

Climate change and broadacre livestock production across southern Australia. 3. Adaptation options via livestock genetic improvement
Andrew D. Moore A B and Afshin Ghahramani AA CSIRO Climate Adaptation National Research Flagship and Plant Industry, GPO Box 1600, Canberra, ACT 2601, Australia.
B Corresponding author. Email: Andrew.Moore@csiro.au
Animal Production Science 54(2) 111-124 https://doi.org/10.1071/AN13052
Submitted: 31 January 2013 Accepted: 19 May 2013 Published: 20 August 2013
Abstract
Climate change is predicted to reduce the productivity of the broadacre livestock industries across southern Australia; to date there has been no formal evaluation of the potential of genetic improvement in cattle or sheep to ameliorate the impacts of changing climates. We used the GRAZPLAN simulation models to assess selection of five traits of sheep and cattle as adaptation options under the SRES A2 global change scenario. Analysis of the breeding strategies was carried out for 25 representative locations, five livestock enterprises and three future years (2030, 2050, 2070). Uncertainty in future climates was taken into account by considering projected climates from four global circulation models. For three sheep enterprises, breeding for greater fleece growth (at constant body size) was predicted to produce the greatest improvements in forage conversion efficiency, and so it was the most effective genetic adaptation option. For beef cow and steer enterprises, breeding for larger body size was most effective; for beef cows, however, this conclusion relied on per-animal costs (including provision of bulls) remaining stable as body size increases. Increased conception rates proved to be less effective but potentially viable as an adaptation in beef cow and crossbred ewe enterprises. In the southern Australian environments that were analysed, our modelling suggests that breeding for tolerance to heat stress is unlikely to improve the performance of livestock production systems even at 2070. Genetic improvement of livestock was able to recover much less of the impact of climate change on profitability at drier locations where the need for adaptation is likely to be greatest. Combinations of feedbase and livestock genetic adaptations are likely to complement one another as the former alter the amount of forage that can be consumed, while the latter affect the efficiency with which consumed forage is converted to animal products. Climate change impacts on pasture production across southern Australia are likely to have only small effects on methane emissions intensity, as are a range of candidate genetic and feedbase adaptations to climate change; methane emissions per hectare in future climates will therefore be driven mainly by changes in livestock numbers due to alterations in pasture productivity.
Additional keywords: breeding, grazing systems, modelling.
Introduction
The broadacre sheep and beef cattle industries in the high-rainfall and cereal–livestock zones of southern Australia together account for ~20% of the nation’s gross value of agricultural production (Australian Bureau of Statistics 2012). Anthropogenic emissions of greenhouse gases, along with other human-induced changes to the earth system, are expected to cause substantial changes in the climate of the farms that produce these livestock in coming decades; for example, projections reported by CSIRO (2007a) for 2050 indicate a warming of ~0.8–2.8°C and decreases in rainfall of 0–20%.
In the first paper of this series (Moore and Ghahramani 2013) we used simulation modelling to show that these climate changes will adversely impact pasture growth and livestock production across most of southern Australia. For the SRES A2 scenario of global change (IPCC 2000) and in the absence of adaptations other than adjusting stocking rates to avoid excessive erosion risk, we estimated that total aboveground net primary productivity would decrease by an average of 9% in 2030, 7% in 2050 and 14% in 2070 from historical levels. We found, however, that the sustainable rates of pasture utilisation would decrease disproportionately, so that projected decreases in operating profit averaged 27% in 2030, 32% in 2050 and 48% in 2070. The proportional reductions in both pasture growth and profitability were greater at locations near the dry margin of the cereal–livestock zone. A second paper (Ghahramani and Moore 2013) explored a range of adaptation options that aimed to alter the amount or pattern of forage supply, or to increase the proportion of forage production that could be safely utilised; we found that increasing soil fertility and (where feasible) the incorporation of lucerne into the feedbase were the most effective adaptations, and that the effectiveness of adaptations tended to be lower in the drier parts of the cereal–livestock zone.
Genetic improvement of sheep to suit Australian conditions has been under way since the early days of European settlement, but until the 1970s breeding of southern Australian beef cattle followed a British lead (Pattie 1973). The introduction of index-based selection methods in the 1980s and their subsequent improvement has led to accelerating genetic progress in participating beef cattle seedstock herds, especially for the Angus breed (Barwick and Henzell 2005) and for terminal sire sheep breeds (Swan et al. 2009). Rates of gain in Merino sheep in recent years have been slower (Swan et al. 2009). The extent to which these genetic improvements in stud animals are reflected in the wider herd and flock is difficult to ascertain from the literature.
Discussion of livestock genetics as an adaptation to climate change in Australia (Howden et al. 2008; Stokes et al. 2010) has focussed mainly on breeding for adaptation to heat stress, primarily for sheep and cattle in the pastoral zone. However, a wide range of genetic improvements – those that improve the efficiency with which livestock convert forage to animal product – are also potentially viable adaptations to the reductions in forage supply that can be expected under future climates. Modern approaches to livestock improvement that are based on quantitative genetics can deliver substantial gains in production per head in commercial flocks and herds (Johnston 2007; Swan et al. 2009).
To date there has been no formal evaluation of the potential of genetic improvement in cattle or sheep to ameliorate the impacts of changing climates, either in Australia or elsewhere. In this paper, therefore, we extend the climate change impacts analysis of Moore and Ghahramani (2013) to examine the potential of livestock genetic improvements to adapt sheep and beef cattle production systems across southern Australia to projected future climates. This analysis is a companion to that in Ghahramani and Moore (2013); in particular, we address the effects of single genetic adaptations in isolation, leaving combination of adaptation approaches to the final paper in the series.
Simulation modelling is the only viable way to integrate the multiple, and spatially variable, effects of changing climate on pasture and animal production with the physiological changes and tradeoffs implied by alternative breeding objectives. We have therefore carried out this climate change adaptation study with the GRAZPLAN simulation models of pasture and livestock systems (Freer et al. 1997; Moore et al. 1997).
Methods
The modelling study reported here extends work reported in companion papers (Ghahramani and Moore 2013; Moore and Ghahramani 2013) that have used similar methods; readers are referred to these papers for further detail.
The GRAZPLAN models
The GRAZPLAN simulation models of the dynamics of grazed temperate grasslands were used for this analysis. These models operate at a daily timestep and consist of a water balance model (Moore et al. 1997), a pasture growth model (Moore et al. 1997) and a model of the intake, nutrition, reproduction and mortality of ruminants (updated from that reported in Freer et al. 1997). These biophysical models are coupled to a flexible representation of the management of grazing systems (Moore et al. 1997). Moore and Ghahramani (2013) provide an account of the wide variety of physical and physiological responses to changing climates that are captured in the GRAZPLAN water balance and pasture model. The ruminant model is based on the Australian feeding standard (CSIRO 2007b). It includes direct effects of increased temperature via reductions in animal intakes on hot days, decreased energy expenditures by livestock in winter and lower peri-natal mortality of lambs (Freer et al. 1997); the last of these effects will also be modified by changes in rainfall during the period that lambs are born. Methane production in the model is predicted with an equation derived from that of Blaxter and Clapperton (1965), in which the proportion of gross energy intake lost as methane varies between 0.06 and 0.09 with the metabolisable energy (ME) content of the diet and the energy balance of livestock.
