

Do vertebral chemical signatures distinguish juvenile blacktip shark (Carcharhinus limbatus) nursery regions in the northern Gulf of Mexico?
Justin P. Lewis A B E , William F. PattersonA University of South Alabama, Department of Marine Sciences, 307 North University Boulevard, Mobile, AL 366888, USA.
B Dauphin Island Sea Lab, 101 Bienville Boulevard, Dauphin Island, AL 36528, USA.
C National Marine Fisheries Service, Southeast Fisheries Science Center, Panama City Laboratory, 3500 Delwood Beach Road, Panama City, FL 32408, USA.
D Electro Scientific Industries, 685 Old Buffalo Trail, Bozeman, MT 59715, USA.
E Corresponding author. Present address: Center for Fisheries Research and Development, Gulf Coast Research Laboratory, The University of Southern Mississippi, 703 East Beach Drive, Ocean Springs, MS 39564, USA. Email: jplewis86@gmail.com
Marine and Freshwater Research 67(7) 1014-1022 https://doi.org/10.1071/MF15088
Submitted: 1 March 2015 Accepted: 1 December 2015 Published: 11 February 2016
Journal Compilation © CSIRO Publishing 2016 Open Access CC BY-NC-ND
Abstract
Identifying and protecting shark nurseries is a common management strategy used to help rebuild overfished stocks, yet we know little about connectivity between juvenile and adult populations. By analysing trace metals incorporated into vertebral cartilage, it may be possible to infer natal origin based on nursery-specific chemical signatures. To assess the efficacy of this approach, we collected juvenile blacktip sharks (Carcharhinus limbatus; n = 93) from four regions in the Gulf of Mexico in 2012 and 2013 and analysed their vertebral centra with laser ablation–inductively coupled plasma–mass spectrometry. We observed significant regional differences in six element : Ca ratios in both 2012 and 2013. Multi-element chemical signatures were significantly different among regions and between year-classes. Year-class-specific linear discriminant function analysis yielded regional classification accuracies of 81% for 2012 and 85% for 2013, although samples were not obtained from all four regions in 2012. Combining year-classes resulted in an overall classification accuracy of 84%, thus demonstrating the usefulness of this approach. These results are encouraging yet highlight a need for more research to better evaluate the efficacy of vertebral chemistry to study elasmobranch population connectivity.
Additional keywords: laser ablation, natal origin, shark nurseries, vertebral chemistry.
Introduction
Nurseries are areas that contribute a disproportionate number of juvenile recruits to adult populations, thus the conservation benefit of protecting nurseries is evident. However, inconsistent application of the term ‘nursery’ prompted the development of guidelines to empirically compare the value of different habitats or systems in the context of population maintenance and growth (Beck et al. 2001; Heupel et al. 2007). Proxies that indicate an area provides favourable conditions for subsequent recruitment to the adult population (e.g. high juvenile density, growth rate and survival) are important for understanding the ecological processes of source versus sink regions (for a review, see Beck et al. 2001). However, the ultimate character that distinguishes a functional nursery is a disproportionately high contribution of recruits to the adult population, thus determining where adults spent their early life is the ultimate test of whether a given habitat or system is, in fact, a nursery.
Identifying nursery areas based on estimates of connectivity to adult populations is particularly challenging for coastal shark species. Conventional mark–recapture studies are limited because recapture rates are typically low (e.g. ≤10%; Kohler and Turner 2001) and home ranges in the order of tens of square kilometres (e.g. Yeiser et al. 2008) make it difficult to mark enough individuals to compensate for high mortality during the first year of life. In the case of bony fishes, natural biogeochemical tags based on chemical constituents of calcified structures (e.g. otoliths, scales, fin rays and bone) provide an alternative approach to study movement patterns and population connectivity (for a review, see Elsdon et al. 2008). During the biomineralisation process, divalent cations, such as Sr2+ and Ba2+, substitute for Ca2+ relative to their environmental availability (Wells et al. 2000), whereas other elements may become incorporated interstitially or associated with the protein matrix (Campana 1999). Thus, the chemical constituents of calcified structures are representative of environmental conditions at the time of deposition. In bony fishes, otoliths are the preferred structure for chemical analysis of natural biogeochemical tags because they are inert once formed and their biomineralisation pattern produces alternating translucent and opaque zones that provide a chronometer that accompanies the recorded chemical histories (Campana 1999). Material at the core of the otolith represents deposition during the first year of life; thus, chemical signatures found in the core can act as a natural tag to be used to estimate the proportion of adults derived from different nurseries (e.g. Vasconcelos et al. 2011).
The incorporation of trace metal impurities into calcified structures of other marine organisms has also been shown to reflect the environmental conditions at the time of mineralisation (e.g. Pitts and Wallace 1994) and may be used to address a variety of ecological questions. Results from recent field (Scharer et al. 2012) and experimental (Smith et al. 2013) studies show some elements deposited in the hydroxyapatite matrix (Ca10(PO4)6(OH)2) of elasmobranch vertebrae are also influenced by abiotic factors (e.g. salinity, water chemistry and temperature). These results indicate that chemical signatures in vertebrae may serve as natural biogeochemical tags of elasmobranch juvenile habitat that could be used to examine sources of recruits to adult populations. However, their usefulness for studying connectivity between juvenile and adult populations is contingent on our ability to accurately classify individuals to their natal nurseries based on vertebral chemical signatures.
In the present study, blacktip sharks (Carcharhinus limbatus) were examined as a model coastal shark species to test whether vertebral chemical signatures distinguish nursery regions in the northern Gulf of Mexico (Gulf) and whether differences can be used to infer nursery origin. The blacktip shark is a cosmopolitan species that is found throughout coastal waters of the Gulf and is one of the more economically important shark species in the region (Southeast Data, Assessment, and Review 2012). Females reproduce biennially following a gestation period of ~12 months, with parturition peaking from March to May in coastal nurseries throughout the Gulf (Baremore and Passerotti 2013). Juvenile blacktips remain in natal coastal embayments up to the first 6 months of life before migrating to wintering grounds (Heupel et al. 2004; Hueter and Tyminski 2007). Tagging data indicate individuals up to the age of 3 years return to their nurseries during summer months (Heupel and Simpfendorfer 2002; Hueter et al. 2005), whereas analysis of mitochondrial (mt)DNA and nuclear DNA microsatellites indicates restricted gene flow that is consistent with female philopatry to natal regions (Keeney et al. 2005). Development of region-specific natural biogeochemical tags based on vertebral chemistry may permit further testing of site fidelity and philopatry for this species, as well as facilitate examination of connectivity between juvenile and adult populations.
