

Grain feeding increases core body temperature of beef cattle
R. H. Jacob A F , V. S. M. Surridge B , D. T. Beatty C D , G. E. Gardner D and R. D. Warner EA Department of Agriculture and Food WA, Baron Hay Court, South Perth, WA 6151, Australia.
B Department of Agriculture and Food WA, Albany Highway, Albany, WA 6330, Australia.
C Meat and Livestock Australia, Building East 1, Office No. E313, Dubai Airport Freezone, Dubai 491, United Arab Emirates.
D School of Veterinary and Biomedical Sciences, Murdoch University, South Street, Murdoch, WA 6150, Australia.
E CSIRO Animal, Food and Health Sciences, Werribee, Vic. 3030, Australia.
F Corresponding author. Email: robin.jacob@agric.wa.gov.au
Animal Production Science 54(4) 444-449 https://doi.org/10.1071/AN13463
Submitted: 6 November 2013 Accepted: 10 February 2014 Published: 6 March 2014
Journal Compilation © CSIRO Publishing 2014 Open Access CC BY-NC-ND
Abstract
The core body temperature and post slaughter loin temperatures of steers fed on grass pasture was compared with those of steers fed a grain-based feedlot diet. The feeding treatments were grass for 300 days (Grass), grass for 150 days then feedlot for 150 days (Short Feedlot) and feedlot for 300 days (Long Feedlot). Temperature telemeters were inserted under the peritoneum of the steers and temperature measured at intervals of 1 h for the 300 days, and then at intervals of 1 min for the 48-h period before slaughter. The pH and temperature decline post mortem was also measured. The carcasses of the feedlot steers were heavier and fatter than those from the Grass-fed steers. The core body temperature of the steers from the feedlot treatments was 0.3–0.4°C higher than for the Grass treatment at the time of slaughter. The loin temperature was higher in the feedlot treatments than the Grass treatment at all times measured post mortem as was the temperature at pH 6. Feedlotting can increase the likelihood of ‘high rigor temperature’ conditions of high temperature and low pH occurring in beef carcasses, due to an increase in core body temperature before slaughter, a decrease in the rate of cooling and an increase in the rate of pH decline post mortem. These effects are possibly due to a combination of a direct effect of feed type on body temperature as well as indirect effects on bodyweight and condition score.
Introduction
High rigor temperature is characterised by muscle temperature and pH conditions being greater than 35°C and less than 6, respectively, and occurs in many beef carcasses in Australia (Hopkins et al. 2007; Warner et al. 2014a). Reduced water-holding capacity (Offer and Knight 1988), reduced tenderisation (Warner et al. 2014b), pale colour (Swatland 2004; Hughes et al. 2014), premature browning during retail display (Ledward 1985) and sarcomere shortening (Honikel et al. 1983) have all been associated with high rigor temperature. High rigor temperature occurs more frequently when cattle are finished on grain for an extended period of time compared with grass pasture (Warner et al. 2014a), although the reason for this is unclear and could be due to differences in either the rate of cooling or the rate of pH decline, both of which are interdependent as reviewed by Jacob and Hopkins (2014).
Many cattle in Australia are fed a grain-based diet during the finishing period before slaughter; the length of which can vary according to market requirements and pasture conditions on-farm. Grass-finished steers with carcass weights of 300–420 kg and 7–22 mm P8 fat (Anonymous 2000) are slaughtered for lower-value Asian markets. Steers destined for Asian (Japan and Korea) supermarkets are grain fed for 120–150 days and slaughtered at carcass weights of 280–400 kg. Steers grain fed for 150– 350 days and slaughtered at carcass weights of 350–450 kg are destined for Asian restaurants and supermarkets, depending on their marble scores (Andrews and Littler 2007).
The rate of carcass cooling could be influenced by body temperature at the time of slaughter as this represents the starting point from which the temperature declines post mortem. The core body temperature of an animal refers to the inner body temperature excluding skin and subcutaneous adipose tissue and is a critical aspect of an animal’s thermal equilibrium (Lefcourt and Adams 1996). Cattle under temperate climates that are well hydrated exhibit a normal core body temperature range from 36.7 to 39.1°C (Cunningham 2002). Throughout the day the body temperature will fluctuate, called rhythmic fluctuation, with body temperature reaching its peak at dusk and drops to the lowest body temperature in the morning (Lefcourt and Adams 1996, 1998).
