

Potential nutritional strategies for the amelioration or prevention of high rigor temperature in cattle – a review
Kristy DiGiacomo A , Brian J. Leury A and Frank R. Dunshea A BA Melbourne School of Land and Environment, The University of Melbourne, Parkville, Vic, 3010, Australia.
B Corresponding author. Email: fdunshea@unimelb.edu.au
Animal Production Science 54(4) 430-443 https://doi.org/10.1071/AN13303
Submitted: 19 July 2013 Accepted: 17 January 2014 Published: 25 February 2014
Journal Compilation © CSIRO Publishing 2014 Open Access CC BY-NC-ND
Abstract
Environmental conditions influence animal production from an animal performance perspective and at the carcass level post-slaughter. High rigor temperature occurs when the animal is hyperthermic pre-slaughter, and this leads to tougher meat. Hyperthermia can result from increased environmental temperature, exercise, stress or a combination of these factors. Consumer satisfaction with beef meat is influenced by the visual and sensory traits of the product when raw and cooked, with beef consumers commonly selecting tenderness of the product as the most important quality trait. High rigor temperature leads to a reduction in carcass and eating quality. This review examines some possible metabolic causes of hyperthermia, with focus on the importance of adipose tissue metabolism and the roles of insulin and leptin. Potential strategies for the amelioration or prevention of high rigor temperature are offered, including the use of dietary supplements such as betaine and chromium, anti-diabetic agents such as thiazolidinediones, vitamin D, and magnesium (Mg) to provide stress relief.
Additional keywords: adipose, fat, heat stress, insulin resistance, leptin, supplementation, TZDs.
Introduction
Farming and abattoir practices need to adapt to climate change, to prevent or reduce the impact of high environmental temperature on meat quality due to processes such as high rigor temperature (Gregory 2010). In Australia, the 75% of beef carcasses have high rigor temperature (Warner et al. 2014a), which shows the importance of finding methods to reduce the incidence. One of the main causes of high rigor temperature appears to be a combination of rapid glycolysis and relatively slow cooling, which can lead to high rigor temperatures and potential toughening (Jacob and Hopkins 2014). Moreover, high rigor temperature may be exacerbated by loss of proteolytic potential due to inactivation of proteolytic enzymes (Kim et al. 2014; Warner et al. 2014a). The mechanisms of high rigor temperature in beef carcasses and the effects on meat quality are reviewed in Kim et al. (2014) and Jacob and Hopkins (2014) in this issue of Animal Production Science. Factors that contribute to elevated body temperature pre-slaughter potentially predispose animals to higher carcass temperature immediately post slaughter (Jacob and Hopkins 2014).
There is mounting evidence that a state of insulin resistance can compromise the ability of an individual to thermoregulate. For example, literature related to rodents and humans suggests that stressors such as infection, inflammation, ageing, heat stress and excessive adipose tissue deposition can all modulate insulin action (Tsiotra and Tsigos 2006). Since all of these stressors are also encountered by beef cattle, we hypothesise that the responses in cattle may be similar. In particular, as cattle fatten and develop intramuscular fat and marbling, which is morphologically and metabolically similar to visceral fat in humans, the responses to stressors may resemble those seen in diabetic humans. In this context, intramuscular fat accumulation has been found to be associated with insulin resistance in humans (Goodpaster et al. 1997; Kim et al. 2006; Komiya et al. 2006). We hypothesise that there are links between insulin resistance, stress and thermoregulation in cattle, and possible links to high rigor temperature.
Effect of age, weight and fatness on plasma metabolites and hormones
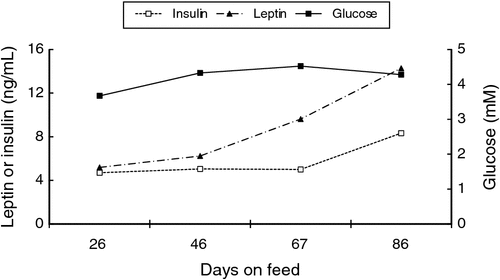
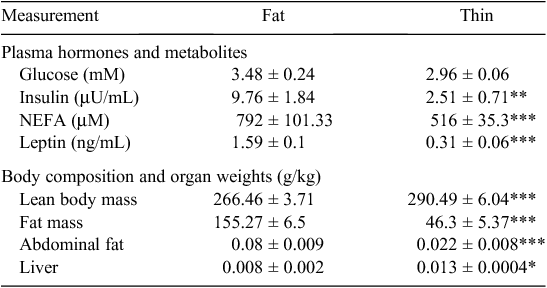
Insulin resistance is defined as a reduced ability to respond to the effects of insulin, especially by peripheral tissue such as muscle and adipose tissues. Since cells must have glucose to survive, the body compensates for insulin resistance by producing additional insulin, resulting in hyperinsulinemia and eventually hyperglycemia. In cattle, blood glucose increases with age (Lorenz 2000), while fasting plasma insulin is positively correlated with liveweight (McCann et al. 1986). When grazing beef cattle are introduced to concentrate feeding in a feedlot, the response is exacerbated and there is an increase in circulating glucose and insulin with time on feed (Fig. 1) (Henry et al. 2000; Hersom et al. 2004c), which is also associated with increasing fatness and adipocyte cellularity (Hersom et al. 2004a, 2004b, 2004c). Feeding of grain is also associated with an increase in core body temperature (Jacob et al. 2014). Fat sheep and cattle have higher plasma concentrations of glucose, insulin and non-esterified fatty acids (NEFA) than their thin counterparts (Table 1) (McCann et al. 1989; McCann et al. 1997; Henry et al. 2000). Moderately fat sheep and cattle exhibit an exaggerated insulin response to intravenous glucose injection (McCann et al. 1989; McCann et al. 1986). Although these differences are most exaggerated in obese ruminants, they also occur when animals grow older, heavier and fatter. For example, Eisemann et al. (1997) found that insulin responsiveness and sensitivity of the hindquarter of beef steers decreased with increasing age and liveweight (275 v. 490 kg), indicating insulin resistance of peripheral tissues as steers get heavier and fatter. Metabolism across the hindlimb or hindquarter is generally considered to reflect skeletal muscle metabolism, although these tissue beds also contain adipose tissue (Boisclair et al. 1993, 1994; Dunshea et al. 1995). Together these data suggest that long periods of grain feeding would increase fat deposition and elevate circulating glucose and insulin, presumably by inducing a state of insulin resistance. This was demonstrated recently, where animals with a higher blood insulin concentration at slaughter had a higher incidence of high rigor temperature (Warner et al. 2014a).
|
|
Another hormone associated with increased body fatness is leptin, a hormone produced by adipocytes that can signal the brain as a satiety factor; and mutations in the leptin gene or in the leptin receptor can lead to obesity (see review by Zieba et al. 2005). Expression of the leptin gene in adipose tissue and circulating leptin concentrations are highly correlated with percentage of body fat in rodents (Soukas et al. 2000) and the degree of obesity in humans (Auwerx and Staels 1998). The concentration of leptin is directly related to the degree of adiposity in cattle (Ehrhardt et al. 2000; Hersom et al. 2004c) and sheep Chilliard et al. 2005). Long-fed cattle and obese sheep tend to reduce their feed intake as they become fatter and leptin may act as a satiety factor to inhibit feed intake in fat animals that have accumulated adipose tissue. In this context, central leptin infusion decreases feed intake in ad libitum fed feed but not in sheep on a restricted feed intake (Henry et al. 1999, 2004).
There is a complex interplay between plasma insulin and leptin. Insulin stimulates plasma leptin in monogastrics such as rodents and humans (Ahima and Flier 2000; Considine 2001), and probably ruminants (Ingvartsen and Boisclair 2001). Plasma leptin appears positively related to plasma glucose and insulin and negatively related to plasma NEFA and somatotropin in periparturient dairy cows (Block et al. 2001). Also, Leury et al. (2003) found that insulin infusion increased plasma leptin during late pregnancy when cows were in positive energy balance but had little effect during early lactation when cows were in negative energy balance. However, central administration of leptin does not alter plasma insulin (Henry et al. 1999), except perhaps in previously well-fed sheep that were fasted for 72 h, in which insulin decreased (Henry et al. 2004). Since finishing beef cows have a highly positive energy balance, the increased plasma leptin could be due to a combination of increased body fatness and increased insulin secretion. In high residual feed intake (RFI) (inefficient) steers, plasma insulin concentrations tend to be higher, perhaps indicating that inefficient steers are less sensitive to the actions of insulin (Richardson et al. 2004).
Therefore, it is reasonable to suppose that as long-fed cattle accumulate subcutaneous, visceral and intramuscular fat, they also become insulin resistant. As a result, plasma insulin, glucose and leptin increase and animals reduce their ability to cope with stressors such as infection, inflammation and heat stress. Although the focus here has been on adipose tissue production of leptin, it is also possible that several other adipose-tissue-derived factors such as adiponectin, tumour necrosis factor α (TNFα) and resistin may also be involved in the induction of insulin resistance in long fed cattle (Komatsu et al. 2005; Roh et al. 2006).
Heat stress reduces the in vitro expression of leptin and its receptor mRNA in bovine peripheral blood mononuclear cells (Lacetera et al. 2009), although the concentrations of leptin and its receptors were increased when lactating dairy cattle were exposed to longer daylengths in vivo (Bernabucci et al. 2006), similar to those seen in summer months. It is therefore suggested that during heat exposure, the secretion of leptin from adipose tissue is enhanced while the secretion of leptin from other tissues, which contribute less to the overall pool of leptin, may be reduced in a homeostatic mechanism that assists to attenuate less critical processes such as immunity (Lacetera et al. 2009).
Insulin resistance and heat tolerance
In addition to the effects that insulin resistance has on tissue carbohydrate metabolism, increasing evidence suggests that diabetic individuals suffer from thermal intolerance, exhibiting an inability to control body temperature (Ohtsuka et al. 1995). In part this is because skin blood flow and skin thickness are reduced in diabetic individuals (Forst et al. 2006), thereby reducing the ability to thermoregulate. In this context, chronic treatment with the insulin sensitiser rosiglitazone increases skin blood flow and improves the ability to thermoregulate in diabetic individuals (Petrofsky et al. 2005).
The fundamental thermoregulatory strategy of any mammal, including beef cattle, is to maintain a body core temperature higher than ambient to allow heat to dissipate via conduction, convection, radiation and evaporation (Collier et al. 2006). Conduction, convection and radiation are referred to as sensible routes of heat loss and they require a thermal gradient to operate. On the other hand, evaporation works on a vapour/pressure gradient, and it is defined as insensible heat loss. When ambient temperature conditions approach body temperature, the only viable route of heat loss is evaporation; if ambient conditions exceed body temperature, heat flow will reverse and the animal becomes a heat sink. In order to assist evaporation from the skin during heat exposure, and thereby dissipate heat, there is an increase in blood flow to, and sweating from, the skin of cattle and other ruminants (Choshniak et al. 1982). Merino sheep, which are more heat-tolerant than British sheep breeds, appear to exhibit greater insulin sensitivity, at least with respect to fat metabolism, than British breeds (Ponnampalam et al. 2005). Interestingly, such differences exist between Merino sheep selected for high and low greasy fleece weights (G. M. Butler, F. R. Dunshea and B. J. Leury, unpubl. data).
One link between insulin resistance, skin blood flow and thermoregulation may be via nitric oxide synthase (NOS). Chronic diabetes, acute hyperglycaemia and NOS inhibition by Nω-nitro-L-argine (L-NNA) all lower heat tolerance in rats (Swiecki et al. 2003). Those authors suggest that, in part, this may be due to the inability to produce heat shock proteins (HSP), as the control rats had greater hepatic and intestinal expression of HSP-72 when subjected to heat stress than the heat-treated diabetic, hyperglycaemic or L-NNA-treated rats. The regulation of blood flow to the skin may be mediated via the central nervous system, at least in part regulated by an isoform of NOS, specifically neuronal NOS (nNOS) (Kellogg et al. 2008). Kellogg et al. (2008) examined the effects of the specific nNOS inhibitor 7-nitro indazole (7-NI), administered by intradermal microdialysis, on vasodilation induced by whole-body heat stress or local skin warming and found that nNOS inhibition reduced the increases skin blood flow during whole body heat stress but not during local skin warming. The differences between the effects of different isoforms of NOS, as well as the differing responses to systemic and local hyperthermia, highlight how complex the vasodilatory response can be (Cals-Grierson and Ormerod 2004). However, muscle nNOS is reduced in individuals that are insulin-resistant (Bradley et al. 2007) but there are few data on skin nNOS activity or contents.
In lactating cattle, heat stress reduced feed intake and increased basal and glucose-stimulated insulin concentrations, while adipose tissue reserves were not mobilised, which is likely a response that allows animals to acclimate to heat, although this is yet to be elucidated (Achmadi et al. 1993; Rhoads et al. 2009; Shwartz et al. 2009; Wheelock et al. 2010). We recently reviewed these responses in lactating dairy cattle in depth (Dunshea et al. 2013), and thus the responses in lactating cattle will not be covered here. Prolactin modulates sweat gland function (Alamer 2011) and is involved in maintaining electrolyte and water balance in the heat-stressed mammalian gut, kidney and mammary gland (Collier et al. 1982; Faichney and Barry 1986). Prolactin increases insulin secretion (Bole-Feysot et al. 1998) while also reducing the number of insulin receptors in adipose tissue (McNamara 1991). Therefore, the increase in plasma prolactin concentrations has a multifaceted influence on the heat-stressed mammal and may be partially responsible for heat-associated insulin resistance. Prolactin may also influence metabolism independent of physiological responses (such as respiration rate) to heat, as we recently demonstrated in sheep exposed to a mild level of heat (~36°C for 6–7 h daily) (DiGiacomo 2011).