Materials and methods
Sample collection and preparation
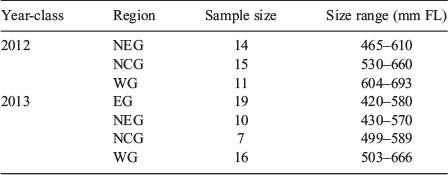
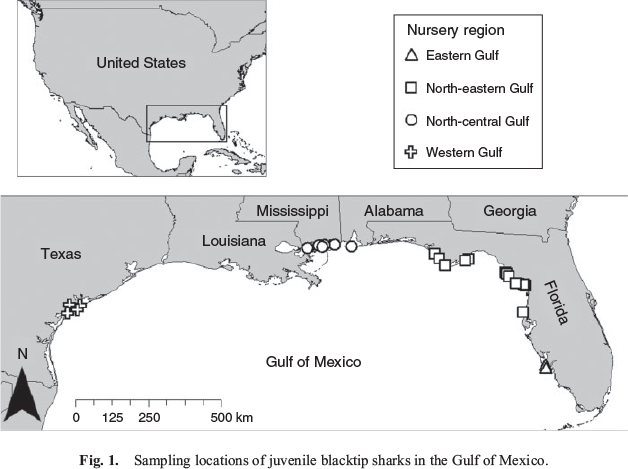
Juvenile blacktip sharks were collected opportunistically in 2012 and 2013 (Table 1) during the Gulf of Mexico States Shark Pupping and Nursery Survey (GULFSPAN). The GULFSPAN survey is a fisheries-independent gill net survey that was initiated by the National Marine Fisheries Service (NMFS) Laboratory in Panama City (FL, USA) in 2003 and occurs annually within several coastal areas of the Gulf. Samples were also obtained from fisheries-independent surveys by the Mote Marine Laboratory (Sarasota, FL, USA), Florida State University Coastal and Marine Laboratory (St Teresa, FL, USA), Dauphin Island Sea Laboratory (Dauphin Island, AL, USA), University of Southern Mississippi Gulf Coast Research Laboratory (Ocean Springs, MS, USA) and Texas Parks and Wildlife Department (Port O’Connor, TX, USA) using gill nets or longlines. Samples of the 2012 and 2013 year-classes were collected from four regions in the Gulf (Fig. 1): (1) the eastern Gulf (EG) off the south-west coast of Florida; (2) the north-eastern Gulf (NEG) from the Big Bend west through the Florida panhandle; (3) the north-central Gulf (NCG) within the state boundaries of Alabama and Mississippi; and (4) the western Gulf (WG) in San Antonio Bay, Texas.
|
|
Thoracic vertebrae were removed in the field or in the laboratory and stored frozen until processed. Excess tissue and haemal and neural arches were excised with a scalpel (Fig. 2a). Individual vertebrae were sectioned along the sagittal plane with a Buehler (Lake Bluff, IL, USA) Isomet low-speed saw, resulting in ~0.5-mm sections (Fig. 2b). Vertebral sections were placed in acid-leached polystyrene cells filled with ultrapure water (18 MΩ cm–1) and sonicated for 1 h. Following sonication, sections were rinsed with ultrapure water, transferred to acid-leached cell wells and dried under a Class 10 laminar flow clean hood.

|
Sample analysis
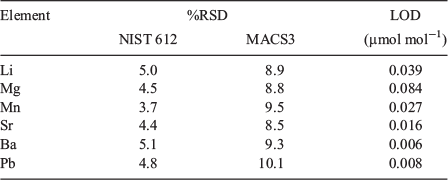
Vertebral chemistry was analysed with laser ablation–inductively coupled plasma–mass spectrometry (LA-ICP-MS). The LA-ICP-MS system consisted of a Nd : YAG NWR213 (Electro Scientific Industries, Portland, OR, USA) laser ablation system coupled with an Agilent Technologies (Santa Clara, CA, USA) 7700x ICP-MS. Helium (flow rate 0.7 L min–1) was used to sweep ablated material from the ablation cell then mixed with argon (flow rate 0.79 L min–1) and transported the ICP-MS. Sample order was randomised and LA-ICP-MS analysis consisted of three parallel laser ablation transects through the corpus calcareum (Fig. 2c). Transects were spaced 120 µm apart and each transect was preablated (laser spot size 100 µm, speed 110 µm s–1, repetition rate 5 Hz, and fluence 0.41 J cm–2) before analysis. Ablation transects consisted of a spot size of 80 µm, a scan speed of 11 µm s–1, a repetition rate of 10 Hz and fluence of 4.0 J cm–2. Background counts were collected 30 s before each transect and subtracted from the signal. Reference materials from the United States Geological Survey (USGS MACS3) and National Institute of Standards and Technology (NIST 612) were ablated every 45 min to evaluate analytical precision and correct for instrument drift. Counts per second (CPS) were converted to units of concentration using the Trace_ElementIS data reduction scheme in Iolite (version 2.5, School of Earth Sciences, University of Melbourne, Melbourne, Australia) using Ca (35 wt% in shark vertebrae; Tillett et al. 2011) as an internal standard. We monitored eight elements (7Li, 24Mg, 43Ca, 44Ca, 31P, 55Mn, 88Sr, 137Ba and 208Pb). Because the concentration of 31P is not certified in the MACS3, it was excluded from subsequent analyses. The concentrations of the remaining elements were normalised to 43Ca (element : Ca) and are expressed as micromoles per mole (Li : Ca, Mn : Ca, Ba : Ca, Pb : Ca) or millimoles per mole (Mg : Ca and Sr : Ca).
Data analysis
The portion of each transect that represented postnatal growth was measured with a Nikon (Tokyo, Japan) SMZ1500 dissecting microscope and Nikon’s NIS-elements BR 4.00.07 imaging software. The minimum amount of postnatal vertebral tissue analysed among samples was 170 µm. Therefore, data from the first 170 µm immediately following the birth mark were averaged among the three transects of each vertebral section to estimate chemical signatures. Assuming a mean birthday of 1 May (Carlson et al. 2006), this region represents ~1 month of life.