Cattle fed high energy diets are more susceptible to heat stress (Gaughan et al. 1997) and stresses such as transport and handling cause an increase in the core body temperature of cattle that may impact on meat quality (Ferguson et al. 2001). Feedlotting is known to increase the risk of heat stress in cattle due to several reasons such as feed type (Robinson and Lee 1947), feed toxins, and cattle type. Rectal temperature along with pulse rate and respiration rate of heifers consuming an 80 : 20 concentrate to roughage ration were observed to increase significantly in comparison to heifers on an all roughage diet (Brosh et al. 1998).
Feeds that have the lowest heat increment are fats and oils, with roughages and forages having the highest heat increments. Different types of carbohydrates have different heat increments with cellulose having a higher heat increment compared with starch and sugar due to greater heat production with acetate compared with propionate metabolism (Armstrong and Blaxter 1956; MacRae and Lobley 1982). Large cattle have a reduced surface area to weight ratio hence a relatively smaller surface area from which to disperse heat from, than smaller cattle (Sjaastad et al. 2003). Therefore, due to animal size and feed composition, it seems likely that feedlot-fed cattle will have higher core temperatures at slaughter compared with grass-fed cattle.
In Australia cattle are slaughtered after a curfew, transport and lairage period that can last for 24–48 h (McLennan 2005), during which time they may have access to water but not feed. Any effects of feed type or metabolic differences due to the finishing period may therefore have diminished by the time of slaughter. This study aimed to compare core body temperatures from cattle finished on grass pasture and grain feeding (feedlot) systems, to determine whether finishing cattle with a grain-based feedlot system might subsequently predispose beef carcasses to high temperature and low pH conditions.
Materials and methods
Animals and treatments
This experiment was conducted on a commercial beef property in the Great Southern region of Western Australia. Eighteen cattle were used in the experiment; all of which were Bos taurus steers, Angus or Angus cross, and ~13 months of age. At the commencement of the experiment the mean liveweight of the cattle was 369 ± 14 kg (mean ± s.e.m.). They were allocated randomly to one of three treatment groups, containing six animals each. The three treatment groups were; Grass, Short Feedlot and Long Feedlot and are described in Table 1.
|
The treatments spanned 11 calendar months beginning on 12 February 2008 and finished when the steers were slaughtered 300 days later on 9 December 2008. The Short Feedlot steers were introduced to the feedlot on 11 July 2008 after 150 days of Grass feeding. In effect, there was an overlap between treatments such that from 12 February 2008 to 11 July 2008 (period 1) the Grass and Short Feedlot treatments had the same feed type, and from 11 July 2008 until 7 December 2008 (period 2) Short Feedlot and Long Feedlot had the same feed type.
The feedlot ration (Table 2) was fed ad libitum after an introduction period of 10 days designed to avoid acidosis. For feedlot treatments, the experimental steers were mixed with 94 similar steers so that the pen contained 100 steers in total, at a stocking density of one steer per 16 m2. The pasture was an annual grass pasture estimated to contain 9 MJ/kg of metabolisable energy and 12% crude protein.

|
Core body temperature measurement
To measure core body temperature, two temperature telemeters (iButton DS1922 L, Maxim Integrated, San Jose, CA, USA) were implanted into the abdominal cavity under the peritoneum of each steer, 8 days before the start of the experiment. One temperature logger was programmed to record once every 60 min for the duration of the feeding program of 300 days (Tlive). The other was programmed to record at intervals of 1 min, to commence before the on-farm curfew, transportation, and lairage periods for a total of 2 days (Tcurfew).
The telemeters were surgically attached to the abdominal wall, under the peritoneum using the method of Beatty et al. (2006). The steers were transported to the Murdoch University Veterinary Hospital for the surgery. Preparation of the temperature telemeters before implantation consisted of wrapping in cellular tape then plumbers tape to bind them together, followed by dipping in an inert wax at least 3–4 times, allowing the wax to dry in between each layer. Lastly, ~60 cm of non-absorbable suture (Vetafil, Bengen, Germany) was tied around the middle of waxed bundle, to allow suturing on to the muscle adjacent to the peritoneal cavity.
