.
The results of Experiment 1 are consistent with those of Petherick et al. (1993), in which hens chose feed over a sand dustbathing substrate in a Y-maze task when feed deprived but did not increase choices for sand over feed when deprived of sand. Petherick et al. (1993) also reported an effect of resource deprivation on speed of movement into the maze arm, where birds were faster when feed deprived than when they were sand deprived. In contrast, the speed of movement data from the current experiment are equivocal. There was a trend (P = 0.092) towards faster movement in the maze out of the choice area, but not out of the start box, by feed-restricted hens in Experiment 1 (feed vs dust) than by unrestricted hens, but there was no evidence of faster movement in feed-restricted hens in Experiment 3 (feed vs social contact). Further, there were tendencies for hens that were social-contact or dust restricted to move faster and require fewer pushes by the handler in Experiment 2 (dust vs social contact) than for unrestricted birds. It is somewhat surprising that no significant effects of restriction were found on speed of movement in the maze, since similar relationships have been reported by both Petherick et al. (1992, in an alleyway) and Petherick et al. (1993, in a Y maze) and by Laine et al. (2007, using the same strain of bird and apparatus, and a method similar to that in the current study). However, speed of movement may be affected by several other factors that differed between the studies (such as the motivation level, as a result of differing resource utility, prior experience and competing motivations (e.g. fear, exploration)). There is some evidence to suggest that, in addition, bird age may also have affected the speed of movement; Channing et al. (2001) reported both decreased activity in older compared with younger birds, and less time spent feeding in older birds (possibly indicative of a reduced feeding motivation in older compared with younger birds). In Petherick et al. (1992) and Petherick et al. (1993), birds were less than 20 weeks old, and in Laine et al. (2007), birds were 28 weeks old; however, in the current study, birds were markedly older at 41 (Experiment 1) to 54 (Experiment 3) weeks of age.
Despite the lack of effect of resource restriction on speed of movement, it does appear that inclusion of speed-of-movement measurement within the current methodology provided some support for the preference results. There is some evidence to suggest that speed was dependent on the resource chosen, with increased speed for resources that were more frequently chosen by birds. In Experiments 1 and 3, birds were quicker to make feed choices than choices for the alternative resource, supporting the notion that birds had a higher motivation for feed than dust and social contact, regardless of prior restriction. In Experiment 2, birds were quicker to make dust choices than social-contact choices, possibly indicating a higher motivation for dust than social contact, despite no evidence of a preference for dust over social contact in this comparison. It is possible that variation in speed of movement reflected variation in motivation to access the resource in the Y maze, consistent with the finding of an effect of dust restriction on motivation to access dust.
Protocols in the current study included physically pushing birds that did not move after the maximum time allocated for a choice to occur. This action may have interfered with the birds’ choice behaviour. However, when trials where pushes were used were removed from the analyses, the main effects did not change, except for dust choice in Experiment 2 where the increase in dust choice in dust-restricted compared with unrestricted birds disappeared when trials with pushes were removed, although a significant effect of dust restriction was still present on the first trials only. Petherick et al. (1990) reported that pushing hens forward out of the start box in a Y-maze apparatus did not appear to be aversive.
The results of Experiment 2 provided some evidence to suggest that the resource of comparison in a Y-maze test influences the relative attractiveness of the test resource. The finding of an effect of dust restriction on dust choice over social contact differs somewhat from the lack of any similar effect of dust restriction reported by Petherick et al. (1993) on dust choice over feed and by Arnold et al. (2007) on dust choice over an empty maze arm. Possibly, the effect of restriction is stronger if the alternative is a less valued resource (i.e. social contact rather than feed). Indeed, this is consistent with intuitive expectations; however, if this were the case, we would expect an effect of dust restriction on dust choice over an empty maze, which was not observed in Arnold et al. (2007). One explanation for this is the impact of previous experience, which differed between these studies, on dustbathing motivation. Expression of dustbathing behaviour in laying hens can be affected by the degree of prior exposure to dustbath substrates (e.g. Petherick et al. 1995; Larsen et al. 2000; Olsson et al. 2002). Although this phenomenon is generally reported in relation to the impact of experience during rearing, recent experience may also play a role. In Arnold et al. (2007), access to dustbaths in the birds’ home cages was relatively arduous (dustbaths were placed above a nestbox, and accessed via a perch), compared with the current study, where dustbaths were located on the floor of the home cages. This may have led to differences between studies in familiarity with the dustbath substrate, and establishment of dustbathing routines in the days leading up to the Y-maze tests. Future research investigating the administration of dustbaths in caged systems should identify or control for the impact of dustbath location on utilisation and subsequent dustbathing preferences.