In common with many biological simulation models, the genetic characteristics of livestock are represented in the ruminant model by a set of 256 ‘genotypic parameters’. A hypothetical livestock genotype can be described by modifying these parameters, and the effect of introducing it into a given livestock production system can then be simulated. Tradeoffs in animal performance due to limited supplies of energy and protein will be accounted for within these simulations.
Variation of the effects of adaptations across space, time and livestock enterprises
The study area (henceforth ‘southern Australia’) is an area of 1.00 million km2 (Fig. 1); it includes 0.33 million km2 of grazed lands that support 84% of Australia’s sheep production and 36% of its beef cattle production by value (Australian Bureau of Statistics 2012). Climates and soil types vary widely; for example, annual rainfall at the locations modelled in this study varies from 299 to 1091 mm and mean annual temperature from 11.6 to 19.1°C (Table 1). Forage types are consequently very diverse (Pearson et al. 1997). Projected climate changes also vary across the study area (Christensen et al. 2007). To capture the likely geographic differences in the effectiveness of different adaptations, we have used a set of 25 representative locations (Fig. 1; Table 1) that were identified using methods described by Moore and Ghahramani (2013). At each of the 25 locations, a representative set of land resources (weather, soils and pastures) was described using the attributes required by the GRAZPLAN simulation models (Moore and Ghahramani 2013; Table 1).
|

|
Climate projections from the CMIP3 global climate models were used in this study. In order to take account of the uncertainty in projected climates, climate projections from four global circulation models (GCMs) were considered for each future year: CCSM3 (Collins et al. 2006), ECHAM5/MPI-OM (Roeckner et al. 2003), GFDL-CM2.1 (Delworth et al. 2006) and UKMO-HadGEM1 (Johns et al. 2006). GCMs were selected on the basis of their overall skill over the Australian continent (CSIRO 2007a) and the availability of the monthly-scale projected weather data required for downscaling. A single future emissions scenario was used; the high-emissions SRES A2 scenario was selected because it is consistent with recent history (Peters et al. 2012). A reference period of historical weather data (1970–99) was simulated (with an atmospheric CO2 concentration of 350 ppm), plus projected future climates for the years 2030, 2050 and 2070 with CO2 concentrations of 451, 532 and 635 ppm, respectively (Houghton et al. 2001).
Historical daily weather data sequences for each location were taken from the Patched Point dataset (Jeffrey et al. 2001). Weather data for each projected climate were constructed using a downscaling technique modified from that of Zhang (2007; for details of the modifications see Moore and Ghahramani 2013). This algorithm was selected because it accounts for changes in precipitation (in particular) at three temporal scales that are important in pasture–livestock systems: inter-annual, between seasons and also the short-term patterns of wet and dry days.
Livestock production systems were specified for each of five enterprises: Merino ewes producing both fine wool and lambs for meat, Merino × Border Leicester cross ewes with an emphasis on lamb production, Angus cows producing yearling or weaner steers and heifers, Merino wethers for fine wool production, and Angus steers. Within each enterprise, the same livestock genotypes were assumed across all locations (Table 3) in order to facilitate comparisons across sites. The same genotypic parameters were used for the Merino ewe and Merino wether enterprises and for the Angus cow and Angus steer enterprises. Representative management policies (livestock replacement, the timing of the reproductive cycle, the sale of young stock and thresholds for supplementary feeding) were described separately for each enterprise × location combination. Details of the livestock management systems can be found in Tables S1 and S2, available as Supplementary Material to this paper.
Adaptation options
As used in this paper, ‘forage conversion efficiency’ is the ratio of gross income from meat and wool to mass of pasture dry matter consumed, expressed in $/kg. In terms of the water-use efficiency framework of Moore et al. (2011), it is computed as.

where f9 denotes the conversion efficiency of pasture to a livestock product and f10 denotes the price of the product.
Several possible adaptations through improved livestock genetics were designed and examined, on the premise that using animals with higher conversion efficiency would mitigate the consequences of a lower amount of consumable pasture growth. There are numerous possible routes to more feed-efficient livestock: the ones modelled here were chosen because they were nominated by livestock producers in a series of workshops (Pattinson 2011).
Higher body size
Maintenance energy requirements of ruminants vary with the 3/4 power of bodyweight, while their maximum rate of intake increases roughly linearly (CSIRO 2007b). Larger animals should therefore use a smaller proportion of the energy in consumed forage for maintenance, leaving more energy for growth, wool production or reproduction. There has been a long-standing trend of increasing livestock body size in Australia; Bell and Moore (2012) noted that the weight of ewes sold for mutton in Australia has increased by 9% in the last 30 years. The body size of an animal breed is represented in the GRAZPLAN ruminant model by a parameter called the ‘standard reference weight’, i.e. the weight of a mature, empty female in average body condition.
Higher fleece weight
There is considerable variation in potential wool growth rates, both between and within sheep breeds (e.g. Atkins 1980). Animals that devote a higher proportion of their energy and protein intake to wool growth can be expected to generate more profit per kilogram of forage eaten, as long as the resources devoted to extra wool growth do not compromise survival or reproduction. Breeding programs that aim to increase fleece weights typically achieve this aim in part through an increase in the body size of livestock; here, however, we consider an increase in fleece weight that is obtained at a constant body size. In the GRAZPLAN ruminant model, this increase is modelled as a change in the ratio of a parameter known as the ‘reference fleece weight’ to the standard reference weight.
Higher conception rates
In ewe and beef cow enterprises, a higher reproduction rate will result in a greater energetic efficiency in the flock or herd as a whole, because it increases the proportion of young, growing livestock. In southern Australian livestock systems, the great majority of ewes and cows conceive each year and survival from conception to birth and of newborn calves is also high; the greatest scope for genetic improvement in overall reproduction rates therefore lies in fecundity and in post-natal survival in sheep. We have focussed on fecundity as an adaptation option, as it applies to both sheep and cattle enterprises. In both sheep and cattle, most of any conception rate increase will have to appear as an increase in the proportion of multiple births; this carries a risk of increased peri-natal mortalities of offspring (Donnelly 1984). In the GRAZPLAN ruminant model, conception rate per oestrus cycle is controlled by a set of nine genotypic parameters that describe the effect of time of year and of relative body size for single, twin and triplet conceptions (Freer et al. 1997).