Leptin and thermogenesis
In rodents, leptin regulates thermogenesis in brown adipose tissue (BAT) directly and indirectly by controlling sympathetic outflow from the brain (Lowell and Spiegelman 2000). In chronically heat-stressed mice, adipose tissue leptin mRNA expression was significantly increased, which was paralleled by an increase in circulating plasma leptin despite the concurrent decrease in feed intake; perhaps indicating that heat treatment itself influences leptin (Morera et al. 2012).
In rats, catabolic states such as starvation (Sivitz et al. 1999) and lactation (Xiao et al. 2004) reduce uncoupling protein 1 (UCP 1) mRNA expression in BAT, and this effect can be reversed by peripheral leptin treatment. Central administration of leptin increases sympathetic nerve activity (Rahmouni and Morgan 2007), while a leptin antagonist blunts high fat feeding induced elevation of UCP 1 mRNA expression (Zhang et al. 2007) in BAT of rats, providing compelling evidence that leptin acts on the brain to regulate diet-induced thermogenesis. However, effects of leptin on skeletal muscle thermogenesis are more equivocal.
Leptin treatment was shown to increase oxygen uptake in explants of murine soleus muscle, indicative of thermogenesis (Dulloo et al. 2002), providing evidence for a direct effect on muscle. In an elegant study, Henry et al. (2008) demonstrated a post-prandial rise in thermogenesis that was evident in gluteal (subcutaneous) fat, visceral (retroperitoneal) fat and skeletal muscle. Importantly, central infusion of leptin markedly increased the post-prandial thermogenesis at all three sites, albeit to varying degrees. Central leptin infusion increased post-prandial thermogenesis to a greater extent in skeletal muscle and visceral fat than in gluteal fat (Henry et al. 2008), and this is associated with increased expression of UCP3 and UCP3 mRNA and increased mitochondrial oxygen consumption due to a shift towards uncoupled respiration (Henry et al. 2011). Furthermore, skeletal muscle and visceral fat exhibited higher baseline temperatures than gluteal fat. Together these data demonstrated that inherently different thermogenic capacities are operative in discrete tissues in sheep. Given that skeletal muscle represents a significant mass within the body and its robust thermogenic capabilities, the energy expended by this tissue is of great importance in the determination of daily energy expenditure.
In the context of what might happen in feedlot cattle, the accumulation of adipose tissue lipid and resultant increased plasma leptin may place a post-prandial thermogenic load that could become even more difficult to dissipate because of increased fat cover and reduced skin blood flow resulting from insulin resistance. This could become exacerbated with the resultant stress and the associated catecholamine-induced heat production around transport and slaughter. It is highly likely that even a small increase in body temperature as the animal enters the kill-chain could be enough to ensure that a carcass becomes classed as high rigor temperature.
Stress and temperature
A main cause of high rigor temperature appears to be a combination of rapid glycolysis and relatively slow cooling, which can lead to high rigor temperatures and potential toughening (Warner et al. 2014b). Moreover, high rigor temperature may cause a loss in proteolytic potential due to inactivation of proteolytic enzymes (Kim et al. 2014). Factors that contribute to elevated body temperature pre-slaughter would potentially predispose animals to higher carcass temperature immediately post slaughter. In the previous section, evidence was provided for impaired thermoregulatory capacity in cattle fed grain for long periods, due to the progressive development of an insulin-resistant state. Thermoregulatory capacity can also be altered by exposure to certain stressors. For instance, exposure to high ambient temperature, exercise (Warner et al. 2005) and psychological stress (Pighin et al. 2014) can result in an elevation of body temperature (core and peripheral temperatures) as a consequence of passive and/or active heat accumulation. In nearly all circumstances, an animal responds to external stimuli or stress by making complex adjustments to physiological processes in an attempt to maintain homeostasis. This normally involves activation of the hypothalamic-pituitary-adrenal (HPA) axis and sympathetic nervous system (SNS). Thus, an insulin-resistant state and/or stress may reduce the capacity of cattle fed grain for long periods to dissipate heat considerably in the immediate pre-slaughter period.
Effect of heat stress and exercise on body temperature
Exercise and thermal stress are known to increase core body temperature in a range of species, and the combination of the two stressors augments the elevation in body temperature (Febbraio et al. 1994). The magnitude of increase in core temperature depends on the severity and duration of each particular stressor. Although little is known about changes in muscle temperature per se during exercise and heat stress in ruminants, in humans it is documented that during exercise intramuscular temperature rises in proportion to the increase in workload, and the rise in intramuscular temperature is augmented during combined exercise and heat stress (Febbraio et al. 1994; Parkin et al. 1999). Warner et al. (2005) showed that lambs exercised for 10 min pre-slaughter had higher post-mortem muscle temperatures in the loin, neck and leg. Thus, it seems reasonable to assume that intramuscular temperature is also elevated in ruminants during exercise and thermal stress.
Effect of psychological stress on body temperature
With respect to thermoregulation in animals, recent research in the area of psychological stressors and effects on body temperature may provide some insight into variation in body temperature in livestock. The ‘psychogenic fever’ response has been most notably demonstrated in humans following physical examinations and after watching disturbing films, as well as in laboratory animals (Takakazu et al. 2001). This was recently demonstrated in lambs, where a rise in core body temperature occurred in response to the stressors associated with pre-slaughter handling such as mustering and handling (Pighin et al. 2014).
Stress can increase core temperature, and this effect can last for weeks, and even years, depending on the severity and duration of the stressor (Godsil et al. 2000; Takakazu et al. 2001; Pardon et al. 2004; Bhatnagar et al. 2006). Body temperature is also known to respond to conditioning, where the animal anticipates adverse environments and experiences, resulting in a rise in core temperature (Godsil et al. 2000; Pardon et al. 2004). This rise in core temperature is caused by a fever-like response, similar to that occurring during illness. An interaction between psychological stress and the fever response is thought to be associated with thermosensitive neurons in the preoptic area of the hypothalamus (that are central to the regulation of body temperature), which are responsive to non-thermal emotions, at least in monkeys (Takakazu et al. 2001). The release of stress hormones (i.e. activation of the HPA axis and stimulation of the SNS) mediates some, but not all, of the mechanisms that increase core temperature.
Some of the body temperature responses to social stress are clearly independent of the HPA response. Bhatnagar et al. (2006) demonstrated that repeated social stress in rats was associated with an increase in body temperature, presumably independent of activation of the HPA axis, since the elevation in core temperature was sustained well after increases in ACTH (adrenocorticotropic hormone) and corticosterone concentrations had returned to their basal levels following the generalised stress response. Interestingly, this repeated stress reduced bodyweight gain but increased feed intake during different periods, meaning that these changes to bodyweight cannot be explained entirely by the changes to feed intake, and may be the result of increased temperatures. An alternative mechanism may be that enhanced noradrenaline release in the hypothalamus during stress increases production of prostaglandin E2 (PGE2), which acts on the central nervous system to increase core temperature (Takakazu et al. 2001).
Skin blood flow decreases in response to stress as blood is redirected towards skeletal muscles and/or because of peripheral vasoconstriction (i.e. SNS-induced). Interestingly, this has also been observed with fear-related stress. In rats, tail and paw temperature decreased in response to fear, presumably due to SNS vasoconstrictor activity, although there was no marked reduction in temperature for other body surfaces (Vianna and Carrive 2005). Thus, these responses are quite differential and probably depend on body shape/conformation, etc.; similar responses have been noted in primates including humans.
Moreover, studies are often conflicting and it is difficult to ascertain whether there are real species differences or the measurement techniques are inadequate or that type of stressor and/or physiological state/animal variation is important. For instance, Ruis et al. (2001) found that gilts subjected to social isolation displayed significantly lower temperature, recorded from the ear and rectum by thermometer, which contrasts with most other studies. This indicates that perhaps species-specific responses occur. Re-exposure to fear-inducing situations in rats increases body temperature, possibly due to vasoconstriction limiting heat loss and metabolic effects generating heat via muscle activity and increased metabolism (Vianna and Carrive 2005). Those authors also found that the increase in body temperature following re-exposure to fear was delayed by 7 min and was higher in sham- than fear-conditioned animals. However, this may depend on the type of stressor, as Godsil et al. (2000) found that rectal temperatures of sham-conditioned rats did not increase, whereas that of animals who received foot shock increased by ~1°C.
Effects of elevated muscle temperature
Both exercise and heat stress can cause significant changes in cardiovascular and thermoregulatory processes; in combination, this can be a significant strain on animals. There has been considerable research in this area in several species including sheep (e.g. Bell et al. 1983) and humans (e.g. Buskirk 1977) but there are virtually no data in large ruminants. By contrast, much less is known about the combined effects of exercise and heat stress on whole animal and muscle metabolism (Febbraio 2001), with most of the available information in the field of human exercise physiology. Both of these stressors per se and especially in combination cause relative hyperthermia in humans and decreased exercise performance (Drust et al. 2005).
A striking metabolic alteration with exercise, at least for submaximal exercise in humans, is the preferential shift towards muscle glycogen utilisation at the expense of triglyceride use. and this appears to involve both oxidative and non-oxidative energy pathways (Febbraio 2001). Accumulation of muscle lactate and plasma lactate is also increased during exercise and heat stress in humans (e.g. Parkin et al. (1999)) and dogs (Kozlowski et al. 1985). This appears related to increased intramuscular glycogenolysis and not to increased uptake and oxidation of blood glucose. More recent studies have shown that while oxidation by muscle of exogenous glucose is reduced under these conditions (Jentjens et al. 2002), a combination of ingested glucose and fructose may improve oxidation of carbohydrate in muscle (Jentjens et al. 2006). Only limited information is available on muscle metabolism during heat stress and/or exercise in large ruminants. Glucose availability in ruminants relies predominantly on gluconeogenesis, so glucose sparing might be expected. However, Pethick et al. (1987) demonstrated that glucose was preferentially absorbed and oxidised over NEFA in the hindlimb of exercising sheep under thermo-neutral conditions. How the imposition of heat stress per se or in combination with exercise influences the pattern of substrate oxidation in ruminants is not known.
The mechanisms responsible for alterations in metabolism during combined exercise and heat stress or heat stress per se have not need elucidated. Several possible mechanisms have been proposed, including a direct temperature effect on enzyme reaction rates (the Q10 effect; Young 1990), reduced oxygen and substrate delivery to muscle as a result of competing demands for blood-flow delivery to the periphery to aid in heat dissipation (Rowell 1974), and an increase in circulating epinephrine concentrations (Yaspelkis et al. 1993).
Probably one of the most important implications of elevated intramuscular temperature per se is the potential effect on key enzymes that ultimately influence muscle metabolism, the so-called Q10 effect (Young 1990). A Q10 value represents the x-fold increase in enzyme reaction rate for every 10°C increase in temperature; this is usually ~2–3 for enzyme-mediated reactions. Thus, for increases in intramuscular temperature of ~1°C, enzyme reaction rates could increase by ~15–20%. The implication for elevated carcass temperature immediately post-slaughter is obvious if glycogen is preferentially utilised pre-slaughter and its metabolism accelerated post-slaughter because of the elevated temperatures.
Stress and heat shock proteins
Considerable research has also recently focused on the role of intracellular HSPs in protecting cell integrity during exposure to stress, in particular hyperthermia. Heat shock proteins are a family of highly conserved proteins of varying molecular weight and are found in nearly all cells and tissues (Lindquist 1986). Some are expressed constitutively in non-stressful situations and others are induced by a range of different stressors, including heat stress and exercise.
One of the most important of the HSPs is HSP70 and its inducible form, HSP72. Activation of the HPA and SNS axes are likely to play an important role in stimulating the synthesis of HSPs (Fleshner and Johnson 2005). These proteins are rapidly induced in response to a range of stressors, including heat stress (Ruell et al. 2004), exercise in humans (Febbraio et al. 2002; Whitham et al. 2006) and psychological stress in rats (Fleshner et al. 2004). Small HSPs are involved in muscle cell maintenance and repair, and the upregulation of some of these proteins, such as HSP27, is related to improved tenderness, juiciness and flavour in beef (Bernard et al. 2007); in transported pigs, a decline in HSP27 expression is associated with a decline in meat quality (Yu et al. 2009). Not only are HSPs rapidly induced intracellularly, but they can also appear in blood, although the exact mechanism for translocation into blood has not been identified, and the importance of elevated circulating HSPs has not been established. Nonetheless, they may be a valuable biomarker of stress in animals.
