

Influence of climate and environment on post-fire recovery of mountain big sagebrush
Zachary J. Nelson A D , Peter J. Weisberg B and Stanley G. Kitchen CA Program in Ecology, Evolution and Conservation Biology, University of Nevada, 1664 N. Virginia Street, MS 186, Reno, NV 89557, USA.
B Department of Natural Resources and Environmental Science, University of Nevada, 1664 N. Virginia Street, MS 186, Reno, NV 89557, USA.
C USDA Forest Service, Rocky Mountain Research Station, Shrub Sciences Laboratory 735 North 500 East, Provo, UT 84606, USA.
D Corresponding author. Present address: Inyo County Water Department, 135 S. Jackson Street, Independence, CA 93526, USA. Email: z.j.nelson2010@gmail.com
International Journal of Wildland Fire 23(1) 131-142 https://doi.org/10.1071/WF13012
Submitted: 21 January 2013 Accepted: 14 August 2013 Published: 14 November 2013
Abstract
In arid and semi-arid landscapes around the world, wildfire plays a key role in maintaining species diversity. Dominant plant associations may depend upon particular fire regime characteristics for their persistence. Mountain shrub communities in high-elevation landscapes of the Intermountain West, USA, are strongly influenced by the post-fire recovery dynamics of the obligate-seeding shrub, mountain big sagebrush (Artemisia tridentata Nutt. ssp. vaseyana [Rydb.] Beetle). This species is a short-distance disperser with a short-lived seedbank, leading to highly variable post-fire recovery times (15–100 years). We investigated the relative importance of site productivity and seasonal climate in explaining the variance in recovery time for 36 fires, comprising a fire chrono-sequence (from 1971 to 2007) for the Great Basin and Colorado Plateau. A. t. vaseyana recovery was positively related to precipitation in the cool season immediately following fire, likely because deep soil-water recharge that persists throughout the growing season enhances first-year seedling survival. Percentage sand fraction positively correlated with recovery rate yet negatively correlated with live cover in unburnt stands. Our data support the hypothesis that post-fire recovery rate of A. t. vaseyana depends on the climatically controlled ephemerality of the regeneration niche, as is likely true for many arid-land shrub species.
Additional keywords: Artemisia tridentata ssp. vaseyana, Colorado Plateau, fire effects, Great Basin, precipitation variability, succession.
Introduction
Arid and semi-arid landscapes of western North America are predicted to experience dramatic transformations in coming decades as a result of increased temperatures, longer growing seasons and increased fire frequency (Rehfeldt et al. 2006; Westerling et al. 2006). In the Intermountain West, USA, big sagebrush (Artemisia tridentata) plays a foundational role in ecosystem organisation. This widespread species occupies a 500 000-km2 area extending from arid valley bottoms to more mesic montane environments (McArthur and Ott 1996). Notable life-history characteristics are small seeds with short-range anemochorous dispersal, seed dormancy to guard against premature germination in unfavourable winter conditions, lack of resprouting capability and perennial foliage that persists through the winter. Three identified subspecies of big sagebrush have different adaptations to water availability, disturbance and freezing hazard. Bonham et al. (1991) classified the three subspecies into Grime’s (1974) competition-stress-ruderal (CSR) model of plant strategies. Wyoming big sagebrush (A. tridentata ssp. wyomingensis Beetle & Young), a stress tolerator (Bonham et al. 1991), occurs in the driest sites, typically on fine-textured soils, has the fastest root elongation rates but slowest overall growth, variable seed production, lack of seed dormancy and slow germination rates (Meyer 1994). The comparatively high seed production, smaller seeds and faster growth rate of basin big sagebrush (Artemisia tridentata Nutt. ssp. tridentata) are characteristics of a ruderal species (Bonham et al. 1991). Its large growth form is associated with its occurrence in deeper soil washes that are susceptible to higher rates of disturbance by flooding and scouring events. Mountain big sagebrush (A. tridentata ssp. vaseyana [Rydb.] Beetle) has characteristics consistent with those of a competitor species (Bonham et al. 1991). It has consistent but low levels of seed production compared to A. t. ssp. tridentata., presumably owing to a need to devote resources to vegetative growth in these high-elevation competitive environments. A shallower maximum root density than the other two subspecies indicates that competition with shallow-rooted heterospecifics may be important.
Future increase in land surface temperature and change in precipitation regimes could change the ecological role of fire across A. tridentata ecosystems, which range from arid valley bottoms to mesic montane locations. The post-fire regeneration dynamics of A. tridentata are also likely to change, as seedling establishment is strongly limited by soil moisture at lower elevations, and by cold temperatures and snow depth at higher elevations (Burke et al. 1989; Perfors et al. 2003; Poore et al. 2009). Understanding the processes that influence post-fire recovery of A. tridentata and the likely effects on community dynamics will be critical for developing adaptive management strategies in a changing environment.
Post-fire recovery of A. tridentata is highly variable. Recovery to pre-fire cover values can take fewer than 15 years or more than 100 years (Baker 2006; 2011). A. tridentata does not resprout following disturbance and is limited to reestablishment from seed (Daubenmire 1975). A. tridentata seeds lack specialised appendages for long-distance dispersal and most seeds move less than 3 m (Welch 2005); the maximum observed dispersal distance is 33 m (Goodwin 1956). Thus, for large-area, high-severity burns that leave no surviving mature plants as seed sources, recovery from the burn perimeter can take decades (Welch 2005, p. 179; Ziegenhagen and Miller 2009). Ziegenhagen and Miller (2009) concluded in a study on A. t. vaseyana, that regeneration from the seedbank is the critical determinant in recovery trajectories after burns where few surviving plants remain as seed sources. Although seeds are short lived and do not form a persistent seedbank (McDonough and Harniss 1974; Young and Evans 1989; Meyer et al. 1990), a small fraction of seed may remain viable for up to 4 years in the soil (Ziegenhagen and Miller 2009; Wijayratne and Pyke 2012). Wijayratne and Pyke (2012) showed that shallow burial of seed prolongs dormancy – an idea suggested by Meyer (1994) stemming from findings that seed retained light requirements for germination even after chill requirements were satisfied (Meyer et al. 1990).