Higher body size of sires
This adaptation option is limited to livestock enterprises where terminal sires are used (here, only the crossbred ewe enterprise). By using a breed of rams with larger body size, the offspring will be larger, thereby increasing the conversion efficiency both of individual young stock and of the production system as a whole. The usefulness of this management option is limited by the risk of dystokia; if fetuses grow too large relative to their mothers, birth becomes difficult and the death rate of lambs or calves increases (Woolliams et al. 1983). The effect of sire body size is represented in the GRAZPLAN ruminant model by assuming that the genotypic parameters describing offspring can be obtained by averaging those in the maternal and paternal genotypes.
Improved tolerance to heat stress
At high temperatures, livestock can reduce their food intake (Weston 2002) and sustained high temperatures during mating can reduce fertility (Amundson et al. 2006). Modelling of the heat balance of livestock (e.g. Howden and Turnpenny 1997) is made difficult by a lack of knowledge of behavioural adaptations, in particular shifts in the timing of grazing within the day. In the GRAZPLAN ruminant model, potential intake of livestock is reduced linearly once maximum temperature exceeds a threshold, but only when minimum temperatures are also high (Freer et al. 1997).
One genetic adaptation that we did not consider was breeding for fleeces with lower fibre diameter. Finer wool receives higher prices, and so breeding for it is a viable strategy for individual producers. Here, however, we consider adaptations that receive widespread adoption, and if the Merino industry as a whole moves to finer wool, then over the decades the price premium is likely to dissipate.
Modelling the adaptations
A key difference between genetic adaptations and the feedbase and livestock management adaptations considered in earlier papers in this series is that the former need to be implemented slowly over time. In other words, we needed to consider gradual improvement of genetics over time, using rates of progress that should be achievable given our knowledge of rates of genetic improvement during the last 20–30 years. We assumed linear rates of genetic gain over time and used selection index theory (Falconer 1981) to place the projected rates of genetic progress for the adaptations (other than heat stress tolerance) on a common basis. Relevant genetic variances and correlations for four traits (adult bodyweight, greasy fleece weight, fibre diameter and conception rate, i.e. offspring born per mother mated; Table 2) were sourced from the literature for the two sheep breeds and for Angus cattle. Fibre diameter was included as we assumed that it would be maintained over time. Selection indices were constructed for higher body size (i.e. equal proportional changes in weight and fleece weight, zero change in fibre diameter and fertility), higher fleece weight (change in fleece weight only) and higher conception rate (change in conception rate only) and rates of genetic gain per unit of selection intensity were calculated from the genetic parameters. A common selection intensity was then chosen that gave a relative rate of increase in body size of Merino sheep of 0.5% in the first year; the corresponding rates of genetic gain calculated for the other traits and breeds are given in Table 3.

|

|
Little or no information is available about the genetic control of heat stress in sheep or cattle, and so the scope for genetic improvement through heat stress was evaluated by modelling alternative genotypes with zero effect of heat stress on intake. Effects of increased temperature on reproduction were neglected, as mating times in the representative management systems generally avoided this risk; temperature-humidity indices during mating reached sustained levels likely to induce stress (Amundson et al. 2006; Marai et al. 2007) only for the sheep enterprises at Dalwallinu at 2050 and 2070.
Simulation experiment
As for our analysis of feedbase adaptation options (Ghahramani and Moore 2013), a factorial simulation experiment was conducted in which the factors were climate scenario (1 + 4 × 3 levels), location (25), livestock enterprise (5), and genetic adaptation option (5). Historical-climate simulations were run over the years 1970–99, while projected-climate simulations were run over 30 years of climate realisations for the nominated future year. For each combination of the factors, a range of stocking rates was modelled and physical and financial outputs from the grazing system were stored from each simulation run. In particular, modelled long-term average methane emissions on a per-hectare basis were recorded for each simulation, as well as for the simulations of feedbase adaptation options reported in Ghahramani and Moore (2013). A long-term rate of operating profit was calculated as the gross margin less overhead costs, an operator allowance and a further adjustment for the capital cost of the livestock required at each stocking rate with a 7% interest rate (Moore and Ghahramani 2013). Costs and prices were computed from average values over the years 2007–11. A fixed operator allowance of $60 000 was assumed for all locations and enterprises, but it was spread over varying farm areas. Because we were analysing scenarios in which adoption of improved genetics was widespread across the industry, we assumed that market competition between studs would prevent relative increases in the price of rams and bulls.
An ‘optimal sustainable stocking rate’ (OSSR) was selected for each location × enterprise × climate × adaptation combination as that which gave highest profit while keeping the frequency of low ground cover (defined as cover <0.70) below location-specific thresholds (Table 1). All results are reported at the OSSR for a given climate, location, enterprise and adaptation scenario.
In order to compare adaptation strategies across locations and enterprises with different financial outcomes, we computed the ‘relative effectiveness’ of an adaptation as the proportion of the negative impact of climate change that it recovers:

where PA denotes long-term average operating profit under a projected climate when an adaptation has been implemented, PN is operating profit under that climate without any adaptation, and PH is operating profit during the historical period. The relative effectiveness of a given increase in profitability will, by definition, be larger at locations where the impact of climate change on profitability is smaller; in circumstances where PN ≥ PH, the relative effectiveness cannot be calculated. System-level efficiency measures from the framework of Moore et al. (2011) were also computed from the biophysical and economic simulation results, in order to gain insight into ways that genetic improvements resulted in more economically viable production systems.
Results
Profitability changes due to the genetic adaptations were much more consistent between GCMs than for the feedbase adaptations reported by Ghahramani and Moore (2013). All results are therefore reported as averages over the GCMs.
Industry-level effectiveness of genetic adaptations for each livestock enterprise
Table 4 shows the changes over time in the relative effectiveness of the different genetic adaptations in restoring profitability of each livestock enterprise, when averaged over the study area. At this industry scale, greater fleece growth was clearly the most effective option for the three sheep enterprises, while selection for larger body size was most effective for the cattle enterprises. Larger body size was of intermediate relative effectiveness in the three sheep enterprises, as was higher conception rate in the crossbred ewe and beef cow enterprises. Apart from tolerance to heat stress, which was not modelled as an incremental improvement, the relative effectiveness of all the genetic adaptations increased from 2030 to 2050 and then declined from 2050 to 2070.

|
For some specific adaptation × enterprise combinations, (greater fleece growth for wethers and larger body size for beef cows and steers), the modelled rates of genetic progress were sufficient to restore average operating profit per hectare to or beyond its historical level. Because most of the adaptations were applicable to only a subset of the enterprises, however, the overall relative effectiveness of each adaptation in restoring the profitability of broadacre livestock production was diluted: the highest industry-level relative effectiveness was 0.59 for greater fleece growth at 2050.