Heat shock proteins and thermo-tolerance
It is conceivable that variation in the rapidity and extent of induction of HSPs between animals in their response to stressors such heat and exercise, and psychological stress, may translate into variation in thermo-tolerance. In humans for instance, heat intolerance appears related to transcriptional malfunction associated with HSP induction, along with other stress-related proteins, which suggests some inherent basis for thermoregulatory dysfunction (Moran et al. 2006). This may have important implications for animal health and performance, but equally, it may have significant implications for intracellular protein integrity (e.g. protection of proteolytic enzymes from heat-induced denaturation) in the carcass post-slaughter. For instance, some animals may be more sensitive to elevated body temperature and have enhanced upregulation of HSP synthesis in the pre-slaughter period, which subsequently protects muscle cell integrity in the carcass. This may help to reduce high rigor temperature and other meat quality defects associated with elevated carcass temperature post-slaughter. For example, in Duroc boars, differences in the genotype at the HSP72 gene appear to influence backfat deposition under different environmental temperatures, although this appears to be most prominent in cooler months (Huang et al. 2004).
In addition to any inherent variation between animals in the induction of HSPs and thermo-tolerance, recent evidence suggests that it may be possible to pre-condition animals to thermal stress, enabling them to cope with subsequent heat-stress events. Although the exact mechanisms for this have not been elucidated, short-term (30-min) hyperthermia preconditioning in mice was associated with enhanced survival rates and elevated hepatic levels of HSP70 (King et al. 2002).
In addition to hyperthermia pre-conditioning, there is now ample evidence in rodents that other forms of pre-conditioning, either alone or in combination with hyperthermia pre-conditioning, may offer significant protection for tissues and cells from subsequent exposure to a range of stressors. One important example is the demonstrated role of L-glutamine (L-Gln) in potentiating the induction of HSP following subsequent heat stress, injury or sepsis (Wischmeyer et al. 2001; Ropeleski et al. 2005; Morrison et al. 2006; Wang et al. 2007). As one of the most abundant amino acids in plasma, L-Gln is synthesised and stored in several tissues, including both lung and skeletal muscle. L-Gln becomes conditionally essential in states of severe stress in which intracellular L-Gln levels can fall by ~50% and plasma concentrations by 30% (Kovacevic and McGivan 1983). A study by Singleton and Wischmeyer (2006) showed that in rats previously administered with L-Gln orally and then exposed to hyperthermic conditions, HSP expression in gut tissues was enhanced, gut permeability was reduced, and plasma endotoxin concentrations were lower than in rats exposed to hyperthermia without L-Gln supplementation. The integrity of the gastrointestinal epithelium is highly susceptible to heat stress, and thus it is not surprising that HSP72 is highly expressed in gut tissue even under thermo-neutral conditions (Ruell et al. 2004), and protection of gut tissue under these conditions plays a vital role in allowing animals to cope with elevated body temperature. By contrast, some tissues decrease HSP expression in response to stress. Rats exposed to restraint stress had decreased HSP90 in liver and spleen tissues, while brain and some peripheral tissues (thymus, adrenal glands and testes) showed no change to their HSP70/90 steady-state levels (Vamvakopoulos et al. 1993).
Heat-stressed cells have compromised mitochondrial function, and thus aerobic metabolism is adversely affected, causing cells to rely more heavily upon glycolytic energy metabolism (Welch 1992). Intriguingly, chaperone proteins (HSPs) are well documented to protect against obesity-induced insulin resistance (e.g. Kurucz et al. 2002; McCarty 2006; Chung et al. 2008; Gupte et al. 2011). While the exact mechanism behind this phenomenon is unidentified, it is thought to involve reduced serum insulin and IGF-1 levels (McCarty 2006) and it is also suggested to be due to the protective effects of HSPs limiting the actions of plasma free radicals (Kurucz et al. 2002). Furthermore, HSPs may offer mitochondrial protection by preventing damage to oxidation processes (Gupte et al. 2008). Intriguingly, people with an inherited reduced capacity to synthesise HSPs, as well as ageing populations, have an increased risk of developing insulin resistance (McCarty 2006; Kavanagh et al. 2009).
Chung et al. (2008) showed the expression of HSP70, but not HSP90, protein in human skeletal muscle to be markedly reduced in insulin-resistant populations, and in the skeletal muscle of type 2 diabetic humans, HSP70 expression was decreased. In the same study, heat therapy protected mice from basal hyperglycemia and hyperinsulinemia otherwise induced by consuming a high-fat diet (Chung et al. 2008). Moreover, diabetic rodents and humans with higher concentrations of HSP70 have higher survival rates (Kavanagh et al. 2009). This leads to the interesting notion that HSP induction may combat insulin resistance. Furthering this idea, liver samples from diabetic animals had significantly less heat shock factor 1 (HSF1, a transcription factor that regulates the expression of HSP) and somewhat less HSP70 than normal animals (Kavanagh et al. 2009). Human skeletal muscle HSP70 is strongly correlated with numerous insulin-stimulated metabolic changes such as decreased lipid oxidation and increased uptake, oxygenation and storage of glucose (Kurucz et al. 2002). In adipose cells, prolonged oxidative stress negatively influenced insulin-stimulated GLUT4 actions, therefore limiting insulin-stimulated increases in glucose transport (Kurucz et al. 2002). Nevertheless, involvement of HSPs in insulin signalling and nutrient partitioning requires further investigation, which may provide insights into how heat affects substrate metabolism at a cellular level.
Dietary interventions to reduce high rigor temperature
Betaine
The most compelling dietary intervention to date is the use of betaine (trimethylglycine), as reviewed by Dunshea et al. (2005). Mammals utilise betaine for two major functions. The first is as a methyl donor (via S-adenosyl-methionine (SAM)), thereby sparing methionine and increasing the available substrates for protein synthesis (Matthews et al. 2001). Second, when not catabolised, betaine is used as an organic osmoprotectant (Fernández et al. 1998; Huang et al. 2007). Bacterial studies show that betaine is able to assist the organism to survive high osmolarities by restoring changes to enzymes and stabilising and assisting in thermoprotection (Caldas et al. 1999). As an osmolyte, betaine accumulates within osmotically stressed cells (Eklund et al. 2005) and betaine uptake can be via active (Na+ Cl–) or passive transporter methods (Craig 2004). This is an important role, as loss of water within a cell can lead to cell damage or catabolism, with osmotic stress interrupting the cell cycle and halting cell proliferation while causing cells to initiate apoptosis.
Dietary betaine has also been shown to improve the growth performance of pigs by reducing the maintenance energy requirement due to its osmolyte properties reducing the need for sodium/potassium pumps (Schrama et al. 2003; Suster et al. 2004). Betaine supplementation (0.125%) significantly lowered serum urea nitrogen in pigs, while total protein levels significantly increased (Huang et al. 2007), and IGF-1, free thyronine (FT3), free thyroxine (FT4) and insulin levels all significantly increased by 40–50% in response to betaine supplementation (Huang et al. 2006).
Betaine can also improve water-holding capacity and reduce drip loss in meat (Dunshea et al. 2005), although in meat lambs, betaine supplementation did not alter the hydration status of the animals at slaughter (Pearce et al. 2008). Also, dietary betaine can improve the integrity of gut mucosal cells and reduce the severity of some enteric infections in poultry (Matthews and Southern 2000; Klasing et al. 2002). Cronje (2005) has suggested that that heat stress and its manifestations are a disease of the gut and that dietary betaine may assist by improving gut integrity.
Most investigations utilising dietary betaine as a nutritional supplement involve pigs and poultry, although there is evidence that dietary betaine can reduce heat stress and improve feed intake and growth performance in beef cattle (Cronje 2005; Loxton et al. 2007). Growing lambs supplemented with 2 g/kg of betaine had significantly reduced fat thickness, while liveweight and muscle area did not differ (Fernández et al. 1998). Importantly, albeit with small numbers, Loxton et al. (2007) found that dietary betaine supplementation reduced the incidence of heat toughening (defined as passing through the MSA temperature–pH window) from 5/6 carcasses to 1/7 carcasses. In sheep, no change to bodyweight and lean muscle content was seen in response to betaine supplementation (Fernández et al. 2000). We recently showed dietary betaine supplementation (0, 2 and 4 g betaine/day) to have dose-dependent physiological responses in sheep exposed to heat (36−43°C) and thermo-neutral conditions (22°C) (DiGiacomo 2011), such that regardless of environmental temperature, rectal temperatures were lower in sheep supplemented 2 g betaine/day and higher in those supplemented 4 g betaine/day compared with control sheep (39.6, 39.8 and 39.7°C, respectively, P = 0.002). Similar responses were seen for respiration rate (113, 102 and 116 breaths/min for 0, 2 and 4 g betaine/day, respectively) and heart rate (95, 88 and 99 beats/min) (DiGiacomo et al. 2012).
In lactating dairy cows, plasma NEFA concentrations decreased linearly with increasing dietary betaine supplementation levels, which may be the direct result of increased insulin concentrations inhibiting NEFA production (Wang et al. 2010a). Increased circulating NEFA concentrations are involved in insulin resistance, and therefore a decrease in circulating NEFA levels due to dietary betaine supplementation may be beneficial (Boston et al. 2008). In mice fed a high-fat diet, dietary betaine supplementation inhibits lipogenesis and reduces insulin resistance by improving hepatic function via improved adipose tissue function and reduced hepatic damage (Wang et al. 2010b). Betaine supplementation reverses insulin resistance in an in vitro model, while in both in vitro and in vivo models, betaine normalises downstream signalling pathways involved in gluconeogenesis, perhaps by improving phosphorylation of early steps in the insulin signalling cascade (Kathirvel et al. 2010). Dietary betaine has been shown to improve insulin resistance in humans, again via the amelioration of hepatic injury (Borgschulte et al. 2008). Finally, human patients suffering non-alcoholic fatty liver disease have improved hepatic function when given a dietary supplementation of betaine (Patrick 2002; Borgschulte et al. 2008;Abdelmalek et al. 2009). Although the mode of action via which betaine elicits these effects is contentious, betaine is thought to ameliorate endoplasmic reticulum stress (Wang et al. 2010b) by converting homocysteine into methionine (a fatty acid) (Abdelmalek et al. 2009), although this is not always the case (Borgschulte et al. 2008).
Other dietary strategies should be directed towards alleviating insulin resistance or the stress around slaughter, for example, the use of antidiabetic agents such as thiazolidinediones (TZD), chromium, zinc and vanadium.
Type II diabetes strategies (chromium, Zn, TZD, vanadium)
Given that our hypothesis is largely based around the premise that the insulin resistance associated with fattening in cattle grain fed long-term can predispose the animal towards thermal instability, then possible solutions may be some of the antidiabetic compounds such as chromium, zinc, TZD and vanadium.
Dietary chromium has been suggested to increase insulin sensitivity (Steele et al. 1977), and over the last 20 years or so, it has been occasionally revisited as a potential means of manipulating fat deposition in humans and farm animals. While chromium chloride has been successful in improving growth performance of poultry (Steele and Rosebrough 1979; Uyanik et al. 2002) or protein-deficient rats (Mertz and Roginski 1969), effects in pigs (Page et al. 1993; Mooney and Cromwell 1995; Shelton et al. 2003) and ruminants (Kegley et al. 1997a; Kegley et al. 2000) have been more equivocal. In part, this may reflect variability in absorption of chromium, since the absorption and utilisation of chromium may depend on incorporation into organic molecules (Evans and Johnson 1980; Kegley et al. 1997b). However, when organic forms of chromium have been used, there have been consistent improvements in insulin sensitivity in ruminants fed dietary chromium. For example, heifers supplemented with chromium propionate had lower serum insulin concentrations following a glucose infusion (in non-fasted animals), although this did not appear to alter glucose uptake, perhaps indicating increased tissue insulin sensitivity (Spears et al. 2012). Holstein calves supplemented with chromium in summer had a reduced insulin response (~30%) to a glucose infusion compared with control animals, suggesting improved efficiency of insulin (Yari et al. 2010). Furthermore, dry matter intake, milk fat and protein yield increased while serum insulin and NEFA concentrations decreased in chromium-supplemented, early-lactation Holsteins during summer, again suggesting an increase in insulin sensitivity (Nikkhah et al. 2011).
Supplemental chromium picolinate increased glucose clearance rate and decreased glucose half-life in calves fed diets based on corn–cottonseed hulls (Bunting et al. 1994). In calves fed milk replacer, supplemental chromium nicotinate slowed the return to basal glucose concentration after an insulin infusion (Kegley et al. 1997b). Several other studies with calves and young cattle have shown improved glucose clearance after either glucose or insulin challenges (Bunting et al. 2000; Depew et al. 1998; Kegley et al. 2000). While little or no work appears to have been done in older cattle, Gardner et al. (1998) found that dietary chromium chelavite reduced GR fat depth and increased the activity of ATP citrate lyase, a marker of the glucose/insulin axis, by 30% in relatively mature sheep (2-year-old) sheep. Therefore, these data suggest that dietary chromium supplementation may be a means of improving insulin sensitivity in cattle fed grain long-term and improve thermoregulation. However, it should be noted that supplemental chromium did not affect rectal temperature after challenge with infectious bovine rhinotracheitis virus in steers. In a succinct review by Hua et al. (2012), the potential mechanisms driving chromium’s effect on improving insulin resistance are explored, and it is noted that recent studies have found chromium to improve the actions of insulin and the insulin-signalling pathway while inhibiting downregulators of insulin signalling.