The steers were fasted overnight and weighed before surgery. Surgery was completed with the animal positioned standing in a veterinary crush. An epidural nerve block provided sedation while a lumbar paravertebral nerve block provided anaesthesia via separate injections of 0.04 mg/kg of xylazine (Ilium/Troy Laboratories, Glendenning, NSW, Australia) made up to 2 mL with 2% plain lignocaine (Ilium/Troy Laboratories). A 20-cm incision was made into the peritoneal cavity in the area of the right paralumbar fossa, where the muscle layers were blunt dissected to the peritoneum. The wax-coated packages containing the telemeters were soaked in chlorhexidine solution for 24 h and then cleaned with distilled water before being stitched onto the peritoneal wall muscle layer. To close the peritoneum and muscle layers of non-absorbable, synthetic suture material (Vetafil, 0.40 mm diameter) was employed. An anti-inflammatory injection of 1.1–2.2 mg/kg of Flunixin (Ilium/Troy Laboratories) and a broad spectrum antibiotic 5–10 mg/kg of oxytet-200 LA (Ilium/Troy Laboratories, containing oxytetracycline 200 mg/mL) was administered at the time of surgery. After surgery, the steers were left to recover and monitored closely for 1 week on the Murdoch University farm then were trucked back to the commercial property. The skin sutures were removed 14 days after surgery.
Slaughter
At the end of the 300-day feeding period, the steers from all treatments were transported to a commercial abattoir, located ~300 km from the farm and slaughtered as a single consignment. During lairage the steers had access to water but not food, such that the total time without feed before slaughter was 48 h. Slaughter consisted of stunning with a captive bolt followed by exsanguination. Temperature telemeters were retrieved from the carcass after evisceration. Loin temperature and pH were measured by a Meat Standards Australia accredited grader at hourly intervals for 5 h post slaughter and again at the time of grading 20 h post slaughter, as described by Warner et al. (2014a). Carcass and fat measurements were taken from abattoir records.
Statistical analyses
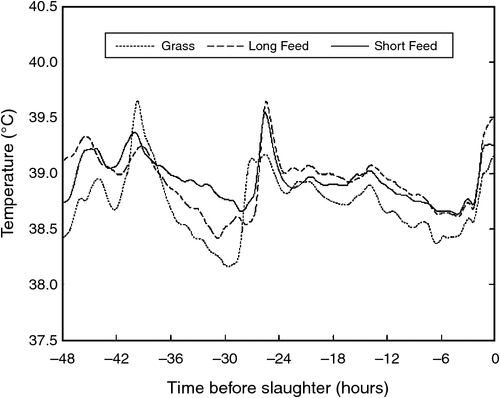
Genstat 15th Edition (32 bit, VSN International Ltd, Hemel Hempstead, UK) was used for all statistical analyses. A linear mixed model was used to test the effect of treatment on core body temperature during the on-farm period (Tlive). The model included treatment (Grass, Short Feedlot and Long Feedlot) and hour of day (1–24) as fixed effects. The random term was month/day.animal. A one-way ANOVA was used to test the difference between treatments for mean core body temperature for the 48-h period before slaughter (Tcurfew) that included on-farm curfew, transport and lairage at the abattoir. Separation of means was determined by calculation of least significant difference (P < 0.05). Carcass grading, ante-mortem and post-mortem data was analysed using one-way ANOVA with no blocking. The plots presented in Fig. 3 are of smoothed data, derived from raw data using the moving averages trendline option in Microsoft Excel, with the period set to 60.
Results
During both treatment periods on-farm there was a significant effect (P < 0.01) of treatment, hour of day and a significant interaction (P < 0.01) between treatment and hour of day for Tlive. A diurnal pattern was evident for Tlive, with the daily minimum being significantly (P < 0.05) lower than the daily maximum temperature for all treatments and treatment periods (Figs 1, 2). For treatment period 1 (12 February 2008–11 July 2008) there was no difference between Grass and Short Feedlot steers for Tlive (Fig. 1). There was a significant difference between Long Feedlot and the other two treatments for the minimum (P < 0.05) but not the maximum Tlive (P > 0.05). The minimum Tlive occurred at 0600 hours for Grass and Short Feedlot treatments and at 0800 hours for the Long Feedlot treatment. The maximum value for Tlive occurred at 1800 hours for all treatments. For treatment period 2 (11 July 2008–9 December 2008), the minimum and maximum values for Tlive treatment were lower for the Grass compared with both the Short Feedlot and Long Feedlot treatments (Fig. 2). The maximum value for Tlive was lower for the Short Feedlot treatment compared with the Long Feedlot treatment. However, there was no difference between Short Feedlot and Long Feedlot treatments for the minimum value of Tlive. The minimum value for Tlive occurred at 0600 hours for the Grass treatment and at 0800 hours for both the Short Feedlot and Long Feedlot treatments. The maximum value for Tlive occurred at 1800 hours for Grass and Short Feedlot treatments and at 1900 hours for the Long Feedlot treatment.