The modelled relative effectiveness of complete tolerance of high temperatures as an adaptation was very small for all enterprises and future years: the highest effectiveness found for this adaptation option (0.02 for steers at 2030) was below the lowest value for all the other adaptations (0.06 for higher conception rate in beef cows at 2030). Adaptation to heat stress is therefore not considered further in these results.
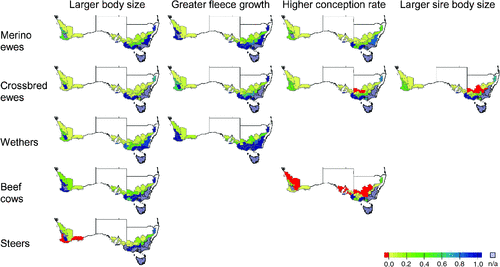
Regional variations in the effectiveness of genetic adaptations
The spatial patterns of the relative effectiveness of each genetic adaptation are shown in Fig. 1 for 2050, the future year at which the effectiveness of genetic adaptation was greatest. It can be seen that the average figures in Table 4 mask significant spatial variation. For example, there are seven regions where higher conception in beef cattle is counter-productive. While larger body size is a highly effective adaptation overall for steers, it has a negative relative effectiveness in 2 of the 25 regions and a relative effectiveness less than 0.2 in another 9 regions. In every case, the adaptations had greater relative effectiveness in the high-rainfall zone, especially in the south-eastern part of the mainland. For Launceston, the Tasmanian location, projected climates increased the profitability of the livestock enterprises and so relative effectiveness could not be calculated.
This gradient in relative effectiveness from wet to dry regions is caused to a large extent by smaller relative impacts on profitability in the higher-rainfall regions (Moore and Ghahramani 2013). In these regions, a smaller proportional increase in profitability is required to recover the effects of changing climate. In particular, the high average relative effectiveness values for the cattle enterprises in Table 4 are caused by the concentration of cattle production in those parts of the study area where the impact of climate change is smaller.
Table 5 shows that when the genetic adaptations are compared within each location × enterprise combination, greater fleece growth is the most effective option for the sheep enterprises and larger body size is the most effective option for the cattle enterprises in nearly every case. For the steer enterprise, this was necessarily the case as only one adaptation was modelled. Table 5 also confirms that even when the locally-best genetic adaptation option is selected, livestock genetic improvement has low potential effectiveness at low-rainfall locations, and that most of the industry-level adaptation effect shown in Table 4 arises from profit increases at wetter locations that have a smaller requirement for adaptation to climate change.

|
The genetic adaptations resulted in only minor changes in stocking rates (when expressed as dry sheep equivalents/ha) and in the total amounts of forage consumed. Fig. 2 shows that the different genetic adaptations did, however, alter the forage conversion efficiencies of the livestock production systems. Breeding for greater fleece growth had the largest overall effects on conversion efficiency, with the gains in conversion efficiency being slightly greater at low-rainfall locations in the ewe enterprises. In the Merino ewe and wether enterprises, the increases in forage conversion efficiency from breeding for greater fleece growth were much greater for the other adaptations. For the crossbred ewe enterprise, however, especially at lower-rainfall locations, the differences between the conversion efficiency gains from breeding for greater fleece growth and breeding for higher conception rate were much smaller and so the most-effective adaptations for this enterprise might alter if the ratio of sheep meat price to wool price were to increase above its 2007–11 level. Breeding for higher conception rate was estimated to increase conversion efficiency of the beef cattle enterprises by a greater amount than breeding for larger body size, even though the changes in profit due to higher conception rate were smaller. In keeping with their relative effectiveness (Table 4), higher conception rate produced greater increases in conversion efficiency in the crossbred ewe enterprise than in the Merino ewe enterprise. Larger body sizes increased forage conversion efficiency of steer enterprises by as much as 20%, but with a good deal of idiosyncratic variability between locations. In the other enterprises – including beef cattle – breeding for larger body size only had small effects on conversion efficiency, generally with larger increases at wetter locations.
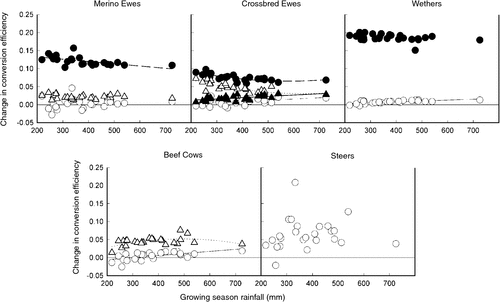
|
 ), and higher sire body size (▲). Conversion efficiencies are computed as dollars of gross income per kg of forage DM consumed and are averages over four global circulation models. Linear or polynomial regressions on growing season (April–October) rainfall are shown where they are significant.
), and higher sire body size (▲). Conversion efficiencies are computed as dollars of gross income per kg of forage DM consumed and are averages over four global circulation models. Linear or polynomial regressions on growing season (April–October) rainfall are shown where they are significant.Changes in methane emissions under projected climates and adaptations
As can be seen from Table 6, modelled methane emissions per hectare fall significantly over the study area under projected future climates, but this is driven almost entirely by reductions in OSSR (Moore and Ghahramani 2013); emissions intensity changes relatively little except in the steer enterprise, where it declines by 4% at 2070. The additional changes in methane emissions per hectare from adapting via livestock genetic improvement are small, but emissions per dollar of income decline in response to these adaptations, especially to breeding for greater fleece growth (which produces the greatest increase in conversion efficiency). For three of the feedbase options examined by Ghahramani and Moore (2013), methane emissions respond differently: adding lucerne to the feedbase, higher soil fertility and the use of confinement feeding all increase OSSR and hence methane emissions per hectare while having little effect on methane emissions intensity. Removing annual legumes from pastures, which is generally an unprofitable option, also increases methane emitted per dollar earned.

|
Discussion
Long-term feasibility of the genetic adaptations
One of the key assumptions in this modelling analysis is that the rates of genetic gain given in Table 3 can be sustained until 2070. Several the projected phenotypes fall within the range observed in existing strains of the modelled breeds: Merino ewes can be found with a standard reference weight of 65 kg (Safari et al. 2007a) or a reference conception rate above 140% (Fogarty 2009), as can meat sheep with a reference conception rate of 170% and Angus cows with a mature weight of 700 kg (Costa et al. 2011). While a 38% increase in fleece weight seems large, half that gain was achieved by index selection over only 10 years in the QPLU$ flock at Trangie (Mortimer et al. 2006). This ‘what-if’ analysis does, however, assume widespread and (for 2030) near-immediate adoption of the relevant breeding goal.