Recruitment has been shown to correlate with timing and magnitude of precipitation (Cawker 1980; Maier et al. 2001). A study in south-west Montana showed reestablishment after mechanical removal was greater on north-west exposures (Johnson and Payne 1968). However, other studies have not found strong correlations between soil, aspect, climate and post-fire recovery for any subspecies (Lesica et al. 2007). As a consequence of the wide ecological amplitude occupied by the species, environmental correlates of post-fire recovery have varied greatly (Baker 2011). Studies from 70 populations across a wide geographic area spanning from the south-west Great Basin to the western Colorado Plateau, indicate that physiological controls of germination phenology have adapted to different climate regimes to allow germination in the season that maximises establishment success (Meyer 1994). Seeds at mild-winter sites are non-dormant with high germination rates at near-freezing temperatures, whereas at cold-winter sites seed dormancy is high, presumably to avoid precocious autumn germination (Meyer and Monsen 1992). Dormancy may play a role in post-fire recovery because for there to be a post-fire recovery response driven by the soil seedbank, a proportion of the previous year’s seedbank must remain dormant through a growing season. Therefore, populations with a propensity for higher rates of dormancy, an adaptation to climate, may also benefit in the post-fire recovery process from a dormant seedbank, which is needed to reestablish populations in areas where reproductively mature individuals and their respective seed crop have been killed by recent fire.
We undertook a regional study that encompasses a range of environmental conditions to detect and quantify key processes influencing post-fire recovery of A. t. ssp. vaseyana ecosystems. This study has a focus on ssp. vaseyana because it is one component of a larger investigation into historical fire regimes of ssp. vaseyana communities that are seral to woodland. Using a space-for-time substitution, or chrono-sequence, approach, we first quantified the influence of time since fire on post-fire recovery of Artemisia tridentata ssp. vaseyana (hereafter referred to as Artemisia) over a regional scale that included the eastern Great Basin, Colorado Plateau and intervening highlands and mountains (i.e. Utah Highlands). We then evaluated the importance of soil and climate characteristics, including potential lagged effects of precipitation before and after the fire event, while controlling for time since fire (Table 1). Our sample of Artemisia stands represents a successional gradient from recently burnt to conifer co-dominance.

|
Methods
Study area
We located historic fires that burnt from 1972 to 2008 in the eastern Great Basin, Colorado Plateau and Utah Highlands (Fig. 1) spanning 1926 to 2788 m in elevation. In these high plateaus, canyons and mountains, Artemisia comprises a dominant component of the shrubland. Superimposed on the Artemisia-dominated shrublands at intermediate elevations are woodlands that include Pinus edulis, P. monophylla, Juniperus scopulorum and J. osteosperma. Quercus gambelii, and Cercocarpus ledifolius woodlands occur above these zones. P. ponderosa occurs in drier montane forests and Pseudotsuga menziesii in wetter sites. Populus tremuloides occurs in middle and upper elevation zones in mesic areas. Co-dominant shrub species include Amelanchier alnifolia, Berberis repens, Ericameria nauseosa, Chrysothamnus viscidiflorus, Eriogonum spp., Gutierrezia sarothrae, Prunus virginiana, Purshia tridentata, Rosa woodsii, Symporicarpos oreophilus and Tetradymia canescens. Common perennial grasses include Achnatherum hymenoides, A. lettermanii, Pseudoroegneria spicata, Poa fendleriana, and Poa secunda. Winter precipitation (October–March) ranged from 12.2 to 40.7 cm and summer precipitation (April–September) ranged from 16.4 to 29.6 cm.

|
Field methods
We stratified sampling of historic fires among the Colorado Plateau (n = 13), Great Basin (n = 10) and Utah Highlands (n = 13) regions where time since fire ranged from 1 to 36 years and fire sizes ranged from ~20 to 35 200 ha. Sample sites were selected to minimise the potential confounding effects of post-fire reseeding efforts and heavy grazing by livestock. To this end, we consulted local experts familiar with rehabilitation treatments and historic grazing patterns and inspected candidate sites for presence of introduced plant species commonly used in seedings, and evidence of grazing pressure (e.g. proximity to water sources, stubble height, bare ground and manure piles) before selection. Only sites without evidence of reseeding or conspicuous grazing affect following the fire were selected for sampling. Approximately one-third of inspected sites were eliminated using this screening process. Failed post-fire broadcast seedings were possible for some sites; however, in these cases the competitive effects of seeded species on natural recovery were likely negligible. We did not attempt to distinguish and quantify heavy, moderate or light grazing, but excluded sites where a grazing effect was evident from the above indicators. For 27 of the 36 burns we were able to pair burnt with unburnt sites outside of the burnt perimeter that had similar slope, aspect and elevation and so were suitable to serve as a comparison of post-fire recovery rates. One site per fire was selected to be representative of the burnt conditions and one representative reference site per fire was selected nearby within 3.8 km of the burn boundary, although in most cases they were much closer (median distance = 301 m, minimum distance = 19 m). In a post-hoc analysis, we compared the soil texture values between paired sites. Paired t-tests indicated soil texture was not directionally different between burnt and unburnt sites (e.g. unburnt having higher % sand than burnt). We subtracted unburnt values for % sand, % clay and % silt from paired burnt sites, and the computed average difference was less than 1% in all pairs. When absolute change was considered (i.e. transforming all difference values to positive), the average absolute difference between burnt and unburnt sites for clay was 3.31% with a median difference of 2.36% and mode of 1.00%. The average difference for silt was 4.09% with a median of 2.36% and mode 0.64%, and for sand, 6.57% with a median of 6.19% and mode of 1.64%. Given the close agreement of soil texture between paired sites, the use of the unburnt sites as a reference for unburnt vegetation conditions was deemed appropriate.