Univariate normality was assessed graphically and homogeneity of variance was tested using Levene’s test. Individual element : Ca ratios that did not meet parametric assumptions were natural log transformed, but transformation of Li : Ca and Pb : Ca for the 2013 year-class did not result in equal variances among regions. Because the results of non-parametric and parametric univariate tests yielded the same significance and produced identical multiple comparison results, we only present the results of parametric tests. One multivariate outlier was identified and removed from the dataset. Graphical assessment of multivariate dispersion using the Euclidian distances confirmed that homogeneity of variances was met for the multivariate dataset when using raw data of Li : Ca and Pb : Ca.
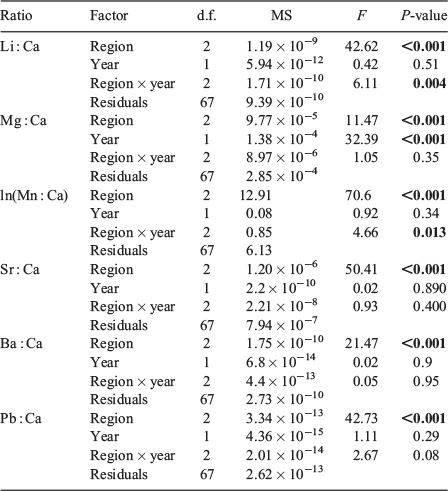
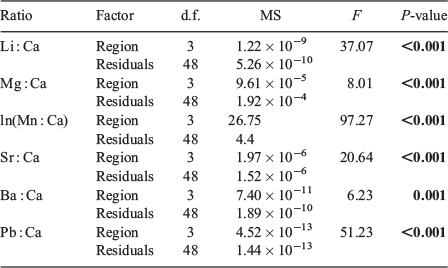
Differences in multi-elemental signatures among regions (excluding the EG, which was only sampled in 2013) and between years were tested with a two-way multivariate analysis of variance (MANOVA) with Pillai’s Trace as the test statistic. Regional and year-class differences for individual element : Ca ratios were tested with a two-way analysis of variance (ANOVA), again for all regions except the EG. A second MANOVA was computed among all regions sampled in 2013, which was followed by an ANOVA for individual element : Ca ratios. Significant pairwise differences in element : Ca ratios within year-classes were identified with Tukey’s honestly significant difference (HSD) tests. A linear discriminant function analysis (LDFA) was computed for each year-class separately, and then for all data combined, to assess the ability of vertebral chemical signatures to distinguish sample regions. The accuracy of LDFA models was assessed via cross-validated classification accuracies. Initial LDFAs included all six elements; however, previous studies of the abiotic and biotic factors that influence vertebral chemistry suggest Mn : Ca is primarily derived from dietary sources and may be influenced by internal nutrient reserves (see Discussion). Thus, LDFAs were also run without Mn : Ca. All statistical analyses were performed in R version 3.2.1 (R Foundation for Statistical Computing, Vienna, Austria).
Results
Each element assayed was consistently above detection limits, with the exception of Pb. Values of Pb below detection were not common (0.1%) within the region of interest and, when present, were replaced with the limits of detection (LOD; Table 2). Relative standard deviations (%RSD) were 3.7–5.1% for the NIST 612 standard and 8.8–10.1% for the MACS3 standard (Table 2).
|
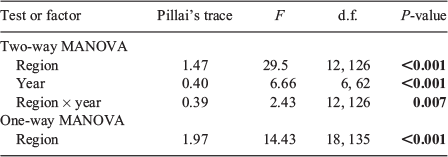
Multi-element chemical signatures differed significantly among the three regions sampled in 2012 and 2013 (F12,126 = 29.5, P < 0.001) and between years (F6,62 = 6.66, P < 0.001), with a significant interaction between factors (F12,126 = 2.43, P = 0.007; Table 3). There was also a significant difference in multi-element signatures among regions sampled in 2013 (F18,135 = 14.43, P < 0.001; Table 3).
|
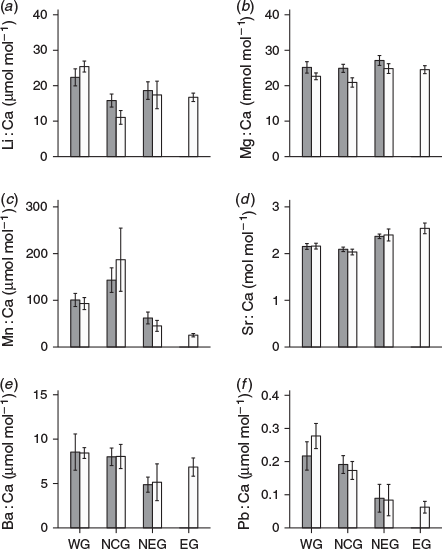
Results of the two-way ANOVA (Table 4) indicated all elemental ratios were significantly different among regions (P < 0.001), but only Mg : Ca exhibited a significant difference between year-classes (P < 0.001). There were significant interactions between region and year-class for Li : Ca and Mn : Ca (P ≤ 0.03). Regional differences in mean element : Ca were also observed among the four regions sampled in 2013 (Table 5) and showed the same general trend observed in 2012 (Fig. 3). Sharks from the NCG tended to have the lowest mean Li : Ca, Mg : Ca and Sr : Ca and highest mean for Mn : Ca, whereas sharks from the WG had the highest mean Li : Ca, Ba : Ca and Pb : Ca. Sharks from the NEG had the highest mean vertebral Mg : Ca and Sr : Ca and lowest values for Ba : Ca and Pb : Ca. Both NEG and EG exhibited similar element : Ca ratios, with the exception of Mn : Ca (P < 0.001). In fact, Mn : Ca was the only elemental ratio that was unique for each region within year-class (P < 0.05).
|
|
|
Mean cross-validated classification accuracies from LDFAs including all elemental ratios were 80, 90 and 88% for 2012, 2013 and both year-classes combined respectively (Fig. 4). Excluding Mn : Ca yielded a slightly higher mean classification accuracy of 81% for 2012. However, the exclusion of Mn : Ca yielded lower mean classification accuracies of 84 and 85% for the 2013 year-class and both year-classes combined respectively (Fig. 4). The lowest classification accuracy was observed for WG samples of the 2012 year-class (54%), whereas the highest classification accuracy was for NCG samples of the 2013 year-class (100%).