Potential usefulness of the genetic adaptations
For many of the location × enterprise combinations there is at least one genetic improvement strategy that will recover much, if not all, of the decline in operating profit. For the projected climates at 2050, adoption of the most-profitable options shown in Table 5 would recover all of the impact on profitability of 36% of livestock enterprises by value, and half or more of the impact on another 12% of livestock enterprises. For about a quarter of livestock enterprises, however, the best adaptation through livestock genetics has a relative effectiveness below 0.2 at 2050. These enterprises are concentrated in the drier regions of the study area, where the quantity of consumable forage decreases substantially under projected future climates (Moore and Ghahramani 2013); in these environments, the increases in forage conversion efficiency required to compensate for the reduced forage availability are simply unobtainable.
For sheep, breeding for greater fleece growth was the most effective of the genetic adaptation strategies, because energy devoted to wool production returns a greater economic value than energy devoted to the growth of sheep meat. As a result, conversion efficiency of forage to income by the sheep production systems was significantly increased by breeding to redirect resources to the fleece at all locations, even for crossbred ewes (Fig. 2). The small increases in energy demand imposed by extra wool growth (0.05–0.2 MJ ME/head.day) did not significantly compromise meat production.
At the production system level, increases in biophysical efficiency from larger animal body sizes were small for all enterprises except steers (6% or less to 2070, averaging 1% at 2050). The gains in operating profit reflected in Table 4 and Fig. 1 were largely produced by lower costs, since costs of rams and bulls, shearing and animal husbandry (but not purchased stock or sale costs) were assumed to remain constant on a per-head basis. At 2050, these cost efficiencies amounted on average to an 8% reduction in cost per hectare for beef cows and crossbred ewes, 6% for wethers and 3% for Merino ewes. The greater conversion efficiency increases found for the steer enterprises presumably arose from all animals in this enterprise being young, growing stock. To the extent that husbandry and shearing costs per head increase with body size, therefore, the relative effectiveness of this genetic change will decline.
The relative effectiveness of breeding for larger body size was higher for the cattle enterprises than for the sheep enterprises (Table 4). This difference was not caused by large differentials in the benefit from higher conversion efficiency (Fig. 2) or savings from having fewer animals to manage, but from the different geographical locations of the industries. Within the study area, cattle production is disproportionately located in the high-rainfall and eastern regions where climate change impact (the denominator of Eqn 2) is smaller, and so a smaller level of relative operating profit increase is required to return profitability to historical levels.
Breeding for increased conception rates was dominated by another genetic option for all the breeding enterprises and locations, but for the enterprises based on meat production (crossbred ewes and beef cattle) it provided a second-best approach. For the crossbred ewe enterprises, breeding for increased conception rates was unique in producing substantially greater increases at low-rainfall locations than at high-rainfall locations (Fig. 2). This response was not, however, large enough to outweigh the larger impact of climate change in lower-rainfall regions, and so the general spatial pattern of relative effectiveness remained (Fig. 1).
The results of the modelling analysis for heat tolerance should be regarded as somewhat preliminary, given the simple nature of the heat stress response functions in the GRAZPLAN ruminant model. Nevertheless, the negligible responses in Table 4 strongly suggest that livestock heat stress is a minor issue in southern Australia compared with those impacts of climate change on broadacre livestock production that arise through pasture growth, as long as shade is present (as implicit in the GRAZPLAN equations). Part of the reason for this is that the hottest locations in the study area (e.g. Dalwallinu) are also dry in summer. Not only does low relative humidity allow animals to thermoregulate more efficiently through evaporative heat loss, but animals’ grazing durations during summer should be relatively short in Mediterranean environments, permitting them to shift their grazing period to the night hours (Silanikove 2000). For some southern Australian locations adjacent to the study area that have summer-dominant rainfall (e.g. the north-western plains of New South Wales), heat stress may have a significant impact.
Differences and similarities compared with feedbase-oriented adaptations
The first and most important difference between the genetic adaptations reported here and the adaptations via the feedbase analysed in a companion paper (Table 5 vs Ghahramani and Moore 2013) is that the genetic adaptations had little or no effect on aboveground net primary productivity and so resulted in only small changes in OSSR. The feedbase adaptations, on the other hand, were useful to the extent that they allowed the OSSR to increase. Because of these different modes of action – which are also reflected in the methane emissions results shown in Table 6 – it is likely that feedbase and genetic adaptations can be combined with little or no dilution of effectiveness.
The main advantage of the feedbase adaptations (especially higher soil fertility) over the genetic adaptations was that they can be employed effectively across all the livestock enterprises considered in our analysis, whereas the genetic adaptations were applicable to only a subset of enterprises (e.g. greater fleece growth) or, in the case of larger body size, had relatively low effectiveness for some enterprises. The main advantage of the genetic adaptations over the feedbase adaptations was that the benefits from the former can be expected to increase over time; as a result, the relative effectiveness of the genetic adaptations in 2070 was comparable to, and in some cases greater than, relative effectiveness in 2030 (Table 4). The effectiveness of the feedbase adaptations, on the other hand, declined over time (table 3 of Ghahramani and Moore 2013).
The most important similarity between the feedbase and genetic adaptations lies in the spatial pattern of their effectiveness (Fig. 1; fig. 3 of Ghahramani and Moore 2013). Despite some of the adaptations giving proportionally larger effects in drier environments (Fig. 2), the greater impact of climate change on OSSR and hence profitability in drier environments means that much larger proportional improvements are needed from adaptation strategies. For a location such as Narrandera, where the amount of safely consumable herbage and hence OSSR is estimated to decline by nearly 60% at 2050, an adaptation strategy needs to more than double economic water-use efficiency to be fully effective, and none of the options examined in our analyses approaches this level.
Methane emissions from broadacre livestock systems under projected climates
From Table 6, it is clear that changes in methane emissions per hectare under projected future climates will be driven by changes in livestock numbers much more than by methane emissions per dry sheep equivalent. Even though the projected future climates result in substantial changes in the amount and pattern of pasture production (Moore and Ghahramani 2013), and also in its species composition (Ghahramani and Moore 2013), they do not alter animals’ energy intake or patterns of weight change sufficiently to produce large shifts in methane emissions per kilogram of forage intake. The genetic adaptations that are the focus of this paper change the total consumption of forage very little, and are therefore also predicted to have little effect on total methane emissions over and above that induced by climate change impacts.
The feedbase-oriented adaptations to climate change analysed by Ghahramani and Moore (2013), including the introduction of substantial areas of summer-growing forages and increases in soil fertility, still leave the intra-annual pattern (rather than amount) of energy available to livestock sufficiently similar to that under historical climate that methane emissions intensity is largely unaffected (Table 6). If widely adopted, the successful feedbase adaptations are predicted to result in increases in livestock numbers that would return total methane emissions towards their historical levels without much affecting emissions intensity.
Alcock and Hegarty (2011), also using the GRAZPLAN models for a single location and historical climate, found that a range of management adaptations could improve emissions intensity by 10–20%. Many of these management changes, however, induced shifts in the average ME content of the feedbase, e.g. through changed mating dates or supplementary feeding. In keeping with our results, they found that changing to a genotype with larger body size had little effect on emissions intensity, but for increases in conception rates they estimated a larger improvement in emissions intensity than found here for 2050 climates at our nearest location (Cootamundra).
