

Genetics of heifer puberty in two tropical beef genotypes in northern Australia and associations with heifer- and steer-production traits
D. J. Johnston A B F , S. A. Barwick A B , N. J. Corbet A C , G. Fordyce A D , R. G. Holroyd A E , P. J. Williams A C and H. M. Burrow A FA Cooperative Research Centre for Beef Genetic Technologies, University of New England, Armidale, NSW 2351, Australia.
B Animal Genetics and Breeding Unit1, University of New England, Armidale, NSW 2351, Australia.
C CSIRO Livestock Industries, Rockhampton, Qld 4702, Australia.
D Queensland Department of Primary Industries and Fisheries, Charters Towers, Qld 4820, Australia.
E Queensland Department of Primary Industries and Fisheries, Rockhampton, Qld 4702, Australia.
F Corresponding author. Email: djohnsto@une.edu.au
Animal Production Science 49(6) 399-412 https://doi.org/10.1071/EA08276
Submitted: 11 November 2008 Accepted: 17 February 2009 Published: 13 May 2009
Abstract
A total of 2115 heifers from two tropical genotypes (1007 Brahman and 1108 Tropical Composite) raised in four locations in northern Australia were ovarian-scanned every 4–6 weeks to determine the age at the first-observed corpus luteum (CL) and this was used to define the age at puberty for each heifer. Other traits recorded at each time of ovarian scanning were liveweight, fat depths and body condition score. Reproductive tract size was measured close to the start of the first joining period. Results showed significant effects of location and birth month on the age at first CL and associated puberty traits. Genotypes did not differ significantly for the age or weight at first CL; however, Brahman were fatter at first CL and had a small reproductive tract size compared with that of Tropical Composite. Genetic analyses estimated the age at first CL to be moderately to highly heritable for Brahman (0.57) and Tropical Composite (0.52). The associated traits were also moderately heritable, except for reproductive tract size in Brahmans (0.03) and for Tropical Composite, the presence of an observed CL on the scanning day closest to the start of joining (0.07). Genetic correlations among puberty traits were mostly moderate to high and generally larger in magnitude for Brahman than for Tropical Composite. Genetic correlations between the age at CL and heifer- and steer-production traits showed important genotype differences. For Tropical Composite, the age at CL was negatively correlated with the heifer growth rate in their first postweaning wet season (–0.40) and carcass marbling score (–0.49), but was positively correlated with carcass P8 fat depth (0.43). For Brahman, the age at CL was moderately negatively genetically correlated with heifer measures of bodyweight, fatness, body condition score and IGF-I, in both their first postweaning wet and second dry seasons, but was positively correlated with the dry-season growth rate. For Brahman, genetic correlations between the age at CL and steer traits showed possible antagonisms with feedlot residual feed intake (–0.60) and meat colour (0.73). Selection can be used to change the heifer age at puberty in both genotypes, with few major antagonisms with steer- and heifer-production traits.
Additional keywords: beef, fertility, puberty, ultrasound, heritability, genetic correlations.
Introduction
Improved female reproductive performance of beef breeds in northern Australia is an important means of increasing profitability (Taylor and Rudder 1986). Several studies have shown that breed differences exist for female fertility traits of tropical genotypes in northern Australia (e.g. Mackinnon et al. 1989; Prayaga 2004). The review of Davis (1993) identified significant within-breed genetic differences for female reproduction traits related to calf output, and results from a large divergent selection study for pregnancy rate in a tropical beef herd generated significant differences in pregnancy rate between the high and low lines (Hetzel et al. 1989; Mackinnon et al. 1990; Davis et al. 1993). However, industry-wide improvement of female reproductive performance by genetic selection has generally proved difficult, mainly because of low heritabilities and the late expression of traits, and difficulties in capturing the necessary joining and reproductive data. Currently, female fertility traits are generally not included in beef genetic-evaluation schemes worldwide, except for days to calving in Australia (Graser et al. 2005) and heifer pregnancy percentage in the USA (Evans et al. 1999). Therefore, inclusion of female fertility traits into beef genetic-evaluation systems may require identification of new traits that can be recorded early in life, are heritable and genetically correlated with the underlying profit trait.
One possible contributor to the observed genetic differences in female reproductive performance is the age at puberty. Breed differences have been reported for the age and weight at puberty (Gregory et al. 1991; Burns et al. 1992; Martin et al. 1992; Thallman et al. 1999) and specifically, Bos indicus breeds (e.g. Brahmans) have been reported to be older at puberty than other breeds (Gregory et al. 1979; Morgan 1981; Bolton et al. 1987; Hearnshaw et al. 1994; Thallman et al. 1999). Several studies have shown that the age at puberty was heritable in Bos taurus breeds of beef cattle (MacNeil et al. 1984; Gregory et al. 1995). Limited estimates exist for Bos indicus genotypes, although in a small study, Vargas et al. (1998) reported a heritability of 0.42 for the age at puberty for Brahmans. For the age at puberty to be useful in a genetic-evaluation scheme it needs to be genetically correlated with female reproductive traits measured in industry herds. However, results are inconclusive. Several studies have shown improved pregnancy or calving rates to be associated with the age at puberty (Laster et al. 1979; Morris et al. 2000; Phocas and Sapa 2004) and Mackinnon et al. (1990) postulated the selection response in female fertility in a divergently selected tropical beef herd was likely due to earlier sexual maturity. Mialon et al. (2001) showed a positive genetic correlation between the age at the first oestrous and the postpartum to return to oestrus interval. However, others (e.g. Dow et al. 1982; Martin et al. 1992; Patterson et al. 1992) have reported no relationship, or unfavourable relationships.
Measuring heifer age at puberty in beef cattle is challenging, particularly on large numbers required for genetic analyses. The two most common methods used to determine the heifer age at puberty are oestrus observation and progesterone assays. Recently, ultrasonography has been used to measure ovarian activity, in particular follicular size and the occurrence of a corpus luteum (CL) in livestock, including cattle (Pierson and Ginther 1988; Griffin and Ginther 1992; Garcia et al. 2002), and consequently could be a practical means for determining puberty in large numbers of heifers. An alternative approach of reproductive tract scoring has also been proposed for pubertal detection in yearling heifers (Andersen et al. 1991, as cited by Martin et al. 1992). The present paper reports results from a large breeding project which aimed to estimate genetic components of whole herd profitability in northern Australia, and to improve production efficiency and product quality, without compromising female performance or adaptation. The primary aim of the present study was to estimate genetic parameters for puberty traits by ovarian ultrasound scanning in two tropical beef genotypes raised in production environments of northern Australia. The study also aimed to estimate genetic relationships between heifer puberty traits and production traits of both heifers and steers, including liveweight and body composition, steer feed intake, net feed intake, and carcass- and meat-quality traits.
Materials and methods
Animals
Females used in the present study were part of a northern Australia breeding project of the Cooperative Research Centre for Cattle and Beef Quality (CRC) described by Burrow et al. (2003). Brahman (BRAH) and Tropical Composite (TCOMP) genotypes were used for the present study, these each being widely used in the subtropical and tropical regions of northern Australia. The TCOMP genotype animals comprised ~50% tropically adapted breeds and 50% non-tropically adapted Bos taurus breeds. On average, the 50% tropically adapted component was approximately one-half derived from the Bos indicus (viz. Brahman) and one-half from tropically adapted Taurine breeds (viz. 24% Africander and 2% N’Dama, through the Senepol). A complete description of the TCOMP genotype by property of origin is presented in Barwick et al. (2009a).