|
Discussion
Results from the present study demonstrate the potential of using natural biogeochemical tags in juvenile blacktip shark vertebrae to distinguish Gulf nursery regions. Although water chemistry and other environmental factors that likely drive these biogeochemical signatures were not available for all regions, spatial differences in environmental conditions were clearly sufficient to produce distinct signatures among regions of interest (for a review, see Elsdon et al. 2008). Earlier attempts to distinguish shark nursery regions based on vertebral biogeochemical signatures had mixed success at similar spatial scales (e.g. hundreds to thousands of kilometres). Smith (2013) reported that vertebral signatures of juvenile scalloped hammerhead sharks (Sphyrna lewini) collected from three nursery areas along the Pacific coast of Mexico produced site-specific classification accuracies ranging from 26 to 80%. Tillett et al. (2011) reported significant differences among the vertebral chemistry of bull sharks (Carcharhinus leucas) among six nurseries in Australia, but noted that there was no difference in the vertebral chemistry of pigeye sharks (Carcharhinus amboinensis). This is not too surprising considering that neonate bull sharks frequent areas of lower salinity than the pigeye shark (Knip et al. 2011). Similar to neonate and juvenile pigeye sharks, blacktip and scalloped hammerhead sharks also prefer higher salinities than neonate bull sharks (Hueter and Tyminski 2007; Parsons and Hoffmayer 2007; Steiner et al. 2007). Thus, it appears that watersheds, basin geology or local biogeochemical cycling were more distinct among regions examined in the present study compared with the previous work by Tillett et al. (2011) and Smith (2013).
The high mobility of juvenile individuals can also create problems when applying natural biogeochemical tags because an assumption of this approach is that samples of a target group or region have not mixed with other groups or regions (Elsdon et al. 2008). Thus, the usefulness of these tags will be constrained by spatial scales over which minimal mixing occurs between adjacent areas of interest. Heupel et al. (2004) reported that young-of-year blacktip sharks in Terra Ceia Bay (FL, USA) had restricted core home ranges (mean monthly 50% kernal densities ≤ 0.5 km2) throughout the summer, but the degree to which they ventured outside their core home range increased in the month of July. Given the relatively large distance between regions in the present study and the high degree of site fidelity and limited home ranges reported by Heupel et al. (2004) for juvenile blacktips, the probability we sampled migrants from adjacent regions is extremely low. Because the activity space of juvenile blacktip sharks increases throughout their first summer of life, focussed sample collection during the early months of the birth season will increase the probability an individual is captured in its natal estuary. By increasing sampling effort in and around putative nurseries, it may be possible to determine the spatial resolution at which one can accurately assign natal origin.
Failing to account for intra-annual variability, especially in populations that exhibit a protracted spawning and birth season, can have a strong effect on classification accuracies and estimates of connectivity (Cook 2011; Smith 2013). In the present study, almost all sharks exhibited temporal changes in vertebral chemistry, which reflect temporal variability in abiotic (temperature and water chemistry) and biotic (expanding home range) factors. Our limited sample size precludes statistical analysis of how classification accuracies vary when sampling different regions along the vertebrae or whether sharks born at different times within an estuary have different natal signatures (Smith 2013). The duration of the birth season for blacktip sharks in the Gulf is ~2 months (Baremore and Passerotti 2013), which is much shorter than that of the scalloped hammerhead population along the Pacific coast of Mexico, which occurs between May and October (Smith 2013). Hamer et al. (2003) reported that separating juveniles into two intra-annual cohorts had a negligible effect on classification accuracies for age-0+ individuals of the snapper (Pagrus auratus) that settle in coastal nurseries of Victoria, Australia, between mid-December and March. Therefore, intra-annual variability may have little effect on classification accuracies of blacktip sharks in the Gulf given the short birth season.
The usefulness of natural biogeochemical tags derived from calcified structures stems from the relationship between water chemistry, other environmental factors and element : Ca ratios in the structures themselves. Under estuarine conditions, bony fish otolith Sr : Ca generally shows a positive correlation with salinity, whereas Ba : Ca typically shows a negative relationship with salinity (Limburg 1995; Elsdon and Gillanders 2005; Macdonald and Crook 2010). Because a large portion of otolith Sr and Ba is derived from the surrounding water (Walther and Thorrold 2006; Webb et al. 2012; Izzo et al. 2015), fluctuations in Sr : Ca and Ba : Ca can be used to infer the relative salinity of environments experienced by a fish during its lifetime (e.g. Limburg 1995). Similarly, Scharer et al. (2012) reported that the Sr : Ca in vertebral sections of the smalltooth sawfish (Pristis pectinata) had a positive correlation with salinity, and Smith et al. (2013) reported that the Ba : Ca in the vertebrae of round stingray (Urobatis halleri) was correlated with Ba in seawater. For juvenile blacktip sharks examined here, there were regional differences in Sr : Ca and Ba : Ca that were inversely related. However, it is difficult to attribute this pattern to regional differences in salinity without water chemistry data because the amount of Sr and Ba in freshwater end members will affect the amount of Sr and Ba available at a particular salinity (Wells et al. 2003; Kraus and Secor 2004). Moreover, several factors, including growth rate, ontogeny, temperature and diet, can affect the relationship between water chemistry and element : Ca deposition in calcified structures (for a review, see Sturrock et al. 2012). Growth rate does not appear to affect Li : Ca, Mg : Ca, Mn : Ca, Sr : Ca and Ba : Ca deposition in the round stingray; however, Mg : Ca, Mn : Ca and Ba : Ca were significantly affected by water temperature (Smith et al. 2013). The work of Smith et al. (2013) represents the only study to focus on the relationship between water chemistry and vertebral element : Ca in elasmobranchs; thus, we know little about the influences mentioned above and the extent of interspecies variability.
It is not necessary to fully elucidate the underlying abiotic and biotic factors driving regional variability in element : Ca ratios of calcified structures to use them as natural biogeochemical tags to examine population connectivity. However, other biologically relevant questions can be addressed if the primary factors influencing vertebral chemistry can be determined. Previously, regional differences in otolith Mn : Ca in gag grouper (Mycteroperca microlepis) and red snapper (Lutjanus campechanus) across the Gulf were attributed to latitudinal differences in soil chemistry (Hanson et al. 2004; Sluis et al. 2012). However, the majority of Mn in elasmobranch soft tissue appears to be derived from dietary sources (Mathews and Fisher 2009), which may explain observed enrichment in vertebral Mn : Ca relative to water Mn : Ca (Smith et al. 2013). An increase in Mn : Ca immediately following birth was present in the vertebrae of sharks from all regions in the present study, thus suggesting a common process influencing vertebral Mn : Ca. It is not clear to what degree the distribution of Mn is driven by the protein content of specific portions of vertebrae (Sturrock et al. 2012). Such transition metals do show an affinity for protein binding sites (Miller et al. 2006) and ~28% of otolith Mn can be bound to water soluble proteins (Izzo et al. 2016). However, there may also be a physiological process at play. The neonatal life stage is particularly stressful for elasmobranchs and can be associated with a decline in body mass (Duncan and Holland 2006) and high mortality (Heupel and Simpfendorfer 2002; Duncan and Holland 2006). Neonates rely on maternal nutrient reserves in the form of an enlarged liver to compensate for the lack of foraging experience (Hussey et al. 2010; Olin et al. 2011). Because the liver plays an important role in Mn homeostasis (Aschner and Aschner 2005; Madejczyk et al. 2009), metabolising liver tissue may result in excess Mn entering the bloodstream that becomes deposited in the vertebrae during the first weeks or months of life. Although Mn : Ca was the only elemental ratio that was significantly different among regions in both years in the present study, the possible link to food availability and maternal investment may result in intra-annual variability in vertebral Mn : Ca in a given area as less fit individuals perish. Thus, it is unknown whether vertebral Mn : Ca ratios of sharks collected early in the year would accurately reflect that of the surviving members of a cohort. Excluding Mn : Ca demonstrated that, although useful for discriminating groups, one can still achieve moderate to high classification accuracies using an array of other minor and trace elements.