|
|
The mean temperature during the ante-mortem period was lower for the Grass treatment compared with both the Short Feedlot and Long Feedlot treatments (Table 3) and this is clearly demonstrated by the plot of moving averages for Tcurfew (Fig. 3). The mean slaughter time was 1110 hours, 1316 hours and 1130 hours for Grass, Short Feedlot and Long Feedlot steers, respectively. There was a significant (P < 0.05) effect of treatment on hot carcass weight and fat score (Table 3). The carcass weight of the steers in the Long Feedlot treatment was higher than for those in the Short Feedlot treatment that were higher than for those in the Grass treatment. Fat score was the same for steers in the Short Feedlot and Long Feedlot treatments and higher for those in the Grass treatment (P < 0.05).
|
|
During the post-mortem period the loin temperature of the Grass-fed steers was lower (P < 0.01) than Short Feedlot and Long Feedlot steers at 1, 2, 3, 4, 5 and 20 h and was not different between Short and Long Feedlot steers at any time (P > 0.05) post slaughter. The loin pH was higher at 4 and 5 h post slaughter for Short Feedlot, Long Feedlot and Grass, respectively, and no different (P > 0.05) at the other time points.
The temperature at pH 6 was lower for the Grass treatment compared with the Short and Long Feedlot treatments (P < 0.01, Table 4), and no difference between Short and Long Feedlot treatments.

|
Discussion
Effect of treatment on core body temperature
Carcasses from feedlot steers were heavier, fatter and had a higher temperature at the time of slaughter than carcasses from Grass-fed steers. Subsequently they cooled at a slower rate during the post-mortem period. Higher muscle temperature post slaughter in the carcasses of grain-fed cattle has previously been attributed to heavier, fatter carcasses and consequent slower temperature decline post slaughter (Warner et al. 2014a). However, in this case, the higher temperature in grain-fed cattle/carcasses was observed during the finishing period, the ante-mortem period, at slaughter, as well as post slaughter. So the effect of feeding treatment was to increase the temperature of the animals when alive as well as to decrease the rate of cooling post mortem. However, the difference in loin temperature between Short Feedlot and Long Feedlot treatments was gone by 2 h post mortem and there was no difference between these treatments for rate of pH decline (Table 4). So it might be argued that the effect of treatment on live animal temperature was less important than the effect on rate of cooling post mortem in relation to rigor temperature.
The effect of feedlotting on the temperature of the live animal appeared to depend on the length of the grain feeding period. This is supported by the relative differences in temperature between treatments being different for treatment periods 1 and 2. In treatment period 1, the feeding regime for Grass and Short Feedlot treatments was effectively the same and there was no difference in core body temperature between these treatments, as could be expected (Fig. 1). The Long Feedlot steers had a higher temperature than Grass and Short Feedlot steers but this occurred for the diurnal minimum only and not the diurnal maximum temperature. The diurnal minimum temperature also occurred later in the day for the Long Feedlot steers. The reason for this is unknown but may have been associated with different feeding behaviours between the treatments. During treatment period 2, the temperature for the Long Feedlot treatment was clearly higher than for the Grass treatment for both minimum and maximum diurnal treatments. While the Short Feedlot treatment was higher than the Grass treatment for maximum diurnal temperature, this value was also lower than for the Long Feedlot treatment. So in summary the difference between grass and grain feeding appeared to begin as an increase in the minimum diurnal temperature and then extended to an increase in maximum diurnal temperature as well.
This difference in time could point to more than one mechanism being influenced by treatment. A possible explanation is the higher energy content of the grain-based ration compared with pasture increased minimum diurnal temperature. As the feeding period continued an effect of bodyweight influenced the ability of animals to lose heat hence both maximum and minimal diurnal temperatures were affected.
The diurnal fluctuation of temperature for cattle is 0.5–1.2°C depending on ambient temperature (Hahn 1999). The heat load of cattle is highest during the day when heat gained via solar radiation and metabolism regularly surpasses heat loss from radiation, convection and evaporation. Consequently heat will be stored in the body and the core body temperature will increase. At night the cooler temperatures reverse the heat flow and the stored heat is dispersed back to the environment resulting in reduction of the core body temperature (Finch 1986). A difference in the minimum diurnal temperature as observed in treatment period 1, suggests the treatment is exerting an effect when a net loss of heat is occurring. Alternatively a difference in maximum diurnal temperature as observed in treatment period 2, suggests that the treatment is exerting an effect when a net gain in heat is occurring.