The cattle were bred in northern Australia on seven cooperator properties (four BRAH and three TCOMP) and at the ‘Belmont’ Research Station (both BRAH and TCOMP). Calves were generated by artificial insemination (AI) and natural service. At each property of origin, calf sex, date of birth, dam identification number and dam year of birth were recorded. Sire parentage was determined by DNA fingerprinting. Genetic linkage, across properties of origin and year within a genotype, was generated by AI. Full genetic-linkage statistics for the heifer data are presented in Barwick et al. (2009b).
Heifer allocation and management
Calves were generated during 4 and 3 years for BRAH and TCOMP, respectively. After weaning each year (average age 6.4 months), the complete calf crop for the project from each property of origin was delivered to the control of the project. Heifers were allocated according to the genotype, property of origin and sire to one of the following four Queensland research stations: ‘Brian Pastures’ (BRIANP), ‘Swans Lagoon’ (SWANS), ‘Belmont’ (BELMONT) or ‘Toorak’ (TOORAK) research stations (see Table 1). Distribution of BRAH heifers was proportionally greater to the harsher environments (SWANS and BELMONT), whereas TCOMP were allocated in greater numbers to the more benign locations (i.e. TOORAK and BRIANP). No BRAH heifers were allocated to BRIANP and no TCOMP heifers were allocated to SWANS. At BELMONT, the BRAH and TCOMP heifers that were born and located there were managed as contemporaries throughout the experiment. Further details of heifer management and of the postweaning heifer locations are given by Barwick et al. (2009b).
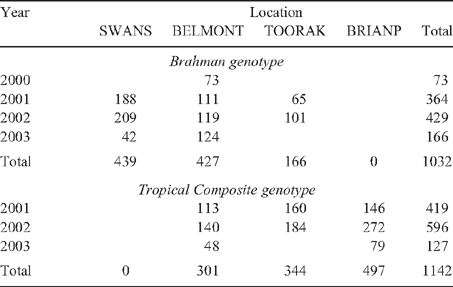
|
At each location, all heifers of the same year of birth were managed as a single group (defined as a cohort). Each cohort was grown out at pasture and joined at ~27 months (i.e. to the first calve as 3-year olds). Slight differences (i.e. less than 2 months) existed in the commencement date of joining across locations within a year, reflecting regional management preferences.
Ovarian measures and measurement procedures
Ovarian scanning
Ovarian activity was assessed in heifers by real-time ultrasound scanning performed by one of four trained operators. At scanning, each ovary was viewed by ultrasound imaging (Aloka SSD-500; Corometrics Medical Systems, Wallington, CT, USA), with linear array 7.5 MHz rectal transducer; or Honda HS-2000V (Honda Electronics, Toyohashi City, Japan), with variable-frequency transducer set at 10 MHz, and the presence of a CL or corpus albicans (CA) was recorded. Training in ovarian scanning was by an experienced ultrasonographer.
Assessment of ovarian activity commenced for a cohort when heifers reached ~200-kg liveweight at 10–12 months of age. Assessments were conducted at intervals of between 3 and 12 weeks, with most being 4–6 weeks. Intervals closer to the start of the assessment (i.e. the first 4 months) for a cohort tended to be longer (approximately every 8 weeks), coinciding with the first ‘dry’ season that heifers experienced after weaning. Following this early period, and coinciding with the ‘wet’ season, the measurements became much more frequent (average interval of 4.6 weeks) and continued for a further 15 months. Some exceptions occurred because of seasonal conditions and availability of technicians. In the 2001 cohorts, assessments were temporarily discontinued following the detection of the first CL or CA.
Reproductive tract size
Reproductive tract size was recorded on heifers when the average cohort age was ~1.5 years, and again approximately 6 months later before the commencement of their first joining. Assessment was performed via rectal palpation to estimate the diameter of the uterine horn, proximal to the bifurcation by manual palpation. The system used was similar, but not identical, to the system described by Andersen et al. (1991), as cited by Martin et al. (1992).
Liveweight and fatness measures
All heifers were weighed, ultrasound-scanned for fat depth at the P8 site (Perry et al. 2001) and body condition scored (Barwick et al. 2009a) at the time of ovarian scanning. Assessors across all locations were trained before the commencement of the study to ensure consistency of all measures and scores. Periodic checks also occurred throughout the experiment to maintain standards.
Trait definitions
For each individual heifer, the age at CL (AGECL) was defined as the age (in days) of the heifer at the first-observed CL (or CA) and was used as an estimate of puberty (i.e. the first confirmed evidence). The date of the first CL for each heifer was then used to identify other measures recorded on the heifer at this time (or within 7 days) and included heifer liveweight at first CL (WTCL), ultrasound scan P8 fat depth at first CL (FATCL) and body condition score at first CL (CSCL) (see Table 2). Three additional traits were defined by first identifying the date of the commencement of the first joining period for each heifer cohort. Reproductive tract size (TSIZE) assessed on average 20 days before the commencement of joining was used with the exception of one BRAH cohort (n = 41) where the scoring occurred 7 months before joining. The other two traits were binary scores that simply classified each heifer (yes = 1, no = 0), regarding the observation of a CL. The first trait was defined as the observation of a CL or CA at any time before, or on, the day of scanning closest to the commencement of joining (CLPRIOR). The second trait was a subset of the first and was defined as the observation of CL or CA on the day of scanning closest to the start of the joining period (CLJOIN).

|
Scan CL-data editing
Checks were performed on all records before analyses. Records from heifers that were sick or unable to be ovarian-scanned (n = 4) and those from heifers who were pregnant without being identified as having a prior CL (n = 6) were removed. Within each cohort, AGECL records more than three standard deviations from the mean were removed (n = 14). A small number (n = 10) of BRAH heifers did not have their first CL observed by the time of analysis, despite being >26 months of age. These received a penalty AGECL record based on the last scanning date at their location plus 30 days.
Heifer growth and composition measures
Heifer growth and body composition traits studied included measures recorded on each heifer at the end of their first postweaning ‘wet’ season (ENDWET) and at the end of their second postweaning ‘dry’ season (ENDDRY), and corresponding to heifer ages of ~18 and 24 months, respectively. These measures were described by Barwick et al. (2009b) and included liveweight (LWT), ultrasound-scanned fat depth at the P8 site (SP8) and over the 12/13th rib (SRIB), scanned area of the M. longissimus thoracis et lumborum (LTL) at the 12/13th rib (SEMA), body condition score (CS), hip height (HH), concentration of the insulin-like growth factor I (IGF-I) in serum and average daily liveweight gain (ADG). ADG was computed by individual animal regressions of liveweight on days for multiple weights recorded during the 6-month period defined for both ENDWET and ENDDRY.