Both experimental studies (Newton et al. 1994; Cantarel et al. 2013) and our modelling results (Ghahramani and Moore 2013) indicate that the legume content of pastures is likely to increase under climate change as long as soil phosphorus fertility is maintained. A recent laboratory study (Purcell et al. 2012) indicates that methane emissions per unit of ME are lower from legume forages than from grasses; if these results translate to the field and to Australian conditions, then the predictions of our analysis with respect to methane emissions intensity will require revision.
Conclusions
Our analysis suggests the most effective genetic-based adaptation options could, if fully adopted, result in recovery of most of industry profitability at 2030 and nearly all of it at 2050 as improvements in conversion efficiency and cost-effectiveness accumulate. The benefits of genetic gain will be spread unevenly across the study area, however, with producers in drier environments being forced to apply genetic improvement to fewer animals. Nonetheless, genetic improvement is a good example of a ‘no-regret’ adaptation to climate change that is likely to be economically beneficial under present-day climate.
The financial calculations in this analysis have used constant prices since future price and cost levels are highly uncertain. In the past, however, gains from livestock breeding have largely been used to compensate for declines in livestock producers’ terms of trade. If the long-term cost-price squeeze continues and is overlaid by the climate change impacts estimated in our analysis, then the incremental adaptations we have examined may be insufficient to maintain the viability of many livestock enterprises.
As noted above, the feedbase-oriented adaptations examined by Ghahramani and Moore (2013) and the genetic adaptations examined affect the water-use efficiency of livestock systems at different points, and so are likely to be effective in tandem. The analysis of combinations of adaptations will be the subject of a final paper in this series.
Acknowledgements
This research was supported by funding from the Australian Government Department of Agriculture, Fisheries, and Forestry under its Climate Change Research Program; Meat & Livestock Australia; Dairy Australia; and Australian Wool Innovation. We are indebted to our colleagues in the Southern Livestock Adaptation 2030 research and development program (Phil Graham, Melissa Rebbeck, Martin Dunstan, Mike Hyder, Richard Eckard, Peter Ball, Doug Alcock, Phil Bowden, Geoff Casburn, Jan Edwards, Ruth Corrigan, Jane Court, Clare Edwards, Nathan Ferguson, Jeff House, Colin Langford, Greg Meaker, Jim Meckiff, Kieran Ransom and Julia Smith) who have shared representative grazing system information with us, and to Russell Pattinson for his coordination of the program.
References
Alcock DJ, Hegarty RS (2011) Potential effects of animal management and genetic improvement on enteric methane emissions, emissions intensity and productivity of sheep enterprises at Cowra, Australia. Animal Feed Science and Technology 166–167, 749–760.| Potential effects of animal management and genetic improvement on enteric methane emissions, emissions intensity and productivity of sheep enterprises at Cowra, Australia.Crossref | GoogleScholarGoogle Scholar |
Amundson JL, Mader TL, Rasby RJ, Hu QS (2006) Environmental effects on pregnancy rate in beef cattle. Journal of Animal Science 84, 3415–3420.
| Environmental effects on pregnancy rate in beef cattle.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28Xht1CnurbE&md5=5ca90781d6dfdf32b029f1f44b2f5de2CAS | 17093236PubMed |
Atkins KD (1980) The comparative productivity of five ewe breeds. 2. Hogget wool production. Australian Journal of Experimental Agriculture and Animal Husbandry 20, 280–287.
| The comparative productivity of five ewe breeds. 2. Hogget wool production.Crossref | GoogleScholarGoogle Scholar |
Australian Bureau of Statistics (2012) 7121.0 – agricultural commodities, Australia, 2010–11. Available at www.abs.gov.au/AUSSTATS/abs@.nsf/Lookup/7121.0Main+Features12010-11 [Verified 24 May 2013]
Barwick SA, Henzell AL (2005) Development successes and issues for the future in deriving and applying selection indices for beef breeding. Australian Journal of Experimental Agriculture 45, 923–933.
| Development successes and issues for the future in deriving and applying selection indices for beef breeding.Crossref | GoogleScholarGoogle Scholar |
Bell LW, Moore AD (2012) Integrated crop–livestock systems in Australian agriculture: trends, drivers and implications. Agricultural Systems 111, 1–12.
| Integrated crop–livestock systems in Australian agriculture: trends, drivers and implications.Crossref | GoogleScholarGoogle Scholar |
Blaxter KL, Clapperton JL (1965) Prediction of the amount of methane produced by ruminants. The British Journal of Nutrition 19, 511–522.
| Prediction of the amount of methane produced by ruminants.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaF28XitFKktg%3D%3D&md5=ec3a76e9bbdd683050d21c878857d138CAS | 5852118PubMed |
Cantarel AAM, Bloor JMG, Soussana JF (2013) Four years of simulated climate change reduces aboveground productivity and alters functional diversity in a grassland ecosystem. Journal of Vegetation Science 24, 113–126.
| Four years of simulated climate change reduces aboveground productivity and alters functional diversity in a grassland ecosystem.Crossref | GoogleScholarGoogle Scholar |
Christensen JH, Hewitson B, Busuioc A, Chen A, Gao X, Held I, Jones R, Kolli RK, Kwon WT, Laprise R, Magaña Rueda V, Mearns L, Menéndez CG, Räisänen J, Rinke A, Sarr A, Whetton P (2007) Regional climate projections. In ‘Climate change 2007: the physical science basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change’. (Eds S Solomon, D Qin, M Manning, Z Chen, M Marquis, KB Averyt, M Tignor, HL Miller) pp. 847–940. (Cambridge University Press: Cambridge)
Collins WD, Bitz CM, Blackmon ML, Bonan GB, Bretherton CS, Carton JA, Chang P, Doney SC, Hack JJ, Henderson TB, Kiehl JT, Large WG, McKenna DS, Santer BD, Smith RD (2006) The Community Climate System Model version 3 (CCSM3). Journal of Climate 19, 2122–2143.
| The Community Climate System Model version 3 (CCSM3).Crossref | GoogleScholarGoogle Scholar |
Costa RB, Misztal I, Elzo MA, Bertrand JK, Silva LOC, Lukaszewicz M (2011) Estimation of genetic parameters for mature weight in Angus cattle. Journal of Animal Science 89, 2680–2686.