Results from the present study suggest that examining connectivity between blacktip shark nurseries and offshore adult populations based on vertebral biogeochemical signatures holds some promise, but one factor that may compromise the effectiveness of such an approach is the unknown metabolic stability of vertebral tissue. Bone in higher vertebrates is constantly remodelled owing to its high healing potential (Kalfas 2001). The limited physiological response of elasmobranchs following natural (Officer et al. 1995) and experimentally induced (Ashhurst 2004) trauma suggests elasmobranchs may not be capable of extensive skeletal remodelling. However, unsuccessful attempts to age sharks based on radionuclide decay (226Ra : 210Pb) may indicate metabolic instability of the tissue, but another explanation would be exogenous uptake of 210Pb during growth band formation (Welden et al. 1987; Fenton 2001). Furthermore, the success of bomb radiocarbon age validation (e.g. Campana et al. 2002; Passerotti et al. 2010, 2014), use of stable isotopes to detect ontogenetic shifts in diet (Estrada et al. 2006) and verification of annual bands using Sr : Ca patterns that correspond to known life history characteristics (Scharer et al. 2012) indicates the corpus calcareum likely remains relatively inert once formed.
Important questions remain surrounding the mechanisms governing mineral dynamics and sites of inclusion (e.g. substitution in calcium phosphate hydroxyapatite matrix, bound to proteins or incorporated interstitially), but the results of the present study demonstrate that vertebral biogeochemical tags can be used to accurately distinguish nursery regions of blacktip sharks and may provide a means to estimate the relative contribution of different areas to the adult stock. By building an atlas of cohort-specific vertebral chemical signatures from different areas, it may be possible to infer the natal origin of adult sharks. This type of information would be particularly useful because some coastal sharks possess life history traits that make them vulnerable to overharvest. Whether protecting nursery areas should be the most important aspect of a management plan will depend on the biology of the species and life stage removed by the fishery (Kinney and Simpfendorfer 2009). However, estimates of connectivity between juvenile and adult populations can provide the insight necessary to evaluate the effectiveness of nursery conservation as a management tool.
Acknowledgements
Funding support for this project was provided by National Oceanic and Atmospheric Administration (NOAA) Fisheries, Panama City Laboratory, the Highly Migratory Species Division of NOAA Fisheries and the National Science Foundation (grant number DBI-1319188 to W. F. Patterson III). The authors thank D. M. Bethea and K. Smith (NOAA Fisheries), W. J. Bubley (Texas Parks and Wildlife Department), J. M. Drymon, M. N. Schrandt, A. M. Koretz and K. C. Gregalis (Dauphin Island Sea Lab) R. D. Grubbs, C. T. Peterson and J. L. Imhoff (Florida State University Coastal and Marine Laboratory), J. M. Hendon, J. M. Higgs and S. Ashworth (Gulf Coast Research Laboratory), R. J. D. Wells (Texas A&M University Galveston), N. M. Whitney and J. Morris (Mote Marine Laboratory) and the many staff members, students and volunteers who participated in field collection. The authors are also grateful for the guidance and advice provided by N. R. Miller (University of Texas Austin) during LA-ICP-MS analyses, and instrument time and technical support provide by C. J. P. O’Connor, J. Wilkins and the staff of the Scientific Division of Electro Scientific Industries (Bozeman, MT, USA).
References
Aschner, J. L., and Aschner, M. (2005). Nutritional aspects of manganese homeostasis. Molecular Aspects of Medicine 26, 353–362.| Nutritional aspects of manganese homeostasis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXpslygur4%3D&md5=4521afab529a4f1eab4484940abbd8ddCAS | 16099026PubMed |
Ashhurst, D. E. (2004). The cartilaginous skeleton of an elasmobranch fish does not heal. Matrix Biology 23, 15–22.
| The cartilaginous skeleton of an elasmobranch fish does not heal.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXksVaku7g%3D&md5=81780c40e8be44b21d9b0c0f92de2dffCAS | 15172034PubMed |
Baremore, I. E., and Passerotti, M. S. (2013). Reproduction of the blacktip shark in the Gulf of Mexico. Marine and Coastal Fisheries 5, 127–138.
| Reproduction of the blacktip shark in the Gulf of Mexico.Crossref | GoogleScholarGoogle Scholar |
Beck, M. W., Heck, K. L., Able, K. W., Childers, D. L., Eggleston, D. B., Gillanders, B. M., Halpern, B., Hays, C. G., Hoshino, K., Minello, T. J., Orth, R. J., Sheridan, P. F., and Weinstein, M. P. (2001). The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates. Bioscience 51, 633–641.
| The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates.Crossref | GoogleScholarGoogle Scholar |
Campana, S. E. (1999). Chemistry and composition of fish otoliths: pathways, mechanisms and applications. Marine Ecology Progress Series 188, 263–297.
| Chemistry and composition of fish otoliths: pathways, mechanisms and applications.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXjtFKmtA%3D%3D&md5=1c1a0b19f44f50ec78fd6669fd952b22CAS |
Campana, S. E., Natanson, L. J., and Myklevoll, S. (2002). Bomb dating and age determination of large pelagic sharks. Canadian Journal of Fisheries and Aquatic Sciences 59, 450–455.