The results of all three experiments in the present study were in general agreement, indicating a strong relative preference for feed over dust and social contact by laying hens. This finding is in agreement with other preference methodologies (e.g. Dawkins 1983), also suggesting that varying the motivational state by applying prior resource restriction enhances Y-maze methodology, allowing interpretation of results in terms of both preference outcomes and impacts of resource utility. Results also indicated the value of including measurements of speed of movement during resource choice in providing information about preferences and relative preference strengths in addition to the choice outcomes. Further, the present study provided empirical confirmation of the importance of interpreting results of Y-maze tests within the context of the resource of comparison, consistent with intuitive expectations. In addition, the study provided some evidence for a stronger motivation for dust, if previously deprived of access, than of social contact, if previously deprived of visual and tactile contact, by laying hens.
It must be noted that to be confident that the outcomes of these preference tests reflect the welfare requirements of the animal, additional evidence, particularly on the occurrence of abnormal behaviour, stress physiology and health when restricted of the resource or the behaviour of interest, are necessary to provide a more comprehensive assessment of the impact of restricting a resource or behaviour on animal welfare (Widowski and Hemsworth 2008).
Acknowledgements
This research was conducted within the Poultry CRC, established and supported under the Australian Government’s Cooperative Research Centres Program.
References
Arnold NA, Ralph C, Petherick JC, Hemsworth PH (2007) The choice behaviour of laying hens for feed and dustbathing. In ‘Proceedings Australian Poultry Science Symposium Vol. 19’. (Ed. P Selle) p. 32. (University of Sydney: Camden)Barnard C (2004) ‘Animal behaviour: mechanism, development, function and evolution.’ (Pearson Education: Harlow, UK)
Bradshaw RH (1992) Conspecific discrimination and social preference in the laying hen. Applied Animal Behaviour Science 33, 69–75.
| Conspecific discrimination and social preference in the laying hen.Crossref | GoogleScholarGoogle Scholar |
Channing CE, Hughes BO, Walker AW (2001) Spatial distribution and behaviour of laying hens housed in an alternative system. Applied Animal Behaviour Science 72, 335–345.
| Spatial distribution and behaviour of laying hens housed in an alternative system.Crossref | GoogleScholarGoogle Scholar | 11348682PubMed |
Dawkins M (1976) Towards an objective method of assessing welfare in domestic fowl. Applied Animal Ethology 2, 245–254.
| Towards an objective method of assessing welfare in domestic fowl.Crossref | GoogleScholarGoogle Scholar |
Dawkins M (1977) Do hens suffer in battery cages? Environmental preferences and welfare. Animal Behaviour 25, 1034–1046.
| Do hens suffer in battery cages? Environmental preferences and welfare.Crossref | GoogleScholarGoogle Scholar |
Dawkins MS (1983) Battery hens name their price: consumer demand theory and the measurement of ethological ‘needs’. Animal Behaviour 31, 1195–1205.
| Battery hens name their price: consumer demand theory and the measurement of ethological ‘needs’.Crossref | GoogleScholarGoogle Scholar |
Duncan IJH (1977) Behavioural wisdom lost. Applied Animal Ethology 3, 193–194.
| Behavioural wisdom lost.Crossref | GoogleScholarGoogle Scholar |
Fraser D, Matthews LR (1997) Preference and motivation testing. In ‘Animal welfare’. (Eds MC Appleby, BO Hughes) pp. 159–174. (CAB International: Wallingford, UK)
Hughes BO (1977) Selection of group size in individual laying hens. British Poultry Science 18, 9–18.
| Selection of group size in individual laying hens.Crossref | GoogleScholarGoogle Scholar |
Hughes BO, Black AJ (1973) The preference of domestic hens for different types of battery cage floor. British Poultry Science 14, 615–619.
| The preference of domestic hens for different types of battery cage floor.Crossref | GoogleScholarGoogle Scholar |
Hutson GD (1981) Sheep movement on slatted floors. Australian Journal of Experimental Agriculture and Animal Husbandry 21, 474–479.