At relatively high levels, zinc exerts potent insulin-mimetic effects, and so dietary zinc has been proposed as a means of manipulating adipogenesis and marbling in cattle (Kawachi 2006). Sasaki et al. (2002) showed that insulin promoted adipocyte proliferation in ovine preadipocyte cells, while Tanaka et al. (2001) found that zinc enhanced adipogenesis in 3T3-L1 adipose cells. Yano et al. (2004) investigated the effect of zinc, either alone or in combination with insulin, on markers of adipogenesis (glycerol-3-phosphate dehydrogenase activity or lipid content) in bovine preadipocytes and found that zinc increased adipogenesis in a dose-dependent manner, although the effect was not as great as insulin. Whether these effects translate into in vivo action or occur in other insulin-dependent tissues is unknown. Also, supplemental zinc has had no effect on plasma glucose Wang (2006) or glucose clearance (Droke et al. 1993) in lambs.
Vanadium has also been reported to possess insulin-mimetic activity on various types of cells and so has been suggested as a means of increasing adipogenesis (Kawachi 2006; Shukla and Bhonde 2008). Although vanadium is used as a treatment for diabetes and it may improve insulin sensitivity in cattle, concerns about toxicity and contamination of the food chain would probably ensure that it is not used (Gummow et al. 2005). However, given that it is used as a low-dose, orally active antidiabetogenic agent in humans (Rustenbeck 2007), if proof-of-concept studies demonstrate that insulin resistance is involved in high rigor temperature, then vanadium may be worth considering as a dietary supplement.
Thiazolidinediones are potent, synthetic ligands for peroxisome proliferator-activated receptor gamma (PPARγ) that potentiate the action of insulin in peripheral tissues, and are used as orally active treatments for diabetes in humans (Krentz and Bailey 2005). Kushibiki et al. (2001) administered TZD to steers injected with TNF-α to induce insulin resistance and observed a decrease in plasma concentrations of NEFA, insulin and glucagon, suggesting that administration of TZD ameliorated insulin action. Administration of TZD in the prepartum period for dairy cattle increased PPARγ mRNA expression and decreased leptin mRNA expression (Schoenberg et al. 2011). Also, Smith et al. (2007) injected TZDs (intrajugular) to lactating cows, which are normally very insulin-resistant, and observed an improvement in insulin sensitivity as evidenced by a decrease in plasma NEFA. Therefore, TZD may be an effective means of improving insulin resistance in cattle fed grain long-term, although it is not known whether TZDs survive the rumen. Also, the use of TZDs for production animals is likely to be cost-prohibitive.
Vitamin D deficiency is associated with increased insulin resistance (Reis et al. 2005), and there is some evidence that vitamin D supplementation in humans may improve insulin sensitivity. Normally sufficient vitamin D is produced after exposure to sunlight, and intuitively, lot-fed cattle in Australia are unlikely to receive no exposure to sunlight. However, dark-skinned individuals need much more sunlight to produce sufficient vitamin D, and dark-skinned or dark-haired cattle are more likely to absorb heat than light-skinned or light-haired cattle. It would also be of interest to observe whether carcasses form dark cattle are more likely to be of high rigor temperature that light cattle. Several studies have shown that vitamin D supplementation can improve tenderness in beef through elevation of serum calcium, calcium being a potent activator of proteases involved in tenderisation (Swanek et al. 1999; Montgomery et al. 2004).
Magnesium to reduce catecholamine secretion and activation (stress relief)
Magnesium (Mg) has a relaxant effect on skeletal muscle and it has been shown to depress skeletal muscle activity by antagonising calcium, which is required for neurotransmitter release and muscle contraction (see review by Dunshea et al. 2005). Studies have shown that dietary magnesium supplementation alleviates the effects of stress by reducing plasma cortisol, norepinephrine, epinephrine and dopamine concentrations (Niemack et al. 1979; Kietzmann and Jablonski 1985). Consequently, studies have been conducted to investigate the influence of dietary Mg supplementation on reducing the effects of stress and improving pork quality. Given that the stress around slaughter may result in a loss of thermoregulation, dietary Mg may assist in reducing heat toughening.
Magnesium supplementation (MgO) has also been shown to reduce the loss of glycogen from muscle in Merino lambs during the post-farm period leading up to slaughter (Gardner et al. 2001). This has led to some supply chains in Australia insisting on high-energy rations containing supplemental MgO being fed to Merino lambs in the 2–4 weeks pre-slaughter. This logic is based on the accepted fact that the Merino breed, on average, has a higher incidence of dark cutting (or high pHu meat) due to greater stress sensitivity and so increased rate of glycogen loss caused by the post-farm stressors that occur immediately pre-slaughter (Gardner et al. 1998). Therefore, dietary Mg may be worth investigating as a means of reducing the heat generation in response to pre-slaughter stress of cattle.
Other
Other targets may include manipulation of NOS through use of NO donors such as arginine or the use of dietary L-Gln to potentiate HSPs. Other targets that are more difficult to manipulate are leptin and UCPs.
Conclusion
A main cause of high rigor temperature appears to be a combination of rapid glycolysis and relatively slow cooling, which can lead to high rigor temperatures and potential toughening. Moreover, high rigor temperature may be exacerbated by a loss in proteolytic potential due to inactivation of proteolytic enzymes. Factors that contribute to elevated body temperature pre-slaughter potentially predispose animals to higher carcass temperature immediately post-slaughter. There is mounting evidence that a state of insulin resistance can compromise the ability of an individual to thermoregulate and this can be exacerbated by stressors. Several dietary treatments might be investigated as means of reducing heat toughening. Most promising are dietary betaine, chromium and magnesium either alone or in combination.
References
Abdelmalek MF, Sanderson SO, Angulo P, Soldevila-Pico C, Liu C, Peter J, Keach J, Cave M, Chen T, McClain CJ, Lindor KD (2009) Betaine for nonalcoholic fatty liver disease: Results of a randomized placebo-controlled trial. Hepatology (Baltimore, Md.) 50, 1818–1826.| Betaine for nonalcoholic fatty liver disease: Results of a randomized placebo-controlled trial.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhs1SjurvI&md5=68a83088856ab185bd2b0e00213fdc36CAS |
Achmadi J, Yanagisawa T, Sano H, Terashima Y (1993) Pancreatic insulin secretory response and insulin action in heat-exposed sheep given a concentrate or roughage diet. Domestic Animal Endocrinology 10, 279–287.
| Pancreatic insulin secretory response and insulin action in heat-exposed sheep given a concentrate or roughage diet.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2cXhs1antbw%3D&md5=3a6c158a3a375756af118ca111c68f01CAS | 8306632PubMed |
Ahima RS, Flier JS (2000) Leptin. Annual Review of Physiology 62, 413–437.
| Leptin.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXltlGrsbs%3D&md5=43d9097da41f57bad15555314c0ec3a6CAS | 10845097PubMed |
Alamer M (2011) The role of prolactin in thermoregulation and water balance during heat stress in domestic ruminants. Asian Journal of Animal and Veterinary Advances 6, 1153–1169.
| The role of prolactin in thermoregulation and water balance during heat stress in domestic ruminants.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XktFajsQ%3D%3D&md5=04ea72835b7af56828717f25a140b30aCAS |
Auwerx J, Staels B (1998) Leptin. Lancet 351, 737–742.
| Leptin.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXitFeit7s%3D&md5=2c70ecbf6f631dbdbde65b40b2cf4ffeCAS | 9504534PubMed |
Bell AW, Hales JR, King RB, Fawcett AA (1983) Influence of heat stress on exercise-induced changes in regional blood flow in sheep. Journal of Applied Physiology 55, 1916–1923.
Bernabucci U, Basirico L, Lacetera N, Morera P, Ronchi B, Accorsi PA, Seren E, Nardone A (2006) Photoperiod affects gene expression of leptin and leptin receptors in adipose tissue from lactating dairy cows. Journal of Dairy Science 89, 4678–4686.
| Photoperiod affects gene expression of leptin and leptin receptors in adipose tissue from lactating dairy cows.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhtlWgsrvO&md5=b0d375741e254df5971c1aa0c7d673e9CAS | 17106100PubMed |
Bernard C, Cassar-Malek I, Le Cunff M, Dubroeucq H, Renand G, Hocquette J-F (2007) New indicators of beef sensory quality revealed by expression of specific genes. Journal of Agricultural and Food Chemistry 55, 5229–5237.
| New indicators of beef sensory quality revealed by expression of specific genes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXmtVKlt7g%3D&md5=f91551cd15f777975726e7b7c12223e7CAS | 17547415PubMed |
Bhatnagar S, Vining C, Iyer V, Kinni V (2006) Changes in hypothalamic-pituitary-adrenal function, body temperature, body weight and food intake with repeated social stress exposure in rats. Journal of Neuroendocrinology 18, 13–24.
| Changes in hypothalamic-pituitary-adrenal function, body temperature, body weight and food intake with repeated social stress exposure in rats.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhvFKmt7g%3D&md5=ea9f66c0442cc59ab81c3cc29eecc353CAS | 16451216PubMed |
Block S, Butler W, Ehrhardt R, Bell A, Van Amburgh M, Boisclair Y (2001) Decreased concentration of plasma leptin in periparturient dairy cows is caused by negative energy balance. The Journal of Endocrinology 171, 339–348.
| Decreased concentration of plasma leptin in periparturient dairy cows is caused by negative energy balance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXosV2htL4%3D&md5=e1928bc6ddc52e952c2e0ac1bff8815fCAS | 11691654PubMed |
Boisclair YR, Bell AW, Dunshea FR, Harkins M, Bauman DE (1993) Evaluation of the arteriovenous difference technique to simultaneously estimate protein synthesis and degradation in the hindlimb of fed and chronically underfed steers. The Journal of Nutrition 123, 1076–1088.
Boisclair YR, Bauman DE, Bell AW, Dunshea FR, Harkins M (1994) Nutrient utilization and protein turnover in the hindlimb of cattle treated with bovine somatotropin. The Journal of Nutrition 124, 664–673.
Bole-Feysot C, Goffin V, Edery M, Binart N, Kelly PA (1998) Prolactin (PRL) and its receptor: Actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. Endocrinology Reviews 19, 225–268.
Borgschulte G, Kathirvel E, Herrera M, French SW, Morgan TR, Morgan K, Bottiglieri T (2008) Betaine treatment reverses insulin resistance and fatty liver disease without reducing oxidative stress or endoplasmic reticulum stress in an animal model of nafld. Gastroenterology 134, A-414–A-415.
| Betaine treatment reverses insulin resistance and fatty liver disease without reducing oxidative stress or endoplasmic reticulum stress in an animal model of nafld.Crossref | GoogleScholarGoogle Scholar |
Boston RC, Roche JR, Ward GM, Moate PJ (2008) A novel minimal model to describe non-esterfied fatty acid kinetics in Holstein dairy cows. The Journal of Dairy Research 75, 13–18.
| A novel minimal model to describe non-esterfied fatty acid kinetics in Holstein dairy cows.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXitVOmu7w%3D&md5=0a970a35569d71dd517bcc2bc250a0d1CAS | 17922935PubMed |
Bradley SJ, Kingwell BA, Canny BJ, McConell GK (2007) Skeletal muscle neuronal nitric oxide synthase μ protein is reduced in people with impaired glucose homeostasis and is not normalized by exercise training. Metabolism: Clinical and Experimental 56, 1405–1411.
| Skeletal muscle neuronal nitric oxide synthase μ protein is reduced in people with impaired glucose homeostasis and is not normalized by exercise training.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhtVOgu7rJ&md5=6a144ea3c9da9cd9a2a06bca0b6e23d6CAS |
Bunting LD, Fernandez JM, Thompson DL, Southern LL (1994) Influence of chromium picolinate on glucose usage and metabolic criteria in growing Holstein calves. Journal of Animal Science 72, 1591–1599.
Bunting LD, Tarifa TA, Crochet BT, Fernandez JM, Depew CL, Lovejoy JC (2000) Effects of dietary inclusion of chromium propionate and calcium propionate on glucose disposal and gastrointestinal development in dairy calves. Journal of Dairy Science 83, 2491–2498.
| Effects of dietary inclusion of chromium propionate and calcium propionate on glucose disposal and gastrointestinal development in dairy calves.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXosVCgsLk%3D&md5=1bbe954dd0ba6bf4b9ab1161b4bdac60CAS | 11104268PubMed |
Buskirk ER (1977) Temperature regulation with exercise. Exercise and Sport Sciences Reviews 5, 45–88.
| Temperature regulation with exercise.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaE1M%2FgtlChtg%3D%3D&md5=6e238b9d9fe0f5437de47f13a2a06aeeCAS | 357161PubMed |
Caldas T, Demont-Caulet N, Ghazi A, Richarme G (1999) Thermoprotection by glycine betaine and choline. Microbiology 145, 2543–2548.