We estimated year-of-last-fire for eight mature Artemisia stands using evidence collected from fire-scarred P. ponderosa trees (live trees, stumps and snags) located within or near existing Artemisia communities (data on file, USDA Forest Service, Shrub Sciences Laboratory, Provo, UT). Samples were surfaced and cross-dated using standard dendrochronological techniques (Stokes and Smiley 1968) and calendar years were assigned to fire scars based upon the position of individual scars within the dated ring sequence of each tree. Year-of-last-fire for these sites ranged from 1752 to 1928. Sampling for Artemisia recovery was conducted along transects that followed topographic contours and thus had consistent elevation. We used the line-intercept method to quantify canopy cover of woody species directly beneath a 100-m transect line and recorded a new start and end location within an individual crown if a break in live vegetation was greater than 10 cm (Boyd et al. 2007).
Distance to the burn perimeter was not used as a predictor variable because for older burns the perimeter seldom was apparent and the occurrence of unburnt patches of varying sizes could not be determined. This prevented a direct estimate of distance to seed source and its effect on the rate of recovery into the burn interior over time. Thus, we used dendrochronological methods to assess whether recovery was limited by seed availability. Adjacent to both burn-area and reference-area transects, we cut cross-sections and dated 25 Artemisia plants from each of five life-stage classes including seedling (no woody branches), J-1 (juveniles under 15 cm), J-2 (juveniles over 15 cm but with no inflorescences), mature (inflorescences on foliage) and senescent (over 50% of the foliage dead). Cross-sections were cut at the root crown, sanded using progressively finer grit until growth rings were clearly visible, and independently cross-dated by a minimum of two analysts (e.g. Ferguson 1964, Poore et al. 2009). We were able to determine pith year for 84% of samples. For the remaining 16% of samples in which pith was not present, years-to-pith were estimated using concentric rings on a transparent overlay, calibrating estimates of missing ring spacing with that of the overlay. Differences between year of inside dated ring and estimated pith were mostly less than 5 years. Although lack of recruitment would not be sufficient to infer a lack of seed availability, occurrence of recruitment is sufficient to conclude that seed was available either from a nearby seed source or from soil-seedbank that survived the fire.
At each transect location, we recorded elevation, slope, slope position (lower, middle, upper, ridge), slope curvature (concave, convex, flat) and aspect. Soil depth was measured with a 50-cm soil probe at 10-m intervals along the 100-m transect and these 10 measurements were averaged for a mean soil depth value. One soil sample was collected with an auger to a depth of 20 cm at 5-m intervals beneath Artemisia shrubs and within interspaces for a total of 20 under-shrub and 20 inter-space samples per 100-m transect. For each group, the 20 samples were mixed together and analysed for pH (Rhoades 1982), total nitrogen (McGeehan and Naylor 1988), total phosphorus (Olsen et al. 1954), % organic matter (Walkley and Black 1934), and exchangeable calcium, magnesium, sodium and potassium (Normandin et al. 1998). A folded aspect value (i.e. cosine transformation) was calculated to represent solar radiation.
GIS methods
For climate input data we used the Parameter–Elevation Regressions on Independent Slopes Model (PRISM), which is an analytical tool that uses point measurements of precipitation, temperature and other climatic factors to produce continuous, digital-grid estimates of monthly climatic parameters (available at 4-km resolution) (Daly et al. 1994). Cool-season (October–March) and warm-season (April–September) monthly precipitation values were extracted to transect locations from the PRISM dataset and summed for each of three consecutive warm seasons (year preceding fire year, year of the fire, year following the fire) and two consecutive cool seasons (year preceding fire, year following fire). The 30-year monthly means (available at 800-m resolution) were also extracted to transect locations and summed for the same months to create cool and warm season average climatic variables.
Data analysis
We used multiple linear regression to model Artemisia recovery as a function of macro-scale environmental variables including region (Great Basin, Utah Highlands and Colorado Plateau), and site-level climatic, topographic and edaphic variables (Table 1). Climatic variables included cool and warm season precipitation immediately preceding and following the fire event (Table 1). To avoid multicollinearity, we calculated Pearson correlation coefficients for the environmental predictor variables and reduced the set of variables when correlations exceeded r = 0.30. Retained predictor variables were checked for linear regression assumptions of linearity, constant residual variance and normality of residuals.
To evaluate the influence of environmental gradients on Artemisia post-fire recovery rate, we compared two different response variables. The first variable was % Artemisia recovery, defined as canopy cover from the burnt transect divided by canopy cover recorded in the unburnt control transect (n = 27) (sensu Lesica et al. 2007). This calculation standardises the recovery rate by site characteristics. Although more productive sites may recover at a faster rate, they also likely have higher unburnt vegetation cover and thus higher cover values to obtain for 100% recovery to unburnt values. The second response variable calculated was % Artemisia cover, a value independent of reference cover values. For this response variable, we were able to add nine sites (n = 36) to the analysis for burns where no suitable unburnt transects were found of similar environmental context.
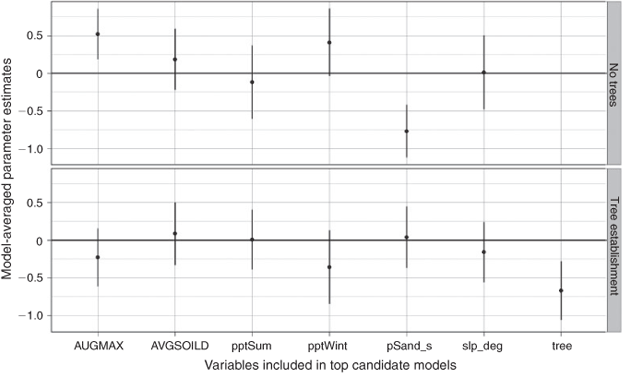
We constructed a set of candidate models for each of four distinct analyses: (1) rate of recovery for burnt sagebrush sites using the % recovery response variable; (2) rate of recovery for burnt sagebrush sites using the % cover variable; (3) sagebrush % cover for sites without evidence of previous fire (unburnt) and without trees present and (4) sagebrush % cover for unburnt sites with varying degrees of tree establishment. We used Akaike’s information criterion corrected for small sample size (AICc) to rank and weight these sets of candidate models (Akaike 1974; Burnham and Anderson 2002). We discuss top models with high AICc weights and report standardised model-averaged parameter coefficients for relative comparisons of influence between variables and between stages of post-fire stand development. All calculations were performed using R version 2.15.1 (R Development Core Team 2012).