Steer growth, and carcass- and meat-quality measures
Records taken on the steer paternal half-sibs of the heifers were used to investigate relationships between heifer measures of puberty and steer production, and carcass- and meat-quality traits. The growth, body composition and feed-intake traits examined are described by Barwick et al. (2009a) and include measures of feed intake collected during the feedlot finishing phase and measures recorded at feedlot exit (EXIT). In brief, steers (n = 2216) were managed in 12 postweaning grow out groups and entered the feedlot at ~400 kg liveweight. They were fed for an average of 119 days on a high-energy feedlot ration and slaughtered at an average liveweight of 568 kg. Measures recorded on steers included LWT, HH, SEMA, SP8, SRIB, CS, IGF-I, scanned percent intramuscular fat (SIMF), daily feed intake (DFI), residual feed intake (RFI) and feed-test average daily gain (ADG).
Steers were slaughtered in one of two commercial abattoirs where several carcass measures were recorded and meat sample was removed from each carcass for subsequent carcass meat-quality measures. Abattoir carcass measures (CARCASS), described by Wolcott et al. (2009), included hot carcass weight (CWT), cold P8 fat depth (P8c), bone-out retail beef yield percentage (RBY) and Meat Standards Australia (MSA) measured rib fat depth (RIB), eye muscle area (EMA), marbling score (MS), ossification score (OSS) and hump height (HMP). Measures of meat quality were all performed on the LTL muscle sample from the Achilles hung side of the carcass (see Wolcott et al. (2009) for a complete description) and included intramuscular fat percentage (CIMF), shear force (SF_A), compression (CMP_A), cooking loss (LOSS_A) and Minolta L* meat colour (L*).
Statistical analyses
Fixed-effect modelling
Significant fixed effects for each heifer puberty trait were identified by the mixed-model procedure in SAS (SAS Institute, Cary, NC, USA). Analyses were first performed separately for each genotype. All initial models included the independent variables of heifer’s birth month, cohort, property of origin and age of dam, and for TCOMP also terms for sire group and dam group (Barwick et al. 2009a). Birth month of the heifer was included to account for differences in both age and seasonal conditions across the calving period, as the average calving period was 4–5 months within an origin, and differences occurred in the starting calving month across origins. Within a cohort and origin subclass, any adjacent birth months that had less than five animals were combined. Age of dam was recorded in years, and when unknown (~15%) was assigned to be the median for the origin. For TCOMP, sire group, dam group (nested within origin) and their interaction were modelled to account for average additive differences between the composite groups and possible differences in the level of non-additive effects in differing combinations of sire groups and dam groups. Sire was included in all models as a random effect. Initial models for each trait included main effects and all first-order interactions. Non-significant (P > 0.05) terms were sequentially removed to yield the final models for each trait. Final models for BRAH included the effects of cohort and birth month for all traits. Origin was significant for all traits except for WTCL and FATCL. The age of dam was significant only for AGECL and WTCL. Interactions between some of these main effects also were significant, mainly interactions with origin. For TCOMP, final models included cohort and origin for all traits. Birth month was significant for all traits except FATCL. Sire group and dam group were significant for AGECL and CLJOIN, and dam group also for TSIZE and CLPRIOR. Interaction terms were also significant, mainly those with cohort.
Significant fixed effects were also identified for each trait by using a combined dataset for BRAH and TCOMP. These models initially included all the significant effects identified above for each genotype, with the addition of terms for genotype and all first-order interactions of effects with genotype. Each model was reduced by removing non-significant (P > 0.05) effects to yield the final model for use in the combined-genotype analyses.
Variance component estimation
Additive genetic variances and heritabilities for the seven heifer puberty traits were estimated in univariate analyses for BRAH and TCOMP separately, by using restricted maximum likelihood procedures in ASReml (Gilmour et al. 1999). All traits were analysed by using an animal model that included the set of fixed effects identified with SAS and random effects of animal and residual. For each trait, analyses were performed with and without a random maternal common environmental effect, and the best fitting model was determined by a log-likelihood ratio test. A relationship matrix (n = 8640) was used that contained up to three generations of both paternal and maternal pedigree when known. In total, 54 BRAH and 51 TCOMP sires were represented, and across both genotypes there was a total of 51 sires having 20 or more daughters with AGECL records.
Genetic correlations among pairwise combinations of the seven puberty traits were estimated in a series of bivariate analyses with ASReml for each genotype separately, by using models described above. Genetic correlations were also estimated in bivariate models between the seven puberty traits and the groups of heifer (i.e. ENDWET and ENDDRY) and steer (i.e. EXIT and CARCASS) production traits. Models for the steer- and heifer-production traits were described by Barwick et al. (2009a, 2009b) and Wolcott et al. (2009).
Model-predicted means
Predicted means for location × genotype and birth-month effects, for each of the puberty traits, were computed in ASReml as linear functions of terms included in the model from the combined-genotype dataset by using the procedure described by Gilmour et al. (2004). The predicted means were averaged across all other fixed-effect levels present. Data on 15 BRAH heifers born in January were excluded from the prediction of genotype means to avoid averaging across unequally represented birth months. The predicted location × genotype means for the location BELMONT allowed the direct comparison of the two genotypes (i.e. BRAH v. TCOMP) and comprised 297 BRAH and 290 TCOMP heifers, representing 32 and 26 sires, respectively. Because there was a predominance of Belmont Red dams at BELMONT, the TCOMP-predicted means were for a sample of the genotype where the contribution of Africander to the tropically adapted component was higher (i.e. ~40% Africander, 1% N’Dama, 10% Brahman) than existed on average in the whole data. The direct genotype comparison was computed with all BRAH and TCOMP heifers that were born and located at BELMONT. At TOORAK, the comparison of genotypes was confounded with preweaning properties of origin and therefore model-predicted means for each trait at TOORAK were estimated within genotype.
Results and discussion
Summary statistics for each of the puberty traits are presented for BRAH and TCOMP in Table 3 and show the mean level and variation in the traits recorded. These summary statistics presented are not adjusted for fixed effects.
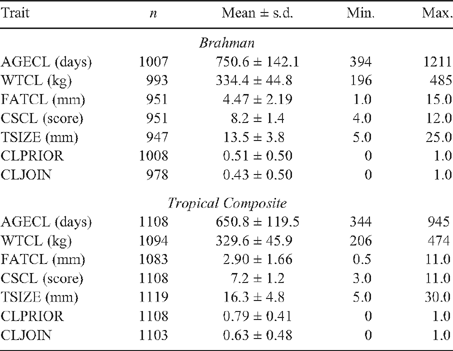
|
Genotype differences
Predicted genotype trait means are presented in Table 4 for each of the heifer puberty traits. BRAH heifers at BELMONT were significantly fatter at first CL (i.e. FATCL difference of 1.5 mm and CSCL of 0.4 score) than were TCOMP at BELMONT. The genotypes were not significantly different for WTCL, AGECL, CLPRIOR and CLJOIN, whereas there was a trend for BRAH to be slightly older at AGECL, with lower percentages for CLPRIOR and CLJOIN, and significantly smaller TSIZE (–1.2 mm). Increased age at puberty in Brahman heifers has been reported in other studies (Gregory et al. 1979; Bolton et al. 1987; Hearnshaw et al. 1994). However, Post and Reich (1980) reported from a small study of mixed tropically adapted breed groups that Brahmans had the youngest age at puberty. Also Burns et al. (1992), in a genotype (i.e. Belmont Red) comparable to the TCOMP, reported a heifer average age at puberty of 583 days and weight at puberty of 319 kg, with 88.2% of heifers estimated to be pubertal into joining at 26 months. These differences in the mean performance, compared with our study, could be due to a range of factors such as seasonal differences, location effects and different methods used to determine the age at puberty. Thus, direct comparison of performance across studies is generally not possible.