Laine S, Arnold NA, Hemsworth PH (2007) Choice behaviour of laying hens: effects of deprivation of feed and dustbath substrate. In ‘Proceedings Australian Poultry Science Symposium Vol. 19’. (Ed. P Selle) pp. 28–31. (University of Sydney: Camden)
Laine SM, Cronin GM, Hemsworth PH, Petherick JC (2008) The effects of quantity of reward on the choice behaviour of laying hens in a Y-maze preference test. In ‘Proceedings of the 42nd congress of the International Society of Applied Ethology, Dublin, Ireland, 5–9 August 2008’. (Eds L Boyle, N O’Connell, A Hanlon) p. 117. (Wageningen Academic Publishers: Wageningen, The Netherlands)
Larsen BH, Vestergaard KS, Hogan JA (2000) Development of dustbathing behavior sequences in the domestic fowl: the significance of functional experience. Developmental Psychobiology 37, 5–12.
| Development of dustbathing behavior sequences in the domestic fowl: the significance of functional experience.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD3M%2FjtFShsQ%3D%3D&md5=02931ffd9c061386beb2fb1e550239f7CAS | 10937656PubMed |
Matthews LR, Ladewig J (1994) Environmental requirements of pigs measured by behavioural demand functions. Animal Behaviour 47, 713–719.
| Environmental requirements of pigs measured by behavioural demand functions.Crossref | GoogleScholarGoogle Scholar |
Olsson IAS, Keeling LJ, Duncan IJH (2002) Why do hens sham dustbathe when they have litter? Applied Animal Behaviour Science 76, 53–64.
| Why do hens sham dustbathe when they have litter?Crossref | GoogleScholarGoogle Scholar |
Petherick JC, Waddington D, Duncan IJH (1990) Learning to gain access to a foraging and dustbathing substrate by domestic fowl: is ‘out of sight out of mind’? Behavioural Processes 22, 213–226.
Petherick JC, Sunderland RH, Waddington D, Rutter SM (1992) Measuring the motivation of domestic fowl in response to a positive and a negative reinforcer. Applied Animal Behaviour Science 33, 357–366.
| Measuring the motivation of domestic fowl in response to a positive and a negative reinforcer.Crossref | GoogleScholarGoogle Scholar |
Petherick JC, Seawright E, Waddington D (1993) Influence of motivational state on choice of food or a dustbathing/foraging substrate by domestic hens. Behavioural Processes 28, 209–220.
| Influence of motivational state on choice of food or a dustbathing/foraging substrate by domestic hens.Crossref | GoogleScholarGoogle Scholar |
Petherick JC, Seawright E, Waddington D, Duncan IJH, Murphy LB (1995) The role of perception in the causation of dustbathing behaviour in domestic fowl. Animal Behaviour 49, 1521–1530.
| The role of perception in the causation of dustbathing behaviour in domestic fowl.Crossref | GoogleScholarGoogle Scholar |
Phillips PA, Thompson BK, Fraser D (1988) Preference tests of ramp designs for young pigs. Canadian Journal of Animal Science 68, 41–48.
| Preference tests of ramp designs for young pigs.Crossref | GoogleScholarGoogle Scholar |
Pollard JC, Littlejohn RP, Suttie JM (1994) Responses of red deer to restraint in a Y maze preference test. Applied Animal Behaviour Science 39, 63–71.
| Responses of red deer to restraint in a Y maze preference test.Crossref | GoogleScholarGoogle Scholar |
Rushen J (1986) Aversion of sheep for handling treatments: paired-choice studies. Applied Animal Behaviour Science 16, 363–370.
| Aversion of sheep for handling treatments: paired-choice studies.Crossref | GoogleScholarGoogle Scholar |
van Rooijen J (1982) The value of choice tests in assessing welfare of domestic animals. Applied Animal Ethology 8, 295–299.
| The value of choice tests in assessing welfare of domestic animals.Crossref | GoogleScholarGoogle Scholar |
Vestergaard K (1982) Dust-bathing in the domestic fowl – diurnal rhythm and dust deprivation. Applied Animal Ethology 8, 487–495.
| Dust-bathing in the domestic fowl – diurnal rhythm and dust deprivation.Crossref | GoogleScholarGoogle Scholar |
Widowski TM, Hemsworth PH (2008) Housing hens to suit their ‘needs’. In ‘Proceedings XXIII World Poultry Congress, 30 June–4 July 2008, Brisbane’. p. 241.