Robinson and Lee (1947) illustrated that a high plane of nutrition results in highly significant increases in rectal temperature, pulse rate and respiratory rate. Rectal temperature along with pulse rate and respiration rate of heifers consuming an 80 : 20 concentrate to roughage ration were observed to increase significantly in comparison to heifers on an all roughage diet (Brosh et al. 1998). In a study performed by Mader et al. (2002) on feeding strategies for managing heat load in feedlot cattle, their conclusion illustrated that ‘limit-feeding feedlot cattle during summer was a successful tool for enhancing animal comfort by alleviating the combined effects of high climatic and metabolic heat load’.
Large cattle have a reduced surface area to weight ratio hence a relatively smaller surface area from which to disperse heat, than smaller cattle (Sjaastad et al. 2003). As the treatment period increased, the difference between treatments for liveweight would also have increased.
The difference between treatments observed at the time of slaughter might have been influenced by the time of slaughter as the Grass steers were slaughtered slightly earlier than Short Feedlot and Long Feedlot steers. The largest difference between mean slaughter times was 1 h 46 min between Grass and Short Feedlot steers, respectively. The diurnal temperature fluctuation seen on-farm was not evident during the curfew period. This was most likely due to fasting during this period. The apparent increase in temperature during the last 2 h before slaughter may have been due to exercise involved in moving animals from lairage into the abattoir. Also the increase in temperature ~24 h before slaughter coincided with the loading and transport period before lairage.
High rigor temperature conditions
High rigor temperature refers to conditions of high temperature and low pH occurring simultaneously during the early post-mortem period (Jacob and Hopkins 2014). The pH data (Table 4) suggests that the rate of post-mortem metabolism was higher in carcasses from the feedlot treatments compared with the Grass treatment. This in addition to the relatively higher temperatures resulted in a higher rigor temperature (temperature at pH 6) for the carcasses from the feedlot treatments. The temperature at pH 6 difference between treatments suggests the loin muscles of feedlot steers went into rigor at an earlier time post slaughter and were under ‘high rigor temperature’ conditions for a longer period compared with those from Grass-fed steers. Some authors suggest that the temperature at which high rigor temperature occurs may be as low as 30°C whereas others suggest it may be 38°C. If a temperature of 30°C and a pH of 6 are taken to be the threshold values for high rigor temperature, then the difference between treatments for the time that high rigor temperature conditions persisted, would likely have been in the order of several hours.
Conclusions
Feedlotting steers increases their risk of ‘high rigor temperature’ because of an increase in core body temperature, a reduction in rate of cooling and an increase in the rate of pH decline post slaughter. These effects are likely due to a combination of effects on body metabolism and size.
Acknowledgements
Special thanks to Athula Naththarampatha and Dr Dean Gutzke for technical assistance and advice. The authors are very grateful for financial assistance from Meat and Livestock Australia for this project.
References
Andrews T, Littler B (2007) Market specifications for beef cattle. Primefact 621. NSW Department of Primary Industry, Orange.Anonymous (2000) ‘Advanced carcase fat measurement, (ACFM), bovine P8, participants workbook.’ (AUS-MEAT Limited: North Murarrie, Qld). Available at http://www.ausmeat.com.au/media/1728/20150-acfm%20bovine%20workbook%202000.pdf [Verified 27 February 2014]
Armstrong DG, Blaxter KL (1956) Heat increments of feeding in ruminants. Nature 177, 1183–1184.
| Heat increments of feeding in ruminants.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaG28%2FoslWhtQ%3D%3D&md5=f734853a106c4bb5ba848803dad152a4CAS | 13334511PubMed |
Beatty DT, Barnes A, Taylor E, Pethick DW, McCarthy M, Maloney SK (2006) Physiological responses of Bos taurus and Bos indicus cattle to prolonged, continuous heat and humidity. Journal of Animal Science 84, 972–985.
Brosh A, Aharoni Y, Degan AA, Wright D, Young BA (1998) Effects of solar radiation, dietary energy, and time of feeding on thermoregulatory responses and energy balance in cattle in a hot environment. Journal of Animal Science 76, 2671–2677.
Cunningham J (2002) ‘Textbook of veterinary physiology.’ (W. B. Saunders Company: Philadelphia, PA)
Ferguson D, Bruce H, Thompson J, Egan A, Perry D, Shorthose W (2001) Factors affecting beef palatability – farmgate to chilled carcass. Australian Journal of Experimental Agriculture 41, 879–891.
| Factors affecting beef palatability – farmgate to chilled carcass.Crossref | GoogleScholarGoogle Scholar |
Finch VA (1986) Body temperature in beef cattle: its control and relevance to production in the tropic. Journal of Animal Science 62, 531–542.