|
The observed increased fatness of BRAH at first CL, yet an age and weight at puberty similar to that in TCOMP heifers, suggests genotype differences in fatness at puberty were simply reflecting the overall tendency of Brahman heifers to be fatter. Barwick et al. (2009b) reported the BRAH heifers were 1.2 mm and 1.3 mm fatter than TCOMP heifers for P8 fat depth at the end of their first postweaning wet (i.e. ENDWET) and at the end of the second dry season (i.e. ENDDRY).
Location differences
Location had a large effect on most of the puberty traits in each genotype (Table 4). For TCOMP, AGECL was similar at BRIANP and TOORAK and both were significantly younger than was the case for heifers at BELMONT. For BRAH, heifers at BELMONT and TOORAK were significantly younger at AGECL than at SWANS. These location trends are generally in line with expected environmental differences, on the basis of differences in heifer growth rates and bodyweights across locations. The possible exception was AGECL at TOORAK, where BRAH were older (but not significantly) than at BELMONT. Hearnshaw et al. (1994) found large nutrition by genotype interaction effects on the age at puberty, where Brahman growth rate did not respond to increasing nutrition compared with other genotypes and had extremely low percentages of heifers pubertal at 22 months of age in a subtropical environment.
For TCOMP, there was a positive association between location means for WTCL and AGECL. For BRAH, however, the mean WTCL declined with increasing AGECL. This may indicate a genotype difference, although is more likely to reflect the influence of location on growth rate, particularly the very low dry-season growth rate at SWANS relative to the other two locations. Several studies have shown that differences in growth rates affect both age and weight at puberty. Yelich et al. (1995) observed that increased growth rate in Angus × Hereford heifers resulted in decreased age at puberty, increased weight, and also increased fatness at puberty, which supports our findings for BRAH (Table 4) although not in TCOMP. Ferrell (1982) reported that slower postweaning growth rate delayed the age at puberty and subsequently reduced pregnancy rates compared with heifers that gained weight rapidly after weaning, suggesting that weight was more important than age in determining puberty. Mackinnon et al. (1989) hypothesised that once sexual maturity was reached in Brahman-based breeds there was little effect of increasing weight at mating on subsequent fertility.
No clear trends in predicted location means were observed for FATCL or CSCL, although for TCOMP the BRIANP heifers were significantly leaner at puberty. For BRAH, heifers at SWANS had significantly higher CSCL than those at TOORAK and BELMONT, whereas heifers at BELMONT had significantly higher FATCL than those at TOORAK and SWANS. Hall et al. (1995) showed heifers fed to gain faster postweaning were heavier, taller and younger at puberty, and that puberty was independent of body fat. These results across locations illustrate that puberty in beef heifers is not simply controlled by weight, growth rate or age alone, but appears to involve a combination of factors relating to physiological age, size and growth rate, and probably also involves body condition for BRAH. The large location effects on puberty traits in the present study also highlight that extrapolation of the observed genotype differences beyond the environment in which they were directly compared (i.e. BELMONT) should not be made. TCOMP were purposely not located at SWANS because it was perceived, and accepted by industry, that they would be too poorly adapted to that environment.
Significant location effects were observed for TSIZE. However, there were no clear trends in either TCOMP or BRAH. The predicted means for CLPRIOR and CLJOIN showed significant differences across locations with each genotype. Heifers at BRIANP had a predicted CLPRIOR mean of 91% compared with 64% at BELMONT. For BRAH, heifers from SWANS and TOORAK had significantly lower CLPRIOR and CLJOIN than those at BELMONT. The trends observed for TCOMP correspond to the expected average environment differences across locations (Barwick et al. 2009b). Similarly for Brahmans, the difference in CLPRIOR means for BELMONT and SWANS was as expected. However, the lower than expected percentage with a CLPRIOR at TOORAK most likely reflects small regional differences in the commencement date of joining, where at TOORAK the heifers were on average 30 days younger than at the other two locations.
Birth-month differences
Birth month had a significant effect on all puberty traits (Table 5). As the heifer’s birth month became later in the calving season (i.e. from August to April) there was a trend for AGECL to increase and TSIZE to decrease. Both FATCL and CSCL increased and there was no observed effect on WTCL. CLPRIOR and CLJOIN both declined as the birth month became later. On average, early born heifers (i.e. August to September) reached puberty by ~20 months of age, coinciding with the end of their first postweaning wet season (i.e. May). For late-born heifers (i.e. February to March), the average age at puberty was delayed until the following May, at ~26 months of age. This suggests that heifers that did not achieve puberty before the start of their second postweaning dry season were significantly delayed in reaching puberty, which can have a dramatic impact on the number of heifers with a CL into mating. The dramatic reduction in the growth rate that can be experienced during the dry season (Barwick et al. 2009b) could be a major factor contributing to the delayed onset of puberty in BRAH heifers. Bolton et al. (1987) reported a decrease in the percentage of heifers pubertal into joining of fall-born compared with spring-born calves and the effect was more pronounced as Brahman percentage increased. Arije and Wiltbank (1971) observed that reduced pasture availability during winter delayed puberty in Hereford heifers, such that early born heifers were older at puberty when the spring flush occurred. In a study by Burns et al. (1992), no significant effect of birth month on the age at puberty was observed, although early born heifers were reported to be heavier at puberty.

|
Additive genetic variances and heritabilities of heifer puberty traits
AGECL, WTCL, FATCL and CSCL were all moderately heritable (Table 6). Additive variances for these traits tended to be larger for BRAH than for TCOMP. Heritability estimates (and approximate standard errors) for AGECL were 0.57 ± 0.12 and 0.52 ± 0.12 for BRAH and TCOMP, respectively, and were slightly higher than a pooled estimate of 0.40 for the age at puberty reported in the review of Martin et al. (1992). Our study differed from other reports in that puberty was determined by ultrasound scanning and no literature estimates of heritabilities were found for the age at puberty determined by this technique. The heritability estimates observed provide good evidence that the ovarian ultrasound-scanning technique used, and the frequency with which the observations were taken, were suitable for determining genetic differences in heifer puberty traits. TSIZE was heritable in TCOMP (0.20 ± 0.09) whereas it was lowly heritable in BRAH (0.03 ± 0.06), most likely reflecting the lower average weight and smaller mean reproductive tract size of BRAH (Table 6). Martin et al. (1992) reported a heritability of prejoining reproductive tract score of 0.28, by using a scoring system that subjectively scored the development of the reproductive tract. We observed the binary traits CLPRIOR and CLJOIN were heritable in BRAH (0.33 ± 0.10 and 0.20 ± 0.09, respectively) whereas they were less heritable in TCOMP (0.13 ± 0.07 and 0.07 ± 0.05), which is likely because the majority of TCOMP heifers had an observed CL before, or on, the day of joining. The genetic variation observed in BRAH for AGECL, CLPRIOR and CLJOIN compared with TCOMP suggests a greater importance of genetic differences in the age at puberty, given the expected influence of these traits on subsequent reproductive performance from their first joining. This is supported by the findings of Phocas and Sapa (2004) who reported a large positive genetic correlation between the percentage pubertal at 15 months and the subsequent calving success in two large European breeds of cattle.