| Bomb dating and age determination of large pelagic sharks.Crossref | GoogleScholarGoogle Scholar |
Carlson, J. K., Sulikowski, J. R., and Baremore, I. E. (2006). Do differences in life history traits exist for blacktip sharks, Carcharhinus limbatus, from the United States South Atlantic Bight and Eastern Gulf of Mexico? Environmental Biology of Fishes 77, 279–292.
| Do differences in life history traits exist for blacktip sharks, Carcharhinus limbatus, from the United States South Atlantic Bight and Eastern Gulf of Mexico?Crossref | GoogleScholarGoogle Scholar |
Cook, G. S. (2011). Changes in otolith microchemistry over a protracted spawning season influence assignment of natal origin. Marine Ecology Progress Series 423, 197–209.
| Changes in otolith microchemistry over a protracted spawning season influence assignment of natal origin.Crossref | GoogleScholarGoogle Scholar |
Duncan, K. M., and Holland, K. N. (2006). Habitat use, growth rates and dispersal patterns of juvenile scalloped hammerhead sharks Sphyrna lewini in a nursery habitat. Marine Ecology Progress Series 312, 211–221.
| Habitat use, growth rates and dispersal patterns of juvenile scalloped hammerhead sharks Sphyrna lewini in a nursery habitat.Crossref | GoogleScholarGoogle Scholar |
Elsdon, T. S., and Gillanders, B. M. (2005). Alternative life-history patterns of estuarine fish: barium in otoliths elucidates freshwater residency. Canadian Journal of Fisheries and Aquatic Sciences 62, 1143–1152.
| Alternative life-history patterns of estuarine fish: barium in otoliths elucidates freshwater residency.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXoslGqs7o%3D&md5=820f1beaf2a24658afa7e44f788b4d6fCAS |
Elsdon, T. S., Wells, B. K., Campana, S. E., Gillanders, B. M., Jones, C. M., Limburg, K. E., Secor, D. H., Thorrold, S. R., and Walther, B. D. (2008). Otolith chemistry to describe movements and life-history parameters of fishes: hypotheses, assumptions, limitations and inferences. Oceanography and Marine Biology – an Annual Review 46, 297–330.
| Otolith chemistry to describe movements and life-history parameters of fishes: hypotheses, assumptions, limitations and inferences.Crossref | GoogleScholarGoogle Scholar |
Estrada, J. A., Rice, A. N., Natanson, L. J., and Skomal, G. B. (2006). Use of isotopic analysis of vertebrae in reconstructing ontogenetic feeding ecology in white sharks. Ecology 87, 829–834.
| Use of isotopic analysis of vertebrae in reconstructing ontogenetic feeding ecology in white sharks.Crossref | GoogleScholarGoogle Scholar | 16676526PubMed |
Fenton, G. E. (2001). Radiometric Ageing of Sharks. FRDC Final Report 1994/021. Fisheries Research and Development Corporation, Canberra, Australia.
Hamer, P. A., Jenkins, G. P., and Gillanders, B. M. (2003). Otolith chemistry of juvenile snapper Pagrus auratus in Victorian waters: natural chemical tags and their temporal variation. Marine Ecology Progress Series 263, 261–273.
| Otolith chemistry of juvenile snapper Pagrus auratus in Victorian waters: natural chemical tags and their temporal variation.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXitVaru7o%3D&md5=c96b8b10981f2a27ec74b8e404a3e411CAS |
Hanson, P. J., Koenig, C. C., and Zdanowicz, V. S. (2004). Elemental composition of otoliths used to trace estuarine habitats of juvenile gag Mycteroperca microlepis along the west coast of Florida. Marine Ecology Progress Series 267, 253–265.
| Elemental composition of otoliths used to trace estuarine habitats of juvenile gag Mycteroperca microlepis along the west coast of Florida.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXkt1KktLw%3D&md5=0b8fccf006ceb320d20621202372e0e4CAS |
Heupel, M. R., and Simpfendorfer, C. A. (2002). Estimation of mortality of juvenile blacktip sharks, Carcharhinus limbatus, within a nursery area using telemetry data. Canadian Journal of Fisheries and Aquatic Sciences 59, 624–632.
| Estimation of mortality of juvenile blacktip sharks, Carcharhinus limbatus, within a nursery area using telemetry data.Crossref | GoogleScholarGoogle Scholar |
Heupel, M. R., Simpfendorfer, C. A., and Hueter, R. E. (2004). Estimation of shark home ranges using passive monitoring techniques. Environmental Biology of Fishes 71, 135–142.
| Estimation of shark home ranges using passive monitoring techniques.Crossref | GoogleScholarGoogle Scholar |
Heupel, M. R., Carlson, J. K., and Simpfendorfer, C. A. (2007). Shark nursery areas: concepts, definition, characterization and assumptions. Marine Ecology Progress Series 337, 287–297.
| Shark nursery areas: concepts, definition, characterization and assumptions.Crossref | GoogleScholarGoogle Scholar |
Hueter, R. E., and Tyminski, J. P. (2007). Species-specific distribution and habitat characteristics of shark nurseries in Gulf of Mexico waters off peninsular Florida and Texas. American Fisheries Society Symposium 50, 193–223.
Hueter, R. E., Heupel, M. R., Heist, E. J., and Keeney, D. B. (2005). Evidence of philopatry in sharks and implications for the management of shark fisheries. Journal of Northwest Atlantic Fishery Science 35, 239–247.
| Evidence of philopatry in sharks and implications for the management of shark fisheries.Crossref | GoogleScholarGoogle Scholar |
Hussey, N. E., Wintner, S. P., Dudley, S. F., Cliff, G., Cocks, D. T., and Aaron MacNeil, M. (2010). Maternal investment and size‐specific reproductive output in carcharhinid sharks. Journal of Animal Ecology 79, 184–193.
| Maternal investment and size‐specific reproductive output in carcharhinid sharks.Crossref | GoogleScholarGoogle Scholar | 19845812PubMed |
Izzo, C., Doubleday, Z. A., Schultz, A. G., Woodcock, S. H., and Gillanders, B. M. (2015). Contribution of water chemistry and fish condition to otolith chemistry: comparisons across salinity environments. Journal of Fish Biology 86, 1680–1698.
| Contribution of water chemistry and fish condition to otolith chemistry: comparisons across salinity environments.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC2Mfps1ensg%3D%3D&md5=2e54b18c3611aa2782c2759c17614afbCAS | 26033292PubMed |
Izzo, C., Doubleday, Z. A., and Gillanders, B. M. (2016). Where do elements bind within the otoliths of fish? Marine and Freshwater Research 67, 1072–1076.