| Estimation of genetic parameters for mature weight in Angus cattle.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhtFWqtrjM&md5=795b5248839041c90ced23a0ee411302CAS | 21478454PubMed |
CSIRO (2007a) ‘Climate change in Australia – Technical Report 2007.’ (CSIRO Marine and Atmospheric Research: Aspendale, Vic.) Available at http://www.climatechangeinaustralia.gov.au/technical_report.php [Verified 24 May 2013]
CSIRO (2007b) ‘Nutrient requirements of domesticated ruminants.’ (CSIRO Publishing: Melbourne)
Delworth TL, Broccoli AJ, Rosati A, Stouffer RJ, Balaji V, Beesley JA, Cooke WF, Dixon KW, Dunne J, Dunne KA, Durachta JW, Findell KL, Ginoux P, Gnanadesikan A, Gordon CT, Griffies SM, Gudgel R, Harrison MJ, Held IM, Hemler RS, Horowitz LW, Klein SA, Knutson TR, Kushner PJ, Langenhorst AR, Lee HC, Lin SJ, Lu J, Malyshev SL, Milly PCD, Ramaswamy V, Russell J, Schwarzkopf MD, Shevliakova E, Sirutis JJ, Spelman MJ, Stern WF, Winton M, Wittenberg AT, Wyman B, Zeng F, Zhang R (2006) GFDL’s CM2 global coupled climate models. Part I: formulation and simulation characteristics. Journal of Climate 19, 643–674.
| GFDL’s CM2 global coupled climate models. Part I: formulation and simulation characteristics.Crossref | GoogleScholarGoogle Scholar |
Donnelly JR (1984) The productivity of breeding ewes grazing on lucerne or grass and clover pastures on the tablelands of southern Australia. III Lamb mortality and weaning percentage. Australian Journal of Agricultural Research 35, 709–721.
| The productivity of breeding ewes grazing on lucerne or grass and clover pastures on the tablelands of southern Australia. III Lamb mortality and weaning percentage.Crossref | GoogleScholarGoogle Scholar |
Falconer DS (1981) ‘Introduction to quantitative genetics.’ 2nd edn. (Longmans Green: London/New York)
Fogarty NM (2009) A review of the effects of the Booroola gene (FecB) on sheep production. Small Ruminant Research 85, 75–84.
| A review of the effects of the Booroola gene (FecB) on sheep production.Crossref | GoogleScholarGoogle Scholar |
Freer M, Moore AD, Donnelly JR (1997) GRAZPLAN: decision support systems for Australian grazing enterprises. II. The animal biology model for feed intake, production and reproduction and the GrazFeed DSS. Agricultural Systems 54, 77–126.
| GRAZPLAN: decision support systems for Australian grazing enterprises. II. The animal biology model for feed intake, production and reproduction and the GrazFeed DSS.Crossref | GoogleScholarGoogle Scholar |
Ghahramani A, Moore AD (2013) Climate change and broadacre livestock production across southern Australia. 2. Adaptation by grassland management. Crop and Pasture Science 64, 615–630.
| Climate change and broadacre livestock production across southern Australia. 2. Adaptation by grassland management.Crossref | GoogleScholarGoogle Scholar |
Gregory KE, Bennett GL, VanVleck LD, Echternkamp SE, Cundiff LV (1997) Genetic and environmental parameters for ovulation rate, twinning rate, and weight traits in a cattle population selected for twinning. Journal of Animal Science 75, 1213–1222.
Houghton JT, Ding Y, Griggs DJ, Noguer M, van der Linden PJ, Xiaosu D (Eds) (2001) ‘Climate change 2001. The scientific basis. Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change (IPCC).’ (Cambridge University Press: Cambridge, UK)
Howden SM, Turnpenny J (1997) Modelling heat stress and water loss of beef cattle in subtropical Queensland under current climates and climate change. In ‘MODSIM ‘97 international congress on modelling and simulation proceedings, 8–11 December, University of Tasmania, Hobart’. (Eds DA McDonald, M McAleer) pp. 1103–1108. (Modelling and Simulation Society of Australia: Canberra)
Howden SM, Crimp SJ, Stokes CJ (2008) Climate change and Australian livestock systems: impacts, research and policy issues. Australian Journal of Experimental Agriculture 48, 780–788.
| Climate change and Australian livestock systems: impacts, research and policy issues.Crossref | GoogleScholarGoogle Scholar |
IPCC (2000) ‘Emissions scenarios. Special Report of the Intergovernmental Panel on Climate Change.’ (Eds N Nakicenovic, R Swart) (Cambridge University Press: Cambridge, UK)
Jeffrey SJ, Carter JO, Moodie KM, Beswick AR (2001) Using spatial interpolation to construct a comprehensive archive of Australian climate data. Environmental Modelling & Software 16, 309–330.
| Using spatial interpolation to construct a comprehensive archive of Australian climate data.Crossref | GoogleScholarGoogle Scholar |
Jeyaruban MG, Johnston DJ, Graser H-U (2009) Estimation of genotype × environment interactions for growth, fatness and reproductive traits in Australian Angus cattle. Animal Production Science 49, 1–8.
| Estimation of genotype × environment interactions for growth, fatness and reproductive traits in Australian Angus cattle.Crossref | GoogleScholarGoogle Scholar |
Johns TC, Durman CF, Banks HT, Roberts MJ, McLaren AJ, Ridley JK, Senior CA, Williams KD, Jones A, Rickard GJ, Cusack S, Ingram WJ, Crucifix M, Sexton DMH, Joshi MM, Dong BW, Spencer H, Hill RSR, Gregory JM, Keen AB, Pardaens AK, Lowe JA, Bodas-Salcedo A, Stark S, Searl Y (2006) The new Hadley Centre Climate Model (HadGEM1): evaluation of coupled simulations. Journal of Climate 19, 1327–1353.
| The new Hadley Centre Climate Model (HadGEM1): evaluation of coupled simulations.Crossref | GoogleScholarGoogle Scholar |
Johnston DJ (2007) Genetic trends in Australian beef cattle – making real progress. Proceedings of the Association for the Advancement of Animal Breeding and Genetics 17, 8–15.
Marai IFM, El-Darawany AA, Fadiel A, Abdel-Hafez MAM (2007) Physiological traits as affected by heat stress in sheep – a review. Small Ruminant Research 71, 1–12.
| Physiological traits as affected by heat stress in sheep – a review.Crossref | GoogleScholarGoogle Scholar |
Moore AD, Ghahramani A (2013) Climate change and broadacre livestock production across southern Australia. 1. Impacts of climate change on pasture and livestock productivity, and on sustainable levels of profitability. Global Change Biology 19, 1440–1455.
| Climate change and broadacre livestock production across southern Australia. 1. Impacts of climate change on pasture and livestock productivity, and on sustainable levels of profitability.Crossref | GoogleScholarGoogle Scholar | 23504950PubMed |
Moore AD, Donnelly JR, Freer M (1997) GRAZPLAN: decision support systems for Australian grazing enterprises. III. Pasture growth and soil moisture submodels, and the GrassGro DSS. Agricultural Systems 55, 535–582.