Results
Climate and site-level environmental correlations
We examined Pearson correlation coefficients to retain a subset of relatively uncorrelated (r < 0.30) variables for inclusion in candidate models. For burnt sites (n = 36), % sand was retained and variables highly correlated with % sand were excluded from model building. These included % clay (r = –0.81), % silt (r = –0.93), Ca (r = –0.72), Mg (r = –0.71), K (r = –0.60), total N (r = –0.60), & organic matter (r = –0.35) and total P (r = –0.35). Thus, % sand serves as a proxy for relatively infertile, coarse-textured soils with low cation exchange capacity. Mean 30-year annual precipitation was strongly correlated with 30-year mean winter precipitation (pptWint) (r = 0.90), before-fire winter precipitation (W0) (r = 0.41) and after-fire winter precipitation (W1) (r = 0.59). Each of these variables was retained for inclusion in separate models to compare their relative influence when considered in tandem with soil variables and time since fire. Percentage sand fraction and winter precipitation were negatively correlated (r = –0.59) and thus were not included in the same model. Soil depth was retained as a predictor, having only marginal negative correlation with % sand (r = –0.28). Warm season precipitation variables (pptSum, G0, G1, G2) were included in models with soil variables but were not included in the same models with cool-season precipitation variables, owing to multicollinearity. August maximum temperature, rather than elevation, was retained for inclusion in models (r = 0.92).
Influences of climate variability and site environment on post-fire Artemisia recovery
We sampled 10 paired transects (burnt and reference) from the Colorado Plateau, nine from the Great Basin and eight from the Utah Highlands. Time since fire (TSF) explained 35% of the between-site variance in canopy recovery:
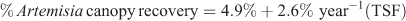
where R2 = 0.35, P = 0.002, n = 27.
Extrapolation using this equation predicts 37 years for full recovery to unburnt cover values. The 95% confidence intervals (±2.05) for the estimate of the slope parameter bracket full recovery to between 24 and 84 years.
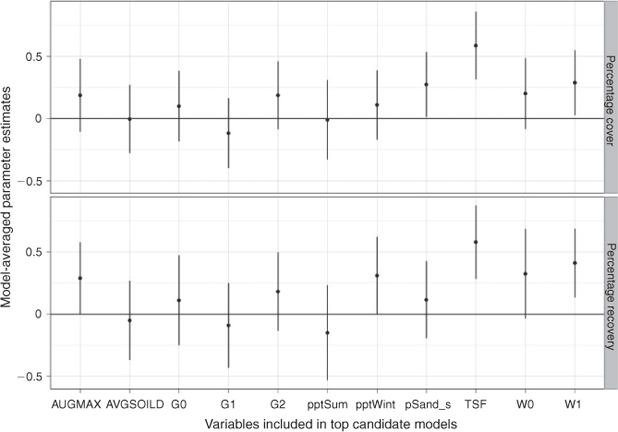
In addition to TSF, W1 positively influenced recovery (Fig. 2). Average pptWint and W0 were highly correlated with this variable but explained less of the between-site variance in absolute cover and had confidence intervals that overlapped zero.
|
Environmental controls on % Artemisia cover after fire
Similar to the recovery response variable scaled to reference values (above), time since fire explained 36% of the variance in % Artemisia cover calculated independent of the paired reference site:
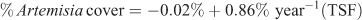
where R2 = 0.36, P < 0.001, n = 36.
Extrapolation of this equation to reach the mean % Artemisia cover for reference sites (24%) would take 27 years. The 95% confidence intervals (±0.42) for the estimate of the slope parameter bracket reaching 24% cover to between 19 and 51 years. The addition of W1 accounted for an additional 6% of the variance in Artemisia cover
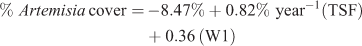
where R2 = 0.41, P < 0.0001, n = 36 (Figs 2, 3).

|
In contrast, 30-year average pptWint only accounted for an additional 1% of the variance (Fig. 2). The addition of % sand fraction in combination with W1 accounted for an additional 8% of the variance:
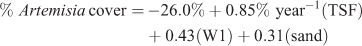
where R2 = 0.49, P < 0.001, n = 36.
The effect of sand fraction should be interpreted with the knowledge that range of sand fraction for these sites spanned only 24 to 71%. Beyond these minimum and maximum values for this study, the relationship is unknown.
Environmental relationships with Artemisia cover in unburnt stands
In treeless plots of apparently unburnt Artemisia stands (n = 18), % sand fraction was negatively associated with Artemisia cover, explaining 37% of the variance:
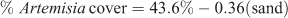
where R2 = 0.37, P = 0.006, n = 18 (Fig. 4).
|
The addition of August maximum temperature explained another 25% of the variance, with warmer sites having greater Artemisia cover:
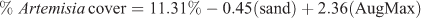
where R2 = 0.62, P < 0.001, n = 18.
In locations where trees had established within Artemisia communities (tree cover range 1–35%), the presence of trees was negatively associated with Artemisia cover and explained 48% of the variance:
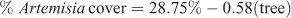
where R2 = 0.48, P < 0.001, n = 18 (Fig. 4).