|
Relatedness of heifer puberty measures
Genetic correlations among AGECL and the other puberty measures are presented in Tables 7 and 8 for BRAH and TCOMP, respectively. Correlations were generally in the same direction for BRAH and TCOMP although the size of the correlations tended to be larger for BRAH. Moderate to strong positive correlations were estimated between each of the puberty traits AGECL, WTCL, FATCL, and CSCL. They show that heifers that were older at AGECL were genetically heavier and fatter when they reached puberty. This is likely due to the fundamental association between the age and the weight. The estimates were of magnitude similar to the 0.52 genetic correlation reported by Laster et al. (1979) between the age and the weight at puberty. The genetic correlations suggest AGECL, WTCL, FATCL and CSCL are related ways of expressing the same physiological event, and that selection for reduced AGECL would lead to correlated reductions in the other measures. It is likely that AGECL is the trait of most importance to a breeding program because of the annual cycle of beef production, particularly in northern Australia. Genetic differences, or correlated changes, in WTCL may also be important for management considerations, in particular the importance of a minimum heifer weight for natural service mating and also the expected ease of calving.

|

|
The moderate heritabilities and additive genetic variances estimated for puberty traits for BRAH and TCOMP suggest it should be possible to change these traits by selection, and studies (Laster et al. 1979; Morris et al. 2000) have shown the age at puberty to be genetically correlated with measures of reproductive performance of the cow. The prerequisite for this, however, will be the availability of a suitable selection criterion that is heritable and measurable early in life. Although AGECL was heritable, the measurement protocols would most likely preclude its measurement across large numbers of animals in industry herds. However, AGECL was highly negatively correlated with CLPRIOR (–1.0 for BRAH and –0.96 for TCOMP) and CLJOIN (–1.0 for BRAH). These estimates suggest that sires whose daughters were genetically younger at AGECL would have daughters with a higher probability of showing a CL before the commencement of their first joining, and for BRAH, a higher probability of a CL observed on a single scan day close to the start of joining. Therefore, it may be possible to develop a simplified scanning protocol to identify the presence of a CL on the basis of the measures of CLPRIOR or CLJOIN, which could be incorporated into a genetic-evaluation system.
Another possible indirect measure of the age at puberty was TSIZE in TCOMP. TSIZE measured before the first joining was genetically correlated with CLPRIOR (0.70) and AGECL (–0.58) and lowly correlated with WTCL, CSCL and FATCL. TSIZE could also provide a relatively inexpensive indirect genetic measure of puberty. The opportunity also exists to improve measurement of this trait by incorporating additional features of the uterine tract, ovaries and possibly the presence of a CL. Enhancements to the scoring of TSIZE may also improve the heritability estimate for BRAH.
Genetic predictors of heifer puberty
Other measures recorded on the heifers and steers may also be genetically correlated with AGECL. These may prove useful as indirect selection criteria and also provide estimates of any trait antagonisms that exist with heifer puberty traits. Table 9 (BRAH) and Table 10 (TCOMP) present estimated genetic correlations of AGECL and associated puberty traits with measures of heifer growth and body composition measures at ENDWET and ENDDRY. Table 11 (BRAH) and Table 12 (TCOMP) present estimated genetic correlations of AGECL and associated puberty traits with measures of steer EXIT traits.

|

|

|

|
Growth and muscling measures
Genetic correlations for measurements of LWT and SEMA, expressed at a constant age, showed they were moderately negatively correlated with AGECL in heifers and less so in steers, e.g. LWT at ENDWET –0.33 for BRAH (Table 9) and –0.38 for TCOMP (Table 10). A similar estimate of –0.32 was reported in Charolais by Mialon et al. (2001). Gregory et al. (1995) reported lower correlations of –0.05 and 0.11, respectively, for 12- and 24-month weights and the age at puberty. In general, the genetic correlations between growth measures and CLPRIOR and CLJOIN were low. For TCOMP, weights at ENDWET (0.54), ENDDRY (0.52) and ENDWET ADG (0.61) were positively correlated with TSIZE, as were measures of liveweight in steers (0.49; Table 12). Our results indicate that selection for increased heifer weight at ENDWET or ENDDRY would genetically decrease the age at puberty and would also result in a small correlated increase in WTCL. For BRAH, correlations between AGECL and heifer measures of HH were generally low, although the correlation was positive with steer EXIT HH (0.50; Table 11). A genetic correlation of 0.25 between the heifer age at puberty and hip height was also reported in Brahmans by Vargas et al. (1998), suggesting for BRAH, that the age at puberty may be influenced by the frame size.
Genetic correlations between the growth rate and heifer puberty traits differed between the genotypes and also with the season of measurement (Tables 9 and 10). For TCOMP, correlations of ADG with AGECL and WTCL were –0.40 and 0.38 at ENDWET, whereas they were 0.08 and 0.49, respectively, at ENDDRY. For BRAH, the correlations were –0.19 and 0.18 at ENDWET, whereas they were 0.58 and 0.56, respectively, at ENDDRY. The genetic correlations between AGECL and steer feedlot ADG were –0.21 and 0.30 for TCOMP (Table 12) and BRAH (Table 11), respectively. These correlations indicate that within Brahmans, those with the genetic potential for high growth rate (i.e. also larger HH and possibly larger mature size), particularly at ENDDRY, will have genetically older AGECL. This is likely to be a function of the large negative genetic correlations reported by Barwick et al. (2009b) in these heifers at ENDDRY between ADG and the measures of fatness (e.g. –0.81 with SRIB) and may also be influenced by the negative genetic correlation observed between IGF-I and AGECL. Martin et al. (1992) also concluded that faster-gaining breeds of larger mature size reach puberty at later ages. There is generally evidence, including in tropical genotypes in northern Australia (Burrow et al. 1991), that selection for weight gain can lead to genetically improved female reproductive rate. It appears that the season or stage at which the growth rate is measured may be important, as a greater dry-season growth rate in the present study had a detrimental genetic effect on BRAH AGECL. However, Fordyce et al. (1988) reported that increasing the rate of weight gain of Brahman cross heifers during the first postweaning dry season increased the probability of conception at 2 years of age.