Cals-Grierson MM, Ormerod AD (2004) Nitric oxide function in the skin. Nitric Oxide 10, 179–193.
| Nitric oxide function in the skin.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXmtVaitrY%3D&md5=584926bb8c3067dbf5fa6694953dad2fCAS | 15275864PubMed |
Chilliard Y, Delavaud C, Bonnet M (2005) Leptin expression in ruminants: Nutritional and physiological regulations in relation with energy metabolism. Domestic Animal Endocrinology 29, 3–22.
| Leptin expression in ruminants: Nutritional and physiological regulations in relation with energy metabolism.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXltVegsr4%3D&md5=194c8411134c59def88e99da6b038935CAS | 15876510PubMed |
Choshniak I, Jenkinson DM, Blatchford DR, Peaker M (1982) Blood flow and catecholamine concentration in bovine and caprine skin during thermal sweating. Comparative Biochemistry and Physiology Part C: Comparative Pharmacology 71, 37–42.
| Blood flow and catecholamine concentration in bovine and caprine skin during thermal sweating.Crossref | GoogleScholarGoogle Scholar |
Chung J, Nguyen A-K, Henstridge DC, Holmes AG, Stanley Chan MH, Mesa JL, Lancaster GI, Southgate RJ, Bruce CR, Duffy SJ, Horvath I, Mestril R, Watt MJ, Hooper PL, Kingwell BA, Vigh L, Hevener A, Febbraio MA (2008) HSP72 protects against obesity-induced insulin resistance. Proceedings of the National Academy of Sciences of the United States of America 105, 1739–1744.
| HSP72 protects against obesity-induced insulin resistance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhvFalsbs%3D&md5=cfd7a9651935744fdf7680ac2c704468CAS | 18223156PubMed |
Collier RJ, Beede DK, Thatcher WW, Israel LA, Wilcox CJ (1982) Influences of environment and its modification on dairy animal health and production. Journal of Dairy Science 65, 2213–2227.
| Influences of environment and its modification on dairy animal health and production.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaL3s7gvFOktg%3D%3D&md5=f336bc4528dcc75affbe293c497eab20CAS | 6759540PubMed |
Collier RJ, Dahl GE, VanBaale MJ (2006) Major advances associated with environmental effects on dairy cattle. Journal of Dairy Science 89, 1244–1253.
| Major advances associated with environmental effects on dairy cattle.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28Xjt1SjsLk%3D&md5=2f5bbe9b1bfbc89034cd50d0670ae57bCAS | 16537957PubMed |
Considine RV (2001) Regulation of leptin production. Reviews in Endocrine & Metabolic Disorders 2, 357–363.
| Regulation of leptin production.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXpt1Wit70%3D&md5=32674b6336d852d42e55739579c85863CAS |
Craig SAS (2004) Betaine in human nutrition. The American Journal of Clinical Nutrition 80, 539–549.
Cronje P (2005) Heat stress in livestock - the role of the gut in its aetiology and a potential role for betaine in its alleviation. Recent Advances in Animal Nutrition in Australia 15, 107–122.
Depew CL, Bunting LD, Fernandez JM, Thompson DL, Adkinson RW (1998) Performance and metabolic responses of young dairy calves fed diets supplemented with chromium tripicolinate. Journal of Dairy Science 81, 2916–2923.
| Performance and metabolic responses of young dairy calves fed diets supplemented with chromium tripicolinate.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXnsV2jt7o%3D&md5=7a2e4b437162ec1a692adc17db5fadc6CAS | 9839235PubMed |
DiGiacomo K (2011) The physiological and metabolic responses to heat in ruminants. PhD Thesis. The University of Melbourne, Vic., Australia.
DiGiacomo K, Simpson S, Leury BJ, Dunshea FR (2012) Dietary betaine improves physiological responses in sheep under chronic heat load in a dose dependent manner. Journal of Animal Science 90, 269
Droke EA, Spears JW, Armstrong JD, Kegley EB, Simpson RB (1993) Dietary zinc affects serum concentrations of insulin and insulin-like growth factor I in growing lambs. The Journal of Nutrition 123, 13–19.
Drust B, Rasmussen P, Mohr M, Nielsen B, Nybo L (2005) Elevations in core and muscle temperature impairs repeated sprint performance. Acta Physiologica Scandinavica 183, 181–190.
| Elevations in core and muscle temperature impairs repeated sprint performance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXhtlOqtrg%3D&md5=f2936a4b4c1e920987790fda25d3dd53CAS | 15676059PubMed |
Dulloo AG, Stock MJ, Solinas G, Boss O, Montani JP, Seydoux J (2002) Leptin directly stimulates thermogenesis in skeletal muscle. FEBS Letters 515, 109–113.
| Leptin directly stimulates thermogenesis in skeletal muscle.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38Xis1eisrs%3D&md5=12dc1645b5e03fb5138cbc546210e9c7CAS | 11943204PubMed |
Dunshea FR, Boisclair YR, Bauman DE, Bell AW (1995) Effects of bovine somatotropin and insulin on whole-body and hindlimb glucose metabolism in growing steers. Journal of Animal Science 73, 2263–2271.
Dunshea FR, D’Souza DN, Pethick DW, Harper GS, Warner RD (2005) Effects of dietary factors and other metabolic modifiers on quality and nutritional value of meat. Meat Science 71, 8–38.
| Effects of dietary factors and other metabolic modifiers on quality and nutritional value of meat.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXmt1Wiu7g%3D&md5=1d5e69a93374b948caec262a6b8bb206CAS | 22064049PubMed |
Dunshea FR, Leury BJ, Fahri F, DiGiacomo K, Hung A, Chauhan S, Clarke IJ, Collier R, Little S, Baumgard LH, Gaughan J (2013) Amelioration of thermal stress impacts in dairy cows. Animal Production Science 53, 965–975.
Ehrhardt R, Slepetis R, Siegal-Willott J, Van Amburgh M, Bell A, Boisclair Y (2000) Development of a specific radioimmunoassay to measure physiological changes of circulating leptin in cattle and sheep. The Journal of Endocrinology 166, 519–528.
| Development of a specific radioimmunoassay to measure physiological changes of circulating leptin in cattle and sheep.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXmslymu7w%3D&md5=cea19416432da32f8be7dd0b166ff7caCAS | 10974646PubMed |
Eisemann JH, Huntington GB, Catherman DR (1997) Insulin sensitivity and responsiveness of portal-drained viscera, liver, hindquarters, and whole body of beef steers weighing 275 or 490 kilograms. Journal of Animal Science 75, 2084–2091.
Eklund M, Bauer E, Wamatu J, Mosenthin R (2005) Potential nutritional and physiological functions of betaine in livestock. Nutrition Research Reviews 18, 31–48.
| Potential nutritional and physiological functions of betaine in livestock.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXntFSktLk%3D&md5=f946c9aecbbd77322ee21b2444cb50e1CAS | 19079893PubMed |
Evans GW, Johnson EC (1980) Zinc absorption in rats fed a low-protein diet and a low-protein diet supplemented with tryptophan or picolinic acid. The Journal of Nutrition 110, 1076–1080.
Faichney GJ, Barry TN (1986) Effects of mild heat exposure and suppression of prolactin secretion on gastro-intestinal tract function and temperature regulation in sheep. Australian Journal of Biological Sciences 39, 85–97.
Febbraio MA (2001) Alterations in energy metabolism during exercise and heat stress. Sports Medicine (Auckland, N.Z.) 31, 47–59.
| Alterations in energy metabolism during exercise and heat stress.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD3M3isFaktA%3D%3D&md5=d3f1af7fdad24be6abfd780f30c5b726CAS |
Febbraio MA, Snow RJ, Stathis CG, Hargreaves M, Carey MF (1994) Effect of heat stress on muscle energy metabolism during exercise. Journal of Applied Physiology 77, 2827–2831.
Febbraio MA, Ott P, Nielsen HB, Steensberg A, Keller C, Krustrup P, Secher NH, Pedersen BK (2002) Exercise induces hepatosplanchnic release of heat shock protein 72 in humans. The Journal of Physiology 544, 957–962.
| Exercise induces hepatosplanchnic release of heat shock protein 72 in humans.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XovF2qt7Y%3D&md5=68f1f80c1e203c8c1845e9f098df600cCAS | 12411538PubMed |
Fernández C, Gallego L, Lopez-Bote CJ (1998) Effect of betaine on fat content in growing lambs. Animal Feed Science and Technology 73, 329–338.
| Effect of betaine on fat content in growing lambs.Crossref | GoogleScholarGoogle Scholar |
Fernández C, López-Saez A, Gallego L, de la Fuente JM (2000) Effect of source of betaine on growth performance and carcass traits in lambs. Animal Feed Science and Technology 86, 71–82.
| Effect of source of betaine on growth performance and carcass traits in lambs.Crossref | GoogleScholarGoogle Scholar |
Fleshner M, Johnson JD (2005) Endogenous extra-cellular heat shock protein 72: Releasing signal(s) and function. International Journal of Hyperthermia 21, 457–471.
| Endogenous extra-cellular heat shock protein 72: Releasing signal(s) and function.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXpslyjsr0%3D&md5=6aa32cdfdfcbd7fae8fda45a87c93c81CAS | 16048842PubMed |
Fleshner M, Campisi J, Amiri L, Diamond DM (2004) Cat exposure induces both intra- and extracellular Hsp72: The role of adrenal hormones. Psychoneuroendocrinology 29, 1142–1152.
| Cat exposure induces both intra- and extracellular Hsp72: The role of adrenal hormones.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXlt1Wjtrs%3D&md5=62fa145d248161ce1b66ba89a3c493e9CAS | 15219638PubMed |
Forst T, Caduff A, Talary M, Weder M, Brandle M, Kann P, Flacke F, Friedrich C, Pfutzner A (2006) Impact of environmental temperature on skin thickness and microvascular blood flow in subjects with and without diabetes. Diabetes Technology & Therapeutics 8, 94–101.
| Impact of environmental temperature on skin thickness and microvascular blood flow in subjects with and without diabetes.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD28%2FosF2lsQ%3D%3D&md5=3e8bd4431be6ffcf8689d4ec0988a6e7CAS |
Gardner GE, Smith G, Pethick DW (1998) Effect of chromium chelavite supplementation on the metabolism of glycogen and lipid in adult Merino sheep. Australian Journal of Agricultural Research 49, 137–146.
| Effect of chromium chelavite supplementation on the metabolism of glycogen and lipid in adult Merino sheep.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXjsFSmtw%3D%3D&md5=f1b0d7566b58617acc8ca02cebf908fcCAS |
Gardner GE, Jacob RH, Pethick DW (2001) The effect of magnesium oxide supplementation on muscle glycogen metabolism before and after exercise and at slaughter in sheep. Australian Journal of Agricultural Research 52, 723–729.
| The effect of magnesium oxide supplementation on muscle glycogen metabolism before and after exercise and at slaughter in sheep.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXlvVCrsLo%3D&md5=0a21a1ae3735fa5f530f7108a76fb380CAS |
Godsil BP, Quinn JJ, Fanselow MS (2000) Body temperature as a conditional response measure for Pavlovian fear conditioning. Learning & Memory (Cold Spring Harbor, N.Y.) 7, 353–356.
| Body temperature as a conditional response measure for Pavlovian fear conditioning.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD3M%2FivFegtg%3D%3D&md5=a24a63fab0e7e9164ad1966c48cfb73aCAS |
Goodpaster BH, Thaete FL, Jean-Aime S, Kelley DE (1997) Subcutaneous abdominal fat and thigh muscle composition predict insulin sensitivity independently of visceral fat. Diabetes 46, 1579–1585.
| Subcutaneous abdominal fat and thigh muscle composition predict insulin sensitivity independently of visceral fat.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXmsVWnuro%3D&md5=0d29151cb7b98ae2ee3a38ab73b9992dCAS | 9313753PubMed |
Gregory NG (2010) How climatic changes could affect meat quality. Food Research International 43, 1866–1873.
| How climatic changes could affect meat quality.Crossref | GoogleScholarGoogle Scholar |
Gummow B, Botha CJ, Noordhuizen JPTM, Heesterbeek JAP (2005) The public health implications of farming cattle in areas with high background concentrations of vanadium. Preventive Veterinary Medicine 72, 281–290.
| The public health implications of farming cattle in areas with high background concentrations of vanadium.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2MnhvVejuw%3D%3D&md5=8e49da6f1e9d33ca5d8caa70beb2e39eCAS | 16140407PubMed |
Gupte AA, Bomhoff GL, Geiger PC (2008) Age-related differences in skeletal muscle insulin signaling: The role of stress kinases and heat shock proteins. Journal of Applied Physiology 105, 839–848.
| Age-related differences in skeletal muscle insulin signaling: The role of stress kinases and heat shock proteins.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhtFygs7fN&md5=b5bbea04a82b8484d7ed443bcaf5db9eCAS | 18599680PubMed |
Gupte AA, Bomhoff GL, Touchberry CD, Geiger PC (2011) Acute heat treatment improves insulin-stimulated glucose uptake in aged skeletal muscle. Journal of Applied Physiology 110, 451–457.
| Acute heat treatment improves insulin-stimulated glucose uptake in aged skeletal muscle.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXis1Cisrs%3D&md5=c252cc1ec4fe5591dbcdefc7fc17679fCAS | 21148343PubMed |
Henry BA, Goding JW, Alexander WS, Tilbrook AJ, Canny BJ, Dunshea F, Rao A, Mansell A, Clarke IJ (1999) Central administration of leptin to ovariectomized ewes inhibits food intake without affecting the secretion of hormones from the pituitary gland: Evidence for a dissociation of effects on appetite and neuroendocrine function. Endocrinology 140, 1175–1182.
Henry BA, Tilbrook AJ, Dunshea FR, Rao A, Blache D, Martin GB, Clarke IJ (2000) Long-term alterations in adiposity affect the expression of melanin-concentrating hormone and enkephalin but not proopiomelanocortin in the hypothalamus of ovariectomized ewes. Endocrinology 141, 1506–1514.