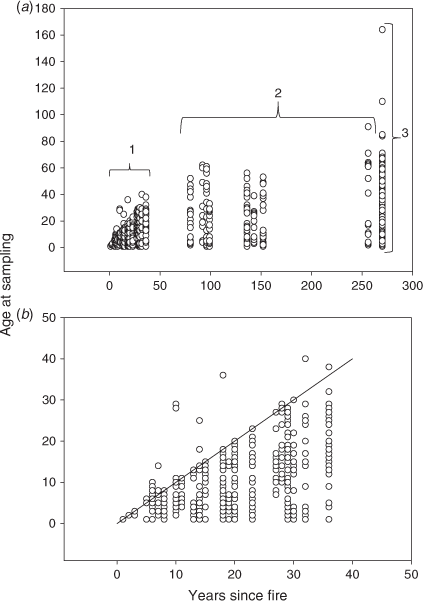
Age distributions
We sampled and dendrochronologically dated 1455 individual Artemisia plants from 72 transects (Fig. 5a). All transects, located at sites that had burnt less than 40 years before sampling (n = 36) in 2007 and 2008, had established Artemisia cohorts within 7 years after the fire (Fig. 5b). Most sites (30 of 36) had cohorts within the first 3 years after fire – including seven sites where individuals were found to have established before the year of the fire, presumably surviving within small unburnt islands near the transect location.
|
Discussion
Post-fire Artemisia regeneration and climate
Plant establishment in semi-arid ecosystems is limited by water availability, which varies spatially as a function of topography and elevation. However, temporal variability (fluctuating resources) may be more important than mean resource availability (Katz and Brown 1992) for controlling the timing and rate of plant recruitment in water-limited ecosystems (Schwinning et al. 2004). The regeneration niche is typically a narrower subset of the conditions than those tolerated by adults, due to factors such as incomplete development of root systems in juveniles and thus reduced access to deep soil water (Grubb 1977). Where populations persist at or near the juvenile survival threshold, post-disturbance recruitment may hinge on whether precipitation in a given year is above or below average, resulting in only ephemeral windows of opportunity for regeneration.
Results of this study suggest that precipitation in the winter following fire, a highly stochastic variable, is of primary importance for defining the ephemeral, post-burn regeneration niche of Artemisia, and hence for quantifying post-fire recovery of Artemisia. Artemisia seeds lack significant carbohydrate reserves (Welch 2005), so water necessary for plant growth and root elongation after germination has a direct influence on probability of surviving summer desiccation. Additionally, recovery of sagebrush was positively correlated with soils with higher sand concentration. The mechanism underlying this effect could be related to seedbank effects or alternatively to competition. Sandier soils may result in greater seed retention and slightly deeper seed burial, which can increase water availability (Chambers 2000). Another mechanism underlying this pattern could be that higher sand fraction insulates seedbanks from fire, improving seedbank survival and potential for establishment; seeds may also be more prone to burial in sandier sites, a process that may be necessary to initiate dormancy until the spring after the fire (Wijayratne and Pyke 2012). Sandier sites also allow deeper infiltration during small to medium precipitation events, possibly providing a competitive advantage to seedlings of deep-rooting species relative to shallow-rooted herbaceous species. Another mechanism might be sparse cover of perennial herbaceous species following fire and more open micro-sites, which promotes sagebrush establishment due to more light and less competition. Negative associations between Artemisia cover and percentage sand fraction in unburnt or fully recovered sites suggest that a positive influence of coarse-textured soils on recovery rate may ultimately be counter-balanced by reduced productivity on these drier and more nutrient-limited sites.
Although we found strong evidence for an effect of the timing of precipitation on post-fire Artemisia recovery, utilising a chrono-sequence approach is correlational in nature and thus we could not control for the effects of unmeasured factors that may also have been important in the processes we studied. Despite this shortcoming, the controlled experimental alternative to a study such as this is infeasible given the long time period that is required to quantify successional processes. The strength of this type of study is to highlight environmental predictor variables that should be considered for further study and in management applications that may benefit from exploratory studies.
Broad-scale variation in patterns of sagebrush dominance during succession
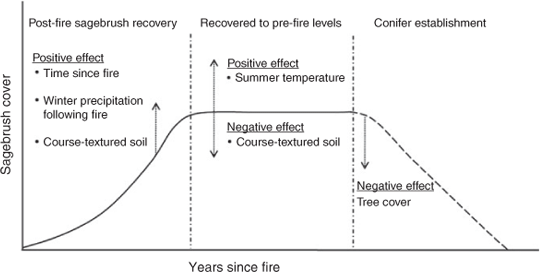
Post-fire sagebrush recovery was positively related to cool-season precipitation and course-textured soil up to 72% sand (Figs 2, 6), but sagebrush cover in unburnt stands was negatively related to percentage sand and positively related to average August maximum temperature (Figs 4, 6). As tree establishment and dominance occurred, sagebrush cover decreased, presumably due to shading and other unmeasured competitive effects (Figs 4, 6). Through the processes of fire and succession, sagebrush abundance can follow a humped-shaped distribution over time where populations increase after fire through seed dispersal and establishment and then decline in the potential advent of conifer colonisation and subsequent establishment and in-filling (Fig. 6). Favourable weather, a stochastic variable, limits establishment of sagebrush and conifer in addition to the spatial proximity of a propagule source.
|
In contrast to our findings, Maier et al. (2001) found recruitment of ssp. vaseyana to be correlated with lower rather than higher than average precipitation in the months of February, April and June. They suggest that higher temperatures possibly associated with the observed lower precipitation in these months, may have favoured recruitment (McDonough and Harniss 1974; Cawker 1980; Young et al. 1991). In colder sites at higher latitudes, such as those in the study of Maier et al. (2001), where plant establishment is limited by spring temperatures and growing season length, years of below-average precipitation and above-average temperatures may enhance recruitment given that overall moisture availability is sufficient for establishment. In contrast, in drier sites at lower latitudes, which are more limited by water availability, higher precipitation may enhance recruitment. This pattern should also be observed along an elevation gradient. In evaluating the lower-elevation spp. wyomingensis, Maier et al. (2001) found that recruitment was positively associated with wetter winters, suggesting either or both thermal insulation from snow cover and deep soil-moisture recharge from winter precipitation could have favoured recruitment in these drier locations. Increased winter precipitation was correlated with recovery at the spp. vaseyana sites we studied. Because spp. vaseyana occurs over a large altitudinal and latitudinal gradient, it is logical that the regeneration niche is limited by moisture in drier, lower elevation or southern latitude sites, but is limited by growing season length and less limited by moisture at higher elevations and more northerly latitudes.
‘Fast-track’ and ‘slow-track’ Artemisia recovery, or a continuum?