Fatness, condition score and IGF-I
Genetic correlations between heifer puberty traits and body composition measures varied between the genotypes and the various measurement stages. In general, estimates for BRAH (Table 9) showed that selection for increased heifer fatness or condition score at either ENDWET (–0.35 for SP8; –0.53 for CS) or ENDDRY (–0.38 for SRIB; –0.43 for CS) would reduce AGECL and also genetically decrease the weight at puberty. Mialon et al. (2001) reported a genetic correlation of –0.57 between the age at the first oestrus and yearling body condition score in Charolais heifers. However, these relationships were not observed for steer measures of fat for BRAH (e.g. 0.04 for SP8 at EXIT; Table 11), and may reflect genetic correlations in fat measures between sexes that were significantly different from one. For example, the genetic correlations of scan P8 fat depth of the heifers with that of the steers at EXIT were 0.79 and 0.60 at ENDWET and ENDDRY, respectively (Barwick et al. 2009b). Genetic correlations for TCOMP between AGECL and fatness measures (Table 10) were low in heifers (e.g. 0.0 and –0.01 for SRIB at ENDWET and ENDDRY, respectively) and steers (e.g. 0.21, 0.13 at EXIT for SP8 and SRIB, respectively; Table 12), suggesting that selection for increased fatness in TCOMP would have little effect on AGECL or WTCL, whereas it would clearly increase FATCL and CSCL. Gregory et al. (1995) also reported no relationship between the age at puberty and condition score.
Values of IGF-I measured in heifers at ENDWET and ENDDRY were both negatively correlated with AGECL, again with estimates for BRAH (Table 9) being significantly more negative (i.e. –0.70 ± 0.13 and –0.43 ± 0.19, respectively) than those for TCOMP (–0.36 ± 0.20 and –0.09 ± 0.28, respectively). IGF-I measured in steers at EXIT was negatively genetically correlated with AGECL for TCOMP (–0.58; Table 12) but not for BRAH (–0.07; Table 11). These results suggest IGF-I may also play a role in the onset of puberty. This is consistent with the review of Wettemann and Bossis (2000) who presented evidence for a role of IGF-I in ovarian function and concluded that reduced levels of IGF-I can cause a cessation of ovulation. However, the reported effect of IGF-I on the onset of puberty in beef heifers is varied. Yilmaz et al. (2006) reported no difference in the heifer age at puberty in small numbers (n = 51) of Angus divergently selected for IGF-I. Whereas Jones et al. (1991), in a small study, reported a significant phenotypic increase in IGF-I in the 56 days before the onset of puberty in Angus, whereas this was not the case for Braford, Simmental or Charolais heifers. However, Yelich et al. (1995) found no significant change in plasma IGF-I at the onset of heifer puberty. The genetic correlations estimated in the present study suggest IGF-I, particularly when measured in heifers at ENDWET, is a good genetic indicator of not only AGECL, but also CLPRIOR, particularly for BRAH (0.75). The utility in a genetic-evaluation system of AGECL, or any of the indirect measures, will depend on their genetic correlation with subsequent measures of female reproductive performance and the direction and magnitude of genetic correlations with other traits of economic importance.
Genetic relationships between steer feed intake, and carcass- and meat-quality traits with heifer puberty traits
In a multiple-trait selection framework it is important to know whether there are sizeable genetic correlations with production and meat-quality traits of the slaughter steer, as well as with aspects of female reproduction. This is particularly so if antagonisms exist that would need to be considered in a selection index. Estimates of genetic correlation between heifer puberty traits and steer exit-feedlot feed-intake measures, and with carcass- and meat-quality (CARCASS) measures are presented in Table 11 (BRAH) and Table 12 (TCOMP).
Feed intake and residual feed intake
Barwick et al. (2009a) reported the heritability of DFI and RFI to be 0.49 and 0.24, respectively, for BRAH and 0.51 and 0.38, respectively, for TCOMP. The estimates of genetic correlation between AGECL and DFI were low in both genotypes (Tables 11 and 12) and for RFI in TCOMP (Table 12). However, for BRAH, RFI was negatively genetically correlated (Table 11) with heifer AGECL (–0.60) and was also moderately to strongly correlated with the other puberty traits. These correlations showed that selection for reduced RFI (i.e. improved feed efficiency) in a steer feedlot-finishing test in BRAH would increase AGECL (and WTCL and FATCL) and reduce CLPRIOR. Improved RFI is genetically correlated (–0.61, Barwick et al. 2009a) with taller steers at exit for BRAH, and HH at EXIT was positively correlated (0.50) with AGECL in the present study. This suggests that there are important, and potentially antagonistic, genetic associations among the mature size, heifer puberty and steer RFI in BRAH, and these do not seem to be greatly influenced by fatness. By contrast, AGECL in TCOMP had little relationship to RFI (0.02), suggesting no such antagonism in this genotype.
Carcass and meat quality
There was little evidence of genetic antagonisms between heifer puberty traits and carcass- and meat-quality measures (Tables 11 and 12). The exception was meat colour (L*) for BRAH, where the genetic correlation with AGECL was 0.73. Of particular interest were the low correlations between tenderness and AGECL (e.g. SF_A = –0.16 and –0.05 for BRAH and TCOMP, respectively), indicating selection for improved meat tenderness and female puberty could occur independently. Wolcott et al. (2009) reported that L* could be considered as an indirect selection criterion for meat tenderness, with a genetic correlation of –0.66 with SF_A. However, the positive genetic correlation between meat colour and AGECL in BRAH indicated that selection for increased L* (i.e. to reduced shear force) would genetically increase AGECL in this breed. This may also be associated with observed correlations in BRAH between L* and heifer ENDWET IGF-I (–0.72) and ENDDRY ADG (0.60) reported by Wolcott et al. (2009), suggesting possible biological associations among measures of meat colour, weight gain and IGF-I concentration.
Some carcass traits had potentially favourably genetic correlations with AGECL. For BRAH, RBY was negatively (–0.55) correlated with AGECL (Table 11), suggesting selection to increase beef-yield percentage would reduce AGECL, and would also genetically increase CLPRIOR (0.66) and CLJOIN (0.83). Laster et al. (1979) reported a negative correlation (–0.70) between the breed-group means of heifer age at puberty and the fat trim percentage. Whereas Mialon et al. (2001) reported no genetic association between the heifer age at the first oestrus and the fat content of male carcasses in Charolais cattle. For TCOMP (Table 12), AGECL was negatively correlated with MS (–0.49) and not correlated with RBY (–0.10), suggesting that selection to increase MS in TCOMP would favour reduced AGECL. However, Bergfeld et al. (1995) reported no differences in the age at puberty in Angus heifers sired by high- and low-marbling EPD sires. The genetic correlation with P8c was also positive with AGECL (0.43) and WTCL (0.64) for TCOMP, suggesting that decreased carcass P8 fatness would be genetically associated with decreased AGECL and WTCL.
Conclusions
Heifer age at puberty is affected by genetic and environmental influences. Age at puberty can be significantly delayed in late-born calves and also in environments that limit growth rates, particularly during the dry season. Therefore, management can be used to reduce the age at puberty by controlling the month of start of calving and its duration, and the nutrition management of the pre-pubertal heifer. Genotype differences existed and could be exploited through the choice of breeds, and within both genotypes sufficient genetic variation exists such that selection could be used to reduce the age at puberty. However, differences between the genotypes Brahman and Tropical Composite in their genetic relationships between traits suggest differences in their biology for mechanisms controlling puberty and the need for separate genetic-evaluation schemes. In general, more genetic variation was observed for Brahman than for Tropical Composite. The genetic differences observed for the age at first CL for Brahman appear to be more important than those for Tropical Composite because of the expected influence of age at puberty on reproductive performance from their first joining, given the lower percentage of Brahman heifers observed with a CL before joining. Although significant genetic variation existed for heifer puberty traits in the Tropical Composite, it is yet to be determined whether they are important as predictors of a lifetime calving performance.