| Where do elements bind within the otoliths of fish?Crossref | GoogleScholarGoogle Scholar |
Kalfas, I. H. (2001). Principles of bone healing. Neurosurgical Focus 10, 1–4.
| Principles of bone healing.Crossref | GoogleScholarGoogle Scholar |
Keeney, D. B., Heupel, M. R., Hueter, R. E., and Heist, E. J. (2005). Microsatellite and mitochondrial DNA analyses of the genetic structure of blacktip shark (Carcharhinus limbatus) nurseries in the northwestern Atlantic, Gulf of Mexico, and Caribbean Sea. Molecular Ecology 14, 1911–1923.
| Microsatellite and mitochondrial DNA analyses of the genetic structure of blacktip shark (Carcharhinus limbatus) nurseries in the northwestern Atlantic, Gulf of Mexico, and Caribbean Sea.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXlslemsb8%3D&md5=af7262c3d63fac51cc2351e5004f736eCAS | 15910315PubMed |
Kinney, M. J., and Simpfendorfer, C. A. (2009). Reassessing the value of nursery areas to shark conservation and management. Conservation Letters 2, 53–60.
| Reassessing the value of nursery areas to shark conservation and management.Crossref | GoogleScholarGoogle Scholar |
Knip, D. M., Heupel, M. R., Simpfendorfer, C. A., Tobin, A. J., and Moloney, J. (2011). Wet-season effects on the distribution of juvenile pigeye sharks, Carcharhinus amboinensis, in tropical nearshore waters. Marine and Freshwater Research 62, 658–667.
| Wet-season effects on the distribution of juvenile pigeye sharks, Carcharhinus amboinensis, in tropical nearshore waters.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXnvFanu7k%3D&md5=cea979b6a3f17b7d775bb2c5e84ae12bCAS |
Kohler, N. E., and Turner, P. A. (2001). Shark tagging: a review of conventional methods and studies. Environmental Biology of Fishes 60, 191–224.
| Shark tagging: a review of conventional methods and studies.Crossref | GoogleScholarGoogle Scholar |
Kraus, R. T., and Secor, D. H. (2004). Incorporation of strontium into otoliths of an estuarine fish. Journal of Experimental Marine Biology and Ecology 302, 85–106.
| Incorporation of strontium into otoliths of an estuarine fish.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXjtVWntbw%3D&md5=7b1b04bb9895fb8a9bab2affe063f61aCAS |
Limburg, K. E. (1995). Otolith strontium traces environmental history of subyearling American shad Alosa sapidissima. Marine Ecology Progress Series 119, 25–35.
| Otolith strontium traces environmental history of subyearling American shad Alosa sapidissima.Crossref | GoogleScholarGoogle Scholar |
Macdonald, J. I., and Crook, D. A. (2010). Variability in Sr : Ca and Ba : Ca ratios in water and fish otoliths across an estuarine salinity gradient. Marine Ecology Progress Series 413, 147–161.
| Variability in Sr : Ca and Ba : Ca ratios in water and fish otoliths across an estuarine salinity gradient.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXht12msLzM&md5=cfd9e1f60ad5bf19da6bcb38eba76372CAS |
Madejczyk, M. S., Boyer, J. L., and Ballatori, N. (2009). Hepatic uptake and biliary excretion of manganese in the little skate, Leucoraja erinacea. Comparative Biochemistry and Physiology Part C Toxicology & Pharmacology 149, 566–571.
| Hepatic uptake and biliary excretion of manganese in the little skate, Leucoraja erinacea.Crossref | GoogleScholarGoogle Scholar |
Mathews, T., and Fisher, N. S. (2009). Dominance of dietary intake of metals in marine elasmobranch and teleost fish. The Science of the Total Environment 407, 5156–5161.
| Dominance of dietary intake of metals in marine elasmobranch and teleost fish.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXptVSjtLs%3D&md5=61c87409bb2023bf88eadaf796885cdfCAS | 19580992PubMed |
Miller, M. B., Clough, A. M., Batson, J. N., and Vachet, R. W. (2006). Transition metal binding to cod otolith proteins. Journal of Experimental Marine Biology and Ecology 329, 135–143.
| Transition metal binding to cod otolith proteins.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XotVKlsw%3D%3D&md5=6016d2fabe754800e5d5c24527808f4aCAS |
Officer, R. A., Clement, J. G., and Rowler, D. K. (1995). Vertebral deformities in a school shark, Galeorhinus galeus: circumstantial evidence for endoskeletal resorption? Journal of Fish Biology 46, 85–98.
| Vertebral deformities in a school shark, Galeorhinus galeus: circumstantial evidence for endoskeletal resorption?Crossref | GoogleScholarGoogle Scholar |
Olin, J. A., Hussey, N. E., Fritts, M., Heupel, M. R., Simpfendorfer, C. A., Poulakis, G. R., and Frisk, A. T. (2011). Maternal meddling in neonatal sharks: implications for interpreting stable isotopes in young animals. Rapid Communications in Mass Spectrometry 25, 1008–1016.
| Maternal meddling in neonatal sharks: implications for interpreting stable isotopes in young animals.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXktFWqsb0%3D&md5=f60f30de2db571fb8b6d408985916944CAS | 21452377PubMed |
Parsons, G. R., and Hoffmayer, E. R. (2007). Identification and characterization of shark nursery grounds along the Mississippi and Alabama gulf coasts. American Fisheries Society Symposium 50, 301–316.
Passerotti, M. S., Carlson, J. K., Piercy, A. N., and Campana, S. E. (2010). Age validation of great hammerhead shark (Sphyrna mokarran), determined by bomb radiocarbon analysis. Fishery Bulletin 108, 346–351.