Henry BA, Goding JW, Tilbrook AJ, Dunshea FR, Blache D, Clarke IJ (2004) Leptin-mediated effects of undernutrition or fasting on luteinizing hormone and growth hormone secretion in ovariectomized ewes depend on the duration of metabolic perturbation. Journal of Neuroendocrinology 16, 244–255.
| Leptin-mediated effects of undernutrition or fasting on luteinizing hormone and growth hormone secretion in ovariectomized ewes depend on the duration of metabolic perturbation.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXivVyltLw%3D&md5=fa6c271ebdcaa8658491b3afdb28a6f9CAS | 15049855PubMed |
Henry BA, Dunshea FR, Gould M, Clarke IJ (2008) Profiling postprandial thermogenesis in muscle and fat of sheep and the central effect of leptin administration. Endocrinology 149, 2019–2026.
| Profiling postprandial thermogenesis in muscle and fat of sheep and the central effect of leptin administration.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXktVeqtr0%3D&md5=b7af09d5d2b27cf1c23f99e51fef4249CAS | 18162524PubMed |
Henry BA, Andrews ZB, Rao A, Clarke IJ (2011) Central leptin activates mitochondrial function and increases heat production in skeletal muscle. Endocrinology 152, 2609–2618.
| Central leptin activates mitochondrial function and increases heat production in skeletal muscle.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXosFaisr0%3D&md5=6cb08a316e6438b16d516816c57f21dfCAS | 21558317PubMed |
Hersom MJ, Horn GW, Krehbiel CR, Phillips WA (2004a) Effect of live weight gain of steers during winter grazing: I. Feedlot performance, carcass characteristics, and body composition of beef steers. Journal of Animal Science 82, 262–272.
Hersom MJ, Krehbiel CR, Horn GW (2004b) Effect of live weight gain of steers during winter grazing: II. Visceral organ mass, cellularity, and oxygen consumption. Journal of Animal Science 82, 184–197.
Hersom MJ, Wettemann RP, Krehbiel CR, Horn GW, Keisler DH (2004c) Effect of live weight gain of steers during winter grazing: III. Blood metabolites and hormones during feedlot finishing. Journal of Animal Science 82, 2059–2068.
Hua Y, Clark S, Ren J, Sreejayan N (2012) Molecular mechanisms of chromium in alleviating insulin resistance. The Journal of Nutritional Biochemistry 23, 313–319.
| Molecular mechanisms of chromium in alleviating insulin resistance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xjslelsr4%3D&md5=245eca730f1e90b29db92edc2c87230fCAS | 22423897PubMed |
Huang SY, Lee WC, Chen MY, Wang SC, Huang CH, Tsou HL, Lin EC (2004) Genotypes of 5′-flanking region in porcine heat-shock protein 70.2 gene affect backfat thickness and growth performance in Duroc boars. Livestock Production Science 85, 181–187.
| Genotypes of 5′-flanking region in porcine heat-shock protein 70.2 gene affect backfat thickness and growth performance in Duroc boars.Crossref | GoogleScholarGoogle Scholar |
Huang QC, Xu ZR, Han XY, Li WF (2006) Changes in hormones, growth factor and lipid metabolism in finishing pigs fed betaine. Livestock Science 105, 78–85.
| Changes in hormones, growth factor and lipid metabolism in finishing pigs fed betaine.Crossref | GoogleScholarGoogle Scholar |
Huang QC, Xu ZR, Han XY, Li WF (2007) Effect of betaine on growth hormone pulsatile secretion and serum metabolites in finishing pigs. Journal of Animal Physiology and Animal Nutrition 91, 85–90.
| Effect of betaine on growth hormone pulsatile secretion and serum metabolites in finishing pigs.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXksFWqs78%3D&md5=843ed5645cb597fa8fb6f16150c075ffCAS | 17355337PubMed |
Ingvartsen KL, Boisclair YR (2001) Leptin and the regulation of food intake, energy homeostasis and immunity with special focus on periparturient ruminants. Domestic Animal Endocrinology 21, 215–250.
| Leptin and the regulation of food intake, energy homeostasis and immunity with special focus on periparturient ruminants.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XhsFWgu7o%3D&md5=f543a458bba8d6c484701a8ca5cb3c20CAS | 11872319PubMed |
Jacob RH, Hopkins DL (2014) Techniques to reduce the temperature of beef muscle early in the post mortem period – a review. Animal Production Science 54, 482–493.
| Techniques to reduce the temperature of beef muscle early in the post mortem period – a review.Crossref | GoogleScholarGoogle Scholar |
Jacob RH, Surridge VSM, Beatty DT, Gardner GE, Warner RD (2014) Grain feeding increases core body temperature of beef cattle. Animal Production Science 54, 444–449.
| Grain feeding increases core body temperature of beef cattle.Crossref | GoogleScholarGoogle Scholar |
Jentjens RLPG, Wagenmakers AJM, Jeukendrup AE (2002) Heat stress increases muscle glycogen use but reduces the oxidation of ingested carbohydrates during exercise. Journal of Applied Physiology 92, 1562–1572.
Jentjens RLPG, Underwood K, Achten J, Currell K, Mann CH, Jeukendrup AE (2006) Exogenous carbohydrate oxidation rates are elevated after combined ingestion of glucose and fructose during exercise in the heat. Journal of Applied Physiology 100, 807–816.
Kathirvel E, Morgan K, Nandgiri G, Sandoval BC, Caudill MA, Bottiglieri T, French SW, Morgan TR (2010) Betaine improves nonalcoholic fatty liver and associated hepatic insulin resistance: A potential mechanism for hepatoprotection by betaine. American Journal of Physiology. Gastrointestinal and Liver Physiology 299, G1068–G1077.
| Betaine improves nonalcoholic fatty liver and associated hepatic insulin resistance: A potential mechanism for hepatoprotection by betaine.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhsFSnsr7I&md5=e722a022d2cfed69bba8fceeea4d211dCAS | 20724529PubMed |
Kavanagh K, Zhang L, Wagner JD (2009) Tissue-specific regulation and expression of heat shock proteins in type 2 diabetic monkeys. Cell Stress & Chaperones 14, 291–299.
| Tissue-specific regulation and expression of heat shock proteins in type 2 diabetic monkeys.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXjvV2gt78%3D&md5=716509fb8e3b8379a42270da1b52f4ccCAS |
Kawachi H (2006) Micronutrients affecting adipogenesis in beef cattle. Animal Science Journal 77, 463–471.
| Micronutrients affecting adipogenesis in beef cattle.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28Xht1egs7jE&md5=12befc0eb5c9b8a88ccf07a3ffa39a72CAS |
Kegley EB, Spears JW, Brown TT (1997a) Effect of shipping and chromium supplementation on performance, immune response, and disease resistance of steers. Journal of Animal Science 75, 1956–1964.
Kegley EB, Spears JW, Eisemann JH (1997b) Performance and glucose metabolism in calves fed a chromium–nicotinic acid complex or chromium chloride. Journal of Dairy Science 80, 1744–1750.
| Performance and glucose metabolism in calves fed a chromium–nicotinic acid complex or chromium chloride.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXls1Kqtbo%3D&md5=9909cc3836d945d839829d872430bcb3CAS | 9276815PubMed |
Kegley EB, Galloway DL, Fakler TM (2000) Effect of dietary chromium-l-methionine on glucose metabolism of beef steers. Journal of Animal Science 78, 3177–3183.
Kellogg DL, Zhao JL, Wu Y (2008) Neuronal nitric oxide synthase control mechanisms in the cutaneous vasculature of humans in vivo. The Journal of Physiology 586, 847–857.
| Neuronal nitric oxide synthase control mechanisms in the cutaneous vasculature of humans in vivo.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXit1ynu7s%3D&md5=e5c83b87317aaccf402a6cd41cc66422CAS | 18048451PubMed |
Kietzmann M, Jablonski H (1985) Blocking of stress in swine with magnesium aspartate hydrochloride. Praktischer Tierarzt 661, 331–335.
Kim C, Park J, Park J, Kang E, Ahn C, Cha B, Lim S, Kim K, Lee H (2006) Comparison of body fat composition and serum adiponectin levels in diabetic obesity and non-diabetic obesity. Obesity (Silver Spring, Md.) 14, 1164–1171.
| Comparison of body fat composition and serum adiponectin levels in diabetic obesity and non-diabetic obesity.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XpslSqsLk%3D&md5=a68ac70aa8758d1856d9d396a43aee7eCAS |
Kim YHB, Warner RD, Rosenvold K (2014) Influence of high pre-rigor temperature and fast pH fall on muscle proteins and meat quality: a review. Animal Production Science 54, 375–395.
| Influence of high pre-rigor temperature and fast pH fall on muscle proteins and meat quality: a review.Crossref | GoogleScholarGoogle Scholar |
King YT, Lin CS, Lin JH, Lee WC (2002) Whole-body hyperthermia-induced thermotolerance is associated with the induction of heat shock protein 70 in mice. The Journal of Experimental Biology 205, 273–278.
Klasing KC, Adler KL, Remus JC, Calvert CC (2002) Dietary betaine increases intraepithelial lymphocytes in the duodenum of coccidia-infected chicks and increases functional properties of phagocytes. The Journal of Nutrition 132, 2274–2282.
Komatsu T, Itoh F, Hodate K, Hazegawa S, Obara Y, Kushibiki S (2005) Gene expression of resistin and TNF-alpha in adipose tissue of Japanese Black steers and Holstein steers. Animal Science Journal 76, 567–573.
| Gene expression of resistin and TNF-alpha in adipose tissue of Japanese Black steers and Holstein steers.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhslWmsrs%3D&md5=defec5848b67ef2c53812adffa8ac453CAS |
Komiya H, Mori Y, Yokose T, Kurokawa N, Horie N, Tajima N (2006) Effect of intramuscular fat difference on glucose and insulin reaction in oral glucose tolerance test. Journal of Atherosclerosis and Thrombosis 13, 136–142.
| Effect of intramuscular fat difference on glucose and insulin reaction in oral glucose tolerance test.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XnvVyrsr8%3D&md5=bc274bb0a17761af8a53b23392419a38CAS | 16835468PubMed |
Kovacevic Z, McGivan JD (1983) Mitochondrial metabolism of glutamine and glutamate and its physiological significance. Physiological Reviews 63, 547–605.
Kozlowski S, Brzezinska Z, Kruk B, Kaciuba-Uscilko H, Greenleaf JE, Nazar K (1985) Exercise hyperthermia as a factor limiting physical performance: Temperature effect on muscle metabolism. Journal of Applied Physiology 59, 766–773.
Krentz AJ, Bailey CJ (2005) Oral antidiabetic agents: Current role in type 2 diabetes mellitus. Drugs 65, 385–411.
| Oral antidiabetic agents: Current role in type 2 diabetes mellitus.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXit1Grsbg%3D&md5=6b5dc63f90dd818d42da7d4998b955deCAS | 15669880PubMed |
Kurucz I, Morva A, Vaag A, Eriksson K-F, Huang X, Groop L, Koranyi L (2002) Decreased expression of heat shock protein 72 in skeletal muscle of patients with type 2 diabetes correlates with insulin resistance. Diabetes 51, 1102–1109.
| Decreased expression of heat shock protein 72 in skeletal muscle of patients with type 2 diabetes correlates with insulin resistance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38Xis1Oju7k%3D&md5=f1d70295ece8ae6dfae25de90a82db5aCAS | 11916932PubMed |
Kushibiki S, Hodate K, Shingu H, Ueda Y, Shinoda M, Mori Y, Itoh T, Yokomizo Y (2001) Insulin resistance induced in dairy steers by tumor necrosis factor alpha is partially reversed by 2,4–thiazolidinedione. Domestic Animal Endocrinology 21, 25–37.
| Insulin resistance induced in dairy steers by tumor necrosis factor alpha is partially reversed by 2,4–thiazolidinedione.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXmt1KltLk%3D&md5=47a664b93259bde06478559b44937f3dCAS | 11524172PubMed |
Lacetera N, Bernabucci U, Basiricò L, Morera P, Nardone A (2009) Heat shock impairs DNA synthesis and down-regulates gene expression for leptin and Ob-Rb receptor in concanavalin a-stimulated bovine peripheral blood mononuclear cells. Veterinary Immunology and Immunopathology 127, 190–194.
| Heat shock impairs DNA synthesis and down-regulates gene expression for leptin and Ob-Rb receptor in concanavalin a-stimulated bovine peripheral blood mononuclear cells.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhsFChtbbF&md5=04e7e120b2059ad7afba9eb38a8938faCAS | 18992945PubMed |
Leury BJ, Baumgard LH, Block SS, Segoale N, Ehrhardt RA, Rhoads RP, Bauman DE, Bell AW, Boisclair YR (2003) Effect of insulin and growth hormone on plasma leptin in periparturient dairy cows. The American Journal of Physiology 285, R1107–R1115.