Gaughan JB, Mader TL, Savage D, Young BA (1997) ‘Time of feeding influence on cattle exposed to heat.’ Nebraska Beef Cattle Reports. (University of Nebraska: Lincoln, NE)
Hahn GL (1999) Dynamic responses to cattle to thermal heat loads. Journal of Animal Science 77, 10–20.
Honikel KO, Roncales P, Hamm R (1983) The influence of temperature on shortening and rigor onset in beef muscle. Meat Science 8, 221–241.
| The influence of temperature on shortening and rigor onset in beef muscle.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL3sXhslOlu7k%3D&md5=083bb9d637249c1d6786ea2df6bc160fCAS | 22055561PubMed |
Hopkins DL, Cassar JA, Toohey ES, Wynn PC (2007) Examination of pH in lot fed beef for Japan. Proceedings of the New Zealand Society of Animal Production 67, 436–440.
Hughes JM, Kearney G, Warner RD (2014) Improving beef meat colour scores at carcass grading. Animal Production Science 54, 422–429.
| Improving beef meat colour scores at carcass grading.Crossref | GoogleScholarGoogle Scholar |
Jacob RH, Hopkins DL (2014) Techniques to reduce the temperature of beef muscle early in the post mortem period – a review. Animal Production Science 54, 482–493.
| Techniques to reduce the temperature of beef muscle early in the post mortem period – a review.Crossref | GoogleScholarGoogle Scholar |
Ledward DA (1985) Post-slaughter influences on the formation of metmyoglobin in beef muscles. Meat Science 15, 149–171.
| Post-slaughter influences on the formation of metmyoglobin in beef muscles.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL28XmslOjsw%3D%3D&md5=c5791c119f84ad5193c4463226eef27fCAS | 22054503PubMed |
Lefcourt AM, Adams WR (1996) Radiotelemetry measurement of body temperatures of feedlot steers during summer. Journal of Animal Science 74, 2633–2640.
Lefcourt AM, Adams WR (1998) Radiotelemetry measurement of body temperature in feedlot steers during winter. Journal of Animal Science 76, 1830–1837.
MacRae JC, Lobley GE (1982) Some factors which influence thermal energy losses during the metabolism of ruminants. Livestock Production Science 9, 447–456.
| Some factors which influence thermal energy losses during the metabolism of ruminants.Crossref | GoogleScholarGoogle Scholar |
Mader TL, Holt SM, Holt GL, Davis MS, Spiers DE (2002) Feeding strategies for managing heat load in feedlot cattle. Journal of Animal Science 80, 2373–2382.
McLennan L (2005) ‘Preslaughter hydration of Western Australian cattle and effects on beef quality, Beef and Sheep Updates.’ (Ed. V Reck) (Department of Agriculture Western Australia: Esperance)
Offer G, Knight P (1988) The structural basis of water holding in meat. In ‘Developments in meat science. Vol. 4’. (Ed. RA Lawrie) pp. 63–243. (Elsevier Applied Science: London, UK)
Robinson KW, Lee DHK (1947) The effects of the nutritional plane upon the reaction of animals to heat. Journal of Animal Science 6, 182–194.
Sjaastad ØV, Hove K, Sand O (2003) Regulation of body temperature. In ‘Physiology of domestic animals’. pp. 598–617. (Scandinavian Veterinary Press: Olso, Norway)
Swatland HJ (2004) Progress in understanding the paleness of meat with a low pH. South African Journal of Animal Science 34, 1–7.
| Progress in understanding the paleness of meat with a low pH.Crossref | GoogleScholarGoogle Scholar |
Warner RD, Dunshea FR, Gutzke D, Lau J, Kearney G (2014a) Factors influencing the incidence of high rigor temperature in beef carcasses in Australia. Animal Production Science 54, 363–374.
| Factors influencing the incidence of high rigor temperature in beef carcasses in Australia.Crossref | GoogleScholarGoogle Scholar |
Warner RD, Thompson JM, Polkinghorne R, Gutzke D, Kearney GA (2014b) A consumer sensory study of the influence of rigor temperature on eating quality and ageing potential of beef striploin and rump. Animal Production Science 54, 396–406.
| A consumer sensory study of the influence of rigor temperature on eating quality and ageing potential of beef striploin and rump.Crossref | GoogleScholarGoogle Scholar |