Passerotti, M. S., Andrews, A. H., Carlson, J. K., Winter, S. P., Goldman, K. J., and Natanson, L. J. (2014). Maximum age and missing time in the vertebrae of sand tiger shark (Carcharias taurus): validated lifespan from bomb radiocarbon dating in the western North Atlantic and southwestern Indian Oceans. Marine and Freshwater Research 65, 674–687.
| Maximum age and missing time in the vertebrae of sand tiger shark (Carcharias taurus): validated lifespan from bomb radiocarbon dating in the western North Atlantic and southwestern Indian Oceans.Crossref | GoogleScholarGoogle Scholar |
Pitts, L. C., and Wallace, G. T. (1994). Lead deposition in the shell of the bivalve, Mya arenaria: an indicator of dissolved lead in seawater. Estuarine, Coastal and Shelf Science 39, 93–104.
| Lead deposition in the shell of the bivalve, Mya arenaria: an indicator of dissolved lead in seawater.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2MXhvVWitrY%3D&md5=2f1ea8bd319494591dee886b017544e6CAS |
Scharer, R. M., Patterson, W. F., Carlson, J. K., and Poulakis, G. R. (2012). Age and growth of endangered smalltooth sawfish (Pristis pectinata) verified with LA-ICP-MS analysis of vertebrae. PLoS One 7, e47850.
| Age and growth of endangered smalltooth sawfish (Pristis pectinata) verified with LA-ICP-MS analysis of vertebrae.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xhs1ansbvK&md5=deaf71fe42eb4eaf488b7eadbe9de876CAS | 23082225PubMed |
Sluis, M. Z., Barnett, B. K., Patterson, W. F., Cowan, J. H., and Shiller, A. M. (2012). Discrimination of juvenile red snapper otolith chemical signatures from Gulf of Mexico nursery regions. Marine and Coastal Fisheries 4, 587–598.
| Discrimination of juvenile red snapper otolith chemical signatures from Gulf of Mexico nursery regions.Crossref | GoogleScholarGoogle Scholar |
Smith, W. D. (2013). Vertebral elemental markers in elasmobranchs: potential for reconstructing environmental history and population structure. Ph.D. Thesis, Oregon State University, Corvallis.
Smith, W. D., Miller, J. A., and Heppell, S. S. (2013). Elemental markers in elasmobranchs: effects of environmental history and growth on vertebral chemistry. PLoS One 8, e62423.
| Elemental markers in elasmobranchs: effects of environmental history and growth on vertebral chemistry.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXhsFOrt77I&md5=be20988421835f7224f1041e40d81216CAS | 24098320PubMed |
Southeast Data, Assessment, and Review (2012). Stock Assessment Report of SEDAR 29, HMS Gulf of Mexico Blacktip Shark. (Southeast Fisheries Science Center: Charleston, SC.) Available at http://sedarweb.org/docs/sar/S29_GOM%20blacktip%20report_SAR_final.pdf [Verified 8 November 2015].
Steiner, P. A., Michel, M., and O’Donnell, P. M. (2007). Notes on the occurrence and distribution of elasmobranchs in the Ten Thousand Islands Estuary, Florida. American Fisheries Society Symposium 50, 237–250.
Sturrock, A. M., Trueman, C. N., Darnaude, A. M., and Hunter, E. (2012). Can otolith elemental chemistry retrospectively track migrations in fully marine fishes? Journal of Fish Biology 81, 766–795.
| Can otolith elemental chemistry retrospectively track migrations in fully marine fishes?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xhtl2ltLvN&md5=9546ccc49c9558b3fe7de3bf4a0f68efCAS | 22803735PubMed |
Tillett, B. J., Meekan, M. G., Parry, D., Munksgaard, N., Field, I. C., Thorburn, D., and Bradshaw, C. J. (2011). Decoding fingerprints: elemental composition of vertebrae correlates to age-related habitat use in two morphologically similar sharks. Marine Ecology Progress Series 434, 133–142.
| Decoding fingerprints: elemental composition of vertebrae correlates to age-related habitat use in two morphologically similar sharks.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXht1GktrjJ&md5=e030bcd9c1c01c00ee47a5acc7a822e1CAS |
Vasconcelos, R. P., Reis-Santos, P., Costa, M. J., and Cabral, H. N. (2011). Connectivity between estuaries and marine environment: Integrating metrics to assess estuarine nursery function. Ecological Indicators 11, 1123–1133.
| Connectivity between estuaries and marine environment: Integrating metrics to assess estuarine nursery function.Crossref | GoogleScholarGoogle Scholar |
Walther, B. D., and Thorrold, S. R. (2006). Water, not food, contributes the majority of strontium and barium deposited in the otoliths of a marine fish. Marine Ecology Progress Series 311, 125–130.
| Water, not food, contributes the majority of strontium and barium deposited in the otoliths of a marine fish.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XntFaltrw%3D&md5=c081c33c5bf8c48f6356395d8f72c789CAS |
Webb, S. D., Woodcock, S. H., and Gillanders, B. M. (2012). Sources of otolith barium and strontium in estuarine fish and the influence of salinity and temperature. Marine Ecology Progress Series 453, 189–199.
| Sources of otolith barium and strontium in estuarine fish and the influence of salinity and temperature.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhtVOmtrrN&md5=741e333ddffda2d19455d220ce8a3a4dCAS |
Welden, B. A., Cailliet, G. M., and Flegal, A. R. (1987). Comparison of radiometric with vertebral band age estimates in four California elasmobranchs. In ‘Age and Growth of fish’. (Eds R. C. Summerfelt and G. E. Hall.) pp. 301–315. (Iowa State University Press: Ames, IA.)
Wells, B. K., Bath, G. E., Thorrold, S. R., and Jones, C. M. (2000). Incorporation of strontium, cadmium, and barium in juvenile spot (Leiostomus xanthurus) scales reflects water chemistry. Canadian Journal of Fisheries and Aquatic Sciences 57, 2122–2129.
| Incorporation of strontium, cadmium, and barium in juvenile spot (Leiostomus xanthurus) scales reflects water chemistry.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXjsFGmuw%3D%3D&md5=5fda68451216b225274edecb5bcf7954CAS |
Wells, B. K., Rieman, B. E., Clayton, J. L., Horan, D. L., and Jones, C. M. (2003). Relationships between water, otolith, and scale chemistries of westslope cutthroat trout from the Coeur d’Alene River, Idaho: the potential application of hard-part chemistry to describe movements in freshwater. Transactions of the American Fisheries Society 132, 409–424.
| Relationships between water, otolith, and scale chemistries of westslope cutthroat trout from the Coeur d’Alene River, Idaho: the potential application of hard-part chemistry to describe movements in freshwater.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXksVGktLk%3D&md5=e00656f5247624767b1651d8c0805c9fCAS |
Yeiser, B. G., Heupel, M. R., and Simpfendorfer, C. A. (2008). Occurrence, home range and movement patterns of juvenile bull (Carcharhinus leucas) and lemon (Negaprion brevirostris) sharks within a Florida estuary. Marine and Freshwater Research 59, 489–501.
| Occurrence, home range and movement patterns of juvenile bull (Carcharhinus leucas) and lemon (Negaprion brevirostris) sharks within a Florida estuary.Crossref | GoogleScholarGoogle Scholar |