Lindquist S (1986) The heat-shock response. Annual Review of Biochemistry 55, 1151–1191.
| The heat-shock response.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL28XltV2lsrY%3D&md5=deda7c87875608198035c2a8c752dc49CAS | 2427013PubMed |
Lorenz I (2000) Retrospective study of serum glucose concentration in cattle with mucosal disease. Journal of Veterinary Medicine A 47, 489–493.
| Retrospective study of serum glucose concentration in cattle with mucosal disease.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD3M%2FmslensA%3D%3D&md5=70df8846a27d0b74b9ba77d9c1d40d7fCAS |
Lowell BB, Spiegelman BM (2000) Towards a molecular understanding of adaptive thermogenesis. Nature 404, 652–660.
Loxton I, Grant TP, Reid DJ, Lawrence RJ (2007) Effects of a supplement containing betaine on feedlot steers exposed to a heat load. In ‘Recent Advances in Animal Nutrition in Australia’, Armidale, NSW, Australia. (Eds P Cronje and N Richards) pp. 201–210. (University of New England: Armidale, NSW)
Matthews JO, Southern LL (2000) The effect of dietary betaine in Eimeria acervulina-infected chicks. Poultry Science 79, 60–65.
Matthews JO, Southern LL, Bidner TD (2001) Estimation of the total sulfur amino acid requirement and the effect of betaine in diets deficient in total sulfur amino acids for the weanling pig. Journal of Animal Science 79, 1557–1565.
McCann JP, Ullmann MB, Temple MR, Reimers TJ, Bergman EN (1986) Insulin and glucose responses to glucose injection in fed and fasted obese and lean sheep. The Journal of Nutrition 116, 1287–1297.
McCann JP, Bergman EN, Reimers TJ (1989) Effects of obesity and ovarian steroids on insulin secretion and removal in sheep. American Journal of Physiology. Endocrinology and Metabolism 256, E116–E128.
McCann JP, Loo SC, Aalseth DL, Abribat T (1997) Differential effects of GH stimulation on fasting and prandial metabolism and plasma IGFs and IGF-binding proteins in lean and obese sheep. The Journal of Endocrinology 154, 329–346.
| Differential effects of GH stimulation on fasting and prandial metabolism and plasma IGFs and IGF-binding proteins in lean and obese sheep.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXltFGqu7c%3D&md5=1381e22dc76f21465ebd16613715a89bCAS | 9291844PubMed |
McCarty MF (2006) Induction of heat shock proteins may combat insulin resistance. Medical Hypotheses 66, 527–534.
| Induction of heat shock proteins may combat insulin resistance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XjvVGluw%3D%3D&md5=e7adca9c76645c3673cf39fef95445daCAS | 16309849PubMed |
McNamara JP (1991) Regulation of adipose tissue metabolism in support of lactation. Journal of Dairy Science 74, 706–719.
| Regulation of adipose tissue metabolism in support of lactation.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3MXitVCitrw%3D&md5=94a8da7947d68d3fd2dc137e338e447cCAS | 2045568PubMed |
Mertz W, Roginski EE (1969) Effects of chromium(III) supplementation on growth and survival under stress in rats fed low protein diets. The Journal of Nutrition 97, 531–536.
Montgomery JL, King MB, Gentry JG, Barham AR, Barham BL, Hilton GG, Blanton JR, Horst RL, Galyean ML, Morrow KJ, Wester DB, Miller MF (2004) Supplemental vitamin D-3 concentration and biological type of steers. II. Tenderness, quality, and residues of beef. Journal of Animal Science 82, 2092–2104.
Mooney KW, Cromwell GL (1995) Effects of dietary chromium picolinate supplementation on growth, carcass characteristics, and accretion rates of carcass tissues in growing-finishing swine. Journal of Animal Science 73, 3351–3357.
Moran DS, Eli-Berchoer L, Heled Y, Mendel L, Schocina M, Horowitz M (2006) Heat intolerance: Does gene transcription contribute? Journal of Applied Physiology 100, 1370–1376.
Morera P, Basiricò L, Hosoda K, Bernabucci U (2012) Chronic heat stress up-regulates leptin and adiponectin secretion and expression and improves leptin, adiponectin and insulin sensitivity in mice. Journal of Molecular Endocrinology 48, 129–138.
| Chronic heat stress up-regulates leptin and adiponectin secretion and expression and improves leptin, adiponectin and insulin sensitivity in mice.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XlvFOkt7k%3D&md5=a67f9fe34801f090d97441ab88f4c3eaCAS | 22279186PubMed |
Morrison AL, Dinges M, Singleton KD, Odoms K, Wong HR, Wischmeyer PE (2006) Glutamine’s protection against cellular injury is dependent on heat shock factor-1. American Journal of Physiology. Cell Physiology 290, C1625–C1632.
| Glutamine’s protection against cellular injury is dependent on heat shock factor-1.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XlvVejtb4%3D&md5=dcbe4709ecf0bf19861cac164c6d9ba4CAS | 16436470PubMed |
Niemack EA, Stockli F, Hausmann E, Sanderegger J, Classen HG, Helbig J (1979) Einflub von magnesium-aspartat-hydrochorid auf kannibalisms, transportsterb und den elec-trolytgehalt im herzen von schueinen. Magnesium Bulletin 3, 195–198.
Nikkhah A, Mirzaei M, Khorvash M, Rahmani HR, Ghorbani GR (2011) Chromium improves production and alters metabolism of early lactation cows in summer. Journal of Animal Physiology and Animal Nutrition 95, 81–89.
| Chromium improves production and alters metabolism of early lactation cows in summer.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXit1ersrc%3D&md5=275c3e330cad6466d51ffc688101c989CAS | 20579184PubMed |
Ohtsuka Y, Yabunaka N, Watanabe I, Noro H, Fujisawa H, Agishi Y (1995) Thermal stress and diabetic complications. International Journal of Biometeorology 38, 57–59.
| Thermal stress and diabetic complications.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK2M3htlWgsw%3D%3D&md5=3735ead561dac90038399529f45fc9f0CAS | 7698855PubMed |
Page TG, Southern LL, Ward TL, Thompson DL (1993) Effect of chromium picolinate on growth and serum and carcass traits of growing-finishing pigs. Journal of Animal Science 71, 656–662.
Pardon M-C, Kendall DA, Perez-Diaz F, Duxon MS, Marsden CA (2004) Repeated sensory contact with aggressive mice rapidly leads to an anticipatory increase in core body temperature and physical activity that precedes the onset of aversive responding. The European Journal of Neuroscience 20, 1033–1050.
| Repeated sensory contact with aggressive mice rapidly leads to an anticipatory increase in core body temperature and physical activity that precedes the onset of aversive responding.Crossref | GoogleScholarGoogle Scholar | 15305872PubMed |
Parkin JM, Carey MF, Zhao S, Febbraio MA (1999) Effect of ambient temperature on human skeletal muscle metabolism during fatiguing submaximal exercise. Journal of Applied Physiology 86, 902–908.
Patrick L (2002) Nonalcoholic fatty liver disease: Relationship to insulin sensitivity and oxidative stress. Treatment approaches using vitamin e, magnesium, and betaine. Alternative Medicine Review 7, 276–291.
Pearce KL, Masters DG, Jacob RH, Hopkins DL, Pethick DW (2008) Effects of sodium chloride and betaine on hydration status of lambs at slaughter. Australian Journal of Experimental Agriculture 48, 1194–1200.
| Effects of sodium chloride and betaine on hydration status of lambs at slaughter.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXptlKlu7g%3D&md5=b9cc1d406eec2f985d6f06136848e487CAS |
Pethick D, Harman N, Chong J (1987) Non-esterified long-chain fatty acid metabolism in fed sheep at rest and during exercise. Australian Journal of Biological Sciences 40, 221–234.
Petrofsky JS, Lee S, Cuneo-Libarona M (2005) The impact of rosiglitazone on heat tolerance in patients with type 2 diabetes. Medical Science Monitor 11, CR562–CR569.
Pighin DG, Brown W, Ferguson DM, Fisher AD, Warner RD (2014) Relationship between changes in core body temperature in lambs and post-slaughter muscle glycogen content and dark-cutting. Animal Production Science 54, 459–463.
| Relationship between changes in core body temperature in lambs and post-slaughter muscle glycogen content and dark-cutting.Crossref | GoogleScholarGoogle Scholar |
Ponnampalam EN, Warner RD, Suster D, Dunshea FR (2005) Breed and nutrition influence the responses to homeostatic signals in lambs. Asia Pacific Journal of Clinical Nutrition 14, S79
Rahmouni K, Morgan DA (2007) Hypothalamic arcuate nucleus mediates the sympathetic and arterial pressure responses to leptin. Hypertension 49, 647–652.
| Hypothalamic arcuate nucleus mediates the sympathetic and arterial pressure responses to leptin.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhslCmsLg%3D&md5=2b31b5ae73b23a86e94040af8b93f89aCAS | 17190874PubMed |
Reis AF, Hauache OM, Velho G (2005) Vitamin d endocrine system and the genetic susceptibility to diabetes, obesity and vascular disease. A review of evidence. Diabetes & Metabolism 31, 318–325.
| Vitamin d endocrine system and the genetic susceptibility to diabetes, obesity and vascular disease. A review of evidence.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXhtFGrurjI&md5=35b8e14f3ff7a3d9986c8a9c86d60a92CAS |
Rhoads ML, Rhoads RP, VanBaale MJ, Collier RJ, Sanders SR, Weber WJ, Crooker BA, Baumgard LH (2009) Effects of heat stress and plane of nutrition on lactating holstein cows: I. Production, metabolism, and aspects of circulating somatotropin. Journal of Dairy Science 92, 1986–1997.
| Effects of heat stress and plane of nutrition on lactating holstein cows: I. Production, metabolism, and aspects of circulating somatotropin.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXlsFynu7k%3D&md5=0ee8f43bfb3f9711856e5007a93e274fCAS | 19389956PubMed |
Richardson EC, Herd RM, Archer JA, Arthur PF (2004) Metabolic differences in angus steers divergently selected for residual feed intake. Australian Journal of Experimental Agriculture 44, 441–452.
| Metabolic differences in angus steers divergently selected for residual feed intake.Crossref | GoogleScholarGoogle Scholar |
Roh S-G, Hishikawa D, Hong Y-H, Sasaki S (2006) Control of adipogenesis in ruminants. Animal Science Journal 77, 472–477.
| Control of adipogenesis in ruminants.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28Xht1egs7jF&md5=ee848ca66362ba1016510b0b0d22b4d4CAS |
Ropeleski MJ, Riehm J, Baer KA, Musch MW, Chang EB (2005) Anti-apoptotic effects of l-glutamine-mediated transcriptional modulation of the heat shock protein 72 during heat shock. Gastroenterology 129, 170–184.
| Anti-apoptotic effects of l-glutamine-mediated transcriptional modulation of the heat shock protein 72 during heat shock.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXntVGrsb8%3D&md5=6b0202a8d51695fb1715625b2d1718cdCAS | 16012946PubMed |
Rowell LB (1974) Human cardiovascular adjustments to exercise and thermal stress. Physiological Reviews 54, 75–159.
Ruell PA, Hoffman KM, Chow CM, Thompson MW (2004) Effect of temperature and duration of hyperthermia on HSP72 induction in rat tissues. Molecular and Cellular Biochemistry 267, 187–194.
| Effect of temperature and duration of hyperthermia on HSP72 induction in rat tissues.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXhtVais7vI&md5=47d65b56940c5f758116545da8d34b66CAS | 15663200PubMed |
Ruis MAW, te Brake JHA, Engel B, Buist WG, Blokhuis HJ, Koolhaas JM (2001) Adaptation to social isolation: Acute and long-term stress responses of growing gilts with different coping characteristics. Physiology & Behavior 73, 541–551.
| Adaptation to social isolation: Acute and long-term stress responses of growing gilts with different coping characteristics.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXlslGnu7c%3D&md5=03dd1032400f4c7fe97b0f6a6e26c838CAS |
Rustenbeck I (2007) Unconventional antidiabetic agents. Medizinische Monatsschrift fur Pharmazeuten 30, 131–137.
Sasaki S, Rho S, Choi K (2002) Regulatory mechanism for proliferation and differentiation in ovine preadipocytes and mature adipocytes. Proceedings of Japanese Society for Animal Nutrition and Metabolism 46, 101–119.
Schoenberg KM, Perfield KL, Farney JK, Bradford BJ, Boisclair YR, Overton TR (2011) Effects of prepartum 2,4-thiazolidinedione on insulin sensitivity, plasma concentrations of tumor necrosis factor-α and leptin, and adipose tissue gene expression. Journal of Dairy Science 94, 5523–5532.
| Effects of prepartum 2,4-thiazolidinedione on insulin sensitivity, plasma concentrations of tumor necrosis factor-α and leptin, and adipose tissue gene expression.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhtl2qsLrM&md5=e864cd5e3e37b00b832b6ca477a7986dCAS | 22032375PubMed |
Schrama JW, Heetkamp MJW, Simmins PH, Gerrits WJJ (2003) Dietary betaine supplementation affects energy metabolism of pigs. Journal of Animal Science 81, 1202–1209.
Shelton JL, Payne RL, Johnston SL, Bidner TD, Southern LL, Odgaard RL, Page TG (2003) Effect of chromium propionate on growth, carcass traits, pork quality, and plasma metabolites in growing-finishing pigs. Journal of Animal Science 81, 2515–2524.