In general, there were few strong genetic indicators of heifer age at puberty, except for IGF-I in Brahman heifers measured at the end of their first postweaning wet season. Genetic relationships indicate that selection to improve heifer age at puberty and steer traits could occur reasonably independently, except for Brahman residual feed intake and meat colour. Other correlations of lower magnitude, given the moderate to large standard errors, could also be economically important.
These results and measures form the basis for further studies examining the genetic associations of puberty traits with tropical adaptation traits, and importantly, their associations with the first and subsequent calving performances of these females. These results will ultimately determine the utility of measuring early-in-life female puberty traits and including them in a genetic-selection scheme.
Acknowledgements
The authors acknowledge the Cooperative Research Centre for Cattle and Beef Quality (and its core partners; The University of New England, NSW Department of Primary Industries, CSIRO and Queensland Department of Primary Industries and Fisheries), the Commonwealth funding through the CRC’s Program, and the generous financial support of Meat and Livestock Australia and the Australian Centre for International Agricultural Research (ACIAR). We acknowledge the tremendous contributions of the Australian Agricultural Co., C&R Briggs, Consolidated Pastoral Co., North Australian Pastoral Co., Stanbroke Pastoral Co., J&SM Halbersater, GE&J McCamley, S Kidman & Co., MDH, and the research stations of AgForce Queensland and QDPI&F. We also gratefully acknowledge all Beef CRC participants, both scientists and technical staff, who contributed to or supported the work, including those involved in cattle management, data collection, laboratory analyses and data handling. We especially acknowledge the contributions of Mick Sullivan, Matt Wolcott, Warren Sim, Peggy Olsson, Tracy Longhurst, Trudy Obst, Steve O’Connor, Russ Tyler, Neil Cooper, Graeme Halford and Andrew McCann.
Andersen KJ,
LeFever DG,
Brinks JS, Odde KG
(1991) The use of reproductive tract scoring in beef heifers. Agri-Practice 12, 4.

Arije GF, Wiltbank JN
(1971) Age and weight at puberty in Hereford heifers. Journal of Animal Science 33, 401–406.
|
CAS |
PubMed |
Barwick SA,
Wolcott ML,
Johnston DJ,
Burrow HM, Sullivan MT
(2009a) Genetics of steer daily and residual feed intake in two tropical beef genotypes, and relationships among intake, body composition, growth and other post-weaning measures. Animal Production Science 49, 351–366.
| Crossref | GoogleScholarGoogle Scholar |
Barwick SA,
Johnston DJ,
Burrow HM,
Holroyd RG,
Fordyce G,
Wolcott ML,
Sim WD, Sullivan MT
(2009b) Genetics of heifer performance in ‘wet’ and ‘dry’ seasons and their relationships with steer performance in two tropical beef genotypes. Animal Production Science 49, 367–382.
| Crossref | GoogleScholarGoogle Scholar |
Bergfeld EGM,
Rasby RJ,
Nielsen MK, Kinder JE
(1995) Heifers sired by bulls with either high or low expected progeny differences (EPDs) for marbling do not differ in age at puberty. Animal Reproduction Science 40, 253–259.
| Crossref | GoogleScholarGoogle Scholar |
Bolton RC,
Frahm RR,
Castree JW, Coleman SW
(1987) Genotype × environment interactions involving proportion of Brahman breeding and season of birth. II. Postweaning growth, sexual development and reproductive performance of heifers. Journal of Animal Science 65, 49–55.
Burns BM,
Howitt CJ,
Webber RJ,
Rudder TH,
Tierney TJ, O’Rourke PK
(1992) Productivity of Hereford, highgrade Simmental and Belmont Red beef heifers in central Queensland 4. Liveweight and age of heifers at puberty. Australian Journal of Experimental Agriculture 32, 1011–1015.
| Crossref | GoogleScholarGoogle Scholar |
Burrow HM,
Seifert GW, Hetzel DJS
(1991) Consequences of selection for weaning weight in zebu, Bos Taurus and zebu × Bos Taurus cattle in the tropics. Australian Journal of Agricultural Research 42, 295–307.
| Crossref | GoogleScholarGoogle Scholar |
Burrow HM,
Johnston DJ,
Barwick SA,
Holroyd RG,
Barendse W,
Thompson JM,
Griffith GR, Sullivan M
(2003) Relationships between carcass and beef quality and components of herd profitability in northern Australia. Proceedings of the Association for the Advancement of Animal Breeding and Genetics 15, 359–362.
Davis GP
(1993) Genetic parameters for tropical beef cattle in northern Australia: a review. Australian Journal of Agricultural Research 44, 179–198.
| Crossref | GoogleScholarGoogle Scholar |
Davis GP,
Corbet NJ,
Mackinnon MJ,
Hetzel DJS,
Entwistle KW, Dixon R
(1993) Response in female fertility and calf growth to selection for pregnancy rate in tropical beef cattle. Australian Journal of Agricultural Research 44, 1509–1521.
| Crossref | GoogleScholarGoogle Scholar |
Dow JS,
Moore JD,
Bailey CM, Foote WD
(1982) Onset of puberty in heifers of diverse beef breeds and crosses. Journal of Animal Science 55, 1041–1047.
| PubMed |
Evans JL,
Golden BL,
Bourdon RM, Long KL
(1999) Additive genetic relationships between heifer pregnancy and scrotal circumference in Hereford cattle. Journal of Animal Science 77, 2621–2628.
|
CAS |
PubMed |
Ferrell CL
(1982) Effects of postweaning rate of gain in onset of puberty and productive performance of heifers of different breeds. Journal of Animal Science 55, 1272–1283.
|
CAS |
PubMed |
Fordyce G,
Holroyd RG,
James TA, Reid DJ
(1988) The effect of post-weaning growth on the fertility of Brahman cross heifers. Proceeding of the Australian Society of Animal Production 17, 396.
Garcia MR,
Amstalden M,
Williams SW,
Stanko RL,
Morrison CD,
Keisler DH,
Nizielski SE, Williams GL
(2002) Serum leptin and its adipose gene expression during pubertal development, the estrous cycle and different seasons in cattle. Journal of Animal Science 80, 2158–2167.
|
CAS |
PubMed |
Gilmour AR,
Cullis BR,
Welham SJ,
Gogel B, Thompson R
(2004) An efficient computing strategy for prediction in linear mixed models. Computational Statistics & Data Analysis 44, 571–586.
| Crossref | GoogleScholarGoogle Scholar |
Graser H-U,
Tier B,
Johnston DJ, Barwick SA
(2005) Genetic evaluation for the beef industry in Australia. Australian Journal of Agricultural Research 45, 913–921.