Baker (2011) conducted a meta-analysis on data from several studies of post-fire recovery of ssp. vaseyana conducted in Montana, north-west Wyoming, and south-west and east Idaho. The combined data suggested qualitative distinctions between ‘fast track’ and ‘slow track’ post-fire recovery trajectories for Artemisia communities. Sixteen sites exhibited ‘fast-track’ behaviour and had reestablished Artemisia to unburnt levels within 25–35 years. In contrast, 40 ‘slow-track’ sites were on pace to recover in 75 years or more, based on extrapolation from observed trajectories. Baker (2011) delineated a line separating the qualitatively different recovery tracks starting at the origin with a slope of ~1.7% recovery year–1. An intermediate space with no data points could be bracketed by lines through the origin with slopes of 1.28–2.28% recovery year–1, although Baker notes that a continuum of recovery rates is also likely. In support of Baker’s continuum hypothesis, our results show a continuum of recovery responses rather than two different tracks. Considering only the 27 burnt sites paired with an unburnt reference, eight sites fall into the slow track, six into the intermediate range and thirteen into the fast track group.
The studies reviewed by Baker (2011) were conducted on the Snake River plain and Wyoming Basin (West 1983), north of the polar front gradient; a regional climate switching point (Miller and Eddleman 2001). South of the switching point, temperatures are generally warmer with more precipitation in the summer and less in the winter (Mitchell 1976; Neilson 1986). Our sites were mainly winter-precipitation dominated but trended towards higher summer precipitation to the southern and eastern portions of our study region. Although latitude, longitude or direct estimates of summer precipitation did not explain a significant portion of the variance in recovery, small sample size limited our power to disentangle the multiple dimensions of climate, soils, region and time. The use of geographic region as an environmental explanatory variable is a surrogate for climate and other unmeasured factors, whereas estimates of precipitation and temperature derived from PRISM provide specific climatic components that could be conceptually linked to mechanisms influencing the recovery process. In a climate change context, there may be reason to expect regional systematic differences in how the regeneration niche is redistributed in time and space, but ultimately these expectations are based on mechanisms linking physiologically based life history traits with climate and soil conditions. Promising avenues of research might couple climate change projections with soil water balance models to simulate the regeneration niche under different scenarios (Schlaepfer et al. 2011). However, thus far the resolution of such models is coarse and of limited applied value for land management. Average temperatures may warm by as much as 5°C by the end of the century for the Great Basin and much of the adjacent area that supports Artemisia (Bachelet et al. 2001; Neilson et al. 2005), a situation that may increase water stress at lower elevations and perhaps decrease cold temperature limitations at higher elevations. The influence of fire-adapted invasive species (Bradley 2010) and fire in these ecosystems may increase as well (Westerling et al. 2006). Further investigation into how the regeneration niche might be redistributed at topographic and regional scales will be important for understanding Artemisia ecosystem dynamics in the context of future climate change.
Management implications
Results indicate that recovery of mountain big sagebrush is most dependent upon favourable weather patterns, particularly winter precipitation, in the year after fire. Thus, burn areas prone to drought (e.g. south or west aspect, low elevation, thin soils) may be at higher risk of delayed recovery than more mesic sites. Insufficient residual seed in the seedbank may also be important, especially where fire intensity was high. Managers can use this knowledge on a case-by-case basis to assess risks to sagebrush recovery and to inform decisions regarding the potential need to artificially seed sagebrush on burnt landscapes. In cases where observed post-fire establishment is judged to be inadequate 2–3 years after fire, recovery may be enhanced by augmenting depleted natural seed reserves using appropriate artificial seeding strategies.
Acknowledgements
We thank Ben Bright, Brian Reeves, Trent Young, Miles Nielson, Todd Granberry and Stephanie Carlson for their dedication to field work, sample processing and data management. Special thanks to Sherel Goodrich, Linda Chappell, Covy Jones and Steve Strong for assistance in site selection and developing site protocols, and to Tod Williams and Dave Stobart for assistance in obtaining permits to conduct the research inside State and National Park boundaries. This study was supported by the Joint Fire Science Program, grant number 06–3-1–17, and is based in part upon work supported by the National Science Foundation under grant number EPS-0814372. For helpful comments on various versions of this article, we express great thanks to Jeanne Chambers, William Longland and two anonymous reviewers.
References
Akaike H (1974) A new look at the statistical model identification. IEEE Transactions on Automatic Control 19, 716–723.| A new look at the statistical model identification.Crossref | GoogleScholarGoogle Scholar |
Bachelet D, Neilson RP, Lenihan JM, Drapek RJ (2001) Climate change effects on vegetation distribution and carbon budget in the United States. Ecosystems 4, 164–185.
| Climate change effects on vegetation distribution and carbon budget in the United States.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXltVKksL4%3D&md5=cf3861627f27ad128a71f6ed21fa89cbCAS |
Baker WL (2006) Fire and restoration of sagebrush ecosystems. Wildlife Society Bulletin 34, 177–185.
| Fire and restoration of sagebrush ecosystems.Crossref | GoogleScholarGoogle Scholar |
Baker WL (2011) Pre-EuroAmerican and recent fire in sagebrush ecosystems. In ‘Greater Sage-Grouse: Ecology and Conservation of a Landscape Species and its Habitats’, Studies in Avian Biology Series, Vol. 38. (Eds ST Knick, JW Connelly) pp. 185–201. (University of California Press: Berkeley, CA)
Blaisdell JP (1953). Ecological effects of planned burning of sagebrush-grass range on the Upper Snake River Plains. United States Department of Agriculture, Technical Bulletin 1075, 39. (Washington, DC)
Bonham CD, Cottrell TR, Mitchell JE (1991) Inferences for life history strategies of Artemisia tridentata subspecies. Journal of Vegetation Science 2, 339–344.
| Inferences for life history strategies of Artemisia tridentata subspecies.Crossref | GoogleScholarGoogle Scholar |
Boyd CS, Bates JD, Miller RF (2007) The influence of gap size on sagebrush cover estimates with the use of line intercept technique. Rangeland Ecology and Management 60, 199–202.