| GRAZPLAN: decision support systems for Australian grazing enterprises. III. Pasture growth and soil moisture submodels, and the GrassGro DSS.Crossref | GoogleScholarGoogle Scholar |
Moore AD, Robertson MJ, Routley R (2011) Evaluation of the water use efficiency of alternative farm practices at a range of spatial and temporal scales: a conceptual framework and a modelling approach. Agricultural Systems 104, 162–174.
| Evaluation of the water use efficiency of alternative farm practices at a range of spatial and temporal scales: a conceptual framework and a modelling approach.Crossref | GoogleScholarGoogle Scholar |
Mortimer S, Taylor P, Atkins K (2006) The Trangie QPLU$ selection lines: responses in clean fleece weight and fibre diameter on completion of ten rounds of selection. In ‘Proceedings of the Trangie QPLUS Open Day, Trangie Agricultural Research Centre. Trangie, Australia, 11 May 2006’. (Ed. CE Pope) pp. 7–11. (NSW Department of Primary Industries: Orange) Available at Available at: www.dpi.nsw.gov.au/__data/assets/pdf_file/0020/153416/trangie-qplus-merinos-open-day-proceedings-2006.pdf [verified 3 June 2013]
Newton PCD, Clark H, Bell CC, Glasgow EM, Campbell BD (1994) Effects of elevated CO2 and simulated seasonal changes in temperature on the species composition and growth rates of pasture turves. Annals of Botany 73, 53–59.
| Effects of elevated CO2 and simulated seasonal changes in temperature on the species composition and growth rates of pasture turves.Crossref | GoogleScholarGoogle Scholar |
Pattie WA (1973) Breeding and genetics. In ‘The pastoral industries of Australia’. (Eds G Alexander, OB Williams) pp. 303–335. (Sydney University Press: Sydney)
Pattinson R (2011) Can southern Australian livestock producers adapt to a changing climate? In ‘Proceedings of the 52nd annual conference of the Grassland Society of Southern Australia’. (Ed. J Hirth) pp. 129–132. (Grassland Society of Southern Australia: Tooborac, Vic.)
Pearson CJ, Brown R, Collins WJ, Archer KA, Wood MS, Petersen C, Bootle B (1997) An Australian temperate pastures database. Australian Journal of Agricultural Research 48, 453–465.
| An Australian temperate pastures database.Crossref | GoogleScholarGoogle Scholar |
Peters GP, Marland G, Le Quere C, Boden TA, Canadell JG, Raupach MR (2012) Rapid growth in CO2 emissions after the 2008–2009 Global Financial Crisis. Nature Climate Change 2, 2–4.
| Rapid growth in CO2 emissions after the 2008–2009 Global Financial Crisis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhs1Gjs7vJ&md5=6f987b2ca31c052301c65feb75b27d28CAS |
Purcell PJ, Grant J, Boland TM, Grogan D, O’Kiely P (2012) The in vitro rumen methane output of perennial grass species and white clover varieties, and associative effects for their binary mixtures, evaluated using a batch-culture technique. Animal Production Science 52, 1077–1088.
| The in vitro rumen methane output of perennial grass species and white clover varieties, and associative effects for their binary mixtures, evaluated using a batch-culture technique.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xhs1Wju73N&md5=391053d6d141cb676b41e70666231feaCAS |
Roeckner E, Bäuml G, Bonaventura L, Brokopf R, Esch M, Giorgetta M, Hagemann S, Kirchner I, Kornblueh L, Manzini E, Rhodin A, Schlese U, Schulzweida U, Tompkins A (2003) ‘The atmospheric general circulation model ECHAM5. Part I: Model description.’ Max Planck Institute for Meteorology Report 349. (Max Planck Institute for Meteorology: Hamburg)
Safari E, Fogarty NM, Gilmour AR (2005) A review of genetic parameter estimates for wool, growth, meat and reproduction traits in sheep. Livestock Production Science 92, 271–289.
| A review of genetic parameter estimates for wool, growth, meat and reproduction traits in sheep.Crossref | GoogleScholarGoogle Scholar |
Safari E, Fogarty NM, Gilmour AR, Atkins KD, Mortimer SI, Swan AA, Brien FD, Greeff JC, van der Werf JHJ (2007a) Across population genetic parameters for wool, growth, and reproduction traits in Australian Merino sheep. 2. Estimates of heritability and variance components. Australian Journal of Agricultural Research 58, 177–184.
| Across population genetic parameters for wool, growth, and reproduction traits in Australian Merino sheep. 2. Estimates of heritability and variance components.Crossref | GoogleScholarGoogle Scholar |
Safari E, Fogarty NM, Gilmour AR, Atkins KD, Mortimer SI, Swan AA, Brien FD, Greeff JC, van der Werf JHJ (2007b) Genetic correlations among and between wool, growth and reproduction traits in Merino sheep. Journal of Animal Breeding and Genetics 124, 65–72.
| Genetic correlations among and between wool, growth and reproduction traits in Merino sheep.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2s3nvVCrsw%3D%3D&md5=774736b0d7f8eb948e6fc042b968602cCAS | 17488356PubMed |
Silanikove N (2000) Effects of heat stress on the welfare of extensively managed domestic ruminants. Livestock Production Science 67, 1–18.
| Effects of heat stress on the welfare of extensively managed domestic ruminants.Crossref | GoogleScholarGoogle Scholar |
Stokes CJ, Crimp SJ, Gifford RM, Ash AJ, Howden SM (2010) Broadacre grazing. In ‘Adapting agriculture to climate change: preparing Australian agriculture, forestry and fisheries for the future’. (Eds CJ Stokes, SM Howden) pp. 171–185. (CSIRO Publishing: Melbourne)
Swan AA, Brown DJ, Banks RG (2009) Genetic progress in the Australian sheep industry. Proceedings of the Association for the Advancement of Animal Breeding and Genetics 18, 326–329.
Weston RH (2002) Constraints on feed intake by grazing sheep. In ‘Sheep nutrition’. (Eds M Freer, H Dove) pp. 27–49. (CSIRO Publishing: Melbourne)
Woolliams C, Wiener G, Macleod NSM (1983) The effects of breed, breeding system and other factors on lamb mortality: 2. Factors influencing the incidence of delayed birth, dystokia, congenital defects and miscellaneous causes of early death. Journal of Agricultural Science (Cambridge) 100, 553–561.
| The effects of breed, breeding system and other factors on lamb mortality: 2. Factors influencing the incidence of delayed birth, dystokia, congenital defects and miscellaneous causes of early death.Crossref | GoogleScholarGoogle Scholar |
Zhang XC (2007) A comparison of explicit and implicit spatial downscaling of GCM output for soil erosion and crop production assessments. Climatic Change 84, 337–363.
| A comparison of explicit and implicit spatial downscaling of GCM output for soil erosion and crop production assessments.Crossref | GoogleScholarGoogle Scholar |