Shukla R, Bhonde RR (2008) Adipogenic action of vanadium: A new dimension in treating diabetes. Biometals 21, 205–210.
| Adipogenic action of vanadium: A new dimension in treating diabetes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXjsFajtr4%3D&md5=f7184fecde1bef4da88e962a240435c0CAS | 17671828PubMed |
Shwartz G, Rhoads ML, VanBaale MJ, Rhoads RP, Baumgard LH (2009) Effects of a supplemental yeast culture on heat-stressed lactating Holstein cows. Journal of Dairy Science 92, 935–942.
| Effects of a supplemental yeast culture on heat-stressed lactating Holstein cows.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXivVakt70%3D&md5=e6d1ca142212c02dbd297b3c4f53cd99CAS | 19233786PubMed |
Singleton KD, Wischmeyer PE (2006) Oral glutamine enhances heat shock protein expression and improves survival following hyperthermia. Shock (Augusta, Ga.) 25, 295–299.
| Oral glutamine enhances heat shock protein expression and improves survival following hyperthermia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28Xis1amsr4%3D&md5=0fefb22568f1ed78b390cf99ecfa04e5CAS |
Sivitz WI, Fink BD, Donohoue PA (1999) Fasting and leptin modulate adipose and muscle uncoupling protein: Divergent effects between messenger ribonucleic acid and protein expression. Endocrinology 140, 1511–1519.
Smith KL, Stebulis SE, Waldron MR, Overton TR (2007) Prepartum 2,4-thiazolidinedione alters metabolic dynamics and dry matter intake of dairy cows. Journal of Dairy Science 90, 3660–3670.
| Prepartum 2,4-thiazolidinedione alters metabolic dynamics and dry matter intake of dairy cows.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXot1OltL4%3D&md5=f1ac8ef1e73b6f37806d1ba8ba5034b5CAS | 17638977PubMed |
Soukas A, Cohen P, Socci ND, Friedman JM (2000) Leptin-specific patterns of gene expression in white adipose tissue. Genes & Development 14, 963–980.
Spears JW, Whisnant CS, Huntington GB, Lloyd KE, Fry RS, Krafka K, Lamptey A, Hyda J (2012) Chromium propionate enhances insulin sensitivity in growing cattle. Journal of Dairy Science 95, 2037–2045.
| Chromium propionate enhances insulin sensitivity in growing cattle.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xkslaqsbg%3D&md5=0277db377aed08ef016c851b53083ea6CAS | 22459849PubMed |
Steele NC, Rosebrough RW (1979) Trivalent chromium and nicotinic acid supplementation for the turkey poult. Poultry Science 58, 983–984.
| Trivalent chromium and nicotinic acid supplementation for the turkey poult.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE1MXkvV2qurY%3D&md5=850f909a1c8f61a320d7bea1083e7979CAS |
Steele NC, Althen TG, Frobish LT (1977) Biological activity of glucose tolerance factor in swine. Journal of Animal Science 45, 1341–1345.
Suster D, Leury BJ, King RH, Mottram M, Dunshea FR (2004) Interrelationships between porcine somatotropin (pST), betaine, and energy level on body composition and tissue distribution of finisher boars. Australian Journal of Agricultural Research 55, 983–990.
| Interrelationships between porcine somatotropin (pST), betaine, and energy level on body composition and tissue distribution of finisher boars.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXnvVKiurk%3D&md5=547e84d4cbe64a23b882b2e9501680e4CAS |
Swanek SS, Morgan JB, Owens FN, Gill DR, Strasia CA, Dolezal HG, Ray FK (1999) Vitamin D3 supplementation of beef steers increases longissimus tenderness. Journal of Animal Science 77, 874–881.
Swiecki C, Stojadinovic A, Anderson J, Zhao A, Dawson H, Shea-Donohue T (2003) Effect of hyperglycemia and nitric oxide synthase inhibition on heat tolerance and induction of heat shock protein 72 kDa in vivo. The American Surgeon 69, 587–592.
Takakazu O, Kae O, Tetsuro H (2001) Mechanisms and mediators of psychological stress-induced rise in core temperature. Psychosomatic Medicine 63, 476–486.
Tanaka S, Takahashi E, Matsui T, Yano H (2001) Zinc promotes adipocyte differentiation in vitro. Asian-Australasian Journal of Animal Sciences 14, 966–969.
Tsiotra PC, Tsigos C (2006) Stress, the endoplasmic reticulum, and insulin resistance. Annals of the New York Academy of Sciences 1083, 63–76.
| Stress, the endoplasmic reticulum, and insulin resistance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXmslKrsw%3D%3D&md5=9059595d3cb1408f367a75653eabad12CAS | 17148734PubMed |
Uyanik F, Atasever A, Özdamar S, Aydin F (2002) Effects of dietary chromium chloride supplementation on performance, some serum parameters, and immune response in broilers. Biological Trace Element Research 90, 99–115.
| Effects of dietary chromium chloride supplementation on performance, some serum parameters, and immune response in broilers.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXntVKrtg%3D%3D&md5=2ab5e7dbae38659509705fc29b3f3db1CAS | 12666829PubMed |
Vamvakopoulos NC, Fukuhara K, Patchev V, Chrousos GP (1993) Effect of single and repeated immobilization stress on the heat shock protein 70/90 system of the rat–glucocorticoid-independent reversible reduction of Hsp90 in the liver and spleen. Neuroendocrinology 57, 1057–1065.
| Effect of single and repeated immobilization stress on the heat shock protein 70/90 system of the rat–glucocorticoid-independent reversible reduction of Hsp90 in the liver and spleen.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK2c%2FltVeqsg%3D%3D&md5=0511bc2426ebddf071ff5238fec8bbc9CAS | 8232764PubMed |
Vianna DML, Carrive P (2005) Changes in cutaneous and body temperature during and after conditioned fear to context in the rat. The European Journal of Neuroscience 21, 2505–2512.
| Changes in cutaneous and body temperature during and after conditioned fear to context in the rat.Crossref | GoogleScholarGoogle Scholar |
Wang RL (2006) Influence of different dietary levels of zinc on performance, vitamin B-12, and blood parameters in lambs. International Journal for Vitamin and Nutrition Research 76, 353–358.
| Influence of different dietary levels of zinc on performance, vitamin B-12, and blood parameters in lambs.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXkvVSgt74%3D&md5=730818e10eac575c7733696bafaa729eCAS |
Wang SJ, Chen HW, Huang MH, Yang RC (2007) Previous heat shock facilitates the glutamine-induced expression of heat-shock protein 72 in septic liver. Nutrition (Burbank, Los Angeles County, Calif.) 23, 582–588.
| Previous heat shock facilitates the glutamine-induced expression of heat-shock protein 72 in septic liver.Crossref | GoogleScholarGoogle Scholar |
Wang C, Liu Q, Yang WZ, Wu J, Zhang WW, Zhang P, Dong KH, Huang YX (2010a) Effects of betaine supplementation on rumen fermentation, lactation performance, feed digestibilities and plasma characteristics in dairy cows. The Journal of Agricultural Science 148, 487–495.
| Effects of betaine supplementation on rumen fermentation, lactation performance, feed digestibilities and plasma characteristics in dairy cows.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXotFeisbw%3D&md5=729ca643d452fb5205416d523dab44b5CAS |
Wang Z, Yao T, Pini M, Zhou Z, Fantuzzi G, Song Z (2010b) Betaine improved adipose tissue function in mice fed a high fat diet: A mechanism for hepatoprotective effect of betaine in nonalcholic fatty liver disease. American Journal of Physiology. Gastrointestinal and Liver Physiology 298, G634–G642.
| Betaine improved adipose tissue function in mice fed a high fat diet: A mechanism for hepatoprotective effect of betaine in nonalcholic fatty liver disease.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXmtlWjtLs%3D&md5=ce02c5bf39194cba6d7d85482085956fCAS | 20203061PubMed |
Warner RD, Ferguson DM, McDonagh MB, Channon HA, Cottrell JJ, Dunshea FR (2005) Acute exercise stress and electrical stimulation influence the consumer perception of sheep meat eating quality and objective quality traits. Australian Journal of Experimental Agriculture 45, 553–560.
| Acute exercise stress and electrical stimulation influence the consumer perception of sheep meat eating quality and objective quality traits.Crossref | GoogleScholarGoogle Scholar |
Warner RD, Dunshea FR, Gutzke D, Lau J, Kearney G (2014a) Factors influencing the incidence of high rigor temperature in beef carcasses in Australia. Animal Production Science 54, 363–374.
| Factors influencing the incidence of high rigor temperature in beef carcasses in Australia.Crossref | GoogleScholarGoogle Scholar |
Warner RD, Thompson JM, Polkinghorne R, Gutzke D, Kearney GA (2014b) A consumer sensory study of the influence of rigor temperature on eating quality and ageing potential of beef striploin and rump. Animal Production Science 54, 396–406.
| A consumer sensory study of the influence of rigor temperature on eating quality and ageing potential of beef striploin and rump.Crossref | GoogleScholarGoogle Scholar |
Welch WJ (1992) Mammalian stress response: Cell physiology, structure/function of stress proteins, and implications for medicine and disease. Physiological Reviews 72, 1063–1081.
Wheelock JB, Rhoads RP, Vanbaale MJ, Sanders SR, Baumgard LH (2010) Effects of heat stress on energetic metabolism in lactating Holstein cows. Journal of Dairy Science 93, 644–655.
| Effects of heat stress on energetic metabolism in lactating Holstein cows.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXht1CjtLY%3D&md5=436ea2252411d8f269eae61d43fc12c7CAS | 20105536PubMed |
Whitham M, Walker GJ, Bishop NC (2006) Effect of caffeine supplementation on the extracellular heat shock protein 72 response to exercise. Journal of Applied Physiology 101, 1222–1227.
| Effect of caffeine supplementation on the extracellular heat shock protein 72 response to exercise.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhtlSksb7J&md5=f89f0b2a275cccb1e06d6bdba8697482CAS | 16794026PubMed |
Wischmeyer PE, Kahana M, Wolfson R, Ren H, Musch MM, Chang EB (2001) Glutamine reduces cytokine release, organ damage, and mortality in a rat model of endotoxemia. Shock (Augusta, Ga.) 16, 398–402.
| Glutamine reduces cytokine release, organ damage, and mortality in a rat model of endotoxemia.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD3MnkslCkug%3D%3D&md5=7ee5021eb1db4f55ba64894072ed7da6CAS |
Xiao XQ, Grove KL, Grayson BE, Smith MS (2004) Inhibition of uncoupling protein expression during lactation: Role of leptin. Endocrinology 145, 830–838.
| Inhibition of uncoupling protein expression during lactation: Role of leptin.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXovFSlsA%3D%3D&md5=0df311190825495093a02a4c2e57a779CAS | 14605003PubMed |
Yano H, Hirai S, Kitagawa M (2004) Recent advances in research for nutritional physiology of beef cattle. In ‘Proceedings of Japanese Society for Animal Nutrition and Metabolism’. pp. 79–104.
Yari M, Nikkhah A, Alikhani M, Khorvash M, Rahmani H, Ghorbani GR (2010) Physiological calf responses to increased chromium supply in summer. Journal of Dairy Science 93, 4111–4120.
| Physiological calf responses to increased chromium supply in summer.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtlCqtrnP&md5=81fb04ad526c86472f9cec7b2fbb1281CAS | 20723686PubMed |
Yaspelkis BB, Scroop GC, Wilmore KM, Ivy JL (1993) Carbohydrate metabolism during exercise in hot and thermoneutral environments. International Journal of Sports Medicine 14, 13–19.
| Carbohydrate metabolism during exercise in hot and thermoneutral environments.Crossref | GoogleScholarGoogle Scholar | 8440539PubMed |
Young AJ (1990) Energy substrate utilization during exercise in extreme environments. Exercise and Sport Sciences Reviews 18, 65–118.
| Energy substrate utilization during exercise in extreme environments.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK3c3pt1Sqsw%3D%3D&md5=ba28a1a290de227e6e22e1f3146fe29dCAS | 2192901PubMed |
Yu J, Tang S, Bao E, Zhang M, Hao Q, Yue Z (2009) The effect of transportation on the expression of heat shock proteins and meat quality of M. longissimus dorsi in pigs. Meat Science 83, 474–478.
| The effect of transportation on the expression of heat shock proteins and meat quality of M. longissimus dorsi in pigs.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhtV2rt7vP&md5=93e1422d5e936830f41c71e45ea51e75CAS | 20416675PubMed |
Zhang J, Matheny MK, Tumer N, Mitchell MK, Scarpace PJ (2007) Leptin antagonist reveals that the normalization of caloric intake and the thermic effect of food after high-fat feeding are leptin dependent. American Journal of Physiology. Regulatory, Integrative and Comparative Physiology 292, R868–R874.
| Leptin antagonist reveals that the normalization of caloric intake and the thermic effect of food after high-fat feeding are leptin dependent.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXjt1Gmsr4%3D&md5=6fcce6923ab34fe6a9bd588d88b5a867CAS | 17023670PubMed |
Zieba DA, Amstalden M, Williams GL (2005) Regulatory roles of leptin in reproduction and metabolism: A comparative review. Domestic Animal Endocrinology 29, 166–185.
| Regulatory roles of leptin in reproduction and metabolism: A comparative review.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXltVehu70%3D&md5=51b171dd869dffff6ab654a44a5536f5CAS | 15927772PubMed |