| The influence of gap size on sagebrush cover estimates with the use of line intercept technique.Crossref | GoogleScholarGoogle Scholar |
Bradley BA (2010) Assessing ecosystem threats from global and regional change: hierarchical modeling of risk to sagebrush ecosystems from climate change, land use and invasive species in Nevada, USA. Ecography 33, 198–208.
| Assessing ecosystem threats from global and regional change: hierarchical modeling of risk to sagebrush ecosystems from climate change, land use and invasive species in Nevada, USA.Crossref | GoogleScholarGoogle Scholar |
Burke IC, Reiners WA, Olson RK (1989) Topographic control of vegetation in a mountain big sagebrush steppe. Plant Ecology 84, 77–86.
| Topographic control of vegetation in a mountain big sagebrush steppe.Crossref | GoogleScholarGoogle Scholar |
Burnham KP, Anderson DR (2002) ‘Model Selection and Multimodel Inference: a Practical Information–Theoretic Approach’, 2nd edn (Springer–Verlag: New York)
Cawker KB (1980) Evidence of climatic control from population age structure of Artemisia tridentata Nutt. in southern British Columbia. Journal of Biogeography 7, 237–248.
| Evidence of climatic control from population age structure of Artemisia tridentata Nutt. in southern British Columbia.Crossref | GoogleScholarGoogle Scholar |
Chambers JC (2000) Seed movements and seedling fates in disturbed sagebrush steppe ecosystems: implications for restoration. Ecological Applications 10, 1400–1413.
Daly C, Neilson RP, Phillips DL (1994) A statistical–topographic model for mapping climatological precipitation over mountainous terrain. Journal of Applied Meteorology 33, 140–158.
| A statistical–topographic model for mapping climatological precipitation over mountainous terrain.Crossref | GoogleScholarGoogle Scholar |
Daubenmire R (1975) Ecology of Artemisia tridentata subsp. tridentata in the state of Washington. Northwest Science 49, 24–35.
Evans RD, Black RA, Link SO (1991) Reproductive growth during drought in Artemisia tridentata Nutt. Functional Ecology 5, 676–683.
| Reproductive growth during drought in Artemisia tridentata Nutt.Crossref | GoogleScholarGoogle Scholar |
Ferguson CW (1964) Annual rings in big sagebrush, Artemisia tridentata. In ‘Papers of the Laboratory of Tree-Ring Research, No. 1’. (University of Arizona Press: Tucson, AZ)
Goodwin DL (1956) Autecological studies of Artemisia tridentata Nutt. PhD thesis, State College of Washington.
Grime JP (1974) Vegetation classification by reference to strategies. Nature 250, 26–31.
| Vegetation classification by reference to strategies.Crossref | GoogleScholarGoogle Scholar |
Grubb PJ (1977) The maintenance of species-richness in plant communities: the importance of the regeneration niche. Biological Reviews of the Cambridge Philosophical Society 52, 107–145.
| The maintenance of species-richness in plant communities: the importance of the regeneration niche.Crossref | GoogleScholarGoogle Scholar |
Johnson JR, Payne GF (1968) Sagebrush reinvasion as affected by some environmental influences. Journal of Range Management 21, 209–213.
| Sagebrush reinvasion as affected by some environmental influences.Crossref | GoogleScholarGoogle Scholar |
Katz RW, Brown BG (1992) Extreme events in a changing climate: variability is more important than averages. Climatic Change 21, 289–302.
| Extreme events in a changing climate: variability is more important than averages.Crossref | GoogleScholarGoogle Scholar |
Lesica P, Cooper SV, Kudray G (2007) Recovery of big sagebrush following fire in southwest Montana. Rangeland Ecology and Management 60, 261–269.
| Recovery of big sagebrush following fire in southwest Montana.Crossref | GoogleScholarGoogle Scholar |
Maier AM, Perryman BL, Olson RA, Hild AL (2001) Climatic influences on recruitment of 3 subspecies of Artemisia tridentata. Journal of Range Management 54, 699–703.
| Climatic influences on recruitment of 3 subspecies of Artemisia tridentata.Crossref | GoogleScholarGoogle Scholar |
McArthur ED, Ott JE (1996) Potential natural vegetation in the 17 conterminous western United States. In ‘Proceedings: Shrubland Ecosystem Dynamics in a Changing Environment’, 23–25 May 1995, Las Cruces, NM. (Eds JR Barrow, ED McArthur, RE Sosebee, RJ Tausch) USDA Forest Service, Intermountain Research Station, General Technical Report INT-GTR-338, pp. 16–28. (Ogden, UT)
McDonough WT, Harniss RO (1974) Effects of temperature on germination in three subspecies of big sagebrush. Journal of Range Management 27, 204–205.
| Effects of temperature on germination in three subspecies of big sagebrush.Crossref | GoogleScholarGoogle Scholar |
McGeehan SL, Naylor DV (1988) Automated instrumental analysis of carbon and nitrogen in plant and soil samples. Communications in Soil Science and Plant Analysis 19, 493–505.
| Automated instrumental analysis of carbon and nitrogen in plant and soil samples.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL1cXitFygur0%3D&md5=8524a9c14c5f8da98e1a0748362fbd45CAS |
Meyer SE (1994) Germination and establishment ecology of big sagebrush: implications for community restoration. In ‘Proceedings – Ecology and Management,of Annual Rangelands’, 18–21 May 1992, Boise, ID. (Eds SB Monsen, SG Kitchen) USDA Forest Service, Intermountain Research Station, General Technical Report INT-GTR-313, pp. 244–251. (Ogden, UT)
Meyer SE, Monsen SB (1992) Big sagebrush germination patterns: subspecies and population differences. Journal of Range Management 45, 87–93.
| Big sagebrush germination patterns: subspecies and population differences.Crossref | GoogleScholarGoogle Scholar |
Meyer SE, Monsen SB, McArthur ED (1990) Germination response of Artemisia tridentata (Asteraceae) to light and chill: patterns of between-population variation. Botanical Gazette 151, 176–183.