Gregory KE,
Laster DB,
Cundiff LV,
Smith GM, Koch RM
(1979) Characterization of biological types of cattle – Cycle III: II. Growth rate and puberty in females. Journal of Animal Science 49, 461–471.
Gregory KE,
Lunstra DD,
Cundiff LV, Koch RM
(1991) Breed effects and heterosis in advanced generations of composite populations for puberty and scrotal traits of beef cattle. Journal of Animal Science 73, 2235–2242.
Gregory KE,
Cundiff LV, Koch RM
(1995) Genetic and phenotypic (co)variances for production traits of female populations of purebred and composite beef cattle. Journal of Animal Science 73, 2235–2242.
|
CAS |
PubMed |
Griffin PG, Ginther OJ
(1992) Research applications of ultrasonic imaging in reproductive biology. Journal of Animal Science 70, 953–972.
|
CAS |
PubMed |
Hall JB,
Staigmiller RB,
Bellows RA,
Short RE,
Moseley WM, Bellows SE
(1995) Body composition and metabolic profiles associated with puberty in beef heifers. Journal of Animal Science 73, 3409–3420.
|
CAS |
PubMed |
Hearnshaw H,
Arthur PF,
Barlow R,
Kohun PJ, Darnell RE
(1994) Evaluation of Bos indicus and Bos taurus straightbreds and crosses. II. Postweaning growth, puberty and pelvic size of heifers. Australian Journal of Agricultural Research 45, 795–805.
| Crossref | GoogleScholarGoogle Scholar |
Hetzel DJS,
Mackinnon MJ,
Dixon R, Entwistle KW
(1989) Fertility in a tropical beef herd divergently selected for pregnancy rate. Animal Production 49, 73–81.
Jones EJ,
Armstrong JD, Harvey RW
(1991) Changes in metabolites, metabolic hormones, and luteinizing hormone before puberty in Angus, Braford, Charolais, and Simmental heifers. Journal of Animal Science 69, 1607–1615.
|
CAS |
PubMed |
Laster DB,
Smith GM,
Cundiff LV, Gregory KE
(1979) Characterization of biological types of cattle (cycle II) II. Postweaning growth and puberty of heifers. Journal of Animal Science 48, 500–508.
|
CAS |
PubMed |
Mackinnon MJ,
Hetzel DJS, Taylor JF
(1989) Genetic environment effects on the fertility of beef cattle in a tropical environment. Australian Journal of Agricultural Research 40, 1085–1097.
| Crossref | GoogleScholarGoogle Scholar |
Mackinnon MJ,
Hetzel DJS,
Corbet NJ,
Bryan RP, Dixon R
(1990) Correlated responses to selection for cow fertility in a tropical beef herd. Animal Production 50, 417–424.
MacNeil MD,
Cundiff LV,
Dinkel DA, Koch RM
(1984) Genetic correlations among sex-limited traits in beef cattle. Journal of Animal Science 58, 1171–1180.
|
CAS |
PubMed |
Martin LC,
Brinks JS,
Bourdon RM, Cundiff LV
(1992) Genetic effects on beef heifer puberty and subsequent reproduction. Animal Science 70, 4006–4017.
|
CAS |
Mialon MM,
Renand G,
Krass D, Menissier F
(2001) Genetic relations between cyclic ovarian activity in heifers and cows and beef traits in males. Genetics Selection Evolution 33, 273–287.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
Morgan JHL
(1981) A comparison of breeds and their crosses for beef production. II. Growth and puberty of heifers. Australian Journal of Agricultural Research 32, 811–824.
Morris CA,
Wilson JA,
Bennett GL,
Cullen NG,
Hickey SM, Hunter JC
(2000) Genetic parameters for growth, puberty, and beef cow reproductive traits in a puberty selection experiment. New Zealand Journal of Agricultural Research 43, 83–91.
Patterson DJ,
Corah LR,
Brethour JR,
Higgins GH, Stevenson JS
(1992) Evaluation of reproductive traits in Bos Taurus and Bos indicus crossbred heifers: relationship of age of puberty to length of the postpartum interval to estrus. Journal of Animal Science 70, 1994–1999.
|
CAS |
PubMed |
Perry D,
Shorthose WR,
Ferguson DM, Thompson JM
(2001) Methods used in the CRC program for the determination of carcass yield and beef quality. Australian Journal of Experimental Agriculture 41, 953–957.
| Crossref | GoogleScholarGoogle Scholar |
Phocas F, Sapa J
(2004) Genetic parameters for growth, reproductive performance, calving ease and suckling performance in beef cattle heifers. Animal Science 79, 41–48.
Pierson RA, Ginther OJ
(1988) Ultrasonic imaging of the ovaries and uterus in cattle. Theriogenology 29, 21–37.
| Crossref | GoogleScholarGoogle Scholar |
Post TB, Reich MM
(1980) Puberty in tropical breeds of heifers as monitored by plasma progesterone. Proceeding of the Australian Society of Animal Production 13, 61–62.
Prayaga KC
(2004) Evaluation of beef cattle genotypes and estimation of direct and maternal genetic effects in a tropical environment. 3. Fertility and calf survival traits. Australian Journal of Agricultural Research 55, 811–824.
| Crossref | GoogleScholarGoogle Scholar |
Taylor WJ, Rudder TH
(1986) Reduction of sale age for export carcases: will it affect beef producers in tropical regions? Proceedings of the Australian Society of Animal Production 16, 379–382.
Thallman RM,
Cundiff LV,
Gregory KE, Koch RM
(1999) Germplasm evaluation in beef cattle – Cycle IV: postweaning growth and puberty of heifers. Journal of Animal Science 77, 2651–2659.
|
CAS |
PubMed |
Vargas CA,
Elzo MA,
Chase CC Jr,
Chenoweth PJ, Olson TA
(1998) Influence of frame size and body composition on performance of Brahman cattle. Journal of Animal Science 73, 2536–2541.
Wettemann RP, Bossis I
(2000) Energy intake regulates ovarian function in beef cattle. Proceedings of the American Society Animal Science 1999, 1–10.
Wolcott ML,
Johnston DJ,
Barwick SA,
Iker CL,
Thompson JM, Burrow HM
(2009) Genetics of meat quality and carcass traits and the impact of tenderstretching in two tropical beef genotypes. Animal Production Science 49, 383–398.
| Crossref | GoogleScholarGoogle Scholar |
Yelich JV,
Wettemann RP,
Dolezal HG,
Lusby KS,
Bishop DK, Spicer LJ
(1995) Effects of growth rate on carcass composition and lipid partitioning at puberty and growth hormone, insulin-like growth factor I, insulin, and metabolites before puberty in beef heifers. Journal of Animal Science 73, 2390–2405.
|
CAS |
PubMed |
Yilmaz A,
Davis ME, Simmen RCM
(2006) Analysis of female reproductive traits in Angus beef cattle divergently selected for blood serum insulin-like growth factor I concentration. Theriogenology 65, 1180–1190.
| Crossref | GoogleScholarGoogle Scholar |
CAS |
PubMed |
1 Animal Genetics and Breeding Unit is a joint venture of New South Wales Department of Primary Industries and the University of New England.