| Germination response of Artemisia tridentata (Asteraceae) to light and chill: patterns of between-population variation.Crossref | GoogleScholarGoogle Scholar |
Miller RF, Eddleman L (2001) Spatial and temporal changes of sage grouse habitat in the sagebrush biome. Oregon State University, Agricultural Experiment Station, Technical Bulletin 151. (Corvallis, OR)
Mitchell VL (1976) The regionalization of climate in the Western United States. Journal of Applied Meteorology 15, 920–927.
| The regionalization of climate in the Western United States.Crossref | GoogleScholarGoogle Scholar |
Neilson RP (1986) High-resolution climatic analysis and southwest biogeography. Science 232, 27–34.
| High-resolution climatic analysis and southwest biogeography.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3cvivVGisQ%3D%3D&md5=6247bb2c0d708ea7d9ef1769ded25e8cCAS | 17773996PubMed |
Neilson RP, Pitelka LF, Solomon AM, Nathan R, Midgley GF, Fragoso J, Lischke H, Thompson K (2005) Forecasting regional to global plant migration in response to climate change. Bioscience 55, 749–759.
| Forecasting regional to global plant migration in response to climate change.Crossref | GoogleScholarGoogle Scholar |
Normandin V, Kotuby-Amacher J, Miller RO (1998) Modification of the ammonium acetate extractant for the determination of exchangeable cations in calcareous soils. Communications in Soil Science and Plant Analysis 29, 1785–1791.
| Modification of the ammonium acetate extractant for the determination of exchangeable cations in calcareous soils.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXlvVSlsLs%3D&md5=9ed3132b83a163723675275ca42eac05CAS |
Olsen SR, Cole CV, Watanabe FS, Dean LA (1954) Estimation of available phosphorus in soils by extraction with sodium bicarbonate. USDA Circular 939, 1–19.
| Estimation of available phosphorus in soils by extraction with sodium bicarbonate.Crossref | GoogleScholarGoogle Scholar |
Perfors T, Harte J, Alter SE (2003) Enhanced growth of sagebrush (Artemisia tridentata) in response to manipulated ecosystem warming. Global Change Biology 9, 736–742.
| Enhanced growth of sagebrush (Artemisia tridentata) in response to manipulated ecosystem warming.Crossref | GoogleScholarGoogle Scholar |
Poore RE, Lamanna CA, Ebersole JJ, Enquist BJ (2009) Controls on radial growth of mountain big sagebrush and implications for climate change. Western North American Naturalist 69, 556–562.
| Controls on radial growth of mountain big sagebrush and implications for climate change.Crossref | GoogleScholarGoogle Scholar |
R Development Core Team (2012) R: a language and environment for statistical computing. (R Foundation for Statistical Computing: Vienna Austria) Available at http://www.R-project.org [Verified 7 September 2013]
Rehfeldt GE, Crookston NL, Warwell MV, Evans JS (2006) Empirical analyses of plant-climate relationships for the western United States. International Journal of Plant Sciences 167, 1123–1150.
| Empirical analyses of plant-climate relationships for the western United States.Crossref | GoogleScholarGoogle Scholar |
Rhoades JD (1982) Soluble salts. In ‘Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties’, Monograph Number 9, 2nd edn. (Eds AL Page, RH Miller, DR Keeney) pp. 167–179. (ASA: Madison, WI)
Schlaepfer DR, Lauenroth WK, Bradford JB (2011) Effects of ecohydrological variables on current and future ranges, local suitability patterns, and model accuracy in big sagebrush. Ecography 4, 374–384.
Schwinning S, Sala OE, Loik ME, Ehleringer JR (2004) Thresholds, memory, and seasonality: understanding pulse dynamics in arid/semi-arid ecosystems. Oecologia 141, 191–193.
| Thresholds, memory, and seasonality: understanding pulse dynamics in arid/semi-arid ecosystems.Crossref | GoogleScholarGoogle Scholar | 15300489PubMed |
Stokes MS, Smiley TL (1968) ‘An Introduction to Tree-Ring Dating.’ (University of Chicago Press: Chicago, IL)
Walkley A, Black IA (1934) An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Science 37, 29–38.
| An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaA2cXitlGmug%3D%3D&md5=c5cfb717fd9b3948312456f82ade74ffCAS |
Welch BL (2005) Big sagebrush: a sea fragmented into lakes, ponds, and puddles. USDA Forest Service, Rocky Mountain Research Station, General Technical Report RMRS-GTR-144. (Fort Collins, CO)
West NE (1983) Western intermountain sagebrush steppe. In ‘Ecosystems of the World. 5. Temperate deserts and semi-deserts’. (Ed. NE West) pp. 351–397 (Elsevier Scientific Publishing Co: New York)
Westerling AL, Hidalgo HG, Cayan DR, Swetnam TW (2006) Warming and earlier spring increase western US forest wildfire activity. Science 313, 940–943.
| Warming and earlier spring increase western US forest wildfire activity.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XotFCitbo%3D&md5=308417c344ec067b31725c73724340afCAS | 16825536PubMed |
Wijayratne UC, Pyke DA (2012) Burial increases seed longevity of two Artemisia tridentata (Asteraceae) subspecies. American Journal of Botany 99, 438–447.
| Burial increases seed longevity of two Artemisia tridentata (Asteraceae) subspecies.Crossref | GoogleScholarGoogle Scholar | 22301894PubMed |
Young JA, Evans RA (1989) Dispersal and germination of big sagebrush (Artemisia tridentata) seeds. Weed Science 37, 201–206.
Young JA, Palmquist DE, Evans RA (1991) Temperature profiles for germination of big sagebrush seeds from native stands. Journal of Range Management 44, 385–390.
| Temperature profiles for germination of big sagebrush seeds from native stands.Crossref | GoogleScholarGoogle Scholar |
Ziegenhagen LL, Miller RF (2009) Postfire recovery of two shrubs in the interiors of large burns in the Intermountain West, USA. Western North American Naturalist 69, 195–205.
| Postfire recovery of two shrubs in the interiors of large burns in the Intermountain West, USA.Crossref | GoogleScholarGoogle Scholar |